

A large proportion of genetic variation in cow and ewe body composition is independent of yearling composition
W. S. Pitchford A *
A *
A Davies Livestock Research Centre, School of Animal and Veterinary Sciences, The University of Adelaide, Roseworthy Campus, Roseworthy, SA 5371, Australia.
Animal Production Science 63(11) 957-962 https://doi.org/10.1071/AN22458
Submitted: 14 December 2022 Accepted: 20 February 2023 Published: 14 March 2023
© 2023 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Context: Commercial beef and sheep producers have adopted using condition score for management of cows and ewes to maximise productivity. Significant premiums are being paid for bulls and rams with higher-fat breeding values based on young animal measurements, with the aim of increased resilience in adult female progeny.
Aims: The aim of this study was to quantify the relationship between adult body condition score and young muscle and fat and also the genetic variation in condition that is independent of yearling traits.
Methods: Published genetic parameters for four large data sets from Angus and tropically adapted cattle, and composite and Merino sheep were used to partition genetic variation in adult condition that is associated with, or independent of, young composition measures.
Key results: One genetic standard deviation in young muscle or fat was associated with approximately just 0.1 adult body condition scores. Approximately ¾ of the genetic variation in adult body condition score (BCS) is independent of genetic variation in young weight and composition traits.
Conclusions and implications: Producers need to be careful with premiums paid for bulls and rams with superior yearling composition traits with the aim of changing adult BCS. The best way to achieve this is for seedstock breeders to record and report breeding values for cow and ewe BCS.
Keywords: cattle, condition score, fat, genetic correlation, heritability, mature, muscle, sheep, yearling.
Introduction
At the start of the Beef CRC Maternal Productivity Project, commercial beef breeders raised concerns that genetic selection programs were focusing too heavily on feedlot and carcass traits, with a negative weighting on fat depth to increase lean meat yield, and cows were becoming too lean as a result. There was concern that this would lead to a decline in maternal productivity, especially during times of feed shortage (Lee et al. 2018; Pitchford et al. 2018a). Thus, the question to be addressed in this paper is how much genetic variation is there in cow body composition independent of genetic variation in yearling composition? The focus herein is on genetic and not total phenotypic variation.
This is the same analysis as reported by Pitchford and Lee (2022) but with more details presented and comparisons with other breeds and species to aid general conclusions to be made. It follows mixed graziers asking how general the conclusions are and to what extent should these change bull and ram purchasing decisions.
Materials and methods
Data set 1. Angus cattle
A genetic covariance matrix was formed using 55 covariances reported by Donoghue et al. (2018) and Hickson and Pitchford (2021) from 2641 cows. There were 10 traits, including four yearling heifer traits (weight (WT), eye-muscle area (EMA), P8 rump-fat and rib-fat depth, and seven cow traits measured at the time of weaning their second calf (WT, EMA, P8 fat, rib fat, height (HT) and body condition score (BCS, 1–5)). The values herein differ slightly from those in Pitchford and Lee (2022) as intramuscular fat (IMF) was not included as a young or adult trait herein; hence, the 55 covariances among 10 traits rather than 78 among 12 traits. Phenotypic variances and heritabilities are presented (Table 1).

