

Brahman and Brahman crossbred cattle grown on pasture and in feedlots in subtropical and temperate Australia. 1. Carcass quality
K. M. Schutt A B D E , H. M. Burrow A , J. M. Thompson A C and B. M. Bindon AA Cooperative Research Centre for Beef Genetic Technologies, C.J. Hawkins Homestead, University of New England, Armidale, NSW 2351, Australia.
B CSIRO Livestock Industries, PO Box 5545, Rockhampton Mail Centre, Qld 4702, Australia.
C Division of Animal Science, University of New England, Armidale, NSW 2351, Australia.
D Present address: ‘Ingaby Station’, St George, Qld 4487, Australia.
E Corresponding author. Email: sk_prewett@activ8.net.au
Animal Production Science 49(6) 426-438 https://doi.org/10.1071/EA08081
Submitted: 3 March 2008 Accepted: 12 February 2009 Published: 13 May 2009
Abstract
Brahmans are known to have poorer carcass quality relative to Bos taurus breeds and crossbreds under temperate environments; however, little is known of their performance in subtropical environments. The Cooperative Research Centre for Cattle and Beef Industry (Meat Quality) initiated a crossbred progeny test experiment to compare straightbred Brahmans with Brahman crossbreds finished on pasture and grain, in subtropical and temperate environments, to carcass quality specifications of Australian domestic and export markets. Brahman, Belmont Red, Santa Gertrudis, Angus, Hereford, Shorthorn, Charolais and Limousin sires were mated to Brahman females in subtropical Queensland over 3 years to produce 1750 progeny. At a common age at slaughter, Charolais crossbreds had the highest hot carcass weight (CWT) but were not significantly heavier than Limousin or British crossbred progeny. At common carcass weights, breeds within breed type (British, Continental, tropically adapted) performed similarly. British and Santa Gertrudis crossbreds had the fattest carcasses and lowest yields. British and Belmont Red crossbreds had the highest intramuscular fat percentage (IMF). Continental crossbreds had the highest retail beef yield (RBY), kilograms of retail primals (RTPM) and percent retail primals (pcRTPM) and leanest carcasses. Brahmans had the lowest CWT, intermediate subcutaneous fat cover, high yields and low IMF. Animals finished in the subtropics on pasture were significantly older, leaner and had higher RBY, RTPM and pcRTPM than subtropical feedlot-finished contemporaries. Temperate feedlot animals had significantly more IMF, less subcutaneous fat at the P8 site and slightly lower yields than subtropical feedlot contemporaries, indicating possible effects of postweaning growth path on fat distribution. Belmont Red crossbreds demonstrated the advantages of adaptation with the highest IMF in both subtropical finishing regimes, while Angus progeny had the highest IMF in the temperate feedlot environment and highest IMF overall when analysed across finishing regimes. Significant interactions were mainly the result of scale effects rather than breed re-ranking for carcass traits across markets and finishing regimes. Therefore, breeds that performed well for certain carcass traits in subtropical environments performed consistently for those traits in temperate environments relative to other sire breeds, regardless of market endpoint or finishing nutrition.
Introduction
Beef eating quality and the consistency of its supply are key issues for Australian domestic and export markets (Bindon 2001; Egan et al. 2001). Despite declining consumption of beef, the domestic market remains the largest single market for Australian beef, commanding 37% of total beef production (Bindon and Jones 2001). The gross value of cattle meat product to the Australian economy in 2004–05 was AU$7.8 billion (Australian Bureau of Statistics 2007). Australia is the world’s largest beef exporter with export markets consuming 43% (891 700 t) of Australia’s bone-out beef in 2005–06 (Australian Bureau of Statistics 2007). Japan is Australia’s most important premium export beef market consuming 19.7% of total beef production in 2005–06, whereas South Korea is the second largest premium export beef market (Australian Bureau of Statistics 2007). By the mid 1980s, the USA realised the potential of the Japanese and South Korean beef markets and has been a major competitor with Australia for these markets ever since. This has forced Australian beef producers to become consumer-driven, focussing on consistency of quality and reliability of supply to maintain market share.
The majority of Australia’s 27.5 million beef cattle are produced in northern states (Queensland 42%, Northern Territory 16%; Australian Bureau of Statistics 2006). Production potential cannot be fully expressed unless animals are highly adapted to their production environment (Frisch 1997). Brahmans and Brahman crossbreds predominate in the tropical, subtropical and grassland regions of northern Australia because of their superior adaptation in harsh environments. Consequently, more than 50% of the national herd is estimated to have some Brahman genes (Bindon 2002). By the late 1980s and early 1990s, Bos indicus × Bos taurus types were supplying most of the export trade from Queensland (Ball and Johnson 1989). However, growth rates were significantly affected by seasonal fluctuations in pasture quality and supply, causing periods of fast weight gain during the wet season and weight stasis or loss during the dry season. This resulted in up to 30% of animals failing to meet target liveweights for Japanese and Korean markets by winter in many finishing areas of Queensland (Jeffery et al. 1993). As a consequence, the feedlot sector expanded in northern Australia to improve growth rates, reduce age at turnoff and improve carcass compliance with market specifications. However, new problems were arising for the northern Australian beef industry, with Brahmans developing a reputation for poor carcass and meat quality and poor performance in feedlots relative to B. taurus breeds and crossbreds, threatening their premium markets.
Considerable research has been reported on British, Continental and Brahman crossbreds grown in temperate environments, one of the most well known and comprehensive research projects being the ongoing Germplasm Evaluation Program conducted at the United States Department of Agriculture (USDA) Meat Animal Research Centre at Clay Center in Nebraska (Cundiff et al. 1998, 2001). However, there is limited research comparing carcass quality of straightbred Brahmans with Brahman × British and Brahman × Continental crossbreds finished in tropical and subtropical environments. Where carcass quality of Brahman × British and Brahman × Continental breeds have been reported in such environments (Peacock et al. 1977; DeRouen et al. 1992) the temperature extremes and rainfall patterns differ significantly from Australian subtropical and grassland environments. Further, Australian cattle production systems, finishing regimes, processing practices and markets differ significantly from those of other countries.
Consequently, the Cooperative Research Centre for Cattle and Beef Industry (Meat Quality) (Beef CRC) initiated the Northern Crossbreeding Program in 1994 to examine genetic and non- genetic factors influencing beef quality. The Crossbreeding Program aimed to compare the carcass quality characteristics of straightbred Brahmans relative to different Brahman crossbred genotypes when finished on pasture and grain, in subtropical and temperate environments, that were best suited to beef quality specifications of Australian domestic and export markets. Preliminary results were reported by Newman et al. (1998, 1999, 2002). The purpose of the present paper is to report the final carcass quality results and their implications for the Australian beef industry. The following two companion papers (Schutt et al. 2009a, 2009b) report feed efficiency and meat quality results from this experiment.
