

Livestock preference and feeding value as key determinants for forage improvement – why not ask the consumers?
Hayley C. Norman A * and David G. Masters A
A * and David G. Masters A
A CSIRO Agriculture and Food, 147 Underwood Avenue, Floreat, WA 6014, Australia.
Animal Production Science 63(12) 1161-1176 https://doi.org/10.1071/AN23140
Submitted: 14 April 2023 Accepted: 6 June 2023 Published: 4 July 2023
© 2023 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
This review presents a new model for a multidisciplinary approach to development of modern forage cultivars to enable high-value meat and wool production in the mixed farming zones of Australia. Many pasture improvement activities focus on biomass production, filling novel agro-ecological niches, fixing nitrogen and phenotypic markers. Key traits that drive profitability of livestock, such as feeding value (FV), and nutritive value (NV), are rarely considered in a timely manner. Farmers are often not seriously engaged until late in the development pipeline. Of the nearly 10 000 publications that reference forage improvement or breeding and Australia, less than 5% also reference ruminant production factors such as FV, metabolisable energy, digestibility, methane or toxicity in the abstract, title or keywords. We make the case that the late engagement of livestock science and farmers results in early discarding of potentially valuable genetic material. This in turn may contribute to market and nutritional limitations that contribute to the high failure rate of new pasture cultivars. Four case studies demonstrated the within and among species variation in FV and NV. The first showed the variability in dry matter digestibility and crude protein of 20 species of herbaceous perennials at different stages of maturity. A second study with annual legumes, grasses and brassicas indicated highly significant differences in the rate of decline of NV over time and across species. The third case study focused on commercial and experimental accessions of lucerne and demonstrated highly significant differences among accessions in NV, with a doubling of livestock growth expected with the lowest and highest NV accessions. These case studies demonstrated the potential risks associated with evaluation of FV and NV after most of the germplasm has been excluded. The final case study described a different model that was used in the identification and commercialisation of a woody perennial species. This process incorporated a multidisciplinary team, farmers and livestock-relevant characteristics from the start. In 9 years, Anameka™ oldman saltbush was selected for higher palatability, 20% higher organic-matter digestibility and eight times more biomass than the average of the original genetic material.
Keywords: cattle, economic value, ley farming, plant breeding, plant quality, production system, silvopastrolism, voluntary feed intake.
Introduction
This paper was prepared for the 2021 Harry Stobbs Memorial lecture for the Australian Association of Animal Sciences. Dr Harry Stobbs was committed to understanding grazing behaviour of ruminants and effective communication between farmers and scientists. In pursuing the Harry Stobbs tenet, this paper will explore the role of livestock in forage development in Australia and draw on some case studies to describe opportunities to enhance plant domestication, breeding and improvement programs. The importance of multidisciplinary approaches will be highlighted and a case for the early inclusion of livestock scientists and systematic measurement of feeding value (FV) during forage development will be presented. Simultaneously, early engagement with farmers and a need to understand the role of the new plant within the mixed farming system will be discussed. Such multidisciplinary approaches can provide more opportunity to capitalise on variation within and among candidate forage species and minimise the failure of many forage cultivars due to a lack of on-farm practicality.
Forage production and improvement in the mixed farming zones
Livestock and pastures in the mixed farming zone
The mixed farming zone (MFZ) in Australia (300–650 mm of annual rainfall) includes some 21 000 farmers and covers approximately 57 million ha of farmland (Bell et al. 2019). This zone is the hub of the red meat industry, providing maternal and finishing capacity from pastures and crops, as well as grain and conserved fodder for use in other areas. Livestock in this zone contribute to the management of seasonal risk, offer opportunities to utilise poorer soils and crop by-products, and may assist cropping through soil improvement, a disease break, water use and weed management (Bell et al. 2019). Despite these benefits, there has been a dramatic decline is sheep numbers in Australia from over 160 million in 1990/1991 to 67.7 million in 2018 (Perret 2015; Meat and Livestock Australia 2019), with the greatest decline in the MFZ. This has been accompanied by greater social and environmental pressures on farming, as well as changes in flock structure. National cattle numbers have remained steady at 23.2 to 26.4 million over the same time period. There has been an increasing trend for a greater emphasis on lamb production in systems, with lambs representing just under 23% of the sheep flock in 1990–1998 and 31% of the flock in 2013–2021 (Meat and Livestock Australia 2023). These factors increase the emphasis for higher FV diets to optimise lamb growth.
The decline in livestock numbers, combined with climate variability, soil acidity, salinity, changing cropping practices and emerging diseases, has led to a decline in pasture systems based on subterranean clover (Trifolium subterraneum L.) and annual medics (Medicago spp.) and the most commonly planted perennial legume, lucerne (alf-alfa, Medicago sativa L.; Fortune et al. 1995; Howieson et al. 2000; Cocks 2001; Hackney et al. 2019). Over the same period, the area cropped has increased by more than 60% (Perret 2015). Together with lower whole-farm stocking rates (Bell and Moore 2012), this has resulted in many pastures becoming a fallow dominated by weedy volunteer species with poor nutritional value and suboptimal break-crop benefits for cereal production (Lawes et al. 2022). Modelling, focused on the MFZ of Western Australia, indicates that fallows are unlikely to increase overall grain productivity, with the exception of very low rainfall, weedy or diseased paddocks (Chen et al. 2023).
There is a strong case for change towards better self-regenerating legume pastures and an increase in sheep numbers in the MFZ. Recent economic analysis, based on whole-farm profit of mixed enterprises, highlighted the value of livestock, indicating that the optimal cropping proportion of the farm is between 40% and 60% (Young et al. 2020). This is a scenario where livestock occupy ~50% of the farm and obtain nutritional benefits from cropped land through grazing young crops and residues. Similarly, others have reported that whole-farm stocking rates are a major driver of profitability, with modelling showing that an increase in stocking rates of 20% from a conservative baseline increases the livestock enterprise gross margin by between 11% and 17% (Friend et al. 2018). The increase in stocking rates needs to be sustainable and will be facilitated by more productive and flexible pastures; otherwise, overgrazing will also decrease whole-farm profit (Young et al. 2022). The major weaknesses of these ley farming systems are the summer–autumn feed gap for livestock and hydrological imbalances that lead to secondary salinisation of soils (Masters et al. 2005; Edwards et al. 2019).
Even without an increase in stocking rate, new and better-adapted perennial legumes have been predicted to increase whole-farm profit in the MFZ by 26% (Bathgate et al. 2009). Perennials offer an opportunity to improve farm profitability by utilising the infrequent summer–autumn rainfall events for production of high-value feed at a time that it is scarce (Ridley and Pannell 2005; Moore 2014; Edwards et al. 2019). In Mediterranean-climate zones of southern and western Australia, pastures providing carry-over or green feed in summer and autumn can have an economic value of up to 10 times that of equivalent quality feed in spring (Morrison and Bathgate 1990); seasonal variation in plant growth and composition significantly limits stocking rates and profitability (Moore et al. 2009). Perennial forages can reduce flexibility in cropping rotations; so, a focus on their use on land that is marginal for crop production, such as acidic, sandy, saline, waterlogged or rocky land is practical and has a lower opportunity cost (Monjardino et al. 2014; Edwards et al. 2019). Perennials may offer additional benefits to livestock systems through provision of shade, shelter and nutrients that assist animals to manage oxidative stress (Masters et al. 2023).
There is some intent to increase livestock numbers in the MFZ; for example, a survey of 170 farmers in New South Wales found that 76% of farmers have long-term goals to increase stocking rates, but their current feedbase limits capacity to support livestock production goals more than 50% of the time (Hackney et al. 2017). This imbalance between intent to increase stocking rates and capacity to meet livestock needs leads to increased economic, animal welfare and environmental risks.
Recent plant improvement
Nichols et al. (2012) reported that there had been 300 forage legume cultivars registered for the temperate and Mediterranean-climate in Australia since 1920, comprising 36 annual and 11 perennial species (Nichols et al. 2012; IP Australia 2022). In the past decade, an additional 20 annual and 20 perennial legume cultivars were registered (IP Australia 2022). The most common types have been lucerne (77 cultivars), annual medics (46 cultivars) and subterranean clover (43 cultivars). When reading the plant breeders’ criteria for commercialisation, the majority has been selected for ‘vigour’, seed-dormancy traits, disease resistance, tolerance to abiotic stresses, nutrient-use efficiency or characteristics of the phenotypes (leaf marks etc.) that allow the cultivar to achieve Plant Breeders Rights by being distinctive. The failure rate for these legume cultivars has been high, the vast majority of these have been superseded or have failed to achieve industry adoption. While it is a major challenge to develop new pasture species that optimise livestock production, persistence (or have traits that lead to inexpensive seed for resowing), have adapted rhizobia (for legumes), are suited to ever-evolving cropping systems and outperform historical cultivars, the needs of livestock appear to have been a very low priority in recent pasture improvement programs. Is the emphasis on distinctiveness, a trait that arguably has little to do with FV, hampering efforts to improve the feedbase in the MFZ of Australia?
