

The liveweight, carcasses and meat quality of intact and castrated male lambs reared under a pasture-fed Australian production system
Shawn R. McGrath A B , M. Bruce Allworth A B , James Stephens C , Michelle L. E. Henry D and Benjamin W. B. Holman E *
A B , M. Bruce Allworth A B , James Stephens C , Michelle L. E. Henry D and Benjamin W. B. Holman E *
A
B
C
D
E
Abstract
Intact male (ram) lambs are often discounted by meat processors because of their perceived defective meat quality.
This study quantified the effects of castration on the liveweight, growth, carcass parameters and the meat quality of male lambs.
Liveweight, growth and carcass data were collected from 133 ram and 132 castrated (wether) lambs reared under a pasture-fed production system. A total of 50 lambs were subsequently selected, balanced by final curfew liveweight and castration status, to investigate additional carcass, meat quality, consumer sensory panel scores and colour stability parameters.
It was found that total average daily gains for ram lambs were highest (242.9 ± 5.2 g/day; P = 0.002), resulting from their higher pre-weaning growth rate of 327.7 ± 9.7 g/day (P < 0.001). Wethers were fattier, having higher estimated intramuscular fat (3.9 vs 3.3 ± 0.1%, respectively) and subcutaneous fat depth (14.7 vs 13.5 ± 0.4 mm, respectively) than ram lambs. Purge loss (P < 0.030) and ultimate pH (P < 0.035) were higher for the meat of wether lambs, but only after 32 days of wet ageing. Consumers scored the meat of wether lambs as significantly more tender (61.0 vs 54.6 ± 2.3%; P = 0.005) and of greater overall liking (65.1 vs 61.1 ± 2.0%; P = 0.044) than that of ram lambs. These, and a comparison of shear force, near infrared spectroscopy for intramuscular fat content and retail colour with consumer thresholds, suggest a general satisfaction with ram and wether lamb meat quality.
Collectively, there seems to be few tangible performance and meat quality advantages to the castration of male lambs.
Prime lamb producers must consider flock management and market preference when deciding on best practice.
Keywords: animal performance, carcass properties, consumer sensory panel, intact ram lambs, lean tissue yield, meat shelf-life, wet ageing period, wethers.
Introduction
Ram (intact male) lambs are often discounted by meat processors because of their perceived defective meat quality and additional processing requirements, when compared with wether (castrated male) lambs. Past research supports this industry practice. For example, Purchas et al. (1979) reported that the meat of wether lambs had greater overall acceptability and lower shear force compared with the meat of ram lambs. Ram lambs, reared under free-range production systems, produced meat that was less tender and with more intense flavour than was found for wether lambs (Hoffman et al. 2020). Furthermore, Crouse et al. (1981) observed that the carcasses from ram lambs were leaner and their meat was less desirable to consumers, when compared with wether lambs. Collectively, disparate meat quality appears to be the major barrier to rearing ram lambs – especially when considering that ram lambs are reported to outperform wether lambs (Seideman et al. 1982; Gravador et al. 2018) and avoid a key animal welfare issue (castration) that may detract from the social licence given to prime lamb producers.
Some lamb producing countries have already made it illegal to castrate ram lambs, citing animal welfare as central to their decision (Bhatti et al. 2019). Other countries may soon follow suit. The ‘Australian Animal Welfare Standards and Guidelines for Sheep’ states the need to undertake any husbandry procedures that may result in significant short-term pain must be assessed against alternative strategies for the long-term welfare of sheep (Animal Health Australia 2016). One solution could be the sale of ram lambs before they reach puberty, and before the onset of maturity-associated meat and behavioural changes, the latter necessitates single-sex flocks and complicates the management of ram lambs (Seideman et al. 1982). This solution is buoyed by advancements made to the growth rates of modern prime lamb breeds, which allows them to reach marketable weights and be slaughtered at a younger age (Gardner et al. 2015; Kelman et al. 2022). Gkarane et al. (2017) found that meat from younger ram lambs had a slightly reduced flavour profile than that of wether lambs, a difference that was intensified as animal age increased. Young et al. (2006) found that the tenderness scores for meat of wether and ram lambs diverged only when they exceeded 300 days of age, although before this time point, ram lamb meat tended to have a less acceptable flavour profile compared with the meat of wether lambs. It is therefore clear that additional research is necessary to encourage industry adoption. Specifically, we must understand if leaving ram lambs intact is a viable option, in Australia, if consumer demands or legislative requirements were to be changed.
The objective of this study was to determine the effects of castration on the liveweight, carcass parameters, and the sensory and physiochemical qualities of meat from male lambs.
Materials and methods
Study design and animal management
The procedures and protocols used for this study were approved by the Animal Care and Ethics Committee of Charles Sturt University (A22039), and adhered to the ‘Best Practice Methodology in the use of Animals for Scientific Purposes’ (National Health and Medical Research Council 2013). The experimental lambs were commercial composite breed animals (Meat Plus Genetics, www.meatplus.com.au) that were sourced, reared and managed on the Charles Sturt University Farm (Wagga Wagga, NSW, Australia). Composite sheep are a multibreed sheep used for prime lamb production. They produce a low-quality fleece that requires shearing, and ewe lambs can be retained as replacement breeders (Bates et al. 2023). Specifically, 300 male lambs from composite ewes, which had been scanned as single bearing, were randomly selected for use in this study. The male lambs were born during July 2021 and marked on 6 August 2021. Lambs were systematically allocated to one of the two treatment groups, with every other male lamb allocated to be left uncastrated (n = 150) or castrated (n = 150) using the Numnuts® system (Small et al. 2020). Lambs were individually identifiable from an electronic ear tag that was attached at lamb marking.
Ewes and lambs were grazed on a dual-purpose cereal crop (Triticum aestivum cv. EGA Wedgetail) without access to roughage or mineral supplements, and as a single flock from lamb marking until weaning on 28 September 2021. Lambs were vaccinated against clostridial, cheesy gland and arthritis disease (7 in 1, GlanEry® Australia) at marking and weaning. Female lambs were removed from the flock at weaning and, thereafter, the experimental lambs (ram and wether) were run as a single flock. Several female lambs were later detected in the flock and removed. Following weaning, male lambs were grazed on pastures dominated by lucerne (Medicago sativa) and barley grass (Hordeum leporinum), and after the barley grass became reproductive, lambs were grazed on a lucerne monoculture. No roughage or mineral supplements were provided to the experimental lambs.