Materials and methods
Experimental design and animals
The Beef CRC Northern Crossbreeding Program was designed to determine genetic and non-genetic factors influencing carcass and meat quality of Brahman and first-cross animals bred in a subtropical environment, and finished on pasture and grain in subtropical and temperate environments. Animals were grown to Australia’s three main market endpoints, being domestic, Korean and Japanese. Full details of the experimental design, measurements and data storage were described by Upton et al. (2001).
Briefly, 1000 Brahman females were mated to produce 1750 Brahman and first-cross progeny over three joinings at two properties, ‘Duckponds’ and Brigalow Research Station, both in subtropical central Queensland. The eight sire breeds selected for the experiment were based on common breeds used in industry. They represented B. indicus (Brahman – purebred control), B. indicus × British-derived (Santa Gertrudis), B. taurus × Sanga-derived (Belmont Red), B. taurus – British (Angus, Hereford, Shorthorn) and B. taurus – Continental (Charolais, Limousin). Brahman, Santa Gertrudis and Belmont Red are tropically adapted breeds.
Selection of sires was primarily by the collaborating seedstock breeders in the Beef CRC Straightbreeding Program, as described by Upton et al. (2001). To compare sire performance relative to industry herds, all sires used in the program were performance-recorded through their breed’s GROUP BREEDPLAN. At the time of selection, carcass and meat quality estimated breeding values (EBV) were not available and hence selection of sires is assumed to have been made without regard for these traits. Common sires were used across the Beef CRC Straightbreeding and Crossbreeding Programs to generate genetic linkages between the programs. Table 1 (derived from table 5 in Upton et al. 2001) shows EBV for growth, fertility and carcass attributes for all sires used in the Crossbreeding Program. It also includes the breed average EBV for animals born in 1997 for each trait, listed in parentheses, showing that sires used in the Crossbreeding Program were close to their specific breed averages for most traits.

|
Artificial insemination (AI) was used to generate the majority of the crossbred progeny, followed by natural mating with back-up bulls of a different breed. To prevent confounding of calves by AI and natural mating, natural mating programs were conducted at the same time as the AI programs. British breeds were not used as back-up sires due to their inability to cope with northern environmental stressors. A full description of AI programs and parentage determination is given by Corbet et al. (1997, 1999). The aim was to produce ~10 steers and 10 heifers per sire. The optimal number of sires, offspring per sire and number of link sires between herds and allocation of animals to treatment combinations are discussed by Robinson (1995). Newman et al. (2002) estimated mean sire EBV (expressed as deviations from the contemporary group) to assess the randomness of sire allocations across contemporary groups, and found that sires were effectively randomly allocated. Table 2 shows the number of sires per breed and the number and sex of progeny generated per breed.
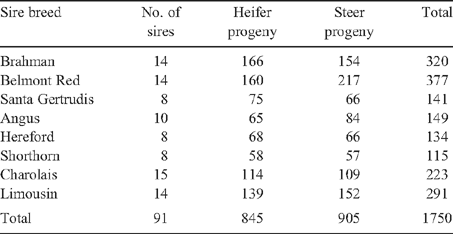
|
Treatments
All male calves were castrated at about 4 months of age. Calves were weaned in 1996, 1997 and 1998 at about 6 months of age. At weaning, calves born at Brigalow Research Station were transferred to ‘Duckponds’. Equal numbers of calves of each sire were assigned by sex, age and weight to one of three market endpoints (domestic, Korean, Japanese), one of two finishing environments (subtropical, temperate), and one of two finishing diets (pasture, feedlot) (Robinson 1995). The finishing treatments are combined and identified as subtropical pasture, subtropical feedlot and temperate feedlot throughout the manuscript. No animals were finished on pasture in the temperate environment. Only steers were finished to Japanese market weights. Following weaning, one-third of the calves were transferred to temperate pasture at Glen Innes Research Station (native and introduced pastures, refer to Ayres et al. (2001)) or ‘Tullimba’ (native wallaby grass, Austrodanthonia spp.) in north-eastern New South Wales to grow-out to feedlot entry weights. In the temperate environment at Glen Innes Research Station, mean daily temperatures ranged from 12.1 to 25.2°C in summer and 0.7 to 13.9°C in winter, with mean relative humidity ranging from 65 and 81% and an average annual rainfall of 848 mm (latitude –29.6952S, longitude 151.6936E, elevation 1060 m; Bureau of Meteorology 2006). In the temperate environment at ‘Tullimba’, mean daily temperatures ranged from 13.4 to 30.3°C in summer and 0.1 to 16.5°C in winter, with an average annual rainfall of 766 mm (latitude –30.1711S, longitude 151.0757E, elevation 654 m; Bureau of Meteorology 2006). After weaning, calves destined for the two subtropical finishing regimes were grown out on buffel grass pasture (Cenchrus ciliaris) at ‘Duckponds’. Mean daily temperatures ranged from 20.7 to 34.8°C in summer and 6.9 to 24.8°C in winter, with mean relative humidity ranging from 33 and 69% and an average annual rainfall of 640 mm (latitude –23.5694S, longitude 148.1756E, elevation 190 m; Bureau of Meteorology 2006).
Animals assigned to feedlot finishing were transferred to their respective feedlots when they reached liveweights of 300 kg for domestic and 400 kg for export markets. Finishing regimes included buffel grass pasture at ‘Duckponds’ (subtropical pasture), ‘Goonoo’ feedlot (subtropical feedlot) or ‘Tullimba’ feedlot (temperate feedlot). Domestic animals were fed for an average of 60 days, whereas export animals were grain fed for a minimum of 90 days (Upton et al. 2001). Diets between feedlots were kept as similar as possible, though commercial constraints resulted in minor differences in diets over the experimental period.
A cohort was defined by a combination of sex, year of birth, market endpoint and finishing regime. All animals within a cohort were managed as a single group during grow-out, finishing, pre-slaughter and slaughter, with the exception of a few cohorts that were slaughtered over 2 consecutive days to facilitate measurement of retail beef yield (RBY). All animals within a cohort were slaughtered when the average weight of their group reached estimated carcass weights of 220, 280 and 330 kg for domestic, Korean and Japanese markets respectively. One-half of all steers finished in the subtropics received repeated implants of 20 mg oestradiol-17β (Compudose 100, Elanco Animal Health, Macquarie Park, NSW, Australia), a hormonal growth promotant (HGP), from ~15 months of age to slaughter. The numbers of implants given to feedlot-finished steers were one, three and four for domestic, Korean and Japanese markets respectively. The numbers of implants given to pasture-finished steers were two, five and seven for domestic, Korean and Japanese markets respectively. Experimental details and results for the HGP experiment were reported by Hunter et al. (2001).