In response to industry consultation that led to a greater understanding to barriers to adoption, recent multidisciplinary programs such as Dryland Pasture Legume Systems aimed to improve pasture legumes for livestock productivity and cropping system benefits in the MFZ (https://grdc.com.au/grdc-investments/investments/investment?code=UMU1805-001RMX). Prior to this, much of the focus has been for traits that reduce costs of establishment and improve persistence and biological nitrogen fixation within cereal production systems. The focus on establishment cost and specific agro-ecological niches was an example of effective communication between the plant breeders and farmers. In the past two decades, there has been a number of novel, self-regenerating (via seed dormancy) species of annual legumes (with appropriate root-nodule rhizobia) domesticated for specific climatic and edaphic niches within systems. In response to farmer concerns regarding the cost of establishment (and associated financial risk), they have been selected to allow seed harvest with conventional cereal-harvesting machines. Biomass production during the establishment year (and subsequent opportunity cost associated with a low-yielding pasture) was another barrier to resowing pastures. Some species can be sown as dormant seed in summer, leading to 10 times greater biomass production in the establishment year (Nutt et al. 2021). This second generation of annual legumes includes cultivars of serradella (Ornithopus compressus L. and O. sativus Brot), biserrula (Biserrula pelecinus L.), bladder clover (Trifolium spumosum L.), gland clover (Trifolium glanduliferum Boiss.) and more recently Trigonella balansae Boiss. and Reuter (Howieson et al. 2000; Loi et al. 2005; Wilmot et al. 2023). These annual species were not selected for FV or nutritional value (NV). As is often the case, biomass production in spring, a time when forage is plentiful, was generally used as a proxy for FV. Pasture breeding objectives tend to be much more complex than the methods applied in animal breeding, where profit-based breeding indices are common (Smith et al. 2014; Thomas et al. 2021).
Designing pasture improvement pathways
There is no doubt that some Australian scientists have conducted world-leading research in domesticating new species to improve profitability of mixed farming systems. However, the high rate of cultivar failure and continued decline in livestock numbers, with simultaneous loss in potential farm productivity and profitability, indicates that we need to challenge traditional forage improvement programs for the MFZ. The 10 steps that are undertaken during conventional annual legume domestication (Fig. 1) were articulated by Mike Ewing, a veteran of pasture legume development (Ewing 2017). A similar framework was described by Nichols et al. (2010). In this model, measurement of FV, NV, and secondary compounds may occur in Step 8 of the pipeline, many years into the domestication or cultivar selection process and when the majority of accessions have been culled. This step appears to be more of a pass/fail step than one aimed at optimisation.
Traditional steps for the domestication and improvement of new pasture species as described in Ewing (2017). Note some of these stages can occur concurrently.

FV is the worth of a forage in terms of animal production, it is a function of how much of the forage is eaten and the utilisation of that forage. NV is defined by the nutrient content or animal response per unit of feed intake (Ulyatt 1973). These characteristics of forage are critical for livestock production, but are sometimes not even considered at all before commercial release. Bell et al. (2019) listed six specific characteristics that have been the focus in the development of the next generation of annual legumes; none of the priorities relates to the value of these plants as feed for livestock. By the time livestock are considered, the number of species or genotypes under evaluation is likely to have reduced from thousands to tens (referred to as ‘advanced material’ in the Ewing model). This appears incongruous, given the pasture has economic value only when converted into meat, milk or wool to complement any other farming-system benefits, such as nitrogen fixation and as a disease break. In this model, there seems to be an inherent belief that ‘green is good’, that livestock requirements are defined by biomass and that all legumes are equally as nutritious and palatable (i.e. no variation in FV or NV) as long as they do not contain secondary compounds that are toxic. Voluntary intake, energy, protein, minerals and plant–animal interactions are largely ignored.
In the model (Fig. 1), farmer input is not overtly sought until Stage 10, when it is time to develop management packages for commercialised material (Ewing 2017). It seems incongruous that the ‘end users’, the farmers, and the livestock they manage have very little early input to the plant domestication and improvement process. Of course, this model does not reflect how all plant breeders operate and there are cases where farmers have been involved much earlier in the improvement pipeline.
Another very recent example of using a narrow focus to reduce the pool of candidate genotypes, assuming that other traits will also be delivered, has been presented in Badgery et al. (2023). In this review, the authors suggested a very similar inverted-triangle framework to develop pasture plants with anti-methanogenic potential. In this case, the first heavy-culling exercise is based on selection of plants with secondary compounds that may have methane-reduction potential (e.g. tannins or saponins), with in vitro screening for methane per unit of DM. The second culling step is for candidate species and genotypes to be tested for agronomic suitability in the field. Animal production factors such as as energy, crude protein, minerals and fibre are considered in the third step with pen feeding trials. Farmers, economists and system modellers are presumably included in the last ‘integrated-system’ step. While it is admirable that a component of animal performance (methane per unit DM) is part of the broad initial screening, this process could just as easily progress a cohort of plants with high tannin that have very low digestibility and bound protein, or high saponin and are not eaten by stock. These options are likely to be poorly adopted by farmers and could negate the initial goal of the breeding program as attainment of target liveweight or condition scores for slaughter could occur at an older age with higher lifetime methane emissions. It is illogical to wait until the pool is shallow before questions regarding farmer needs and how the cultivar fits within a farming system are addressed. At the very least, the authors should assess methane emissions in the context of plant metabolisable energy (ME) rather than DM.
The examples provided above are based on legume development and the reader may be tempted to believe that this thinking is the exception and not the norm. Of the 9647 publications identified from a Web of ScienceTM search (May 2022 (Clarivate 2022) defined by ‘forage improvement OR pasture improvement OR pasture breeding OR forage breeding AND Australia’, less than 5% reference ruminant production factors such as ‘FV’ ‘ME’, ‘digestibility’, ‘crude protein’, ‘methane’ or ‘toxicity’ in the abstract, title or keywords (Table 1). ‘Intake’ is mentioned more than any other component of FV or NV (9.2%), yet it is the hardest trait to measure in grazing ruminants. Acknowledging that this method is crude and the search engine picks up erroneous publication, the numbers are still indicative of a disconnect between plant-improvement activities and the needs of livestock.
| Search topic | n | % |
|---|---|---|
| Forage improvement OR pasture improvement OR pasture breeding OR forage breeding AND Australia | 9647 | |
| +Intake, or | 891 | 9.2 |
| +Crude protein, or | 466 | 4.8 |
| +Feeding value, or | 245 | 2.5 |
| +Nutritional value, or | 217 | 2.2 |
| +Methane, or | 179 | 1.9 |
| +Liveweight gain, or | 99 | 1.0 |
| +Digestibility, or | 98 | 1.0 |
| +Toxicity, or | 68 | 0.7 |
| +Metabolisable energy, or | 60 | 0.6 |
| +Secondary compounds | 3 | 0.0 |
When reading these papers, it is rare for FV or NV to be measured near the start of plant selection. The major exception is when there is a known toxicity that the plant improvement program is attempting to overcome, such as isoflavones in subterranean clover (Gildersleeve et al. 1991) or tannins in lotus (Kelman et al. 2007). Another exception is studies where mixed swards were managed to have higher legume content to improve FV (Reed 2014); however, these efforts focused on composition of taxonomic groups, not optimising FV or NV of the individual components. Aspects of NV are more likely to be assessed in perennial species for intensive dairy production systems. For example, energy value can be improved by gene-technology methods leading to perennial ryegrass genotypes that produce more water-soluble carbohydrates (Rasmussen et al. 2014) or lucerne with altered lignin content (Zhao and Dixon 2014). FV is generally assumed by measuring dry matter production from ungrazed swards at peak biomass in spring (e.g. Loi et al. 2015) and only considered in the very last step as a means of meeting duty of care requirements of novel species prior to commercialisation (Norman et al. 2005; Masters et al. 2006; Wilmot et al. 2023). There appears to be little effort to match measurement of FV to the critical times in the livestock production cycle when it has the highest consequence.
Composition of essential minerals is rarely considered from the perspective of livestock. Masters et al. (2019) reported that, in a significant proportion of recently commercialised pasture plants, concentrations of some minerals were insufficient to support high levels of growth and reproduction for at least part of the year. The mineral data were generated from a range of species growing in two locations (Adelaide, South Australia and Brookton, Western Australia) over two seasons. The authors went further to intimate that narrow agronomic selection criteria, such as selection for phosphorus-utilisation efficiency in plants, may increase the risk of mineral deficiency in grazing livestock. Wilmot et al. (2023) found that a new Trigonella balansae cultivar, tested at only one location in one season, did not meet the minimum sulfur, phosphorus, manganese, molybdenum and copper requirements for mature sheep.
Opportunities from earlier consideration of livestock in the pasture improvement pipeline
Reed (1994) stated that ‘plant breeding and the merit-testing of grass cultivars in multi-site small-plot studies should be supported with some animal production trials if plant improvement is to maintain credibility and remain focused on feasible, commercially relevant goals’ (p. 277). This thinking seems to be the exception, rather than the norm, especially for annual species for the MFZ where persistence or phenotypic distinction traits dominate selection. When FV and NV are assessed late in the plant improvement pipeline, much of the potential for livestock production may be lost if genotypes with elite FV and NV traits have been discarded earlier in the selection process. If there is little variation among and within species, this is a logical approach. To address this assumption, data from four Australian studies that provide information on within, and among species, variation in FV or NV are considered. The studies include variability in components of NV among perennial legume species, variation in NV decline after flowering in annual legumes grasses and brassicas, variability within lucerne breeding lines and variability in ‘wild’ saltbush populations.