Liveweight and growth
Experimental lamb liveweights were recorded, first and immediately after marking (marking weight), using a lambing box, which weighed lambs as they exited the lamb marking cradle, and again at weaning (weaning weight) using portable, electronic sheep scales fitted with an alloy weigh crate. Both these weights were without curfew, as lambs were yarded with their dams. Curfew refers to a period wherein lambs were held off-feed, with ad libitum access to drinking water only. Two lambs were reported to have died post-weaning, with a ram lamb with arthritis in the hindquarters euthanised and another lamb found dead after becoming flystruck. No management problems were identified by the farm manager during the experiment; however, the importance of correct drafting and the prevalence of mounting behaviour post-weaning was noted. Lambs were weighed after an overnight curfew on 9 November 2021 to estimate the number of lambs that would reach the target liveweight of ≥38.0 kg (liveweight + ADG × days until slaughter). A total of 133 ram and 132 wether lambs achieved this target weight, and were therefore designated for slaughter (n = 265). Final curfew liveweights were recorded on 8 December 2021, after a period off-feed. Pre-weaning (preADG, 51 days), post-weaning (postADG, 72 days) and total average daily weight gain (ADG; 123 days) were calculated by dividing the difference in liveweights by the number of days lapsed.
Slaughter and carcass evaluation
The 265 experimental lambs were slaughtered on 16 December 2021 (5–6 months of age) as a single flock at a commercial Australian abattoir, following an overnight lairage period. Carcasses were eviscerated, dressed and inspected in accordance with standard industry practices. Hot standard carcass weight (HSCW) was recorded at ~30 min post-mortem and before assessment with a commercial dual energy X-ray absorptiometer (Scott Automation and Robotics, Australia). This provided an estimation of individual carcass composition, in terms of percentage lean muscle, fat and bone tissue (Connaughton et al. 2020). Intramuscular fat content was estimated (estimated-IMF) using a MEQ Probe (MEQ Solutions, Australia). The curfewed live animal weight and HSCW were used to calculate the dressing percentage. Medium voltage electrical stimulation was applied to each carcass before they entered the holding chiller (Pearce et al. 2010).
A cohort of 50 lambs was selected from the 265 experimental lambs, prior to slaughter and transportation to the abattoir. Half of these were ram lambs, and the other half were wether lambs, selected so both groups (n = 25/group) were balanced by final liveweight. For these cohort carcasses only, the tissue depth on the girth rib site, being the lateral surface of the 12th rib and 110 mm from the midline, was measured using a specialised knife (mm). At 24 h post-mortem, the cohort carcasses were fabricated (boned out) and the loin saddles collected. The exposed surface, at the 12th rib, of the left longissimus lumborum (LL) was measured with a metal ruler, and the subcutaneous fat depth (mm) and eye muscle area (cm2) were calculated (Hopkins et al. 1992a). Fresh colour measurements were made using a chromameter (CR-400, Minolta, Japan) with an 8-mm aperture, and using Illuminant D65 and 10° standard observed settings. Triplicate readings were made for each sample, with care made to avoid fatty deposits and connective tissue when (re)positioning the chromameter. Mean L*, a* and b* values were reported and used to calculate hue and chroma (AMSA 2012). Final pH (pH at 24 h post-mortem) was measured at this same site, using a spear-type gel electrode and calibrated pH meter (IJ-44 and WP-80 respectively, TPS, Australia). The pH meter was calibrated using pH 4.0 and pH 6.8 U standard buffers at muscle approximate temperatures. Both LL were removed, denuded of silverskin and then dissected for wet ageing or analysis.
Sample dissection and storage
Each left LL was halved, and these were assigned to each of two wet ageing periods (5 or 32 days), with allocation balanced by portion (caudal or cranial ends). Portions were vacuum packaged and aged at 2–3°C. Once aged, samples prepared for the analysis of cooking loss, shear force and ultimate pH were frozen at −25°C. Samples were also prepared for the analysis of purge loss and drip loss, albeit these were assessed using fresh (unfrozen) samples.
From each right LL, the cranial portion was immediately removed, frozen at −80°C, and designated for the analysis of total moisture and intramuscular fat content (IMF). The remainder of the muscle was vacuum packaged and wet aged for 5 days under the same aforementioned conditions. Sample slices for the analysis of colour stability were removed from the medial portion of the muscle and assessed fresh (unfrozen). Sample slices for sensory evaluation were prepared from the remainder of the muscle, and these were frozen at −25°C until analysis.
Meat quality
The percentage change is sample weight after wet ageing was calculated as purge loss (%). Samples were weighed without vacuum packaging, and before measuring the final weight, samples were first patted dry with paper towel.
A core of 2.5 cm in diameter was prepared from each sample and placed into individual EZ-DripLoss tubes (Danish Meat Research Institute, Denmark). These were held refrigerated for 72 h, and the percentage change in sample core weight was used to calculate drip loss (%) (Holman et al. 2020a).
Sample blocks of 80.7 ± 12.1 g (mean ± s.d.) were cooked, from frozen and in vacuo, for 35 min and using a water bath set to 71°C. Samples were cooked in five batches (preparation batches), with allocation to these randomised albeit balanced lamb status and wet ageing period. Cooked samples were immediately submerged and held in an ice slurry for 30 min, to halt the cooking process (Hopkins and Thompson 2001). The packaging was then removed, and samples patted dry with paper towel. The change in sample weight before and after cooking was reported as cooking loss (%). Cooked samples were placed into resealable plastic bags and refrigerated overnight to standardise their temperature to 3–4°C.
Cuboidal strips with a cross-sectional area of 1 cm2 were prepared and tested using a texture analyser (model LRX; Lloyd Instruments, UK) that was fitted with a Warner Bratzler ‘vee-shaped’ blade, set with a crosshead speed of 20 cm/min (Holman et al. 2015a). The peak force needed to cut each sample strip, when the cutting line was positioned perpendicular to the myofibril orientation, was recorded. The average of six technical replicates was reported as the shear force (N).
Frozen samples of 1 g were homogenised with 6 mL of buffer (5 mM iodoacetate, 150 mM KCl, pH 7.0 at 4°C), and incubated in a 20°C water bath until temperatures equilibrated (Dransfield 1994). The ultimate pH (PHU; U) was then measured using a pH meter that was fitted with a spear-type electrode (Ionode IJ-44 and smartCHEM-CP; TPS, Australia), and calibrated using 4.0- and 6.8-U standard buffers.
Five slices of <1 mm were removed from each sample parallel to the orientation of myofibrils, and these were analysed using laser diffraction (Bouton et al. 1973). The average was reported as the sarcomere length (μm).
Samples of ~25 g placed were into a freeze-drying unit (ScanVac CoolSafe; LaboGene ApS., Denmark) that was set to −50°C. These were removed once their weight had stabilised (±0.02 mg, when measured after a 24-h interval), and the change in weight was used to calculate the total moisture (%). Freeze-dried samples were ground using a sample mill (model 1095, Knifetec; FOSS Pacific Ltd. Australia), and frozen at −80°C for further analyses. The intramuscular fat (NIR-IMF) of each freeze-dried and ground sample was determined using the near infrared (NIR) method of Bailes et al. (2022) and a multipurpose NIR analyser (MPA II Bruker, USA).