Slaughter protocols and carcass measurements
Details of the pre- and post-slaughter protocols, and carcass and meat quality measurements were reported by Perry et al. (2001). Best practice pre- and post-slaughter procedures were applied. The left side of each carcass was used for sample collection and yield measurements. Measurements of hot carcass weight (CWT) and hot subcutaneous fat depth at the P8 site (HOTP8) were taken before chilling. Carcasses were placed in chillers within 1 h of slaughter and hung by the Achilles tendon. Carcasses were quartered 20–24 h after slaughter. Abattoir personnel broke the carcass down to 17 boneless primal cuts and recorded untrimmed weights. Primals were then prepared to commercial specifications as described by Perry et al. (2001). The RBY was calculated as described in Table 3. Due to disruption of commercial processing procedures, not all animals were measured for every carcass trait, resulting in different numbers of animals having records for the various traits. After July 1997, abattoir conditions precluded full yield determination on all animals, with crossbred animals measured for yield at 3 mm fat cover only (Perry et al. 2001). From early 1998 a modified yield measurement protocol was used because of disruptions to the commercial chain. Hence, RBY and kilograms of retail primals (RTPM) were only recorded for the 1996 calf crop. Intramuscular fat percentage (IMF) was determined for the M. longissimus thoracis et lumborum using solvent extraction with Soxhlet apparatus or near infrared spectrophotometry (Perry et al. 2001). Traits reported in this paper are described in Table 3.

|
Statistical analyses
Least squares means for fixed effects were estimated using the generalised linear model (GLM) procedure of SAS (SAS 2000). Initially a mixed model procedure was used fitting fixed effects of breed, market, finish, sex, year of birth, herd of origin and HGP treatment. Partial confounding of fixed effects and slaughter groups and missing subclasses prevented the model from converging. A second model was developed where each independent variable was fitted separately, and a second independent variable accounting for all remaining fixed effects was included. Sire nested within breed was fitted as a random effect in the breed analysis and was also used as the error term to test breed differences. The models ignored abattoir and slaughter day effects because slaughter day was completely nested within market weight and finishing regime. Abattoir was confounded within finishing environment (subtropical, temperate). The multiple range test devised by Duncan (1955) and extended by Kramer (1957) was used to test for significant differences between means.
Beef markets dictate that animals are sent to slaughter based on liveweight and subsequent carcass weight. Therefore, CWT was used as the covariate to adjust all other carcass traits to a common basis for comparison. Data were analysed across sex. Age at slaughter (AGE), HOTP8, IMF, RBY, RTPM and percent retail primals (pcRTPM) were adjusted to a common CWT for breed and finish effects and a common CWT within market endpoint for market and sex within market effects. British crossbreds were older at slaughter than other genotypes, with the exception of Santa Gertrudis crosses, as they were all bred by AI (P < 0.05). Consequently, CWT was adjusted for AGE for breed effects only. CWT was unadjusted for finish, market and sex within market effects. Steers implanted with HGP were excluded from analyses for finish and sex effects due to confounding with location and sex. Subsequently, only 1473 animals were analysed for finish and sex effects. The final models were:
Sire breed effects on CWT:
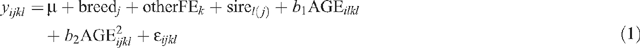
where yijkl is the observation for a dependent variable for animal i, μ is the overall mean, breedj is the effect of the jth sire breed, otherFEk is the effect of the kth group that accounts for all other fixed effects concatenated into one variable (sex || market || finish || year of birth || herd of origin || HGP treatment), sirel(j) is the random effect of the lth sire nested within the jth breed, AGEijkl is effect of age at slaughter fitted as a quadratic covariate, and ॉijkl is the residual error term.
Sire breed effects on AGE, HOTP8, IMF, RBY, RTPM and pcRTPM:
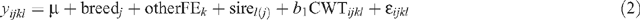
where yijkl is the observation for a dependent variable for animal i, μ is the overall mean, breedj is the effect of the jth sire breed, otherFEk is the effect of the kth group that accounts for all other fixed effects concatenated into one variable (sex || market || finish || year of birth || herd of origin || HGP treatment), sirel(j) is the random effect of the lth sire nested within the jth breed, CWTijkl is the linear effect of carcass weight of the animal fitted as a covariate, and ॉijkl is the residual error term.
Finishing regime effects on CWT:

where yijk is the observation for a dependent variable for animal i, μ is the overall mean, finishj is the effect of the jth finishing regime, otherFEk is the effect of the kth group that accounts for all other fixed effects concatenated into one variable (breed || sex || market || year of birth || herd of origin; HGP treatment was excluded), and ॉijk is the random residual error. Similar models were used to determine market endpoint and sex within market effects on CWT.
Finishing regime effects on AGE, HOTP8, IMF, RBY, RTPM and pcRTPM:
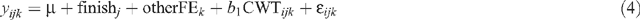
where yijk is the observation for a dependent variable for animal i, μ is the overall mean, finishj is the effect of the jth finishing regime, otherFEk is the effect of the kth group that accounts for all other fixed effects concatenated into one variable (breed || sex || market || year of birth || herd of origin; HGP treatment was excluded), CWTijk is the linear effect of carcass weight of the animal fitted as a covariate, and ॉijk is the random residual error. Similar models were used to determine market endpoint and sex within market effects on AGE, HOTP8, IMF, RBY, RTPM and pcRTPM.
Least squares means were estimated for two-way interactions between all fixed effects. Of particular interest were breed × finish, breed × market and market × finish interactions. Interactions were calculated with concatenated fixed effects models using the GLM procedure of SAS (SAS 2000). The two main effects being evaluated were fitted separately and a concatenated variable comprising all remaining fixed effects was included. Interactions for CWT were adjusted for AGE, while all other carcass traits were adjusted for CWT. For example, the model used to test the breed × finish interaction effects on CWT was:
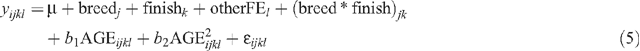
where yijkl is the observation for a dependent variable for animal i, μ is the overall mean, breedj is the effect of the jth sire breed, finishk is the effect of the kth finishing regime, otherFEl is the effect of the lth group that accounts for all other fixed effects concatenated into one variable (sex || market || year of birth || herd of origin), (breed * finish)jk is the interaction for the jth sire breed and kth finishing regime, AGEijkl is effect of age at slaughter fitted as a quadratic covariate, and ॉijkl is the random residual error.
