

Growth traits associated with drought survival, recovery and persistence of cocksfoot (Dactylis glomerata) under prolonged drought treatments
M. Abdollahi Bakhtiari A , F. Saeidnia A B , M. M. Majidi A and A. Mirlohi AA Department of Agronomy and Plant Breeding, College of Agriculture, Isfahan University of Technology, Isfahan 84156-83111, Iran.
B Corresponding author. Email: f.saeednia@ag.iut.ac.ir
Crop and Pasture Science 70(1) 85-94 https://doi.org/10.1071/CP18473
Submitted: 21 April 2018 Accepted: 26 October 2018 Published: 24 December 2018
Abstract
The effect of prolonged drought treatments on persistence, growth traits, drought survival and post-drought recovery was investigated in two sets of differently managed genotypes of cocksfoot (Dactylis glomerata L.). In total, 72 genotypes (two sets of 36 managed for seed and forage harvest, respectively) were evaluated for agro-morphological traits in the field during 2013–15 under normal and deficit irrigation regimes. In the fourth year (2016), irrigation was withheld in both irrigation treatments for 2 months and then plants were re-watered to investigate the effect of prolonged drought regimes on traits related to post-drought recovery. The deficit irrigation regime decreased persistence and recovery. These reductions were lower in the seed-management than the forage set, which indicates that delaying the first harvest of the seed-management treatment to maximise seed production led to lower impact of drought stress. The seed-management treatment also had lower persistence. The forage-management treatment had higher recovery under normal irrigation, whereas under deficit irrigation, the seed-management treatment had higher recovery. Association analysis showed the possibility of selecting genotypes having high values of persistence and drought tolerance. Results also showed a negative correlation between days to flowering and recovery after drought, indicating that selection for earliness may improve survivability and persistence of these plants. Superior genotypes with higher forage production and better recovery, persistence and drought tolerance may be recommended for development of synthetic cultivars.
Additional keywords: orchardgrass, regrowth, survivability.
Introduction
Drought is a primary abiotic constraint affecting crop production worldwide and will become increasingly important with climate change and shortages of freshwater associated with population growth (Hu and Xiong 2014; Lobell et al. 2014). This emphasises the need for suitable plant species or genotypes that are able to survive and produce under drought-stress conditions. Plants have evolved sophisticated defence mechanisms to adapt to drought stress, such as drought escape, drought avoidance and dehydration tolerance mechanisms (Zlatev 2005; Touchette et al. 2007). All of these mechanisms serve to improve the efficiency of water uptake and use, or to reduce loss (Wang and Huang 2004; Pirnajmedin et al. 2015).
Identification and selection of plant genotypes with survival, recovery and sustainable performance during prolonged drought periods form one of the main objectives of plant-breeding programs for arid and semi-arid regions. Perennial forage grasses such as cocksfoot (also known as orchard grass, Dactylis glomerata L.) can be a valuable alternative to annuals under water-deficit conditions (Saeidnia et al. 2016). Cocksfoot is a long-lived, perennial, cross-pollinating grass species that plays an important role in the livestock industry and in the sustainable development of environments worldwide (Stewart and Ellison 2011; Jiang et al. 2014).
Forage yield and persistence, which is defined as ‘the ability to survive successive summer droughts and retain forage productivity’ (Annicchiarico et al. 2011; Pecetti et al. 2011), are two of the most important objectives in forage-grass breeding (Sanada et al. 2010; Van Minnebruggen et al. 2015). In crops such as cocksfoot that are harvested multiple times during one growing season, both forage yield and persistence are related to regrowth. A good capacity to regrow after cutting is an essential factor to forage yield and its stability throughout the season (Van Minnebruggen et al. 2015). Furthermore, most efforts to improve persistence in forage grasses have focused on selecting tolerance to biotic and abiotic stresses (Pirnajmedin et al. 2015; Saeidnia et al. 2016, 2017), selection of genotypes with high root expansion (Norton et al. 2006; Karcher et al. 2008; Nie et al. 2008; Shaimi et al. 2009; Chai et al. 2010), or determination of appropriate selection criteria to obtain high-yielding cultivars with a high level of persistence (Pirnajmedin et al. 2017). However, it is also known that poor regrowth can result in low persistence. Therefore, higher persistence and recovery in forage grasses are also associated with the ability to regrow from meristems on rhizomes and from deep root systems that can exploit remaining soil water (Nie et al. 2008).