Colour stability
Samples of 3-cm depth were prepared and placed onto individual black foam trays, these were positioned so that myofibrils had a perpendicular orientation to the measured surface. Each sample was overwrapped with PVC film, permitted to bloom for 45 min (under refrigeration) and then measured using a spectrophotometer (45/0-L, HunterLab Associates Laboratory, PRC). The spectrophotometer had a 25-mm aperture, and used Illuminant D-65 and 10° standard observer settings (Holman et al. 2015b). After this initial measurement (Day 0), three additional measurements were made, at daily intervals (Day 1–3), between which the samples were held under refrigeration and continuous lighting (NEC 58 W tubes that delivered ~1000 lux to the sample surface). Colour parameters (L*, a* and b*) were recorded and used to calculate hue and chroma values (CIE 1977; AMSA 2012). Reflectance data were also recorded and used to calculate the ratio of reflectance at 630 and 580 nm (R630/580).
Consumer sensory panel
The protocol of Thompson et al. (2005a) was used to analyse the sensory quality of the samples. Specifically, five slices of 1-cm depth were removed from each right-side LL. These were to be halved upon cooking, so that each sample could be tasted by 10 different consumers. Each consumer tasted eight samples, and this equated to 64 consumers volunteering to participate in three sensory panels (1 of 20 and 2 of 22). Informed consent was obtained from all participants involved in the study, with the protocol approved by the Human Research Ethics Committee of Charles Sturt University (H22006). Demographic information was recorded for each consumer (Table 1). Sample allocation was designed so that lamb status was represented (balanced) in each sensory panel and for each consumer. The order in which samples were tasted by a consumer was randomised. Although no experimental sample was lost or omitted, it was necessary to include one non-experimental sample of lamb LL for two consumers (two tastings in total), so as to preserve the aforementioned design elements.
| Demographic categories | % | |
|---|---|---|
| Age (years) | ||
| 18–25 | 67 | |
| 26–30 | 8 | |
| 31–39 | 3 | |
| 40–55 | 9 | |
| ≥55 | 13 | |
| Sex | ||
| Female | 69 | |
| Male | 31 | |
| Smoker | ||
| No | 89 | |
| Yes | 5 | |
| Occupation | ||
| Farmer | 2 | |
| Professional | 17 | |
| Sales and personal service | 2 | |
| Student | 69 | |
| Technical | 8 | |
| Tradesperson | 2 | |
| Red meat consumption | ||
| Daily | 12 | |
| 4–5 times per week | 27 | |
| 2–3 times per week | 38 | |
| Weekly | 16 | |
| Fortnightly | 3 | |
| Monthly | 2 | |
| Less often | 2 | |
| Attitude to red meat | ||
| I enjoy red meat. It is an important part of my diet | 48 | |
| I like red meat. It is a regular part of my diet | 33 | |
| I do eat some red meat although it would not worry me if I didn’t | 19 | |
| I rarely/never eat red meat | – | |
| Preferred level of doneness | ||
| Rare | 27 | |
| Medium | 59 | |
| Well done | 14 | |
| Annual income (AUD) | ||
| <$45,000 | 27 | |
| $45,000–80,000 | 27 | |
| >$80,000 | 45 | |
Some of the 64 participants did not wish to answer specific questions; therefore, the sum percentages do not always equal 100.
Samples were thawed under refrigeration for 24 h prior to the sensory panel. These were cooked to an internal endpoint temperature of 71°C using a clam shell grill (GR-4A; Cuisinart Griddler, USA) set to 220 ± 10°C. Temperatures were verified using an infrared HACCP thermometer (model 8838; AZ Instrument Corp.). Cooked sample slices were immediately halved and served warm to the assigned consumer. Consumers scored each sample on unstructured 10-cm line scales that were bookended with ‘not tender’ versus ‘extremely tender’; ‘not juicy’ versus ‘extremely juicy’; ‘dislike flavour extremely’ versus ‘like flavour extremely’; and ‘dislike extremely overall’ versus ‘like extremely overall’. Categorical ranks of sample quality were also recorded by each consumer, scoring samples as: (1) Awful; (2) Unsatisfactory; (3) Good everyday quality; (4) Better than everyday quality; or (5) Premium quality. Consumers were instructed to cleanse their palate between each tasting with dry water crackers and water.
Statistical analysis
Data were analysed using Genstat (23rd Edition, VSN International, www.vsni.co.uk). Liveweight, preADG, postADG, ADG and carcass data were analysed using linear mixed models fitted with the fixed effects of castration and the random effects of animal. Carcass data included HSCW; percentage bone, fat and lean tissue; dressing; estimated-IMF; and fat depth; and only for the selected 50 carcasses, eye muscle area, subcutaneous fat depth, fresh colour, girth rib tissue depth and pH at 24 h post-mortem. Meat quality data were analysed using linear mixed models fitted with the fixed effects of castration status, ageing period and their interaction, as well as the random effects of animal. Meat quality data included cooking loss, drip loss, NIR-IMF, purge loss, sarcomere length, shear force, total moisture and PHU. Ageing period was not a factor tested for the IMF, sarcomere length and total moisture variables, and was omitted from the model. The pre-cook sample weight was included as a covariate for the analysis of cooking loss and shear force data. Sensory scores for flavour, juiciness, tenderness, overall liking and quality rank were analysed using linear mixed models fitted with the fixed effects of castration, and the random effects of sensory panel, consumer, animal, sample slice, tasting order and their interactions. The level of significance was set at 5%, and non-significant terms were dropped from the final models. Least significant difference (l.s.d.) was used to find differences between predicted means.
Results
Liveweight and rate of growth
The weaning and final curfew liveweights of ram lambs were higher than observed for wether lambs (P < 0.05; Table 2). Ram lamb preADG (327.7 ± 9.7 g/day) and ADG (242.9 ± 5.2 g/day) were highest (P < 0.05). There were no significant castration status effects on marking liveweight or postADG (Table 2).
| Liveweight and growth parameters | Castration status | s.e.m. | P-value | ||
|---|---|---|---|---|---|
| Ram | Wether | ||||
| Marking liveweight (kg) | 14.4 | 14.4 | 0.5 | 0.919 | |
| preADG A (g/day) | 327.7a | 289.9b | 9.7 | <0.001 | |
| Weaning liveweight (kg) | 31.1a | 29.1b | 0.8 | 0.014 | |
| postADG B (g/day) | 188.5 | 187.5 | 7.1 | 0.776 | |
| Final curfew liveweight (kg) | 44.6a | 42.5b | 0.9 | 0.018 | |
| ADG C (g/day) | 242.9a | 226.3b | 5.2 | 0.002 | |
Means, standard error (s.e.m.), and level of significance (P-values) are included.
Means within rows with different lowercase letters were significantly different (P < 0.05).