The model used to test the breed × finish interaction effects on AGE, HOTP8, IMF, RBY, RTPM and pcRTPM was:
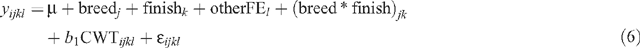
where yijkl is the observation for a dependent variable for animal i, μ is the overall mean, breedj is the effect of the jth sire breed, finishk is the effect of the kth finishing regime, otherFEl is the effect of the lth group that accounts for all other fixed effects concatenated into one variable (sex || market || year of birth || herd of origin), CWTijkl is the linear effect of carcass weight of the animal fitted as a covariate, and ॉijkl is the random residual error.
Results
Sire breed effects
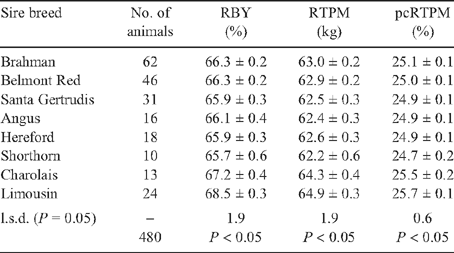
Least squares means for the effect of sire breed of animals in all markets and finishing regimes are shown in Tables 4 and 5. There were significant differences between sire breeds for all traits. Differences between breeds in CWT of more than 10 kg were significant (P < 0.05). Progeny of tropically adapted sire breeds had the lowest CWT, with Brahman carcasses weighing up to 16% less (P < 0.001) than Continental and British crossbreds. There was no difference between British and Continental crossbreds for CWT.

|
|
At common CWT, Brahman-, Belmont Red- and Continental-sired crossbreds were younger at slaughter (P < 0.05) than Santa Gertrudis and British crossbreds. This was probably due to the fact that the Santa Gertrudis and British sires were unable to be used for natural mating following the AI programs, and hence their progeny were older at slaughter than the other crosses.
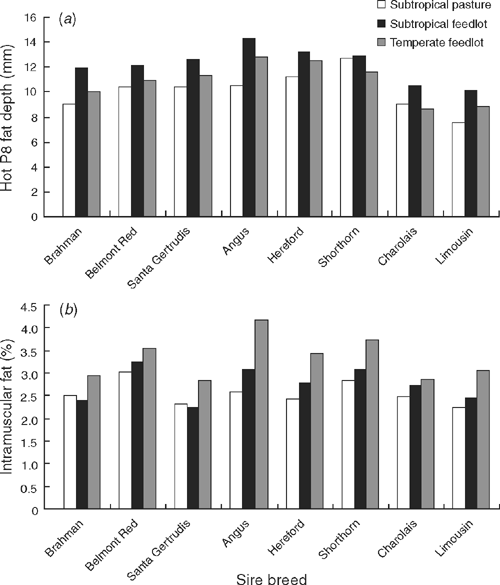
Continental crossbreds had the leanest carcasses (P < 0.001), whereas British and Santa Gertrudis sired progeny had the highest HOTP8 across markets and finishing regimes (Fig. 1a). Most of the fatness effect for British crosses was due to higher subcutaneous fat cover in heifers. Progeny by Belmont Red and Brahman sires had intermediate HOTP8 levels. British and Belmont Red crossbreds had more IMF (P < 0.001) than other genotypes across markets and finishing regimes. Brahman and Santa Gertrudis crosses had the lowest IMF but were not significantly different from the Continental crosses. Belmont Red crossbreds ranked highest for IMF in both subtropical finishing regimes (Fig. 1b). Angus crossbreds had the highest IMF overall when analysed across finishing regimes, and had 27% more IMF than the average of Santa Gertrudis crosses and straightbred Brahman progeny at common carcass weights.
|
Continental crossbreds ranked highest for the three yield traits across all three finishing regimes and markets (Table 5). Limousin crosses had higher (P < 0.05) RBY, RTPM and pcRTPM than British and tropically adapted crosses. There was no significant difference in yields between Charolais crosses and all other crosses. Brahman and Belmont Red sired progeny had intermediate yields, while Santa Gertrudis crosses were similar to the British crosses. Shorthorn sired cattle yielded 2.8% lower RBY and 2.7 kg less RTPM than Limousin sired progeny.
Pasture v. feedlot finishing
Pasture-finished animals were 210 and 229 days older at slaughter (averaged across markets; P < 0.001) than their half-sibs finished in subtropical and temperate feedlots respectively. Pasture-finished animals differed significantly from subtropical feedlot animals for most carcass traits with 2.1 mm less HOTP8, 0.9% more RBY, 1.0 kg more RTPM and 0.6% more pcRTPM (Tables 6 and 7). While pasture-finished animals had 0.7% less IMF than subtropical feedlot contemporaries, the differences were not significant. However, when an atypical Korean heifer slaughter group finished on subtropical pasture (n = 50) was excluded from the analysis, pasture-finished animals had less IMF (P < 0.001) than feedlot animals. Similar trends were apparent for the contrast between subtropical pasture v. temperate feedlot animals, however they did not differ for HOTP8.

|
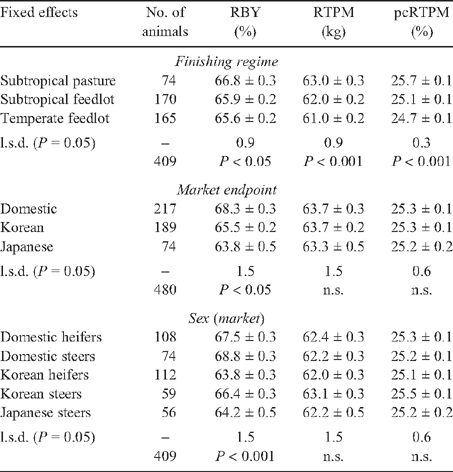
|
Subtropical v. temperate finishing environments
Subtropical feedlot animals had higher (P < 0.001) RTPM and pcRTPM but did not differ in RBY relative to temperate contemporaries (Table 7). Subtropical feedlot animals averaged 1.4 mm more HOTP8 (P < 0.001) than temperate feedlot contemporaries, whereas temperate feedlot animals had 0.54% more IMF (P < 0.001) than subtropical feedlot animals (Table 6). Differences between northern (subtropical) and southern (temperate) feedlots for HOTP8 and IMF are shown in Fig. 1.
An atypical domestic heifer slaughter group finished in the temperate feedlot (n = 53) was responsible for the significant difference in IMF between subtropical and temperate feedlots (2.74 and 3.28% respectively; P < 0.001). Although this slaughter group had a higher mean for IMF than other heifer groups, the measurements were normally distributed and considered genuine.