Improving the persistence and survivability of perennial species is critical in areas with prolonged periods of drought. In addition, rapid recovery of damaged plant tissues and the regrowth of new tissues following drought stress are important in perennial grass management. Post-drought recovery is largely related to preservation of growing points during drought stress, compensatory growth in surviving tissues and the rate of mobilisation of root reserves (Chai et al. 2010). In semi-arid rainfed regions, intense summer droughts stop all production of plants. In these regions, the most relevant criterion for plant tolerance is drought survival, i.e. the ability of plants to remain alive during summer and recover when rehydration occurs. Moreover, in such species, summer growth is inversely correlated with survival and persistence (Volaire et al. 1998). Therefore, the most important strategy is not the ability to produce during drought, but the ability to survive and to recover rapidly after autumn rains (Volaire et al. 2014). Because this aspect of drought tolerance has received limited research attention, it is essential to understand the underlying mechanisms in order to develop perennial forage varieties suitable for arid and semi-arid regions (Volaire and Lelièvre 1997; Volaire et al. 1998).
Efficient recovery from drought may prove more important than plant growth during a dry season because it enables a species to persist in swards or pastures and improve its competition with less drought-resistant species (Kanapeckas et al. 2008). However, in some regions of the world such as Iran, forage species are usually used for seed production in the first harvest and hay production or pasture in the following harvests (Saeidnia et al. 2016, 2018). Therefore, both the seed and forage can be valuable, depending on the utilisation purpose. Pasture-management strategies such as seed and forage harvest should receive attention for production and environmental reasons (Kemp et al. 2000; Dowling et al. 2006).
There is a lack of information on the persistence, survival and recovery of cocksfoot genotypes subjected to prolonged drought conditions and on the association of these factors with morphological traits and drought tolerance. Moreover, little information is available on the impact of pasture-management strategies such as seed and forage harvest on the persistence and regrowth of cocksfoot genotypes after prolonged drought stress. This study was conducted to (i) evaluate genetic variation for persistence, survival and post-drought recovery of Iranian cocksfoot genotypes; (ii) determine the relationship between morphological traits, drought tolerance and post-drought recovery, and identify an appropriate combination of these traits for selection of suitable genotypes for future studies; and (iii) compare the effects of seed harvest and forage harvest on recovery, persistence and productivity of cocksfoot.
Materials and methods
Experimental site
The experiment was conducted at the research farm of Isfahan University of Technology, Isfahan, Iran (32°30ʹN, 51°20ʹE) on a Typic Haplargid, silty clay loam soil. The soil is calcareous, non-saline and non-sodic with 390 g kg–1 of calcium carbonate equivalent, 4.0 g kg–1 of organic carbon, and 0.77 g kg–1 of total nitrogen, with pH 8.3. Monthly temperatures (minimum and maximum) and rainfall of this region during the trial are shown in Fig. 1. The mean annual precipitation is 125 mm and temperature 16.3°C. In this region, there is no rain in the summer and crops must be irrigated over that season.

|
Plant materials and field evaluation
Field management during 2013–15
The genotypes of orchardgrass used in this study were randomly selected from a diverse nursery population made by polycross in a set of 25 parental genotypes. The parental genotypes of the polycross were randomly selected from a previously constructed nursery established in 2006, which mainly consisted of natural ecotypes of orchardgrass from wide geographical regions of Iran, and some foreign natural ecotypes (Supplementary Materials table S1, available at the journal’s website). These parental genotypes were clonally propagated in a greenhouse, and transferred to an isolated polycross nursery in March 2007 to produce new genetic combinations. In June 2009, seeds of each genotype from all of the replications were harvested separately and mixed together in equal proportions, and then a random sample of each seed mixture was taken and the samples were bulked to produce 25 half-sib families. Polycross seeds from the 25 half-sib families were grown in plastic boxes in a greenhouse during winter 2010. Established seedlings were space-planted in the field according to a randomised complete block design with four replications in March 2010. Every plot contained 20 plants of each half-sib family. Therefore, a large population containing ~2000 plants was created.
Thirty-six single plants were randomly selected within this population and assigned to the seed-management treatment, and 36 single plants were randomly selected and allocated to the forage-management treatment (Fig. 2). These genotypes were clonally propagated in a greenhouse during winter 2012 and were space-planted in the field in two separate experiments (for normal and deficit irrigation regimes) in a randomised complete block design with 12 replications in March 2012. In each plot, six clones were planted in each row, with a distance of 50 cm between and within rows, and evaluated under two levels of irrigation (as two experiments close together): a normally watered environment, and an intense-water-deficit environment. Under the normally irrigated regime, plants were watered when 50% of total available soil moisture was depleted from the root-zone. Under the deficit irrigation regime, water was applied when 90% of total available soil moisture was depleted from the root-zone, following accepted methods of determination of evapotranspiration (Allen et al. 1998). Water deficit was applied during the growing season in each year of the experiment from 1 May to 1 October. Irrigation intervals during the growing season and between the two irrigation regimes varied depending on the weather conditions. To determine the amount of irrigation water needed to restore the soil moisture deficit to the field capacity and to determine the times of irrigation, soil moisture content was also measured based on standard gravimetric methods (Clarke Topp et al. 2008) at three depths (0–20, 20–40, and 40–60 cm). Irrigation depth (I, cm) was determined according to the following equation:
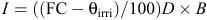

|
where FC is soil gravimetric moisture percentage at field capacity, θirri is soil gravimetric moisture percentage at the time of irrigation, D is root-zone depth, and B is soil bulk density at the root-zone (1.4 g cm–3). Water volumes that should be applied in each irrigation regime were calculated by multiplying the irrigation depth by the total area of plots under each irrigation regime. Water was delivered from a pumping station via polyethylene pipe and the volumes applied were measured with a volumetric counter. The actual depth of irrigation (Ig) was calculated according to the following equation:
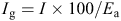
where Ea is the irrigation efficiency (%), assumed as 75% during the growing season (Fig. 3).