Carcass parameters and value
For the experimental lambs, the NIR-IMF (P < 0.001) and fat depth (P = 0.007) were found to be higher for wether lambs than for ram lambs (Table 3). There were no castration effects observed on carcass composition, dressing percentage or HSCW of male lambs (P > 0.05). Some wether lambs (9%) received a price bonus for having an estimated IMF > 5%, whereas no ram lambs qualified for this bonus. Omitting the AUD0.50 per kg HSCW discount industry applied to the carcasses of ram lambs, there was no significant effect of castration status on carcass value (P > 0.05). The average value of carcasses from ram and wether lambs was found to be A$196.20 and A$192.90, respectively.
| Carcass parameters | Castration status | s.e.m. | P-value | ||
|---|---|---|---|---|---|
| Ram | Wether | ||||
| Composition | |||||
| Bone tissue (%) | 14.6 | 14.5 | 0.2 | 0.360 | |
| Fat tissue (%) | 28.7 | 29.3 | 0.5 | 0.159 | |
| Lean tissue (%) | 56.7 | 56.2 | 0.3 | 0.125 | |
| Dressing (%) | 52.6 | 52.6 | 0.4 | 0.867 | |
| Estimated IMF (%) | 3.3b | 3.9a | 0.1 | <0.001 | |
| Fat depth (mm) | 13.5b | 14.7a | 0.4 | 0.007 | |
| Hot standard carcass weight (kg) | 24.6 | 24.2 | 0.5 | 0.360 | |
Means, standard error (s.e.m.) and level of significance (P-values) are included.
Means within rows with different lowercase letters were significantly different (P < 0.05).
For the cohort lambs (Table 4), wether carcasses were found to have significantly higher fat tissue (30.5 ± 0.9% vs 27.8 ± 0.9%, respectively) and lower lean tissue (56.8 ± 0.6% vs 55.4 ± 0.6%, respectively) percentages than the carcasses of ram lambs. Fresh colour measures of yellowness (b*) and intensity (hue) were highest for the meat of wether lambs (P < 0.05). The pH at 24 h post-mortem was found to be higher for the ram lambs, when compared with wether lambs (5.52 ± 0.04 vs 5.37 ± 0.04, respectively; P < 0.001).
| Carcass parameters | Castration status | s.e.m. | P-value | ||
|---|---|---|---|---|---|
| Ram | Wether | ||||
| Curfew liveweight (kg) | 47.8 | 47.6 | 1.6 | 0.910 | |
| Composition | |||||
| Bone tissue (%) | 14.5 | 14.2 | 0.2 | 0.282 | |
| Fat tissue (%) | 28.8b | 30.5a | 0.9 | 0.048 | |
| Lean tissue (%) | 56.8a | 55.4b | 0.6 | 0.024 | |
| Dressing (%) | 53.3 | 53.4 | 0.8 | 0.973 | |
| Eye muscle area (cm2) | 14.8 | 13.5 | 0.7 | 0.101 | |
| Fat depth (mm) | 13.6 | 14.3 | 0.8 | 0.422 | |
| Subcutaneous fat depth (mm) | 3.1 | 3.8 | 0.5 | 0.170 | |
| Fresh colour | |||||
| L* | 38.3 | 38.9 | 0.5 | 0.308 | |
| a* | 17.5 | 17.2 | 0.4 | 0.520 | |
| b* | 8.1b | 8.9a | 0.4 | 0.047 | |
| Chroma | 42.1 | 42.5 | 0.5 | 0.393 | |
| Hue | 24.7b | 27.2a | 0.9 | 0.009 | |
| Grth rib tissue depth (mm) | 21.7 | 21.6 | 2.2 | 0.957 | |
| Hot standard carcass weight (kg) | 25.5 | 25.4 | 0.9 | 0.945 | |
| pH at 24 h | 5.52a | 5.37b | 0.04 | <0.001 | |
Means, standard error (s.e.m.) and level of significance (P-values) are included.
Means within rows with different lowercase letters were significantly different (P < 0.05).
Meat quality
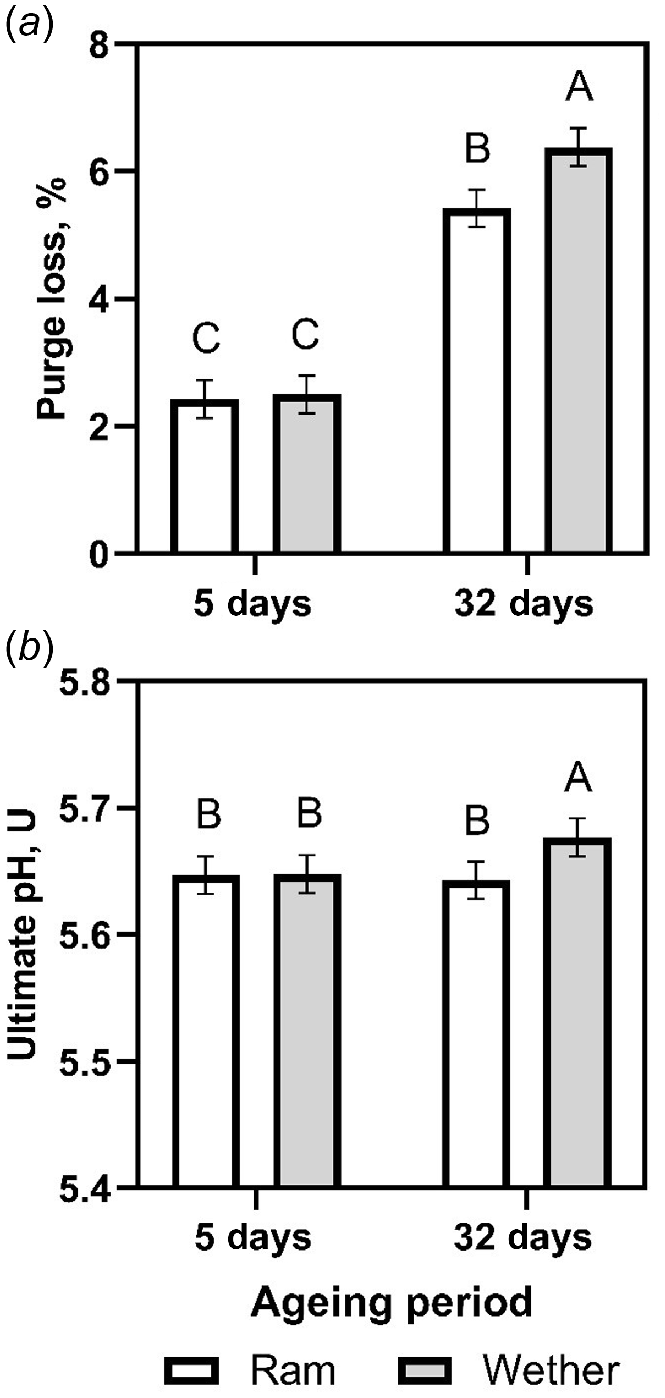
Purge loss was higher for the 32-day wet aged meat of wether lambs, when compared with that of ram lambs (Fig. 1a; P = 0.030). Ultimate pH was highest for 32-day wet aged meat of wether lambs (Fig. 1b; P = 0.035). Purge loss was highest for the meat of wether lambs, independent to the wet ageing period (P = 0.031). The shear force (25.1 ± 0.8 N vs 28.8 ± 0.8 N, respectively) and purge loss (2.5 ± 0.2% vs 5.9 ± 0.2%, respectively) of 32-day wet aged lamb meat were significantly higher than observed after a 5-day wet ageing period (Table 5). Drip loss was highest for 5-day aged lamb meat (P < 0.001).