In an analysis excluding the two atypical heifer slaughter groups, there was no significant difference between subtropical and temperate feedlots for IMF (n = 1085). IMF for subtropical pasture, subtropical feedlot and temperate feedlot were 2.39 ± 0.05, 2.91 ± 0.05 and 3.03 ± 0.06 respectively. Therefore, while results are genuine, the apparent environmental effect on marbling may have been caused by variables confounded within finishing environment (abattoir, slaughter day) and should be treated with caution given the limitations of the data. Subtropical feedlot animals had 1.3 mm more HOTP8 (P < 0.001) than temperate feedlot animals. Exclusion of the atypical slaughter groups from analyses for the other fixed effects caused little change to rankings or significance, with the exception of sex within market effects where the sexes no longer differed for IMF at domestic weights, and Korean heifers had more IMF (P < 0.001) than domestic heifers.
Market endpoint effects
Animals slaughtered for the domestic market had less HOTP8 (P < 0.05) and higher RBY (P < 0.05) than animals slaughtered for Korean and Japanese markets, but differences in IMF, RTPM and pcRTPM between markets were not significant (Tables 6 and 7). When the two atypical slaughter groups (n = 103) were excluded from the analysis, domestic animals had less IMF (P < 0.05) than Korean and Japanese market animals. Korean market animals were fatter (HOTP8) than Japanese market animals due to the absence of heifers in the Japanese market.
Higher IMF and lower RBY were the two main differences between the two heavier market weights. Sex within market results show HOTP8 increased to Korean weights and then plateaued for steers, whereas IMF deposition was not significant between domestic and Korean weights, but increased significantly between Korean and Japanese weights (Tables 6 and 7). As steers aged from Korean to Japanese market weights, IMF increased by 0.54% (P < 0.001) and RBY decreased by 2.2% (P < 0.001), but HOTP8, RTPM and pcRTPM did not change significantly.
Sex effects
Heifers and steers were managed separately following weaning, so direct comparisons are not possible. However, the results show interesting trends (Tables 6 and 7). Heifers took 26 and 46 days longer (P < 0.001) than steers to finish to domestic and Korean market weights respectively. Trends for fat deposition were consistent across sexes; however, heifers had more (P < 0.001) subcutaneous and intramuscular fat than steers at common market weights. Conversely, Korean steers had higher RBY (P < 0.001) than heifers, although there was no difference for RTPM or pcRTPM between the sexes.
Herd of origin effects
Herd of origin had a significant effect on unadjusted CWT (P < 0.001), with animals born at ‘Duckponds’ being heavier at slaughter (281 ± 1 v. 274 ± 1) than those born at Brigalow Research Station. When CWT was adjusted to a common AGE, herd of origin effects remained significant with 7 days difference in AGE (P < 0.001).
Year effects
There were significant differences between years of birth for carcass traits measured over more than one year including AGE, HOTP8 and IMF (P < 0.001). Calves born in 1998 were older at slaughter and leaner than the 1996 and 1997 calf crops, whereas IMF was lowest in 1996. There was no significant effect of year of birth on adjusted or unadjusted CWT.
Interactions
Most significant interactions were caused by scale effects rather than breed re-ranking for carcass traits. Breed × finish interactions were significant for AGE (P < 0.05), CWT (adjusted for AGE; P < 0.001) and IMF (P < 0.05). Fig. 1b shows the breed × finish interaction for IMF. The breed × finish interaction for IMF remained significant after removing the two atypical heifer slaughter groups (n = 103) that distorted IMF results for finish effects. Fig. 1b shows IMF differences between finishing regimes were greatest for British crossbreds, particularly when finished in the temperate feedlot. Brahman and Santa Gertrudis sired progeny marbled slightly better on subtropical pasture relative to subtropical feedlot. Breed × finish interactions were not significant for HOTP8, RBY, RTPM and pcRTPM.
Breed × market interactions were significant for AGE (P < 0.05), CWT (unadjusted; P < 0.001) and IMF (P < 0.001). Angus crosses were superior for IMF when finished to Japanese market weight relative to the other markets and crossbreds. There was little difference in IMF between markets for Continental crosses. Limited breed re-ranking within traits occurred between markets, and where it did occur, it was within breed type. Breed × market interactions were not significant for HOTP8, RBY, RTPM and pcRTPM.
Market × finish interactions were significant (P < 0.001) for AGE, CWT, HOTP8, IMF, RTPM and pcRTPM. There was no market × finish interaction for RBY. Animals slaughtered for the domestic market had significantly less HOTP8 and significantly more IMF when finished in the temperate feedlot relative to the subtropical feedlot, whereas animals slaughtered for Korean and Japanese markets performed similarly for both traits in both feedlot environments.
Discussion
Sire breed effects
Large differences exist between breeds for most bioeconomic traits as a result of different selection goals in different breeds (Gregory et al. 1982; Cundiff et al. 1986). Hence selection of the most suitable breed should be done before selection within that breed (Van Vleck et al. 1992). When considered across all carcass traits examined in this study, Brahman crossbreds had superior carcass quality relative to straightbred Brahmans. Brahman crossbreds performed to expectations, that is, in agreement with sire breed effects on carcass quality reported in reviews by Marshall (1994, 1999) and Cundiff and Gregory (1999). Despite these reviews being mainly based on animals finished in temperate environments, sire breed trends applied to this experiment. These reviews generally agree that B. indicus breeds have moderate lean-to-fat ratios and below average marbling, British breeds have low lean-to-fat ratios and relatively high marbling, and Continental breeds have above average lean-to-fat ratios and average marbling. In this experiment, breeds within breed type (British, Continental) performed similarly. Therefore, cattle breeders should be able to select any breed within breed type examined in this study and achieve predictable outcomes for carcass quality traits.
Carcass attributes of B. taurus and B. taurus × B. indicus cattle grown in temperate environments are well documented. Despite the inability to directly compare production systems and crosses, and inconsistent selection and definitions of traits examined, crossbred carcass traits from this experiment are generally consistent in direction, significance and sire breed ranking to trends from Cycle V (including Brahman sires × B. taurus dams) and Cycle VII (including Continental and British sires × B. taurus dams) of the USDA Meat Animal Research Centre Germplasm Evaluation Program (Cundiff et al. 1998, 2001). In contrast, however, no difference between Angus and Hereford sires were observed for IMF in our experiment. Our results also differed from those of Comerford et al. (1988) who reported significant differences in marbling score (30-point scale, lower = less marbling) between Limousin × Brahman and straightbred Brahmans (12.6 ± 0.7 and 9.4 ± 0.6), and non-significant differences in fat thickness at the 12th rib site between Limousin × Brahman and Hereford × Brahman crosses relative to straightbred Brahmans. In an experiment reported by Huffman et al. (1990) where Angus × Brahman steers (50% Brahman v. 75% Brahman content) were finished on grain to hot carcass weights of 310 and 318 kg, dressing percentage (62.6 v. 63.4%), adjusted fat over the rib eye (1.20 v. 1.24 cm) and USDA yield grade (3.1 v. 3.1) were not different. However, the 75% Brahman content steers had lower marbling scores than 50% Brahman content steers (P < 0.05). These results support our findings for Santa Gertrudis crosses relative to British × Brahman crosses for HOTP8, IMF and RBY.