|
No data were recorded during 2013 (plant establishment year). In 2014 and 2015, half of the genotypes (genotypes 1–36) were randomly assigned to forage evaluation and the rest (genotypes 37–72) to seed evaluation. Days to ear emergence (DEE) and days to anthesis (DTA) were recorded as the time from 1 March until appearance of three panicles and onset of pollen shedding, respectively, in each plant and in each year. Number of stems per plant was recorded at the pollination stage. The distance from the plant base to the top of the three tallest panicles at full anthesis was considered as the plant height. Crown diameter was measured as the plant width remaining after the first harvest. Degree of winter growth (DWG) was scored from 1 (weakest) to 9 (most robust) according to the plant viability, vitality, canopy size and appearance at the end of the cool season, in each year.
In the forage-management treatment, three harvests of aboveground biomass were undertaken. The first harvest was in late spring after pollination, the second in late summer, and the third in late autumn. In the seed-management treatment, the first cut was assigned to seed harvest and the second and third cuts were allocated to forage harvest. At each harvest, the biomass of each plant was cut manually at 5 cm above the ground, dried at 75°C for 48 h and then weighed to obtain dry matter yield (DMY). Percentage DMY was calculated as the ratio of fresh forage yield of a genotype to dry forage yield of the same genotype, multiplied by 100. The stress tolerance index (STI) (Fernandez 1992) for each genotype was calculated based on dry forage yield under normal and deficit irrigation regimes according to the following equation:
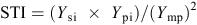
where Ysi is the yield of the ith genotype in the deficit irrigation regime, Ypi is the yield of the ith genotype under normal irrigation, and Ymp is the mean yield over all genotypes in the normal irrigation regime.
Field management during drought in 2016 (withholding irrigation)
In 2016, after 3 years of field evaluation, all genotypes were assessed for drought recovery. In June 2016, irrigation was withheld from both previously irrigated regimes (normal and deficit irrigation) to impose a severe drought for 60 days until complete desiccation of foliage. Plants were then irrigated every week to assess recovery. After 6 weeks of rewatering, recovery-related traits including recovery yield (RY), degree of recovery (DR), recovery ratio (RR) and percentage of recovery (PR) were measured. RY was recorded by measuring the dry forage yield of each genotype after withholding irrigation and rewatering. DR was visually scored on a scale of 0–9, where green and fully hydrated leaves were rated 9 and desiccated brown or dead leaves rated as 0. RR was calculated as the proportion of forage yield of a genotype before withholding irrigation to the forage yield of the same genotype after withholding irrigation and then rewatering. PR was calculated as percentage of live plants of a genotype that have shown recovery after rewatering. Persistence of cocksfoot genotypes was calculated as the difference in forage yield from year 4 (2016) to year 2 (2014).
Statistical analyses
Data were tested for normality by Kolmogorov–Smirnov test, and homogeneity of variance was tested by Bartlett’s test. Differences between environments, years and genotypes and their interactions were examined by using analysis of variance (ANOVA). A split-plot in time (year) model was used for the combined analysis of data obtained during 2 years under two irrigation regimes, according to Steel and Torrie (1980) and Nguyen and Sleper (1983). In addition, a combined ANOVA was performed on recovery-related traits. Comparison of means was undertaken by using the least significant difference (l.s.d.) test at P = 0.05 (Steel and Torrie 1980). The phenotypic correlation (rp) between two traits was calculated as follows:
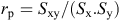
where Sxy is the phenotypic covariance for the characters x and y, and Sx and Sy are the standard deviations for traits x and y, respectively.
Principal component analysis (PCA) was performed to reduce the multiple dimensions of data space (Johnson and Wichern 2007) by using Statgraphics version 15.2.11 (Statgraphics Technologies, The Plains, VA, USA).