The effect of castration status (ram vs wether) and ageing period (5 or 32 day) on the (a) purge loss and (b) ultimate pH values of male lamb longissimus lumborum muscles. Predicted means and s.e. (bars) are plotted. Columns with different letters were significantly different (P < 0.05).
| Meat quality parameters | Castration status | s.e.m. | Ageing period | s.e.m. | P-value | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Ram | Wether | 5 days | 32 days | Status | Ageing | Interaction | ||||
| Cooking loss (%) | 23.6 | 22.9 | 0.4 | 23.4 | 23.0 | 0.4 | 0.602 | 0.240 | 0.198 | |
| Drip loss (%) | 0.7 | 0.7 | 0.1 | 0.9z | 0.5y | 0.1 | 0.680 | 0.001 | 0.084 | |
| NIR-MF (%) | 2.8 | 3.2 | 0.2 | – | – | – | 0.093 | – | – | |
| Purge loss (%) | 3.9b | 4.4a | 0.2 | 2.5y | 5.9z | 0.2 | 0.031 | <0.001 | 0.030 | |
| Sarcomere length (μm) | 1.7 | 1.8 | < 0.1 | – | – | – | 0.051 | – | – | |
| Shear force (N) | 27.9 | 26.0 | 1.2 | 28.8z | 25.1y | 0.8 | 0.116 | <0.001 | 0.123 | |
| Total moisture (%) | 74.7 | 74.5 | 0.2 | – | – | – | 0.342 | – | – | |
| Ultimate pH | 5.65 | 5.66 | 0.02 | 5.65 | 5.66 | 0.01 | 0.231 | 0.113 | 0.035 | |
Means, standard error (s.e.m.) and level of significance (P-values) are included.
Means within fixed effects and within rows with different lowercase letters were significantly different (P < 0.05).
Colour stability
There were no significant effects of castration status by display period interactions on colour parameters (P > 0.05). L*, a*, b* and chroma were found to increase between Day 0 and 1 of display, and thereafter decline (P < 0.001; Table 6). Hue increased with each increase to the display period (P < 0.001), and R630/580 decreased with each increase to the display period (P < 0.001).
| Colour parameters | Display period (day) | s.e.m. | P-value | ||||
|---|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | ||||
| L* | 38.0c | 40.4a | 39.1b | 39.0b | 0.2 | <0.001 | |
| a* | 15.1b | 16.6a | 15.3b | 13.7c | 0.2 | <0.001 | |
| b* | 13.0c | 16.9a | 16.1a | 15.3b | 0.1 | <0.001 | |
| hue | 40.6d | 45.6c | 46.6b | 48.2a | 0.3 | <0.001 | |
| Chroma | 20.0c | 23.7a | 22.2b | 20.5c | 0.2 | <0.001 | |
| R630/580 A | 4.4a | 3.5b | 2.8c | 2.5d | 0.1 | <0.001 | |
Consumer sensory panel
The meat of wethers was scored as more tender (P = 0.005) and of greater overall liking (P = 0.044) than the meat of ram lambs (Table 7). The consumer scores for flavour, juiciness, tenderness and overall liking were >50% for meat of ram and wether lambs. All of the lamb meat was considered to be of ‘good everyday quality’ (Table 7).
| Consumer sensory panel scores | Castration status | s.e.m. | P-value | ||
|---|---|---|---|---|---|
| Ram | Wether | ||||
| Flavour (%) | 63.7 | 66.3 | 1.8 | 0.144 | |
| Juiciness (%) | 56.6 | 57.3 | 2.7 | 0.797 | |
| Tenderness (%) | 54.6b | 61.0a | 2.3 | 0.005 | |
| Overall liking (%) | 61.1b | 65.1a | 2.0 | 0.044 | |
| Quality rank A, 1–5 | 3.3 | 3.3 | 0.1 | 0.369 | |
Means, standard error (s.e.m.) and level of significance (P-values) are included.
Means within rows with different lowercase letters were significantly different (P < 0.05).
Discussion
The ram lambs were found to grow faster than wether lambs, and thereby had a heavier final curfew liveweight, apparent from the absence of any significant difference in marking (initial) liveweights. These findings confirm those presented in the literature (Seideman et al. 1982; Lee et al. 1990; Mahgoub and Lodge 1994; Gravador et al. 2018). Similar to the current study, Purchas (1978) found that preADG were highest for ram lambs, and that postADG were comparable between ram and wether lambs. These outcomes are likely because of the substantive reduction to plasma testosterone and androstenedione concentrations that occurs when male lambs are castrated (Wellington et al. 2003). These androgens are important for protein production and to support animal growth (Schanbacher et al. 1980; Hopkins et al. 1992b; Arnold et al. 1997). It is noted that wether lambs still have androgen levels sufficient to support these outcomes, albeit at a lesser rate – as observed in the current study.
It was unexpected to find differences in liveweight only at pre-weaning, although this has been reported elsewhere (Purchas 1978). PostADG for both ram and wether lambs in this study were only moderate, and it may be that other factors, such as nutrition, meant that the rams were unable to express their potential for higher growth rates. Champness et al. (2021) reported the opportunity to improve growth rates of lambs grazing lucerne by supplementation with coarse salt – research completed on the same farm as the current study (University Farm, Wagga Wagga, NSW, Australia). A greater difference in production through higher postADG and subsequent increase in sale value would be one lever that could encourage uptake of ram lamb production. Lower IMF levels observed for ram lambs, in comparison with wether lambs, resulted in the carcass values being similar for ram and wether lambs. This means that ram lambs would require heavier carcass weights to overcome this pricing factor. Of greater influence, however, would be the discounts currently applied at Australian abattoirs for ram lambs, which actively discourage ram lamb production for the majority of commercial producers. Understanding why such discounts are applied, how production systems could be changed or lamb age assurance programs, so that discounts are not necessary, would be required to enable on-farm and practical change.
Across the experimental lamb carcasses, estimated IMF and subcutaneous fat deposits were found to be higher for wether lambs. These could be the result of growth rate and final curfew liveweight differences observed between the ram and wether lambs. For the cohort lambs, which were selected for comparable final curfew liveweight, the wether lamb carcasses were again found to be fattier, having higher fat tissue and lower lean tissue percentages when compared with ram lambs. Kemp et al. (1972) support this finding, having reported that fat coverage over the spine and over the LL of lamb carcasses is not affected by the interaction between castration status and liveweight. These results confirm the dose-dependent relationship between testosterone and carcass fat deposition that has been reported for male lambs (Schanbacher et al. 1980; Seideman et al. 1982; Sales 2014). Carcass fatness may in turn impact on its thermal properties when refrigerated (Smith et al. 1976), and therefore the rate of temperature decline and post-mortem glycolysis in the muscles. This was observed in the current study from the variations to final pH between ram and wether lambs. Final pH is also associated with fresh colour (Calnan et al. 2016), thereby offering a potential pathway for the difference in yellowness and colour intensity (hue) observed in the meat of wether and ram lambs.