Breed effects were generally consistent in direction and ranking with other Brahman crossbred studies conducted in subtropical environments in the USA. Peacock et al. (1977) reported intermediate fat depths and carcass yields of Brahman steers (6.3 mm and 50.79%) relative to Angus × Brahman (9.9 mm and 49.99%) and Brahman × Charolais (5.8 mm and 51.34%) steers grown in South Central Florida. DeRouen et al. (1992) reported that relative to straightbred Brahmans, Angus × Brahman, Hereford × Brahman and Charolais × Brahman crosses slaughtered at average carcass weights of 271, 265 and 290 kg across four generations averaged 41, 35 and 60 kg higher hot carcass weights, 18, 16 and 31 kg higher retail yields, 4, 4 and 2 mm greater fat thickness, and 0.8, 0.6 and 0.5 units higher marbling scores when finished on ryegrass pasture and feedlot in subtropical Louisiana. Our results were in agreement in direction and ranking with DeRouen et al. (1992), with the exception of straightbred Brahmans having 1.4 mm greater P8 fat thickness than Charolais × Brahman, and more RBY and RTPM than Angus × Brahman (0.2% and 0.6 kg) and Hereford × Brahman (0.4% and 0.3 kg) crosses.
Pasture v. feedlot finishing
Under subtropical conditions, our experiment showed feedlot finishing has advantages for turning cattle off several months earlier with higher fat cover and marbling, and little sacrifice in RBY, RTPM and pcRTPM, relative to pasture-finished animals. This is consistent with findings by Robinson et al. (2001) who reported B. taurus steers finished on pasture in temperate New South Wales had less HOTP8 and IMF than their grain-finished contemporaries. This has important implications in northern Australia where seasonal pasture availability and quality often results in animals being held over for another dry season in order to reach target market weights. Similarly, DeRouen et al. (1992) recommended grain feeding for a minimum of 90 days after ryegrass grazing in subtropical Louisiana to ensure satisfactory carcass quality (hot carcass weight, retail yield, fat thickness, marbling score and quality grade) in Brahman and Brahman × B. taurus crosses.
Results of this crossbreeding program were similar in direction and significance to results from the linked Beef CRC Straightbreeding Program for HOTP8 (10.46 and 12.98 mm), IMF (2.06 and 2.64%), and RBY (68.10 and 66.47%) for animals finished on subtropical pasture and subtropical feedlot respectively (Reverter et al. 2003). In the Straightbreeding Program, animals finished on pasture and feedlot in temperate environments differed for all traits except HOTP8, which is consistent with results from this study for animals finished on subtropical pasture and temperate feedlot.
Bennett et al. (1995) reported steers with ≤ 50% Brahman content finished on rhizome peanut tropical grass pasture in Florida took longer to finish (197 v. 155 days, P < 0.001), had less fat over the rib eye (8.3 v. 11.4 mm, P < 0.01), lower USDA quality grade (P < 0.01) and lower dressing percentage (P < 0.001) than grain-finished steers in Texas. However, in contrast to our results, they found significantly lower marbling scores for forage-finished steers (P < 0.001) and no difference in yield grade between finishing regimes. The higher retail yields of the pasture-finished Brahman crosses from our experiment align with the results from a two-, three- and four-breed rotational crossbreeding experiment including Angus, Brahman, Charolais and Hereford breeds (DeRouen et al. 1992). However, their Brahman crosses had similar fat thickness and significantly lower marbling scores than contemporaries finished on grain.
As ranking of breeds within breed type remained consistent for each carcass trait across pasture and feedlot finishing in our experiment, changing breeds to target different finishing regimes is not necessary for northern Australian cattle production systems. Reverter et al. (2003) reported little evidence of genotype × environment interactions from the Straightbreeding Program, lending further weight to our conclusion. Furthermore, Hearnshaw et al. (1988) reported Brahman × Hereford animals were consistently superior in growth and carcass traits to straightbreds and backcrosses across three different temperate environments including unimproved and improved pastures and feedlot. The authors concluded that breed rankings were the same irrespective of postweaning environment. In contrast, in a New Zealand study comparing eight carcass quality traits (including hot carcass weight, subcutaneous fat depth and marbling score) of 54 pasture-finished and 148 feedlot-finished Angus steers, Charteris et al. (1997) reported significant (P = 0.05) sire × finishing environment interactions for most carcass traits studied.
Subtropical v. temperate finishing environments
The main differences between finishing environments were significantly higher subcutaneous fat depth of subtropical feedlot animals relative to significantly higher marbling of temperate feedlot animals. Effects of abattoir and slaughter day (not included in the model due to confounding) may have contributed to these differences. Significant differences between subtropical and temperate finishing environments were also reported in the Straightbreeding Program between Brahman, Belmont Red and Santa Gertrudis breeds for HOTP8 (Oddy et al. 1997b; Kelly et al. 2001; Reverter et al. 2003) and scanned P8 fat depth (Johnston et al. 2003). Temperate feedlot-finished crossbreds had 0.54% higher IMF (P < 0.001) and 1.4 mm less HOTP8 than subtropical feedlot-finished crossbreds. Tropically adapted straightbreds from the Straightbreeding Program had consistent results, with temperate feedlot-finished animals having 0.87% more IMF and 3.6 mm less HOTP8 than tropical feedlot straightbreds (Reverter et al. 2003).
Significant differences in HOTP8 and IMF between finishing environments suggest environmental effects on fat distribution in the carcass. However, this hypothesis cannot be tested within the scope of this experiment. Growth path differences could be responsible for differences in fat deposition between environments, including differences in grow-out pasture quality and compensatory gain effects resulting from seasonal pasture quantity and quality fluctuations in the subtropical environment during the grow-out phase. Age at time of nutritional restriction, severity and length of restriction, diet quality following realimentation, and length of realimentation before finishing are all likely to influence body composition, fat partitioning and feed efficiency (Keele et al. 1992; Ball et al. 1997; Oddy et al. 1997a, 1997b). Reverter et al. (2003) suggested higher fat deposition in animals eating high energy diets may have been the cause of the differences in HOTP8 and IMF between environments in the Straightbreeding Program, with fat distribution being favoured in the lumbo-sacral region against the thorax in subtropical environments as part of the thermoregulatory process. Growth path differences between north (subtropical) and south (temperate) were also suggested. Kelly et al. (2001) concluded that grow-out nutrition and geographic location significantly affected fatty acid composition of subcutaneous beef fat, where cattle grown in a tropical environment were older, fatter and had higher percentages of saturated fats relative to animals grown out in a temperate environment. Differences between temperate and tropical locations in ambient temperature, nutrition during the grow-out phase, and growth path were implicated in these results. Robinson et al. (2001) observed that steers that had better grow-out nutrition tended to have more intramuscular fat at Japanese weights, which is consistent with our crossbred results. Whether results from other studies can be extrapolated to explain why animals finished in the subtropical feedlot environment had more subcutaneous fat and less marbling than contemporaries finished in the temperate feedlot environment remains conjecture.