Results
The ANOVA showed significant (P < 0.01) differences between the normal and deficit irrigation regimes for all of the measured traits (Supplementary Materials tables S2 and S3). The effect of genotype was significant for all traits, indicating significant variation among the selected genotypes. Genotype × irrigation regime effects were also significant for all traits except persistence. Results from the ANOVA also revealed that the effects of year, year × irrigation regime and genotype × year were significant for most of the traits (Supplementary Materials tables S2 and S3).
With regard to morphological and agronomic traits in 2014–15 (Table 1), some mean values decreased under deficit irrigation (plant height, number of stems, DMY, crown diameter and DWG) and some significantly increased (DEE, DTA and percentage DMY for forage-management treatment; and percentage DMY cut 3 for seed-management treatment).

|
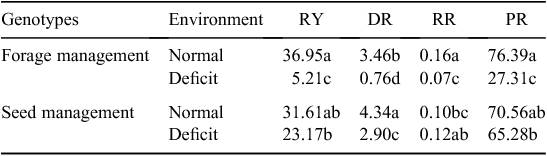
After withholding irrigation and rewatering in 2016, the recovery-related traits RY, DR, RR and PR generally decreased under deficit irrigation compared with the normal irrigation environment (by 86%, 78%, 56% and 64%, respectively, for forage management; and 27% decrease, 33% decrease, 20% increase and 7% decrease for seed management; Table 2); the reductions were greater for the forage-management treatment than the seed-management treatment.
|
Biplots of STI v. persistence under normal and deficit irrigation regimes are presented in Fig. 4a and b, respectively; the highest values of STI were observed for genotypes 66, 69, 70, 44 and 38, and the lowest values for genotypes 1, 34, 27, 15 and 18. The persistence of cocksfoot varied significantly among genotypes under both irrigation regimes (Fig. 5a, b); genotypes 30, 3 and 23 had the highest persistence under normal irrigation, and genotypes 33, 23 and 38 had the highest persistence under deficit irrigation. Genotypes managed for forage production generally had better persistence than genotypes managed for seed production. Comparing irrigation treatments, 34 genotypes tested under the normal irrigation regime and 22 under deficit irrigation were less persistent after the prolonged drought of 2016.

|

|
In year 4 of the experiment (2016), after withholding irrigation and then rewatering, 16 genotypes were completely dehydrated and had died in the forage-management treatment with a deficit irrigation regime. By contrast, all of the genotypes recovered and survived in the seed-management treatment with deficit irrigation and in the seed- and forage-management treatments with normal irrigation, (Supplementary Materials table S4).
Under the deficit irrigation regime, DMY cuts 1–3 and DWG had significant and positive correlations with RY, DR and PR and negative correlations with persistence; these correlations also existed under normal irrigation, except for DMY cut 2 with RY and PR and DWG with persistence under normal irrigation (Table 3). Under deficit irrigation regime, DEE, DTA, crown diameter and persistence had significant and negative correlations with RY, DR and PR. Moreover, DTA had negative correlations with RY and DR under normal irrigation and a positive correlation with persistence under both normal and deficit irrigation (Table 3). STI had a significant and positive correlation with persistence under both irrigation regimes (data not shown); therefore it is possible to identify genotypes having high values of persistence and drought tolerance. In this respect, some genotypes including 1, 2, 12, 14, 19, 22, 23 and 33 were identified as superior (Fig. 4a, b).

|
The PCA revealed that the first two components accounted for >48% and 64% of the total variation under normal and deficit irrigation, respectively. Under the normal irrigation (Fig. 6a), the first principal component (PC1) had positive correlations with DEE, DTA, percentage DMY cut 3 and persistence, and negative correlations with plant height, number of stems, DMY cuts 1–3, DWG, RY, DR, PR and STI. Hence, PC1 was named ‘maturity component’ (Supplementary Materials table S5). The second principal component (PC2), which had positive correlations with crown diameter, RY, RR, PR and persistence, and negative correlation with STI, was considered to represent ‘survival and recovery vigour’ (Supplementary Materials table S5). Selection of genotypes with low PC1 and high PC2 would increase drought tolerance and improve the capability of survival and recovery of cocksfoot. In this respect, genotypes 2, 3, 4, 22, 23 and 24 from Iran, genotype 6 from Hungary and genotype 29 from the Netherlands were superior, with high yield and high recovery. Genotypes 32, 35, 39, 43, 46, 59, 71 and 72 from Iran and genotypes 41 and 64 from Hungary had high yield and low recovery. Finally, genotypes 1, 8, 11, 20, 27, 28, 34, 36 and 66 from Iran, genotypes 13, 15 and 44 from Hungary, and genotype 31 from the Netherlands showed low yield and low recovery.