Case study 1: variation in digestibility within and among perennial legume species
A diverse range of commercial and late research pipeline herbaceous perennials were grown at a single site in Adelaide over two seasons (Norman et al. 2021). The experiment included 30 accessions from 20 species, planted in replicated blocks. Several cutting regimes were imposed. Every month plots were assessed for plant growth stage, biomass production and samples were taken to predict indicators of NV by using calibrated near-infrared spectroscopy (NIRS; Norman et al. 2021). Fig. 2 shows the variation in mean dry matter digestibility (DMD) and crude protein (CP, total nitrogen × 6.25). Data are presented at the vegetative, flowering and mature (ripe seed) physiological stages.
Mean dry matter digestibility (DMD, % ± s.e., top) and crude protein (CP) content (% ± s.e., bottom) of 30 accessions of perennial legumes at the vegetative (black symbols), reproductive (grey symbols) and mature (white symbols) phases. There were significant (P < 0.001) differences among accessions at each maturity stage and l.s.d. (5%) values from the ANOVA are presented for comparison. Reproduced with permission from Norman et al. (2021); © 2021 John Wiley & Sons Ltd.

At the vegetative stage of maturity, DMD of the legumes ranged from 47% to 73% and the CP content ranged from 15% to 26% DM (with a mean of 21% DM). The species with the highest DMD included white clover (Trifolium repens L.) and strawberry clover (Trifolium fragiferum L.). Even in the vegetative stage of growth, greater birdsfoot trefoil (Lotus uliginosus Schkuhr), Australian trefoil (Lotus australis Andrews), running postman (Kennedia prostrata R.Br.), hairy canary clover (Dorycnium hirsutum L.) and erect canary clover (Dorycnium rectum L.) had DMD values below 55%, which is the energy requirement for maintenance of a mature non-reproducing sheep (Freer et al. 1997). Three of these are commercialised forage cultivars. It is worth noting that the low FV Lotus and Dorycnium species would pass the first cull and be included in the Badgery et al. (2023) framework because they contain high tannin levels. There were no significant differences in DMD at the vegetative stage among the four lucerne accessions (mean 67.6%) or among the three sulla (Hedysarum coronarium L.) accessions (mean of 63.1%). There was a significant difference among the sainfoin (Onobrychis viciifolia Scop.) accessions (65.4% and 60.4% for cvv. Othello and Shoshone respectively).
As expected, the DMD and CP declined as plants matured because older leaves and stems presumably had higher cell-wall content than did young leaves (Minson 1990; Elgersma and Søegaard 2018). The rate of decline in NV among the accessions varied. The accessions that had a greater than 10% unit decline in DMD associated with progression to reproduction included lucerne and birdsfoot trefoil (Lotus corniculatus L.). At the mature development stage, DMD ranged from 40% (hairy canary clover) to 66% (white clover).
Models such as GrazFeed™ (CSIRO, Version 5.0.7., Canberra, Australia; Freer et al. 1997) allow for prediction of the relative performance of any class of ruminant grazing the forage accessions. For this study, the model class of sheep was a 6-month-old Merino wether (43 kg liveweight with 3 cm of wool growth) ‘offered’ various forages at 2 kg DM/day using the roughage-supplement function (Norman et al. 2021). The model predicted intake on the basis of the DMD and CP of the forage. Risks of bloat or influence of secondary compounds that affect intake and digestion were not accounted for. Sheep grazing days/ha were calculated from total biomass/predicted daily voluntary intake; these numbers are therefore aspirational (for comparative purposes). After 205 days of growth in the second season after sowing, white clover (cv. Storm) yielded 15 t DM/ha and had potential to support 6-month-old Merino wethers for more than 3500 grazing days/ha, with an estimated 192 g/day of liveweight change and 19.4 g/day of clean wool growth. This species will not persist in the MFZ. Lucerne (cv. SARDI 7, Series 2) offered more than 6500 sheep grazing days/ha, with a lower predicted liveweight gain and wool growth (110 g/day and 16 g/day of clean wool growth). Sulla (cv. Wilpena) offered three times more grazing days than the white clover (11 000 grazing days/ha); however, the NV was such that only modest liveweight gain (22 g/day) was predicted. These data highlighted the importance of understanding the farming-system context, class of stock and the goals of farmers.
Without energy supplementation, the 6-month-old Merino wethers offered hairy canary clover were predicted to lose 100–120 g/day of liveweight (Norman et al. 2021). Was there better material that was overlooked in pursuit of agronomic goals? Would hairy canary clover have been commercialised if there had been NV testing earlier in the development pipeline? The very poor energy value of these species was noted prior to commercialisation (Davies and Lane 2003; Bell et al. 2006); however, in 2011 the cv. Canaritas was commercialised on the basis of high seed production and ease of harvest (Lane et al. 2004; Nichols et al. 2012). This case study reiterates the importance of systematically measuring NV, in parallel with agronomic traits, in forage-improvement programs. This does not need to be onerous; NIRS methods to predict NV are relatively accurate and inexpensive (Norman et al. 2020), and many studies have found that DMD is a trait that is stable across environments (at similar maturity stages), when compared with biomass or even CP content (Shenk and Westerhaus 1982; Carpenter and Casler 1990; Cherney and Volenec 1992).
Case study 2: variation in pattern of digestibility decline during maturity of annual legumes, grasses and brassicas
In a study that was run in parallel to the perennial legume research above, 90 accessions/cultivars from 66 species of annual forages (legumes, grasses and forbs) were grown in replicated plots in Adelaide over two seasons. Biomass was sampled every 2–3 weeks over the growing season and during senescence, dry matter production was determined, and samples were analysed for DMD and other nutritional traits The pattern of change in DMD and CP during the plant lifecycle followed a logistic breakdown pattern. Digestibility curves are presented for nine cultivars, representing five annual legumes, four grasses/cereals and two brassicas (Fig. 3). All curve fits were significant (P < 0.001) and accounted for 90–98% of variation in the raw data.
Logistic decay curves demonstrating changes in DMD in annual legumes (top) and grasses and brassicas (bottom) as they mature. Plants were sown on 12 June 2012 and the first sampling (vegetative phase) was 76 days later on 26 August. Day 190 is the 18 December when all plants had senesced.
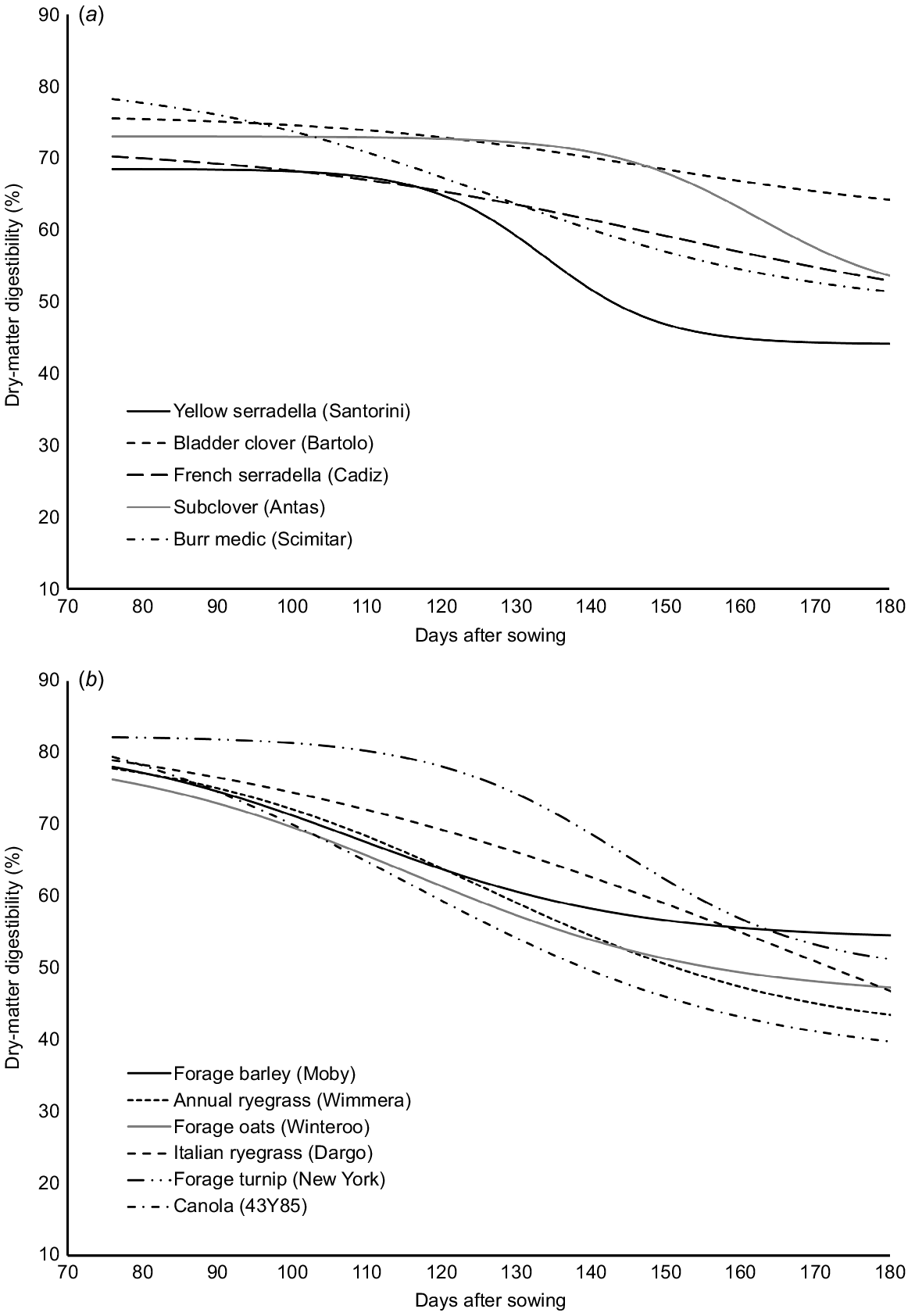
There were clear and significant differences in initial DMD, rate of DMD loss over time and the final DMD of senesced material. All but one cultivar had DMD values above 70% in the vegetative phase, and there were few differences among the cultivars at this stage. DMD decline with maturity was variable; the forage oat (cv. Winteroo) declined at a faster rate than did other species and reached a point of inflection (where rate of decline was highest) at 115 ± 5 days after sowing. The yellow serradella had a point of inflection at 134 ± 5 days. Cultivars that had a relatively slow decline in DMD included subclover (cv. Antas 162 ± 2 days), forage turnip (cv. New York 144 ± 5 days) and bladder clover (cv. Bartolo 148 ± 6 days). The bladder clover cultivar had the highest DMD after senescence (63%). At this time, the forage oat, Italian ryegrass (cv. Dargo), canola, and yellow serradella all had DMD values of <45%.