Breed rankings within breed type did not change across finishing environments, with the exception of a breed × finish interaction for IMF. This supports the earlier conclusion that cattle producers in subtropical and grassland environments in northern Australia do not need to change existing breeding programs to target different finishing environments. In the Straightbreeding Program including 320 Brahmans from the Crossbreeding Program, Johnston et al. (2003) found no evidence of genotype × environment interactions for weight and body composition traits from Angus, Hereford, Shorthorn, Murray Grey, Brahman, Belmont Red and Santa Gertrudis purebreds (n = 7655). Johnston et al. (2003) concluded that changing the production system had significant effects on phenotypic expression of growth and body composition traits, but had little effect on the underlying genetic expression and subsequent ranking of sires. Newman et al. (2002) also concluded that crossbreeding could result in some re-ranking of sire performance for weight-related traits, but there was little expected change in carcass traits.
Market endpoint effects
The effect of increasing age on carcass quality attributes is well known. As bodyweight increases towards an animal’s mature weight the proportion of fat increases, the proportion of muscle decreases slightly or remains constant, and the proportion of bone decreases (Ball et al. 1997). The Crossbreeding Program results agreed in direction and significance to those reported by Reverter et al. (2003) for the Beef CRC Straightbreeding Program, where Brahman, Belmont Red and Santa Gertrudis straightbreds had lower RBY and higher fatness (IMF, HOTP8 and 12/13th rib fat thickness) at heavier market endpoints. Ball and Johnson (1989) reported similar results for a Queensland study involving male and female Brahman crossbreds.
Similarly to our results, Comerford et al. (1988) reported consistent differences between sexes for 12th rib fat thickness, marbling score and yield grade (P < 0.01) in a diallel breeding experiment including Limousin, Polled Hereford and Brahman beef cattle finished on grain and slaughtered at 445 days of age. Therefore, in markets where a premium is paid for marbling, results suggest heifers could be slaughtered at a younger age and lighter weight than steers to achieve similar carcass quality and a slight advantage for RBY. Though the commercial reality of putting this into practice may be prohibitive at present due to the primary market focus being on carcass weight rather than carcass quality attributes.
Implications for industry breeding programs
The important outcome from this experiment is that once commercial cattle breeders have settled on a genotype suited to their subtropical production environment, they do not need to change breeds to target different markets, different nutritional regimes (pasture v. grain) or different finishing environments (subtropical v. temperate). Generally, a sire breed that performs well for a particular trait in one market or finishing environment, will perform consistently for that trait relative to other sire breeds in a different market or finishing environment. Furthermore, all genotypes tested in this experiment complied with basic weight and fat specifications for the markets to which they were targeted.
Use of tropically adapted breeds including Belmont Red and Santa Gertrudis is advantageous where terminal crossbreeding programs prevent use of straightbred British or Continental sires because of severe environmental stressors. In adverse environments, establishing a Brahman × B. taurus crossbreed would allow the retention of adaptation and survival traits, coupled with the advantages of improved carcass quality relative to straightbred Brahmans. Breeders could expect similar carcass quality from established crossbreeds or multi-breed composites relative to this terminal crossbreeding experiment, with the exception of a possible increase in AGE because of reduced heterosis effects on carcass weight. Apart from faster growth rates, a major benefit of crossbreeding is the complementary blending of breed characteristics which can overcome genetic antagonisms between some production traits that can be encountered from within-breed selection.
Feedlot finishing hastens growth allowing animals to reach market specifications for carcass weight, fatness and yield traits significantly earlier than pasture finishing. This can be particularly useful in northern Australia where marked and variable wet and dry seasons cause large fluctuations in pasture availability and quality, resulting in lengthy pasture finishing regimes often spanning more than one wet season before animals reach target carcass weights. Temperate finishing environments may have benefits for increased marbling and reduced subcutaneous fat depth over tropical finishing environments.
Acknowledgements
The authors gratefully acknowledge the significant efforts of all Beef CRC staff involved in breeding and managing the experimental animals, field data collection at ‘Duckponds’, ‘Goonoo’, ‘Tullimba’ and ‘McMaster’, abattoir data collection, laboratory meat quality analyses, collation of project data and maintenance of the CRC database. The following donors of Brahman breeding cows for use in the project are also gratefully acknowledged: Hillgrove Pastoral Co., Australian Agricultural Co., North Australian Pastoral Co., Stanbroke Pastoral Co., Queensland and Northern Territory Pastoral Co., Consolidated Pastoral Co., Heytesbury Pastoral Co., Peter Hughes and Acton Land and Cattle Co. Generous funding for the project was provided by Meat and Livestock Australia through Project NAP.3.104.
Ayres JF,
Dicker RW,
McPhee MJ,
Turner AD,
Murison RD, Kamphorst PG
(2001) Post-weaning growth of cattle in northern New South Wales. 1. Grazing value of temperate perennial pasture grazed by cattle. Australian Journal of Experimental Agriculture 41, 959–969.
| Crossref | GoogleScholarGoogle Scholar |
(Emerald Post Office), http://www.bom.gov.au/climate/averages/tables/cw_056006.shtml (Bundarra Post Office) and http://www.bom.gov.au/climate/averages/tables/cw_056013.shtml (Glenn Innes Agricultural Research Station) [Verified 18 February 2006]
Charteris PL,
Garrick DJ, Morris ST
(1997) Sire by finishing environment interactions for beef cattle carcass and meat quality traits. Proceedings of the New Zealand Society of Animal Production 57, 192–195.

Comerford JW,
Benyshek LL,
Bertrand JK, Johnson MH
(1988) Evaluation of performance characteristics in a diallel among Simmental, Limousin, Polled Hereford and Brahman beef cattle. II. Carcass traits. Journal of Animal Science 66, 306–316.
|
CAS |
PubMed |
Corbet NJ,
Miller RG,
Fraser PF,
Lloyd AG, Burrow HM
(1997) Sire breed effects on gestation length of calves in the subtropics. Proceedings of the Association for the Advancement of Animal Breeding and Genetics 12, 300–303.