|
Under the deficit irrigation regime (Fig. 6b), PC1 had positive correlations with DEE, DTA, percentage DMY cut 3 and persistence, and negative correlations with plant height, number of stems, DMY cuts 1–3, DWG, RY, DR, RR, PR and STI; therefore it was considered the maturity component (Supplementary Materials table S5). PC2 had positive correlations with percentage DMY cuts 1 and 2, RR and persistence; hence it was considered to represent survival and persistence vigour (Supplementary Materials table S5). Selection of genotypes with low PC1 and high PC2 can improve drought tolerance, survival and persistence of cocksfoot and would develop earliness. In this respect, genotypes 2, 9, 23, 48, 63 and 67 from Iran and genotype 33 from Hungary were the superior genotypes with high yield and high recovery. Genotypes 25, 37, 50, 57 and 72 from Iran and genotypes 7, 14, 41, 45, 55, 60, 62 and 65 from Hungary had high yield and low recovery. Finally, genotypes 38, 49, 59, 66 and 69 from Iran, genotypes 44, 58, 61 and 64 from Hungary, and genotype 68 from the Netherlands showed low yield and low recovery.
Discussion
Drought tolerance in terms of plant productivity has been investigated extensively in many crop species, but dehydration tolerance or drought survival in perennial forage grasses has received limited attention. Improving the persistence and survivability of perennial species is vital in regions with prolonged periods of drought. In addition, rapid recovery of damaged plant tissues and the regrowth of new tissues following prolonged drought stress are important in perennial grass management (Chai et al. 2010). This study examined the persistence and regrowth of cocksfoot genotypes over 4 years under two management treatments, one to maximise seed production and the other to maximise forage production. Significant differences were observed among genotypes for all measured traits, suggesting that there is considerable genotypic variation in the studied germplasm. This can facilitate selection of genotypes with variable drought tolerance, recovery after severe drought and persistence.
In the present study, under the deficit irrigation regime, the forage-management treatment showed more reduction in recovery after prolonged drought than the seed-management treatment. Because the water deficit applied was identical in the two management treatments before withholding irrigation, it can be said that early harvest of plants as forage at the first cut increased the impact of drought stress, weakening the plant vigour and therefore causing greater reduction of recovery in the forage-management treatment with deficit irrigation. By contrast, delaying the first harvest of the seed-management treatment long enough to maximise seed production has reduced the impact of drought and increased the tolerance of these genotypes to deficit irrigation.
The range of persistence in crop species depends on factors such as the plant genotypes, management (e.g. fertiliser, grazing management, weed and pest control, irrigation) and environmental factors (e.g. rainfall, light, temperature, soil type) (Cullen et al. 2005; Nie et al. 2008). In the present study, persistence of genotypes decreased in the deficit irrigation regime compared with normal irrigation. It seems that reduction of persistence due to deficit irrigation can be associated with reduced crown diameter and possibly fewer stems, which reduce plant vigour over the years. Moreover, the forage-management treatment showed positive persistence, whereas the seed-management treatment mostly showed negative persistence. Persistence of perennial grasses relies on the maintenance of tiller populations. If each tiller is not replaced in the following growing season, the population declines. Tiller density is therefore an important indicator of population survival (Cullen et al. 2005). Moreover, Culvenor and Simpson (2014, 2016) stated that grazing management can be used as a strategy for improvement of persistence in perennial grasses. They concluded that persistence and continuing productivity will be better ensured in the face of the likely concurrence of stresses such as drought and overgrazing through the use of genotypes with grazing and/or soil stress-tolerance traits combined with good management.
Results of the recovery experiment revealed that all of the recovery-related traits declined under the deficit irrigation regime compared with normal irrigation. Because the water deficit was applied identically in all three years before withholding irrigation, it can be concluded that prolonged drought stress reduced the persistence and recovery of plants (Saeidnia et al. 2017). However, high genetic variation was found among the genotypes for all of the recovery-related traits, indicating good potential for genetic study of these traits and possibility of selection of genotypes with variable recovery and survival vigour in this germplasm collection.
Information on the association of morphological traits with recovery, persistence and drought tolerance is very limited in cocksfoot. Positive associations were observed between recovery after prolonged drought stress and dry forage yield and its components under both normal and deficit irrigation, indicating that more productive genotypes have better recovery vigour. Similar results were found in tall fescue (Festuca arundinacea) (Pirnajmedin et al. 2017). Moreover, DTA had significant negative correlation with recovery and positive correlation with persistence under both irrigation regimes. This indicates that a decrease in flowering time is associated with a decrease in persistence and increase in post-drought recovery. These results were expected because a decrease in flowering time maintains the vigour of plants during their lifetime and therefore leads to improved plant recovery.