Consumers ranked the meat of wethers as being more tender and of greater overall liking than the meat of ram lambs. These findings may reflect the effect of testosterone on collagen synthesis and turnover in lamb muscles. Wether muscles are reported to have lower collagen concentrations compared with those of ram lambs (Miller et al. 1989), and collagen concentrations are associated with lamb meat tenderness (Starkey et al. 2016). Although collagen type and concentration were not analysed in the current study, other laboratory measures for tenderness (shear force, sarcomere length, NIR-IMF) indicated no difference between the meat of wether and ram lambs. The comparative contributions of collagen to estimating consumer acceptance of red meat tenderness is reportedly lesser than the contributions of shear force, particle size and sarcomere length (Holman et al. 2020b). Further, past research has found the meat of wether lambs has higher overall acceptability and comparable tenderness to meat of ram lambs (Purchas et al. 1979). Hoffman et al. (2020) reported no difference in the tenderness and higher flavour scores for the meat of ram lambs, compared with wether lambs, and when reared under a free-range (extensive) production system. These reports and the use of an untrained consumer sensory panel could suggest a ‘halo’ effect was observed; whereby consumers have misassigned scores to a specific organoleptic parameter because of the influence of another (e.g. flavour and tenderness; Pannier et al. 2018). Consumer sensitivity or preferences for lamb meat could have likewise affected sensory panel scores (Thompson et al. 2005b; O’Reilly et al. 2020), meaning that the demographics of the sensory panel must be considered when translating these findings to other populations. Alternatively, by balancing each castration status group by liveweight, the potential influence of their interaction on cohort lamb sensory and meat quality parameters may have been obfuscated.
Purge loss and PHU were highest for wether lamb meat, but only after 32 days of wet ageing, with no other meat quality parameters affected by castration status. Research has demonstrated that most water is held within the myofibrils (Huff-Lonergan and Lonergan 2005) – these being protein structures that degrade with increases to the wet ageing period, and thereby offer less resistance to the negative pressure applied during vacuum packaging (Warner 2023). The aforementioned difference in collagen concentration suggests there are more myofibrils within the structural matrix of wether meat, and that the degradation of these protein structure, unlike collagen, would reduce the amount of bound water and contribute to the observed increase in purge loss. Microbial proliferation is also enhanced when meat has high levels of water activity or when meat is held for long wet ageing periods (Mills et al. 2014). These characteristics, measured herein as purge loss, could have contributed to the variance in PHU observed between aged wether and ram lamb meat, with lactic acid bacterium especially associated with anaerobic storage periods and changes to the pH of meat products (Mills et al. 2014; Pothakos et al. 2015). It was observed, however, that the real difference in purge loss and PHU values between wether and ram lamb meat were minor, and that these results aligned those reported in the literature (De Brito et al. 2016; Coombs et al. 2017a; Holman et al. 2024).
Colour stability was not impacted by castration status. This is important, as intrinsic and extrinsic factors have been shown to affect the colour stability of lamb meat, including animal genotype, feeding systems, methods for slaughter, post-mortem storage and more (Mortimer et al. 2010; Calnan et al. 2016; Coombs et al. 2017b; Ramanathan et al. 2020; Nemati et al. 2024). The broad basis for these effects is their impact on lamb biochemistry – specifically, the ‘redox shifts in myoglobin forms that infer redness (bloom) with the accumulation of oxymyoglobin and the gradual browning or discolouration with the accumulation of metmyoglobin’ (Suman and Joseph 2013). The absence of any castration status effects confirmed that meat myoglobin biochemistry is comparable between ram and wether lambs (Dransfield et al. 1990; Sales 2014). The observed changes to colour parameters across the display period confirms the trends reported in the literature for lamb LL displayed under these same conditions (De Brito et al. 2016; Warner et al. 2017; Holman et al. 2021). Further, when compared with consumer thresholds, it can be observed that both ram and wether lamb meat, in this study, maintained a satisfactory colour (a* values > 14.5) for a display period of up to 2 days (Khliji et al. 2010).
Conclusions
This study demonstrated that ram lambs grow faster than wether lambs, during the pre-weaning period, and achieve heavier liveweights at slaughter. Within a cohort balanced by final curfew liveweight, wether lamb carcasses were found to be fattier, having higher fat tissue and lower lean tissue percentages that may have contributed to their variance in final pH and fresh colour. Purge loss and PHU were higher for the meat of wether lambs, but only a wet aged period of 32 days post-mortem. Untrained consumer sensory scores ranked the meat of wethers as being more tender and of greater overall liking than the meat of ram lambs – even though few of the measured meat quality parameters were observed to be affected by lamb castration status. Furthermore, colour stability was not impacted by castration status, with trends reflecting past research of the effects of display period. These current findings would be complemented by future research of age at slaughter by castration status effects on male lamb meat quality parameters. Although the sampled cohort numbers, sensory panel demographics/preferences and the extensive feed base used in this study may be limiting factors, this study confirmed there to be few tangible production and meat quality advantages to the castration of male lambs. Prime lamb producers must, however, consider flock management (e.g. the ability to manage entire males separately from ewe lambs) and market preference when deciding whether to castrate.
Declaration of funding
This study was funded by the Fred Morley Centre and Graham Centre for Agricultural Innovation (now Gulbali Institute).
Acknowledgements
The authors acknowledge the staff of Charles Sturt University Farm, Gundagai Meat Processors and NSW Department of Primary Industries. The technical contributions of Dr Amy Bates, Dr Emma Lynch, Lynne Hayes, Jake Bourlet, Claire Marriott and Kristy Bailes to this study were particularly appreciated.
References
Animal Health Australia (2016) The Australian animal welfare standards and guidelines for sheep. Animal Health Australia. Available at www.animalwelfarestandards.net.au
Arnold AM, Peralta JM, Thonney ML (1997) Effect of testosterone on differential muscle growth and on protein and nucleic acid concentrations in muscles of growing lambs. Journal of Animal Science 75(6), 1495-1503.
| Crossref | Google Scholar | PubMed |
Bailes KL, Meyer RG, Piltz JW (2022) Prediction of the intramuscular fat and protein content of freeze-dried ground meat from cattle and sheep using near-infrared spectroscopy (NIRS). International Journal of Food Science & Technology 57(4), 2249-2256.
| Crossref | Google Scholar |
Bates AL, McGrath SR, Allworth MB, Robertson SM, Refshauge G (2023) A cross-sectional study of commercial ewe management practices for different sheep breeds across Southern Australia. Animals 13(3), 388.
| Crossref | Google Scholar | PubMed |
Bhatti MA, Williams T, Hopkins DL, Asheim LJ, Steinheim G, Campbell M, Eik LO, Wynn PC, Ådnøy T (2019) Adapting seasonal sheep production to year-round fresh meat and halal market in Norway. Sustainability 11(6), 1554.
| Crossref | Google Scholar |
Bouton PE, Carrol FD, Harris PV, Shorthose WR (1973) Influence of pH and fiber contraction state upon factors affecting the tenderness of bovine muscle. Journal of Food Science 38(3), 404-407.