Corbet NJ,
Miller RG,
Bindon BM,
Burrow HM,
D’Occhio MJ,
Entwistle KW, Fitzpatrick LA
(1999) Synchronization of estrus and fertility in zebu beef heifers treated with three estrus synchronization protocols. Theriogenology 51, 647–659.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
DeRouen SM,
Franke DE,
Bidner TD, Blouin DC
(1992) Two-, three-, and sfour-breed rotational crossbreeding of beef cattle: carcass traits. Journal of Animal Science 70, 3665–3676.
|
CAS |
PubMed |
Duncan DB
(1955) Multiple range test and multiple F test. Biometrics 11, 1–42.
| Crossref | GoogleScholarGoogle Scholar |
Egan AF,
Ferguson DM, Thompson JM
(2001) Consumer sensory requirements for beef and their implications for the Australian beef industry. Australian Journal of Experimental Agriculture 41, 855–859.
| Crossref | GoogleScholarGoogle Scholar |
Hearnshaw H,
Barlow R,
Blackwood I,
Doyle P,
Hoffman W, Wilson C
(1988) Growth and carcass characteristics of Bos indicus and Bos taurus cattle and their crosses in temperate Australia. Proceedings of the Association for the Advancement of Animal Breeding and Genetics 7, 452–455.
Huffman RD,
Williams SE,
Hargrove DD,
Johnson DD, Marshall TT
(1990) Effects of percentage Brahman and Angus breeding, age-season of feeding and slaughter end point on feedlot performance and carcass characteristics. Journal of Animal Science 68, 2243–2252.
|
CAS |
PubMed |
Hunter RA,
Burrow HM, McCrabb GJ
(2001) Sustained growth promotion, carcass and meat quality of steers slaughtered at three liveweights. Australian Journal of Experimental Agriculture 41, 1033–1040.
| Crossref | GoogleScholarGoogle Scholar |
Jeffery M,
James T, Loxton I
(1993) The effect of grain feeding on growth rates and carcase attributes of grazing steers in central Queensland. Recent Advances in Animal Nutrition in Australia 9, 4A.
Johnston DJ,
Reverter A,
Burrow HM,
Oddy VH, Robinson DL
(2003) Genetic and phenotypic characterisation of animal, carcass, and meat quality traits from temperate and tropically adapted beef breeds. 1. Animal measures. Australian Journal of Agricultural Research 54, 107–118.
| Crossref | GoogleScholarGoogle Scholar |
Keele JW,
Williams CB, Bennett GL
(1992) A computer model to predict the effects of level of nutrition on composition of empty body gain in beef cattle. I. Theory and development. Journal of Animal Science 70, 841–857.
|
CAS |
PubMed |
Kelly MJ,
Tume RK,
Newman SA, Thompson JM
(2001) Environmental effects on the fatty acid composition of subcutaneous beef fat. Australian Journal of Experimental Agriculture 41, 1023–1031.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
Kramer CY
(1957) Extension of multiple range tests to group correlated adjusted means. Biometrics 13, 13–18.
| Crossref | GoogleScholarGoogle Scholar |
Marshall DM
(1994) Breed differences and genetic parameters for body composition traits in beef cattle. Journal of Animal Science 72, 2745–2755.
|
CAS |
PubMed |
Newman S,
Burrow HM,
Sim W,
Williams P,
Kelly M,
Menzies D, Farrell E
(1998) Growth and carcass traits of F1 steers and heifers fed for three markets at two locations in Australia. Journal of Animal Science 76(Suppl. 1), 77.
Newman S,
Burrow HM,
Shepherd RK, Bindon BM
(1999) Carcass yield traits of grass- and grain-finished Brahman crosses for domestic and export markets. Proceedings of the Association for the Advancement of Animal Breeding and Genetics 13, 231–234.
Newman S,
Reverter A, Johnston DJ
(2002) Purebred-crossbred performance and genetic evaluation of postweaning growth and carcass traits in Bos indicus × Bos taurus crosses in Australia. Journal of Animal Science 80, 1801–1808.
|
CAS |
PubMed |
Oddy VH,
Ball AJ, Pleasants AB
(1997a) Understanding body composition and efficiency in ruminants: a non-linear approach. Recent Advances in Animal Nutrition in Australia 11, 209–222.
Oddy VH,
Herd RM,
Ball AJ,
Dicker RW,
Robinson DL,
Skerritt J,
Hammond AJ, Thompson JM
(1997b) Prior performance of cattle influences current and future performance, meat yield and quality characteristics: Implications for genetic evaluation. Proceedings of the Association for the Advancement of Animal Breeding and Genetics 12, 738–741.
Perry D,
Shorthose WR,
Ferguson DM, Thompson JM
(2001) Methods used in the CRC program for the determination of carcass yield and beef quality. Australian Journal of Experimental Agriculture 41, 953–957.
| Crossref | GoogleScholarGoogle Scholar |
Reverter A,
Johnston DJ,
Perry D,
Goddard ME, Burrow HM
(2003) Genetic and phenotypic characterisation of animal, carcass, and meat quality traits from temperate and tropically adapted beef breeds. 2. Abattoir carcass traits. Australian Journal of Agricultural Research 54, 119–134.
| Crossref | GoogleScholarGoogle Scholar |
Robinson DL
(1995) Design of the CRC straightbred genetics experiments. Proceedings of the Association for the Advancement of Animal Breeding and Genetics 11, 541–545.
Robinson DL,
Oddy VH,
Dicker RW, McPhee MJ
(2001) Post-weaning growth of cattle in northern New South Wales. 3. Carry-over effects on finishing, carcass characteristics and intramuscular fat. Australian Journal of Experimental Agriculture 41, 1041–1049.
| Crossref | GoogleScholarGoogle Scholar |
Schutt KM,
Arthur PF, Burrow HM
(2009a) Brahman and Brahman crossbred cattle grown on pasture and in feedlots in subtropical and temperate Australia. 3. Feed efficiency and feeding behaviour of feedlot-finished animals. Animal Production Science 49, 452–460.
| Crossref | GoogleScholarGoogle Scholar |
Schutt KM,
Burrow HM,
Thompson JM, Bindon BM
(2009b) Brahman and Brahman crossbred cattle grown on pasture and in feedlots in subtropical and temperate Australia. 2. Meat quality and palatability. Animal Production Science 49, 439–451.
| Crossref | GoogleScholarGoogle Scholar |
Upton W,
Burrow HM,
Dundon A,
Robinson DL, Farrell EB
(2001) CRC breeding program design, measurements and database: methods that underpin CRC research results. Australian Journal of Experimental Agriculture 41, 943–952.
| Crossref | GoogleScholarGoogle Scholar |
Van Vleck LD,
Hakim AF,
Cundiff LV,
Koch RM,
Crouse JD, Boldman KG
(1992) Estimated breeding values for meat characteristics of crossbred cattle with an animal model. Journal of Animal Science 70, 363–371.
|
CAS |
PubMed |