Non-reproducing ewes grazing the bladder clover on the 1 October (spring) were predicted to voluntarily consume about 1.2 kg DM/day and gain 110 g/day of liveweight (GrazFeed™ model, Freer et al. 1997). Ewes offered the yellow serradella were predicted to consume 1.1 kg of DM/day and gain a third of the liveweight (35 g/day). The contrast is much greater on the 1 December (summer), when sheep offered the bladder clover were predicted to consume 1.15 kg of DM/day and gain 50 g/day of liveweight, in contrast to sheep offered yellow serradella who were likely to consume only 0.5 kg DM/day and, subsequently, lose 140 g/day of liveweight without energy supplementation. While these are predictions and do not account for the ability of individual animals to select a higher-quality diet among the plant components, the differences are substantial.
There are several insights from these data. Measurement of FV needs to occur at a time when the pasture species are going to have the greatest value within the farming system or when they are most likely to be grazed. Ideally, it is determined across an annual plant’s lifecycle and accounts for impact of grazing and forage conservation practices. In many studies where pasture quality is measured, it is undertaken in spring at peak biomass when differences are smaller. In mixed farming systems, a significant portion of the spring biomass is under-utilised, while nutrient shortages are greatest in summer and autumn (Moore et al. 2009). For species that are not conserved or intensively rotationally grazed, there is a case for more intensive scrutiny of DMD and CP decline through reproduction and senescence. This study highlighted the opportunity for bladder clover as a standing dry pasture for finishing late lambs in early summer. This strategy needs to consider the fate of the seeds that are consumed if a viable seedbank for regeneration is to be maintained. In this case, ~40% of bladder clover seeds that are eaten by sheep remain viable (H. Norman, unpubl. data) and this means that the sheep would not extract all of the energy that is on offer. However, the gap in NV that would need to be bridged by the use of supplements such as grain would be substantially less for lambs consuming bladder clover residue than for those consuming the residues of many other legume species evaluated.
While FV has generally been neglected as a selection criterion, bladder clover and several other species that have been commercialised in the past 15 years have quality traits that improve livestock profitability. Hackney et al. (2021) compared 19 legume cultivars and found that total herbage production, and its timing and quality differed significantly among species. As in the example presented above, these differences were not predicted by rate of maturity (days to flowering). Thomas et al. (2021) calibrated models using the data from the legumes presented in Fig. 3. They found that pastures with higher NV led to a reduced requirement for supplementary feeding of sheep. Importantly, variation in NV had a greater effect on supplementary feeding costs and gross margins than did differences in biomass production among the annual legume species (Thomas et al. 2021). Differences in the levels of supplementary feeding required in systems with the various legumes was most pronounced during the period from May to July (late autumn to winter). Monjardino et al. (2022) used farming-system models to quantify the relative profitability of subterranean clover, burr medic (Medicago polymorpha L.), bladder clover, French serradella, and biserrula for the medium-rainfall MFZ of Western Australia. The modelling indicated that bladder clover and biserrula respectively offer a 27% and 37% improvement in profitability when compared with a subterranean clover-based pasture system. In contrast, McGrath et al. (2021) compared lambs grazing lucerne, subclover and biserrula, and found that lamb performance was lower than predicted from the nutritional value of the biomass. They hypothesised that this was associated with low levels of photosensitisation or fermentation issues associated with biserrula. Additional feed at the start of the season and a slower loss of NV at the end of the season were important profit drivers for the livestock enterprise (Monjardino et al. 2022). Economic benefits could be achieved through greater utilisation of land under pastures and a higher whole-farm stocking rates, while still maintaining a significant cropping program. Predicted benefits were small at low whole-farm stocking rates. Pastures with higher nutritional value could lead to reduced net emissions per dry sheep equivalent if they allow ruminants to reach finishing weight faster or improve reproductive efficiency (Monjardino et al. 2022).
Case study 3: variability in digestibility within lucerne breeding lines
Lucerne is the most widely sown perennial legume in the MFZ (Bathgate and Pannell 2002). Productivity and persistence of lucerne in dryland systems is challenged by drought during summer, low moisture-storage capability of coarse-textured soils, poor tolerance of soil acidity, intolerance of waterlogging and poor grazing tolerance (Cocks 2001; Humphries and Auricht 2001). Lucerne is considered the ‘queen of forages’, with roughly 10 000 years of coevolution with humans and ruminants. Despite the commercialisation of 77 cultivars in Australia (Nichols et al. 2012; IP Australia 2022), lucerne remains a priority for plant breeders. Breeding objectives have included tolerance of acidic soils, winter activity, grazing tolerance, stem thickness or leaf to stem ratios and aphid tolerance.
A 2011 study compared nutritive traits of 35 commercial and experimental accessions of lucerne, grown in replicated blocks in Adelaide (Norman et al. 2013). The plots were sampled during the vegetative growth phase, 4 weeks after the first biomass cut, and analysed for nutritional traits. There were no significant differences in biomass production among the accessions (unfortunate as early vigour is often a breeding objective); however, there were significant differences in DMD, CP and fibre content (Table 2). The variation in DMD (64–72%) was biologically and economically significant, given similar levels of biomass production. CP ranged from 18% to 23%, and all accessions met the estimated CP requirements of reproducing ewes and growing lambs. Lucerne is generally offered to the class of stock with the highest nutritional requirements. A pregnant Merino ewe (Day 100 of gestation) offered the accession with the highest DMD is predicted to eat 1.2 kg DM/day and grow at a rate of 30 g/day liveweight (GrazFeed™ model; Freer et al. 1997). In contrast, the same ewe eating the accession with the lowest DMD would eat 0.97 kg DM/day and lose 16 g/day. For mature, non-reproducing sheep, the difference in weight gain would be three-fold (18 g gain/day on the lowest-quality lucerne and 63 g/day for the highest-quality lucerne). There is clearly a role for measuring nutritional value in lucerne development.
| Item | DMD (%) | CP (%DM) | NDF (%DM) | ADF (%DM) | DM (g/quadrat) |
|---|---|---|---|---|---|
| Mean | 69.5 | 22.7 | 31.6 | 24.4 | 114 |
| Lowest | 64.2 | 17.4 | 26.9 | 21.1 | 74 |
| Highest | 72.5 | 23.8 | 40.2 | 31.0 | 145 |
| P-value | <0.001 | <0.001 | <0.001 | <0.001 | >0.05 |
Case study 4: variability in nutritional value of a ‘wild’ species of oldman saltbush and the opportunity to use sheep for rapid screening in forage improvement
Case study 4 demonstrated a different approach, one where livestock were included from the beginning of a domestication program. Oldman saltbush (Atriplex nummularia L.), an Australian woody shrub, has become a key species for rehabilitation and use of saline land and other soils that are unprofitable to crop in the MFZ. Oldman saltbush occurs as a dominant species in widespread communities over a 4000 km2 range in arid and semi-arid zones of Australia (Parr-Smith 1982). Because it is adapted to an arid climate, it persists comfortably in the MFZ where rainfall is higher and more regular. The majority of oldman saltbush stands in Australia were generated from seed collected from wild populations. FV of oldman saltbush is constrained by low to moderate biomass production, low to moderate digestibility of organic matter, excessive salt and/or sulfur accumulation and plant secondary compounds such as oxalates (Norman et al. 2004, 2010; Masters et al. 2007; Al Daini et al. 2013). Despite these limitations, saltbush is a valuable source of CP, sulfur, vitamin E and some essential minerals (Ben Salem et al. 2010; Pearce et al. 2010; Fancote et al. 2013; Norman et al. 2019). A collaborative research project, involving farmers, agronomists, livestock scientists and system modellers, identified low FV, particularly ME content, as the key constraint that limited profitability of saltbush systems (Thomas et al. 2009). Pre-experimental modelling indicated that improving shrub organic-matter digestibility by 10% would increase profits by three times the increment associated with increasing biomass production by 10%, or reducing the cost of establishment by 10% (O’Connell et al. 2006). In addition, farmers wanted to improve the relative palatability of the shrub, to reduce overgrazing of the understorey with associated soil erosian risks.