The distribution of genotypes on the biplot of PCA indicated that, under both irrigation regimes, the analysis has separated seed- and forage-management genotypes. The results of PCA revealed negative correlations between phenological traits (DEE and DTA) and post-drought recovery under both normal and deficit irrigation regimes. This suggests that selection for earliness may indirectly improve recovery of this germplasm, which is consistent with the results of Volaire and Lelièvre (1997). The results also showed positive correlations of STI with recovery-related traits under both irrigation regimes. These findings demonstrate that drought-tolerant genotypes have better recovery after prolonged drought stress. Results of PCA also verified positive correlations of phenological traits with persistence. This suggests that selection for late-flowering genotypes may identify the more persistent ones. Previous studies indicated that early-flowering ecotypes of cocksfoot had more ability to survive severe drought (Volaire et al. 1998; Shaimi et al. 2009). Based on the distribution of studied genotypes on the biplot of PCA, genotypes with higher forage production, better recovery and superior persistence and drought tolerance were identified.
In conclusion, considerable genetic variation was observed for forage productivity and its components, and persistence and recovery-related traits, under both normal and deficit irrigation regimes, indicating high potential for improving these traits through targeted selection in breeding programs. Persistence and post-drought recovery were reduced in the deficit irrigation regime. These reductions were lower in the seed-management treatment than the forage-management treatment. Under normal irrigation, forage management showed higher recovery than seed management, whereas under deficit irrigation, seed management showed higher recovery than forage management. Moreover, the seed-management treatment had lower persistence than the forage-management treatment. Results of this study also showed that the early-flowering genotypes were capable of more regrowth and better recovery. DWG and dry forage yield and its components were associated with post-drought recovery under both irrigation regimes and could be useful criteria for identifying suitable genotypes for arid and semi-arid regions. Based on the association of STI with persistence, and applying PCA method, genotypes 2, 19, 22 and 23 from Iran and genotype 33 from Hungary were identified as the superior genotypes. They combined higher forage production, better recovery and superior persistence and drought tolerance, and therefore can be recommended for the development of synthetic cultivars.
Conflicts of interest
The authors declare no conflicts of interest.
References
Allen RG Pereira LS Raes D Smith M (1998 ) ‘Crop evapotranspiration: guidelines for computing crop requirements.’ FAO Irrigation and Drainage Paper No. 56. (Food and Agriculture Organization of the United Nations: Rome)Annicchiarico P, Pecetti L, Bouzerzour H, Kallida R, Khedim A, Porqueddu C, Simões NM, Volaire F, Lelièvre F (2011) Adaptation of contrasting cocksfoot plant types to agricultural environments across the Mediterranean basin. Environmental and Experimental Botany 74, 82–89.
| Adaptation of contrasting cocksfoot plant types to agricultural environments across the Mediterranean basin.Crossref | GoogleScholarGoogle Scholar |
Chai Q, Jin F, Merewitz E, Hung B (2010) Growth and physiological traits associated with drought survival and post- drought recovery in perennial turfgrass species. Journal of the American Society for Horticultural Science 135, 125–133.
Clarke Topp C, Parkin GW, Ferre TPA (2008) Soil water content. In ‘Soil sampling and methods of analysis’. (Eds MR Carter, EG Gregorich) pp. 939–961. (Canadian Society of Soil Science: Pinawa, MB, Canada)
Cullen BR, Chapman DF, Quigley PE (2005) Persistence of Phalaris aquatica in grazed pastures. Plant and tiller population characteristics. Australian Journal of Experimental Agriculture 45, 41–48.
| Persistence of Phalaris aquatica in grazed pastures. Plant and tiller population characteristics.Crossref | GoogleScholarGoogle Scholar |
Culvenor RA, Simpson RJ (2014) Persistence traits in perennial pasture grasses: the case of phalaris (Phalaris aquatica L.). Crop & Pasture Science 65, 1165–1176.
| Persistence traits in perennial pasture grasses: the case of phalaris (Phalaris aquatica L.).Crossref | GoogleScholarGoogle Scholar |
Culvenor RA, Simpson RJ (2016) Interaction of plant genotype and management in the persistence of a perennial grass exposed to grazing and soil fertility stresses. Grass and Forage Science 71, 540–558.
| Interaction of plant genotype and management in the persistence of a perennial grass exposed to grazing and soil fertility stresses.Crossref | GoogleScholarGoogle Scholar |
Dowling PM, Michalk DL, Kemp DR, Millar GD, Priest SM, King WM, Packer IJ, Holst PJ, Tarleton JA (2006) Sustainable grazing systems for the Central Tablelands of New South Wales. 2. Effect of pasture type and grazing management on pasture productivity and composition. Australian Journal of Experimental Agriculture 46, 457–469.