| Crossref | Google Scholar |
Calnan H, Jacob RH, Pethick DW, Gardner GE (2016) Production factors influence fresh lamb longissimus colour more than muscle traits such as myoglobin concentration and pH. Meat Science 119, 41-50.
| Crossref | Google Scholar | PubMed |
Champness MR, McCormick JI, Bhanugopan MS, McGrath SR (2021) Sodium deficiency in lucerne (Medicago sativa) forage in southern Australia and the effect of sodium and barley supplementation on the growth rate of lambs grazing lucerne. Animal Production Science 61(11), 1170-1180.
| Crossref | Google Scholar |
CIE (1977) CIE recommendations on uniform color spaces, color-difference equations, and metric color terms. Color Research & Application 2(1), 5-6.
| Crossref | Google Scholar |
Connaughton SL, Williams A, Anderson F, Kelman KR, Gardner GE (2020) Dual energy X-ray absorptiometry precisely and accurately predicts lamb carcass composition at abattoir chain speed across a range of phenotypic and genotypic variables. Animal 14(10), 2194-2202.
| Crossref | Google Scholar | PubMed |
Coombs CEO, Holman BWB, Collins D, Friend MA, Hopkins DL (2017a) Effects of chilled-then-frozen storage (up to 52 weeks) on lamb M. longissimus lumborum quality and safety parameters. Meat Science 134, 86-97.
| Crossref | Google Scholar | PubMed |
Coombs CEO, Holman BWB, Friend MA, Hopkins DL (2017b) Long-term red meat preservation using chilled and frozen storage combinations: a review. Meat Science 125, 84-94.
| Crossref | Google Scholar | PubMed |
Crouse JD, Busboom JR, Field RA, Ferrell CL (1981) The effects of breed, diet, sex, location and slaughter weight on lamb growth, carcass composition and meat flavor. Journal of Animal Science 53(2), 376-386.
| Crossref | Google Scholar |
De Brito GF, McGrath SR, Holman BWB, Friend MA, Fowler SM, van de Ven RJ, Hopkins DL (2016) The effect of forage type on lamb carcass traits, meat quality and sensory traits. Meat Science 119, 95-101.
| Crossref | Google Scholar | PubMed |
Dransfield E (1994) Optimisation of tenderisation, ageing and tenderness. Meat Science 36(1–2), 105-121.
| Crossref | Google Scholar | PubMed |
Dransfield E, Nute GR, Hogg BW, Walters BR (1990) Carcass and eating quality of ram, castrated ram and ewe lambs. Animal Science 50(2), 291-299.
| Crossref | Google Scholar |
Gardner GE, Williams A, Ball AJ, Jacob RH, Refshauge G, Hocking Edwards J, Behrendt R, Pethick DW (2015) Carcase weight and dressing percentage are increased using Australian sheep breeding values for increased weight and muscling and reduced fat depth. Meat Science 99, 89-98.
| Crossref | Google Scholar | PubMed |
Gkarane V, Allen P, Gravador RS, Diskin MG, Claffey NA, Fahey AG, Brunton NP, Farmer LJ, Moloney AP, Monahan FJ (2017) Effect of castration and age at slaughter on sensory perception of lamb meat. Small Ruminant Research 157, 65-74.
| Crossref | Google Scholar |
Gravador RS, Pace E, Mooney BR, Jaeger SR, Gkarane V, Fahey AG, Brunton NP, Claffey NA, Allen P, Diskin MG, Moloney AP, Farmer LJ, Monahan FJ (2018) A consumer study of the effect of castration and slaughter age of lambs on the sensory quality of meat. Small Ruminant Research 169, 148-153.
| Crossref | Google Scholar |
Hoffman LC, Claasen B, van der Merwe DA, Cloete SWP, Cloete JJE (2020) The effects of production system and sex on the sensory quality characteristics of dorper lamb. Foods 9(6), 725.
| Crossref | Google Scholar |
Holman BWB, Alvarenga TIRC, van de Ven RJ, Hopkins DL (2015a) A comparison of technical replicate (cuts) effect on lamb Warner–Bratzler shear force measurement precision. Meat Science 105, 93-95.
| Crossref | Google Scholar | PubMed |
Holman BWB, Ponnampalam EN, van de Ven RJ, Kerr MG, Hopkins DL (2015b) Lamb meat colour values (HunterLab CIE and reflectance) are influenced by aperture size (5mm v. 25mm). Meat Science 100, 202-208.
| Crossref | Google Scholar | PubMed |
Holman BWB, Alvarenga TIRC, Hopkins DL (2020a) The effect of fibre orientation, measurement interval and muscle on lamb meat drip loss values. Meat Science 161, 107959.
| Crossref | Google Scholar |
Holman BWB, Collins D, Kilgannon AK, Hopkins DL (2020b) Using shear force, sarcomere length, particle size, collagen content, and protein solubility metrics to predict consumer acceptance of aged beef tenderness. Journal of Texture Studies 51(4), 559-566.
| Crossref | Google Scholar | PubMed |
Holman BWB, Hayes RC, Newell MT, Refshauge G, McGrath SR, Fowler SM, Shanley AR, Hopkins DL (2021) The quality and mineral composition of the longissimus lumborum and semimembranosus muscles from lambs fed perennial or annual wheat forage with or without lucerne. Meat Science 180, 108564.
| Crossref | Google Scholar |
Holman BWB, Refshauge G, Newell MT, Hopkins DL, Hayes RC (2024) The carcass characteristics and quality of meat from lambs grazing perennial wheat with different companion legumes (clover, serradella, lucerne) or a mineral supplement. Meat Science 216, 109549.
| Crossref | Google Scholar | PubMed |
Hopkins DL, Thompson JM (2001) The relationship between tenderness, proteolysis, muscle contraction and dissociation of actomyosin. Meat Science 57(1), 1-12.
| Crossref | Google Scholar | PubMed |
Hopkins DL, Gilbert KD, Pirlot KL, Roberts AHK (1992a) Elliotdale and crossbred lambs: growth rate, wool production, fat depth, saleable meat yield, carcass composition and muscle content of selected cuts. Australian Journal of Experimental Agriculture 32(4), 429-434.
| Crossref | Google Scholar |
Hopkins DL, Jackson RB, Roberts AHK (1992b) Comparison of a modified cryptorchid treatment and castration: effect on growth, wool production, posthitis, testosterone production and development of masculine characteristics. Australian Journal of Experimental Agriculture 32(4), 443-446.
| Crossref | Google Scholar |
Huff-Lonergan E, Lonergan SM (2005) Mechanisms of water-holding capacity of meat: the role of postmortem biochemical and structural changes. Meat Science 71(1), 194-204.
| Crossref | Google Scholar | PubMed |
Kelman KR, Alston-Knox C, Pethick DW, Gardner GE (2022) Sire breed, litter size, and environment influence genetic potential for lamb growth when using sire breeding values. Animals 12(4), 501.
| Crossref | Google Scholar |
Kemp JD, Shelley JM, Jr, Ely DG, Moody WG (1972) Effects of castration and slaughter weight on fatness, cooking losses and palatability of lamb. Journal of Animal Science 34(4), 560-562.