A collection of oldman saltbush germplasm was initiated in 2006, with 60 000 seeds from two subspecies collected from 27 sites (provenances) across Australia (Hobbs and Bennell 2008). The collection was grown across three nursery sites in Condobolin (New South Wales), Tammin (Western Australia) and Monarto (South Australia). Each plantation contained approximately 20 000 individuals that were randomly allocated into groups of four half-sib plants (plants originating from seeds collected from the same shrub) within eight replicate blocks. Survival and growth were assessed prior to grazing with flocks of 1-year-old sheep at stocking densities of 25 and 50 sheep/ha. Sheep grazing pressure allowed for subsequent measurement of recovery from grazing (a trait associated with long-term biomass production). Approximately mid-way through grazing, individual shrubs were ranked for defoliation by using a 1–5 scale where 0 is undefoliated, 1 is 20% defoliated, 2 is 40% defoliated, 3 is 60% defoliated, 4 is 80% defoliated and 5 is 100% defoliated. Across the three experimental sites, sheep demonstrated a consistent and statistically significant preference for specific provenances (Fig. 4a–c). Relative palatability data were therefore collected inexpensively, with only a single additional visual assessment at each site.
(a–c) Regression of sheep preferences (as expressed through the mean percentage defoliation score measured mid-way through grazing) among provenances of oldman saltbush grown in three locations. (d) The relationship between mean organic-matter digestibility of leaves and mean defoliation score among provenances at the Monarto site. For each linear regression, the coefficient of determination (R2) was significant (P < 0.01).
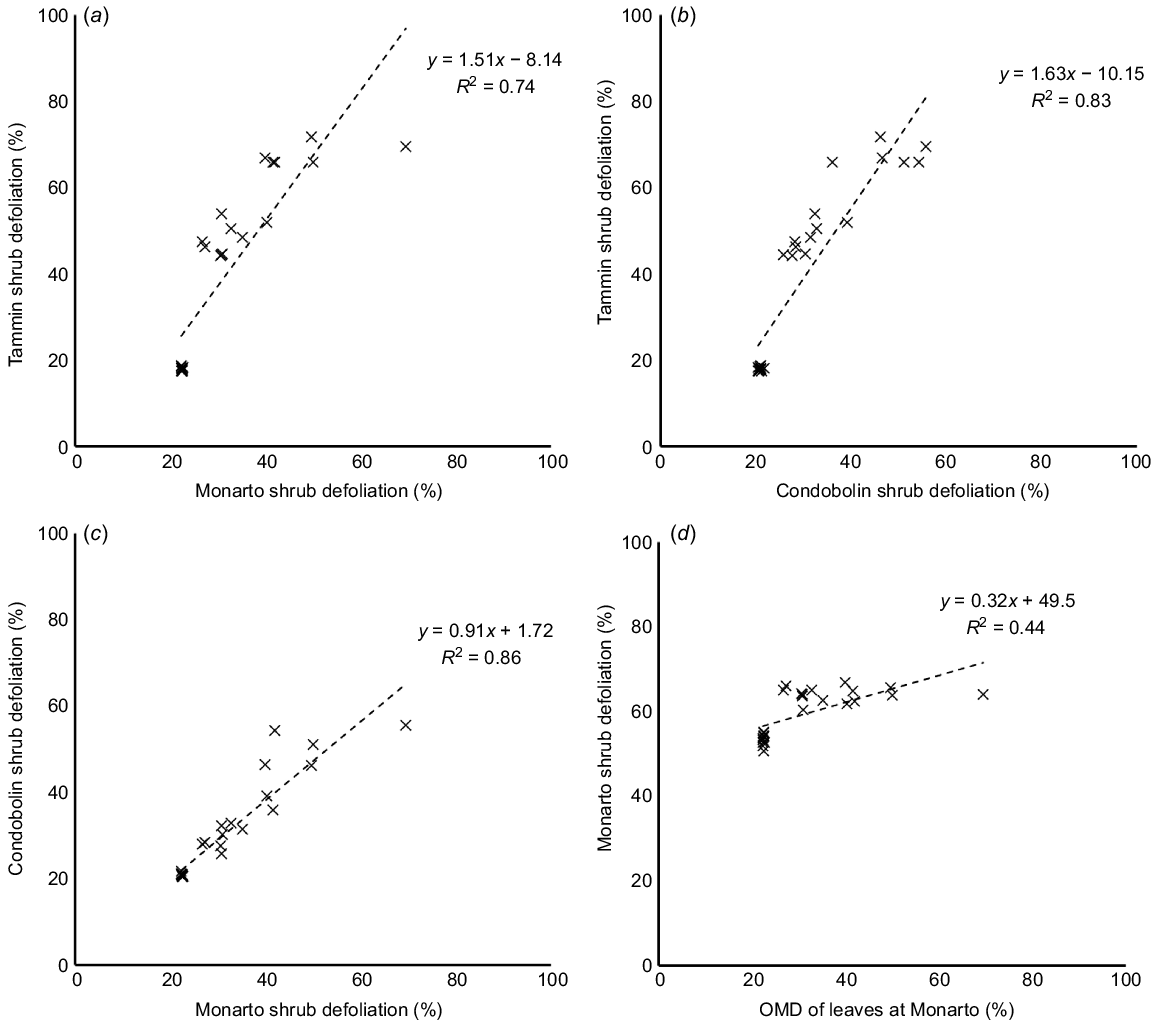
Leaves of the shrubs at the Monarto site (n = 20 shrubs per provenance) were subsampled and analysed for NV, by using laboratory and NIRS methods developed and calibrated for Australian shrubs (Norman and Masters 2010; Norman et al. 2010). Organic matter digestibility (OMD) rather than DMD was used to compare energy potential among plants and provenances, to account to the high and variable salt content among plants (Masters 2015). There was significant variation in the leaves among provenances in OMD, salt, CP, minerals and biomass growth. There was a positive and statistically significant correlation between OMD and preference, with OMD accounting for 44% of the variation in preference (Fig. 4d). The first implication of this finding is that use of sheep to indicate preference offered an opportunity to ‘short-cut’ the task of sampling and analysing many individual plants for OMD; something that is time consuming and expensive in terms of human labour resources in a plant breeding project. The second implication is that if the team had focused on biomass as the primary selection tool while narrowing the germplasm, it is likely they would have forgone gains in energy value, which is the key trait driving profitability.
After consideration of palatability, biomass and NV, 90 of the 60 000 shrubs (the best 30 from each nursery site) were selected and vegetatively cloned as cuttings. When this cohort was compared with the average of the original collection, they had 20% higher OMD, were consistently preferred by sheep and produced eight times more biomass than the mean of the original collection. These cuttings were planted into replicated blocks at three experimental sites across Australia and assessed for agronomic and nutritional traits over 3 years. Again, sheep were used to rank the genotypes for relative palatability, and this was included with agronomic traits as a selection criterion. In the final phase of the work, the best 12 genotypes were identified, vegetatively cloned and planted in replicated block experiments at 13 diverse sites across southern Australia. Genotypes were again assessed for survival, productivity and nutritional value. The most promising four genotypes and an industry standard were harvested and fed to sheep in a final energy and nitrogen metabolism experiment.
This led to commercialisation of Anameka™ (a clonal cultivar) in 2015, just 9 years after the establishment of the collection. Whole-farm economic analysis suggests that Anameka™ can double the profitability of saltbush plantations on farms and reduce risk as the plants are resilient to drought. While this provides a good example of using the ‘animal consumer’ to assist in plant domestication, the multidisciplinary team, working with farmers, enabled other benefits to be described. If improved profitability of Anameka™ systems results in greater rates of adoption, the environmental health of agricultural landscapes in southern Australia can also be improved. Being both perennial and in active growth through the summer and autumn period, saltbush systems have been shown to reduce leakage of rainwater to water tables, thus potentially reducing the effects of dryland salinity (Barrett-Lennard et al. 2005; Bennett et al. 2012). This is an example of how integrating livestock production in farming systems can lead to better environmental outcomes (Masters et al. 2010).
In summary, 9 years is a very short time for domestication and commercialisation of a dioecious, octoploid and woody perennial plant that can have a lifespan of several decades. What contributed to the success of the Anameka™ project were the efforts of a multi-disciplinary team including farmers, tree nursery proprietors and whole farm-system modelling to determine a clear goal, development of methods for accurate and rapid prediction of NV, incorporation of animal nutritional and preference priorities and a well structured on-farm field-testing program. More than 6 million Anameka have now been planted by farmers across southern Australia.
Conclusions
The purpose of this review has been to examine the process of pasture plant breeding and development from a livestock perspective. While we have focused on sheep production examples, the same principles apply to forages that are developed for cattle industries. The case studies that we presented demonstrated that the selection of pasture species has often been based on an assumption that green forage has a consistent FV and NV across plant species and genotypes within species. Priorities have therefore been primarily agronomic, or crop-focused (e.g. nitrogen fixation), with biomass production often the only livestock consideration. This in part has been enabled by incongruent priorities of the livestock and grains sectors. It may also be an artifact of previous flock structures, where carrying larger numbers of wethers, at slower growth rates, may have been a goal in wool-focused systems. It is clear from the information presented that there are very significant differences in NV within and among pasture species and that this often extends into the period of senescence. Livestock offered similar amounts of feed from different pasture species may either gain or lose liveweight. Reducing candidate species from thousands to tens (or less) before livestock, the ultimate consumers, greatly narrows the potential benefits of forage improvement programs.