| Sustainable grazing systems for the Central Tablelands of New South Wales. 2. Effect of pasture type and grazing management on pasture productivity and composition.Crossref | GoogleScholarGoogle Scholar |
Fernandez GCJ (1992) Effective selection criteria for assessing plant stress tolerance. In ‘Proceedings International Symposium on Adaptation to Food Crops to Temperature and Water Stress’. (Ed. CC Kuo) pp. 257–270. (Asian Vegetable Research and Development Center: Shanhua, Taiwan)
Hu H, Xiong L (2014) Genetic engineering and breeding of drought-resistant crops. Annual Review of Plant Biology 65, 715–741.
| Genetic engineering and breeding of drought-resistant crops.Crossref | GoogleScholarGoogle Scholar |
Jiang LF, Qi X, Zhang XQ, Huang LK, Ma X, Xie WG (2014) Analysis of diversity and relationships among orchardgrass (Dactylis glomerata L.) accessions using start codon-targeted markers. Genetics and Molecular Research 13, 4406–4418.
| Analysis of diversity and relationships among orchardgrass (Dactylis glomerata L.) accessions using start codon-targeted markers.Crossref | GoogleScholarGoogle Scholar |
Johnson RA, Wichern DW (2007) ‘Applied multivariate statistical analysis.’ (Prentice Hall International: Upper Saddle River, NJ, USA)
Kanapeckas J, Lemeziene N, Stukonis V, Tarakanovas P (2008) Drought tolerance of turfgrass genetic resources. Biologija 54, 121–124.
| Drought tolerance of turfgrass genetic resources.Crossref | GoogleScholarGoogle Scholar |
Karcher DE, Richardson MD, Hignight K, Rush D (2008) Drought tolerance of tall fescue populations selected for high root/shoot ratios and summer survival. Crop Science 48, 771–777.
| Drought tolerance of tall fescue populations selected for high root/shoot ratios and summer survival.Crossref | GoogleScholarGoogle Scholar |
Kemp DR, Michalk DM, Virgona JM (2000) Towards more sustainable pastures: lessons learnt. Australian Journal of Experimental Agriculture 40, 343–356.
| Towards more sustainable pastures: lessons learnt.Crossref | GoogleScholarGoogle Scholar |
Lobell DB, Roberts MJ, Schlenker W, Braun N, Little BB, Rejesus RM, Hammer GL (2014) Greater sensitivity to drought accompanies maize yield increase in the U.S. Midwest. Science 344, 516–519.
| Greater sensitivity to drought accompanies maize yield increase in the U.S. Midwest.Crossref | GoogleScholarGoogle Scholar |
Nguyen HT, Sleper DA (1983) Theory and application of half sib matings in forage grass breeding. Theoretical and Applied Genetics 64, 187–196.
| Theory and application of half sib matings in forage grass breeding.Crossref | GoogleScholarGoogle Scholar |
Nie ZN, Miller S, Moore GA, Hackney BF, Boschma SP, Reed KFM, Mitchell M, Albertsen TO, Clark S, Craig AD, Kearney G, Li GD, Dear BS (2008) Field evaluation of perennial grasses and herbs in southern Australia. Persistence, root characteristics and summer activity. Australian Journal of Experimental Agriculture 48, 424–435.
| Field evaluation of perennial grasses and herbs in southern Australia. Persistence, root characteristics and summer activity.Crossref | GoogleScholarGoogle Scholar |
Norton MR, Volaire F, Lelievre F (2006) Summer dormancy in Festuca arundinacea Schreb; the influence of season of sowing and a simulated mid-summer storm on two contrasting cultivars. Australian Journal of Agricultural Research 57, 1267–1277.
| Summer dormancy in Festuca arundinacea Schreb; the influence of season of sowing and a simulated mid-summer storm on two contrasting cultivars.Crossref | GoogleScholarGoogle Scholar |
Pecetti L, Annicchiarico P, Abdelguerfi A, Kallida R, Mefti M, Porqueddu C, Simo˜es NM, Volaire F, Lelie’vre F (2011) Response of Mediterranean tall fescue cultivars to contrasting agricultural environments and implications for selection. Journal of Agronomy & Crop Science 197, 12–20.
| Response of Mediterranean tall fescue cultivars to contrasting agricultural environments and implications for selection.Crossref | GoogleScholarGoogle Scholar |
Pirnajmedin F, Majidi MM, Gheysari M (2015) Root and physiological characteristics associated with drought tolerance in Iranian tall fescue. Euphytica 202, 141–155.