| Crossref | Google Scholar |
Khliji S, van de Ven R, Lamb TA, Lanza M, Hopkins DL (2010) Relationship between consumer ranking of lamb colour and objective measures of colour. Meat Science 85(2), 224-229.
| Crossref | Google Scholar | PubMed |
Lee GJ, Harris DC, Ferguson BD, Jelbart RA (1990) Growth and carcass fatness of ewe, wether, ram and cryptorchid crossbred lambs reared at pasture: effects of weaning age. Australian Journal of Experimental Agriculture 30(6), 743-747.
| Crossref | Google Scholar |
Mahgoub O, Lodge GA (1994) Growth and body composition of Omani local sheep 1. Live-weight growth and carcass and non-carcass characteristics. Animal Science 58(3), 365-372.
| Crossref | Google Scholar |
Miller LF, Judge MD, Diekman MA, Hudgens RE, Aberle ED (1989) Relationships among intramuscular collagen, serum hydroxyproline and serum testosterone in growing rams and wethers. Journal of Animal Science 67(3), 698-703.
| Crossref | Google Scholar | PubMed |
Mills J, Donnison A, Brightwell G (2014) Factors affecting microbial spoilage and shelf-life of chilled vacuum-packed lamb transported to distant markets: a review. Meat Science 98(1), 71-80.
| Crossref | Google Scholar | PubMed |
Mortimer SI, Van Der Werf JHJ, Jacob RH, Pethick DW, Pearce KL, Warner RD, Geesink GH, Hocking Edwards JE, Gardner GE, Ponnampalam EN, Kitessa SM, Ball AJ, Hopkins DL (2010) Preliminary estimates of genetic parameters for carcass and meat quality traits in Australian sheep. Animal Production Science 50(12), 1135-1144.
| Crossref | Google Scholar |
Nemati Z, Amirdahri S, Asgari A, Taghizadeh A, Siddiqui SA, Besharati M, Alirezalu K, Holman BWB (2024) Feeding pomegranate pulp to Ghezel lambs for enhanced productivity and meat quality. Veterinary and Animal Science 24, 100356.
| Crossref | Google Scholar | PubMed |
O’Reilly RA, Pannier L, Gardner GE, Garmyn AJ, Luo H, Meng Q, Miller MF, Pethick DW (2020) Influence of demographic factors on sheepmeat sensory scores of American, Australian and Chinese consumers. Foods 9(4), 529.
| Crossref | Google Scholar |
Pannier L, Gardner GE, O’Reilly RA, Pethick DW (2018) Factors affecting lamb eating quality and the potential for their integration into an MSA sheepmeat grading model. Meat Science 144, 43-52.
| Crossref | Google Scholar | PubMed |
Pearce KL, Van De Ven R, Mudford C, Warner RD, Hocking-Edwards J, Jacob R, Pethick DW, Hopkins DL (2010) Case studies demonstrating the benefits on pH and temperature decline of optimising medium-voltage electrical stimulation of lamb carcasses. Animal Production Science 50(12), 1107-1114.
| Crossref | Google Scholar |
Pothakos V, Devlieghere F, Villani F, Björkroth J, Ercolini D (2015) Lactic acid bacteria and their controversial role in fresh meat spoilage. Meat Science 109, 66-74.
| Crossref | Google Scholar | PubMed |
Purchas RW (1978) Some effects of nutrition and castration on meat production from male Suffolk cross (border leicester-romney cross) lambs: I. growth and carcass quality. New Zealand Journal of Agricultural Research 21(3), 367-376.
| Crossref | Google Scholar |
Purchas RW, O’Brien LE, Pendleton CM (1979) Some effects of nutrition and castration on meat production from male Suffolk cross (border leicester-romney cross) lambs: II. Meat quality. New Zealand Journal of Agricultural Research 22(3), 375-383.
| Crossref | Google Scholar |
Ramanathan R, Hunt MC, Mancini RA, Nair MN, Denzer ML, Suman SP, Mafi GG (2020) Recent updates in meat color research: integrating traditional and high-throughput approaches. Meat and Muscle Biology 4(2), 1-24.
| Crossref | Google Scholar |
Sales J (2014) Quantification of the effects of castration on carcass and meat quality of sheep by meta-analysis. Meat Science 98(4), 858-868.
| Crossref | Google Scholar | PubMed |
Schanbacher BD, Crouse JD, Ferrell CL (1980) Testosterone influences on growth, performance, carcass characteristics and composition of young market lambs. Journal of Animal Science 51(3), 685-691.
| Crossref | Google Scholar | PubMed |
Seideman SC, Cross HR, Oltjen RR, Schanbacher BD (1982) Utilization of the intact male for red meat production: a review. Journal of Animal Science 55(4), 826-840.
| Crossref | Google Scholar |
Small AH, Jongman EC, Niemeyer D, Lee C, Colditz IG (2020) Efficacy of precisely injected single local bolus of lignocaine for alleviation of behavioural responses to pain during tail docking and castration of lambs with rubber rings. Research in Veterinary Science 133, 210-218.
| Crossref | Google Scholar | PubMed |
Smith GC, Dutson TR, Hostetler RL, Carpenter ZL (1976) Fatness, rate of chilling and tenderness of lamb. Journal of Food Science 41(4), 748-756.
| Crossref | Google Scholar |
Starkey CP, Geesink GH, Collins D, Hutton Oddy V, Hopkins DL (2016) Do sarcomere length, collagen content, pH, intramuscular fat and desmin degradation explain variation in the tenderness of three ovine muscles? Meat Science 113, 51-58.
| Crossref | Google Scholar | PubMed |
Suman SP, Joseph P (2013) Myoglobin chemistry and meat color. Annual Review of Food Science and Technology 4, 79-99.
| Crossref | Google Scholar | PubMed |
Thompson JM, Gee A, Hopkins DL, Pethick DW, Baud SR, O’Halloran WJ (2005a) Development of a sensory protocol for testing palatability of sheep meats. Australian Journal of Experimental Agriculture 45(5), 469-476.
| Crossref | Google Scholar |
Thompson JM, Pleasants AB, Pethick DW (2005b) The effect of design and demographic factors on consumer sensory scores. Australian Journal of Experimental Agriculture 45(5), 477-482.
| Crossref | Google Scholar |
Warner RD, Kearney G, Hopkins DL, Jacob RH (2017) Retail colour stability of lamb meat is influenced by breed type, muscle, packaging and iron concentration. Meat Science 129, 28-37.
| Crossref | Google Scholar | PubMed |
Wellington GH, Hogue DE, Foote RH (2003) Growth, carcass characteristics and androgen concentrations of gonad-altered ram lambs. Small Ruminant Research 48(1), 51-59.
| Crossref | Google Scholar |
Young OA, Lane GA, Podmore C, Fraser K, Agnew MJ, Cummings TL, Cox NR (2006) Changes in composition and quality characteristics of ovine meat and fat from castrates and rams aged to 2 years. New Zealand Journal of Agricultural Research 49(4), 419-430.
| Crossref | Google Scholar |