There is a belief among plant breeders that that measurement of nutritional value or relative palatability is expensive when compared with traditional genotype × environment agronomic experiments. This is not true for several reasons. The first is that many studies have found that DMD is a trait that is relatively stable across environments (at similar maturity stages), compared with biomass or even CP content (Shenk and Westerhaus 1982; Carpenter and Casler 1990; Cherney and Volenec 1992). Comparative measurement at just one or two sites over the lifecycle of the plant should be sufficient to identify material with higher energy values. Measurement costs have been reduced through the development of multi-species NIRS calibrations that allow rapid, inexpensive and accurate prediction of DMD, fibre profiles, CP and ash (Norman et al. 2020). These have been expanded further to include additional traits, plant secondary compounds and key minerals (H. Norman and R. Meyer, pers. comm.). Unfortunately, use of NIRS for plants in the field (avoiding the drying and grinding steps), is unlikely to provide a useful tool due to significant prediction errors associated with ambient light, moisture and sample heterogeneity. Most plant improvement activities invest considerable resources in collecting, drying and weighing quadrat cuts to compare dry matter production; sending this material for NIRS analysis represents a relatively minor cost compared with the total expense and effort. Equally, grazing research sites to assess livestock preferences and plant recovery from grazing can be managed with low additional costs. The challenge is to foster active, multidisciplinary collaborations among plant breeders, agronomists, farmers and livestock scientists.
The examples of potential benefits in these case studies have been narrowly focused on feed intake and feed utilisation. Ultimately, new forage genotypes must be viewed in a systems context. A feedbase with more digestible biomass for more of the year offers options to increase stocking rate, reduce supplementary feed costs, increase productivity and more flexibility in stock turnoff. Involvement of farmers, one of the other ‘consumers’ of the product, is critical to success. Witcombe et al. (2006) reviewed farmer collaboration in international public-sector breeding programs. They concluded that participatory techniques can be used to achieve high client orientation and meet client needs more effectively. Further, the lack of explicit effort in many international public-sector breeding programs is a real disadvantage as evidenced by farmers continuing to grow landraces or obsolete varieties because new, recommended varieties do not adequately meet their requirements (Witcombe et al. 1998).
Emerging consumer interest in the role of livestock-derived foods to provide a variety of essential nutrients and other health-promoting compounds must also be considered (https://www.dublin-declaration.org/; Leroy et al. 2023). Forage scientists have a role to ensure that forages provide nutrients that are essential to both livestock and human health, and that production systems are ethical and environmentally sustainable.
A new approach
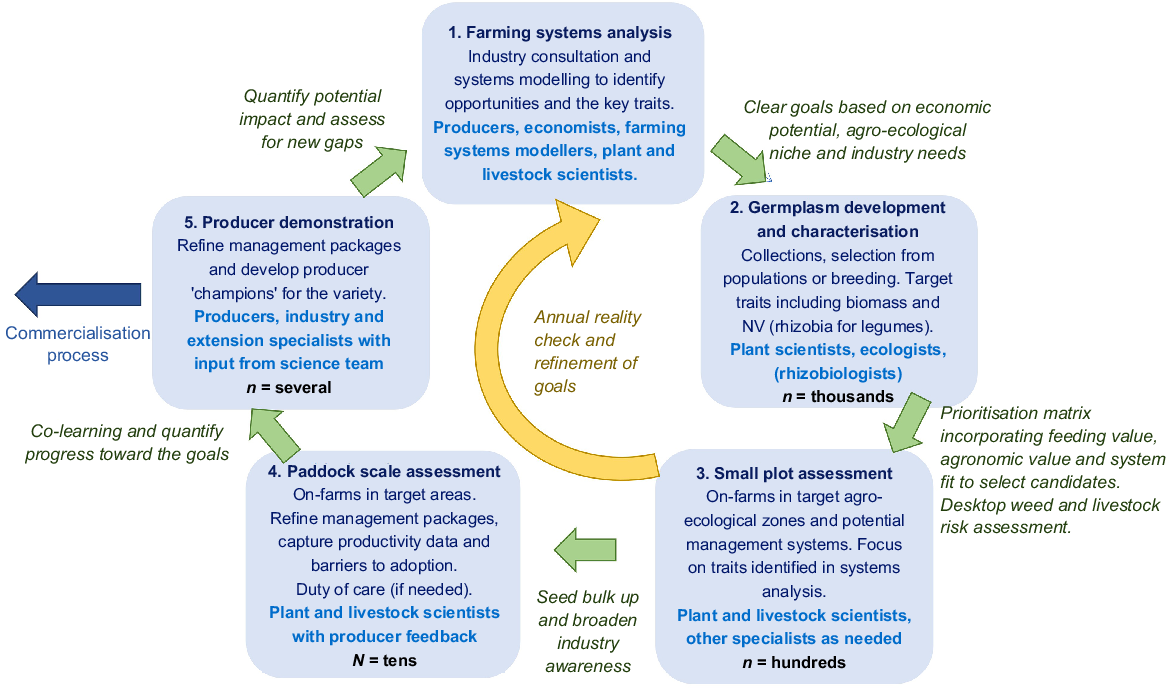
The inverted-triangle approach to pasture and forage improvement in Australia (as described by Nichols et al. 2010; Ewing 2017; Badgery et al. 2023) uses a very narrow focus to reduce the pool of candidate genotypes. This relies on many assumptions regarding the needs of the client, livestock requirements and the fit of the new variety within a broader farming system. A different plant improvement model is advocated with greater farmer consultation, whole farm-system modelling, NV measurement of a broad range of candidate species/genotypes and, where possible, nutritional preference or grazing livestock are incorporated early in the selection process (Fig. 5). The first step is to identify and prioritise opportunities or gaps within farming systems and this should involve system modellers, economists, plant and livestock scientists. The outcome of this step needs to be road-tested with farmers and consultants to achieve some understanding of potential scale, possible barriers to adoption and other benefits that may be achieved. With a clear understanding of the opportunity or gap, a wide range of germplasm is assembled and rapidly screened in the second step, preferably in the target environment. Traits to assess include biomass production, NV at key plant growth stages (through rapid and inexpensive NIRS methods), persistence and/or seed production and anything else that was deemed important in the first stage and is easy to quantify (e.g. phenotypic markers, spiny pods, seed retention etc.). For some species, this may be the stage to utilise livestock preferences to identify elite material. Germplasm can be mapped onto a selection matrix that describes relative agronomic value, relative feeding value and relative climatic and edaphic adaptation. This matrix, combined with desktop assessments of secondary compounds (positive and negative), potential weed risk and (if appropriate) rhizobial needs, is then used to narrow the pool of candidate species and/or genotypes. The smaller pool of candidates can then be tested in replicated small-plot experiments across the target zones. Herbicide-tolerance experiments are conducted. An active feedback loop to the first stage, using data for model sensitivity analysis, allows for information that is collected in the initial stages to be quickly re-assessed in a systems context. Using this approach, assumptions regarding the plant selection goal can be tested and novel opportunities that were not part of the original goal can be explored. Seed bulk-up enables the fourth stage of small paddock assessment on farms. Management packages are developed through agronomic experiments, and grazing studies conducted. Duty of care experiments may be required if the species is new to agriculture. The impact of the plant on meat eating quality and nutrient profile should be considered. The final stage before commercialisation is paddock-scale demonstration with leading farmers to further refine management packages and to generate ‘producer champions’ for the product. Data and farmer feedback are collected to quantify the benefits, support extension activities and re-assess the need for further plant improvement. Where forage plants are developed for mixed farming systems, it is essential that traits that affect both livestock and crop systems are measured simultaneously.
Data availability
The data that support these case studies cannot be publicly shared due to ethical or privacy reasons but may be shared upon reasonable request to the corresponding author if appropriate.
Declaration of funding
The oldman saltbush improvement research was funded by the CRC Future Farm Industries. We also acknowledge support (at various stages) by Meat and Livestock Australia, Australian Wool Innovation, Australian Department of Agriculture and Fisheries, South Australian Government and a large cohort of regional farmer and land management groups. The legume examples were drawn from project number: 01200.042; B.CCH.6540, a collaboration between the University of Western Australia (UWA), South Australian Research and Development Institute (SARDI) and CSIRO. The work was funded through the Australian Department of Agriculture and Water Resources ‘Filling the Research Gap’ program and managed as part of the National Livestock Methane Program by Meat and Livestock Australia.
Acknowledgements
HN thanks the late (and sadly missed) Dr Sarah Blumer, and the organising committee of the 33rd Australian Association of Animal Sciences Conference for the honour of presenting the 2021 Harry Stobbs Memorial lecture. Thanks go to Matt Wilmot, Belinda Hackney, John Howieson and Elizabeth Hulm for providing valuable constructive feedback on the manuscript. The saltbush and legume research described has involved many partners from CSIRO, New South Wales Department of Primary Industries, Murdoch University, WA Department of Primary Industries and Regional Development, UWA, Western Australian Department of Parks and Wildlife, Department of Water Land and Biodiversity Conservation in South Australia and SARDI. Contributors to the projects described in the case studies include Matt Wilmot, Elizabeth Hulm, Ed Barrett-Lennard, Mike Bennell, Belinda Hackney, Robert Harrison, Joshua Hendry, Trevor Hobbs, John Howieson, Steve Hughes, Alan Humphries, Peter Jessop, Angelo Loi, Richard Mazanec, David McKenna, Ram Nair, Bradley Nutt, Wayne O’Sullivan, David Peck, John Piltz, Dean Revell, Alan Rintoul, Trevor Rowe, Greg Sweeney, Dean Thomas, Andrew Toovey, Phillip Vercoe, Ron Yates and Paul Young. We are very grateful to Dustin and Lisa McCreery from Chatfield’s Tree Nursery and to our many host farmers for research input and support. This includes the York family (Tammin), the Butcher family (Brookton), Tighe family (Clackline and Kellerberrin), and the Walsh and Lehmann families from Cranbrook.