| Root and physiological characteristics associated with drought tolerance in Iranian tall fescue.Crossref | GoogleScholarGoogle Scholar |
Pirnajmedin F, Majidi MM, Saeidi G, Gheysari M, Volaire F, Barre P, Osivand AH, Sarfaraz D (2017) Persistence, recovery and root traits of tall fescue genotypes with different flowering date under prolonged water stress. Euphytica 213, 269
| Persistence, recovery and root traits of tall fescue genotypes with different flowering date under prolonged water stress.Crossref | GoogleScholarGoogle Scholar |
Saeidnia F, Majidi MM, Mirlohi A, Shahidaval S (2016) Selection for productivity, persistence and drought tolerance in orchardgrass. Euphytica 212, 111–130.
| Selection for productivity, persistence and drought tolerance in orchardgrass.Crossref | GoogleScholarGoogle Scholar |
Saeidnia F, Majidi MM, Mirlohi A, Manafi M (2017) Productivity, persistence and traits related to drought tolerance in smooth bromegrass. Plant Breeding 136, 270–278.
| Productivity, persistence and traits related to drought tolerance in smooth bromegrass.Crossref | GoogleScholarGoogle Scholar |
Saeidnia F, Majidi MM, Mirlohi A (2018) Genetic potential to improve seed and forage yield simultaneously in smooth bromegrass under water deficit conditions. Euphytica 214, 42
| Genetic potential to improve seed and forage yield simultaneously in smooth bromegrass under water deficit conditions.Crossref | GoogleScholarGoogle Scholar |
Sanada Y, Gras MC, Van Santen E (2010) Cocksfoot. In ‘Fodder crops and amenity grasses. Handbook of plant breeding’. (Eds BU Boller, K Posselt, DF Veronesi) pp. 317–328. (Springer: New York)
Shaimi N, Kallida R, Volaire F, Saidi N, Faiz CA (2009) Summer dormancy and drought survival of Moroccan ecotypes of orchardgrass. Crop Science 49, 1416–1424.
| Summer dormancy and drought survival of Moroccan ecotypes of orchardgrass.Crossref | GoogleScholarGoogle Scholar |
Steel RGD, Torrie JG (1980) ‘Principles and procedures of statistics.’ (McGraw–Hill Book Company: New York)
Stewart AV, Ellison NW (2011) Dactylis. In ‘Wild crop relatives: Genomic and breeding resources: Millets and grasses’. (Ed. C Kole) pp. 73–87. (Springer: Berlin)
Touchette BW, Iannacone LR, Turner GE, Frank AR (2007) Drought tolerance versus drought avoidance: a comparison of pant-water relations in herbaceous wetland plants subjected to water withdrawal and repletion. Wetlands 27, 656–667.
| Drought tolerance versus drought avoidance: a comparison of pant-water relations in herbaceous wetland plants subjected to water withdrawal and repletion.Crossref | GoogleScholarGoogle Scholar |
Van Minnebruggen A, Roldan-Ruiz I, Van Bockstaele E, Haesaert G, Cnops G (2015) The relationship between architectural characteristics and regrowth in Trifolium pratense (red clover). Grass and Forage Science 70, 507–518.
| The relationship between architectural characteristics and regrowth in Trifolium pratense (red clover).Crossref | GoogleScholarGoogle Scholar |
Volaire F, Lelièvre F (1997) Production, persistence and water-soluble carbohydrate accumulation in 21 contrasting populations of Dactylis glomerata L. subjected to severe drought in the south of France. Australian Journal of Agricultural Research 48, 933–944.
| Production, persistence and water-soluble carbohydrate accumulation in 21 contrasting populations of Dactylis glomerata L. subjected to severe drought in the south of France.Crossref | GoogleScholarGoogle Scholar |
Volaire F, Thomas H, Bertagne N, Bourgeois E, Gautier MF, Lelievre F (1998) Survival and recovery of perennial forage grasses under prolonged Mediterranean drought. II. Water status, solute accumulation of dehydrin transcripts in bases of immature leaves. New Phytologist 140, 451–460.
| Survival and recovery of perennial forage grasses under prolonged Mediterranean drought. II. Water status, solute accumulation of dehydrin transcripts in bases of immature leaves.Crossref | GoogleScholarGoogle Scholar |
Volaire F, Barkaoui K, Norton MR (2014) Designing resilient and sustainable grasslands for a drier future: adaptive strategies, functional traits and biotic interactions. European Journal of Agronomy 52, 81–89.
| Designing resilient and sustainable grasslands for a drier future: adaptive strategies, functional traits and biotic interactions.Crossref | GoogleScholarGoogle Scholar |
Wang Z, Huang B (2004) Physiological recovery of Kentucky bluegrass from simultaneous drought and heat stress. Crop Science 44, 1729–1736.
| Physiological recovery of Kentucky bluegrass from simultaneous drought and heat stress.Crossref | GoogleScholarGoogle Scholar |
Zlatev ZS (2005) Effects of water stress on leaf water relations of young bean plants. Journal of Central European Agriculture 6, 5–14.