References
Al Daini H, Norman HC, Young P, Barrett Lennard EG (2013) The source of nitrogen (NH4+ or NO3) affects the concentration of oxalate in the shoots and the growth of Atriplex nummularia (oldman saltbush). Functional Plant Biology 40, 1057-1064.
| Crossref | Google Scholar |
Badgery W, Li G, Simmons A, Wood J, Smith R, Peck D, Ingram L, Durmic Z, Annette Cowie A, Humphries A, Hutton P, Winslow E, Vercoe P, Eckard R (2023) Reducing enteric methane of ruminants in Australian grazing systems – a review of the role for temperate legumes and herbs. Crop & Pasture Science
| Crossref | Google Scholar |
Barrett-Lennard EG, George RJ, Hamilton G, Norman HC, Masters DG (2005) Multi-disciplinary approaches suggest profitable and sustainable farming systems for valley floors at risk of salinity. Australian Journal of Experimental Agriculture 45, 1415-1424.
| Crossref | Google Scholar |
Bathgate A, Pannell DJ (2002) Economics of deep-rooted perennials in Western Australia. Agricultural Water Management 53, 117-132.
| Crossref | Google Scholar |
Bathgate A, Revell C, Kingwell R (2009) Identifying the value of pasture improvement using wholefarm modelling. Agricultural Systems 102, 48-57.
| Crossref | Google Scholar |
Bell LW, Moore AD (2012) Integrated crop–livestock systems in Australian agriculture: trends, drivers and implications. Agricultural Systems 111, 1-12.
| Crossref | Google Scholar |
Bell LW, Moore GA, Ewing MA, Ryan MH (2006) Production, survival and nutritive value of the perennial legumes Dorycnium hirsutum and D. rectum subjected to different cutting heights. Grass & Forage Science 61, 60-70.
| Crossref | Google Scholar |
Ben Salem H, Norman HC, Nefzaoui A, Mayberry DE, Pearce KL, Revell DK (2010) Potential use of oldman saltbush (Atriplex nummularia Lindl.) in sheep and goat feeding. Small Ruminant Research 91, 13-28.
| Crossref | Google Scholar |
Carpenter JA, Casler MD (1990) Divergent phenotypic selection response in smooth bromegrass for forage yield and nutritive-value. Crop Science 30, 17-22.
| Crossref | Google Scholar |
Chen C, Fletcher A, Ota N, Oliver Y, Lawes R (2023) Integrating long fallow into wheat-based cropping systems in Western Australia: spatial pattern of yield and economic responses. Agricultural Systems 204, 103561.
| Crossref | Google Scholar |
Cherney JH, Volenec JJ (1992) Forage evaluation as influenced by environmental replication: a review. Crop Science 32, 841-846.
| Crossref | Google Scholar |
Clarivate (2022) Web of ScienceTM. Available at https://www.webofscience.com/wos/woscc/basic-search
Cocks PS (2001) Ecology of herbaceous perennial legumes: a review of characteristics that may provide management options for the control of salinity and waterlogging in dryland cropping systems. Australian Journal of Agricultural Research 52, 137-151.
| Crossref | Google Scholar |
Davies SR, Lane PA (2003) Seasonal changes in feed quality of Dorycnium spp. In ‘The 11th Australian Agronomy Conference’, Geelong, Victoria. (Eds M Unkovich, GJ O’Leary) (Australian Society of Agronomy: Horsham, Vic., Australia). Available at http://www.regional.org.au/au/asa/2003
Edwards T, Howieson J, Nutt B, Yates R, O’Hara G, Van Wyk BE (2019) A ley-farming system for marginal lands based upon a self-regenerating perennial pasture legume. Agronomy for Sustainable Development 39, 13.
| Crossref | Google Scholar |
Elgersma A, Søegaard K (2018) Changes in nutritive value and herbage yield during extended growth intervals in grass–legume mixtures: effects of species, maturity at harvest, and relationships between productivity and components of feed quality. Grass and Forage Science 73, 78-93.
| Crossref | Google Scholar |
Fancote CR, Vercoe PE, Pearce KL, Williams IH, Norman HC (2013) Backgrounding lambs on saltbush provides an effective source of Vitamin E that can prevent Vitamin E deficiency and reduce the incidence of subclinical nutritional myopathy during summer and autumn. Animal Production Science 53, 247-255.
| Crossref | Google Scholar |
Fortune JA, Cocks PS, McFarlane CK, Smith FP (1995) Distribution and abundance of annual legume seeds in the wheatbelt of Western Australia. Australian Journal of Experimental Agriculture 35, 189-197.
| Crossref | Google Scholar |
Freer M, Moore AD, Donnelly JR (1997) GRAZPLAN: Decision support systems for Australian grazing enterprises: II. The animal biology model for feed intake, production and reproduction and the GrazFeed DSS. Agricultural Systems 54, 77-126.
| Crossref | Google Scholar |
Friend M, Robertson S, Kennedy A, Thomas D (2018) Developing a mixed farming systems RD&A program. Final report to Meat and Livestock Australia. Appendix 2. p. 1117. Available at https://www.mla.com.au/download/finalreports?itemId=3650
Gildersleeve RR, Smith GR, Pemberton IJ, Gilbert CL (1991) Detection of isoflavones in seedling subterranean clover. Crop Science 31, 889-892.
| Crossref | Google Scholar |
Hackney B, Edward C, Piltz J, Friend M, Howieson J (2017) Producer perceptions of feedbase limitations to livestock production and requirements for amelioration. In ‘Proceedings of the 18th Australian Society of Agronomy Conference’, 24–28 September 2017, Ballarat, Vic., Australia. (Australian Society of Agronomy Inc.)
Hackney BF, Jenkins J, Powells J, Edwards CE, De Meyer S, Howieson JG, Yates RJ, Orgill SE (2019) Soil acidity and nutrient deficiency cause poor legume nodulation in the permanent pasture and mixed farming zones of south-eastern Australia. Crop & Pasture Science 70, 1128-1140.
| Crossref | Google Scholar |
Hackney B, Rodham C, Dyce G, Piltz J (2021) Pasture legumes differ in herbage production and quality throughout spring, impacting their potential role in fodder conservation and animal production. Grass and Forage Science 76, 116-133.
| Crossref | Google Scholar |
Hobbs TJ, Bennell M (2008) Agroforestry species profiles for lower rainfall regions of southeastern Australia FloraSearch. 1b. Report to the Joint Venture Agroforestry Program (JVAP) and Future Farm Industries CRC. Published by RIRDC, Canberra, ACT, Australia. Available at https://rirdc.infoservices.com.au/downloads/07-080
Howieson JG, O’Hara GW, Carr SJ (2000) Changing roles for legumes in Mediterranean agriculture: developments from an Australian perspective. Field Crops Research 65, 107-122.
| Crossref | Google Scholar |
Humphries AW, Auricht GC (2001) Breeding lucerne for Australia’s southern dryland cropping environments. Australian Journal of Agricultural Research 52, 153-169.
| Crossref | Google Scholar |
IP Australia (2022) ‘Plant breeders rights database.’ (IP Australia, Australian Government: Canberra, ACT, Australia) Available at www.ipaustralia.gov.au/get-theright-ip/plant-breeders-rights
Kelman WM, Blumenthal MJ, O’Connor JW, Borchard PA (2007) Lotus uliginosus (greater lotus) breeding line LUX97: a high seed producing, low condensed tannin population. Australian Journal of Experimental Agriculture 47, 116-118.
| Crossref | Google Scholar |
Lawes R, Mata G, Richetti J, Fletcher A, Herrmann C (2022) Using remote sensing, process-based crop models, and machine learning to evaluate crop rotations across 20 million hectares in Western Australia. Agronomy for Sustainable Development 42, 120.
| Crossref | Google Scholar |
Leroy F, Smith N, Adesogan AT, Beal T, Iannotti L, Moughan PJ, Mann N (2023) The role of meat in the human diet: evolutionary aspects and nutritional value. Animal Frontiers 13(2), 11-18.
| Crossref | Google Scholar |
Loi A, Howieson JG, Nutt BJ, Carr SJ (2005) A second generation of annual pasture legumes and their potential for inclusion in Mediterranean-type farming systems. Australian Journal of Experimental Agriculture 45, 289-299.
| Crossref | Google Scholar |
Loi A, Franca A, Nutt BJ, Yates RJ, D’Antuono MF, Howieson JG (2015) Important ecological traits for selecting Biserrula pelecinus L. (biserrula) genotypes for their potential introduction into agricultural systems. Grass and Forage Science 70, 519-529.
| Crossref | Google Scholar |
Masters DG, Norman HC, Barrett-Lennard EG (2005) Agricultural systems for saline soil: The potential role of livestock. Asian-Australasian Journal of Animal Sciences 18, 296-300.
| Google Scholar |
Masters DG, Mata G, Revell CK, Davidson RH, Norman HC, Nutt BJ, Solah V (2006) The effects of Prima gland clover (Trifolium glanduliferum Boiss cv Prima) consumption on sheep production and meat quality. Australian Journal of Experimental Agriculture 46, 291-297.
| Crossref | Google Scholar |
Masters DG, Benes SE, Norman HC (2007) Biosaline agriculture for forage and livestock production. Agriculture, Ecosystems & Environment 119, 234-248.
| Crossref | Google Scholar |
Masters DG, Norman HC, Thomas DT (2019) Minerals in pastures: are we meeting the needs of livestock? Crop & Pasture Science 70, 1184-1195.
| Crossref | Google Scholar |
Masters DG, Blache D, Lockwood AL, Maloney SK, Norman HC, Refshauge G, Hancock SN (2023) Shelter and shade for grazing sheep: implications for animal welfare and production and for landscape health. Animal Production Science 63, 623-644.
| Crossref | Google Scholar |
McGrath SR, Sandral GA, Sundermann L, Quinn JC, Weston LA, Friend MA (2021) Liveweight and carcass characteristics of White Dorper and Crossbred lambs grazing lucerne, subterranean clover, biserrula or a choice of subterranean clover plus biserrula in southern Australia. Animal Production Science 61, 1151-1159.
| Crossref | Google Scholar |
Meat and Livestock Australia (2019) Industry projections 2019: Australian sheep. Available at https://www.mla.com.au/globalassets/mla-corporate/prices--markets/documents/trends--analysis/sheep-projections/mla_australian-sheep-industry-projections-2019.pdf [Accessed 22 September 2022]
Meat and Livestock Australia (2023) Market information statistics database. Available at https://www.mla.com.au/prices-markets/Trends-analysis/mla--awi-sheepmeat-survey/ [Accessed 28 March 2023]
Monjardino M, Bathgate A, Llewellyn R (2014) Opportunities for plant improvement to increase the value of forage shrubs on low-rainfall mixed farms. Crop & Pasture Science 65, 1057-1067.
| Crossref | Google Scholar |
Monjardino M, Loi A, Thomas DT, Revell CK, Flohr BM, Llewellyn RS, Norman HC (2022) Improved legume pastures increase economic value, resilience and sustainability of crop-livestock systems. Agricultural Systems 203, 103519.
| Crossref | Google Scholar |
Moore AD (2014) The case for and against perennial forages in the Australian sheep–wheat zone: modelling livestock production, business risk and environmental interactions. Animal Production Science 54, 2029-2041.
| Crossref | Google Scholar |
Moore AD, Bell LW, Revell DK (2009) Feed gaps in mixed-farming systems: insights from the Grain & Graze program. Animal Production Science 49(10), 736-748.
| Crossref | Google Scholar |
Nichols PGH, Revell CK, Humphries AW, Howie JH, Hall EJ, Sandral GA, Ghamkhar K, Harris CA (2012) Temperate pasture legumes in Australia: their history, current use, and future prospects. Crop & Pasture Science 63, 691-725.
| Crossref | Google Scholar |
Norman HC, Freind C, Masters DG, Rintoul AJ, Dynes RA, Williams IH (2004) Variation within and between two saltbush species in plant composition and subsequent selection by sheep. Australian Journal of Agricultural Research 55, 999-1007.
| Crossref | Google Scholar |
Norman HC, Masters DG, Rintoul AJ, Wilmott MG, Jayasena V, Loi A, Revell CK (2005) The relative feeding value of a new pasture legume, eastern star clover (Trifolium dasyurum) compared to subterranean clover (Trifolium subterraneum). Australian Journal of Agricultural Research 56, 637-644.
| Crossref | Google Scholar |
Norman HC, Revell DK, Mayberry DE, Rintoul AJ, Wilmot MG, Masters DG (2010) Comparison of in vivo organic matter digestibility of native Australian shrubs to in vitro and in sacco predictions. Small Ruminant Research 91, 69-80.
| Crossref | Google Scholar |
Norman HC, Hughes SJ, Hulm E, Humphries AW, Oldach K, Revell DK, Durmic Z, Vadhanabhuti J, Vercoe PE (2013) Improving the feeding value of dryland lucerne in Australia. In ‘Proceedings of the 22nd International Grasslands Congress’, 14–19 September 2013, Sydney, NSW, Australia. (New South Wales Department of Primary Industry)
Norman HC, Duncan EG, Masters DG (2019) Halophytic shrubs accumulate minerals associated with antioxidant pathways. Grass and Forage Science 74, 345-355.
| Crossref | Google Scholar |
Norman HC, Hulm E, Humphries AW, Hughes SJ, Vercoe PE (2020) Broad near-infrared spectroscopy calibrations can predict the nutritional value of >100 forage species within the Australian feedbase. Animal Production Science 60, 1111-1122.
| Crossref | Google Scholar |
Norman HC, Humphries AW, Hulm E, Young P, Hughes SJ, Rowe T, Peck DM, Vercoe PE (2021) Productivity and nutritional value of 20 species of perennial legumes in a low-rainfall Mediterranean-type environment in southern Australia. Grass and Forage Science 76, 134-158.
| Crossref | Google Scholar |
Nutt BJ, Loi A, Hackney B, Yates RJ, D’Antuono M, Harrison RJ, Howieson JG (2021) ‘Summer sowing’: a successful innovation to increase the adoption of key species of annual forage legumes for agriculture in Mediterranean and temperate environments. Grass and Forage Science 76, 93-104.
| Crossref | Google Scholar |
O’Connell M, Young J, Kingwell R (2006) The economic value of saltland pastures in a mixed farming system in Western Australia. Agricultural Systems 89, 371-389.
| Crossref | Google Scholar |
Pearce KL, Norman HC, Hopkins DL (2010) The role of saltbush-based pasture systems for the production of high quality sheep and goat meat. Small Ruminant Research 91, 29-38.
| Crossref | Google Scholar |
Rasmussen S, Parsons AJ, Xue H, Liu Q, Jones CS, Ryan GD, Newman JA (2014) Transcript profiling of fructan biosynthetic pathway genes reveals association of a specific fructosyltransferase isoform with the high sugar trait in Lolium perenne. Journal of Plant Physiology 171, 475-485.
| Crossref | Google Scholar |
Reed KFM (1994) Improved grass cultivars increase milk and meat production: a review. New Zealand Journal of Agricultural Research 37, 277-286.
| Crossref | Google Scholar |
Reed KFM (2014) Perennial pasture grasses: an historical review of their introduction, use and development for southern Australia. Crop & Pasture Science 65, 691-712.
| Crossref | Google Scholar |
Ridley AM, Pannell DJ (2005) The role of plants and plant-based research and development in managing dryland salinity in Australia. Australian Journal of Experimental Agriculture 45, 1341-1355.
| Crossref | Google Scholar |
Shenk JS, Westerhaus MO (1982) Selection for yield and quality in orchardgrass. Crop Science 22, 422-425.
| Crossref | Google Scholar |
Smith KF, Ludemann C, Lewis CD, Malcolm B, Banks RG, Jacobs JL, Fennessy PF, Spangenberg GC (2014) Estimating the value of genetic gain in perennial pastures with emphasis on temperate species. Crop & Pasture Science 65, 1230-1237.
| Crossref | Google Scholar |
Thomas DT, White CL, Hardy J, Collins JP, Ryder A, Norman HC (2009) An on-farm evaluation of the capability of saline land for livestock production in southern Australia. Animal Production Science 49, 79-83.
| Crossref | Google Scholar |
Thomas DT, Flohr BM, Monjardino M, Loi A, Llewellyn RS, Lawes RA, Norman HC (2021) Selecting higher nutritive value annual pasture legumes increases the profitability of sheep production. Agricultural Systems 194, 103272.
| Crossref | Google Scholar |
Wilmot MG, Norman HC, Hendry J, Young P, Hulm E, Harrison R, Toovey A, Spiers J (2023) Sheep grazing Trigonella balansae had productivity, health and meat quality similar to sheep grazing subterranean clover or French serradella. Animal Production Science 63, 152-167.
| Crossref | Google Scholar |
Witcombe JR, Packwood AJ, Raj AGB, Virk DS (1998) The extent and rate of adoption of modern cultivars in India. In ‘Seeds of choice: making the most of new varieties for small farmer’. (Eds JR Witcombe, DS Virk, J Farrington) pp. 53–58. (Oxford IBH: New Delhi, India; and Intermediate Technology Publications: London, UK)
Witcombe JR, Gyawali S, Sunwar S, Sthapit BR, Joshi KD (2006) Participatory plant breeding is better described as highly client-oriented plant breeding. II. Optional farmer collaboration in the segregating generationss. Experimental Agriculture 42(1), 79-90.
| Crossref | Google Scholar |
Young M, Kingwell R, Young J, Vercoe P (2020) An economic analysis of sheep flock structures for mixed enterprise Australian farm businesses. Australian Journal of Agricultural and Resource Economics 64, 677-699.
| Crossref | Google Scholar |
Young M, Vercoe PE, Kingwell RS (2022) Optimal sheep stocking rates for broad-acre farm businesses in Western Australia: a review. Animal Production Science 62, 803-817.
| Crossref | Google Scholar |
Zhao Q, Dixon RA (2014) Altering the cell wall and its impact on plant disease: from forage to bioenergy. Annual Review of Phytopathology 52, 69-91.
| Crossref | Google Scholar |