|
Data set 2. Tropical Composite cattle
The second data set utilised was from Barwick et al. (2009) and Wolcott et al. (2014a, 2014b), comprising 1094 Tropical Composite and 1016 Brahman cows. The same 10 traits were used as for the Angus data. The heritabilities and genetic correlations among yearling (ENDWET) traits came from Barwick et al. (2009), the heritabilities and genetic correlations within mature traits came from Wolcott et al. (2014a) and the correlations between yearling and mature traits came from Wolcott et al. (2014b). While Wolcott et al. (2014a) reported separate variances and heritabilities for Composite and Brahman cows, the genetic correlations were reported from a combined analysis. Given the variances were similar and the conclusions herein are driven primarily by the correlations, only the Tropical Composite variance components were used and presented (Table 1).
Data set 3. Lambpro Maternal Composite sheep
Walkom et al. (2016) reported genetic correlations between WT and composition traits of young (post-weaning) and adult Composite ewes, utilising 4886 records. P8 or rump fat was not measured, so data were available for eight traits only. Heritabilities were quite variable among the data sets (Table 1).
Data set 4. Merino sheep
Walkom and Brown (2017) reported genetic parameters for lamb and adult growth and composition traits for 13 698 ewes. The traits were far more limited than in the other three data sets, with just WT and BCS applicable for adult traits. Lamb traits included WT, eye-muscle depth and fat depth. Walkom and Brown (2017) did not report genetic correlations among the three lamb traits (post-weaning WT, eye-muscle depth and fat depth), so these correlations were taken from Brown and Swan (2016) which was not exactly the same data set, but was substantial (n = 42 769).
For each data set, the covariance matrix was not positive definite and so was bent using nearPD in the Matrix package v1.2-17 within R, originally developed by Jens Oehlschlaegel (https://github.com/joehl), using the algorithm of Higham (2002). Genetic correlations were calculated from the bent covariances and are reported for the four data sets (Table 2).

|
A key piece of information for commercial sheep and cattle producers is the change in adult (A) body composition (ΔA) associated with young (Y) estimated breeding values (EBVs or ASBVs). This was calculated by multiplying the regression coefficient by the genetic standard deviation to standardise the response, as follows:
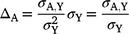
The primary aim of this analysis is to estimate the amount of genetic variation in cow or ewe BCS that is independent of (conditional on) the multiple growth and composition measures recorded on young animals (yearling cattle and post-weaning sheep). The covariance matrix is presented with Y representing the four young (yearling or post-weaning) measures and A representing the six adult measures. The method of calculating conditional adult genetic covariances is as follows:


where Σ11 is the young genetic covariance matrix, Σ22 is the adult genetic covariance and Σ12 is the genetic covariances between young and adult traits. Remaining variances in adult traits have been reported directly (Table 3) and as proportions of original genetic variances (Fig. 1).

|

|
Results
In Angus cattle, heritability of yearling WT and EMA were moderate, with P8 and rib fat being highly heritable (Table 1). WT, fat depths and HT were also highly heritable in adults, with EMA being moderately heritable and BCS being low–moderate (0.13). In Tropical Composite cattle, heritabilities of young WT and muscle were higher than for Angus, but lower for young fat traits. All adult traits were highly heritable, except BCS (0.27). With the exception of HT, heritabilities in the Lambpro Composite sheep were much lower than for both cattle and Merino sheep. Heritabilities of BCS were similar to cattle in the Composite (0.17) and Merino sheep (0.15) data sets.
As expected, the two fat depth measures (P8 and rib) were highly genetically correlated with each other when measured in young or adult cattle (Table 2). WT was moderately to highly correlated with EMA or eye-muscle depth in cattle and sheep. WT was highly correlated between young and adult animals, and especially so for Merinos (0.98). Adult BCS was highly correlated with adult muscle and fat, but not young measures of muscle and fat. Adult HT was highly correlated with WTs but not composition traits including BCS.
Change in adult traits associated with genetic variation in young traits was reasonably consistent across species and breeds (Fig. 1). Adult condition score increased at the rate of an average just 0.08 scores/σG (0.02 excluding Lambpro) for EMA or eye-muscle depth, and 0.11 scores/σG (0.08 excluding Lambpro) for fat depth.
In Angus cattle, about half of the variation in cow WT (51%), HT (39%) and fat depth (P8 53%, rib 69%) was independent of variation in the yearling traits (Table 3, Fig. 2). Surprisingly, the least variation was for HT which was not one of the yearling traits included in the conditional calculation, but yearling composition traits are describing variation in HT. The majority (81%) of the genetic variation in cow muscle and condition score (81%) was independent of variation in yearling traits.

|
While WT was repeatable (little independent genetic variation) between young and adult cattle and sheep, genetic variation in BCS of Tropical Composite cattle was also largely independent (72%) of yearling WT and composition traits (Table 3, Fig. 2). The same was the case for Merino sheep (79%), although was much lower for Lambpro Composite sheep (30%).
Discussion
Cow and ewe BCS is genetically related to composition traits in young (yearling and post-weaning) animals. Accioly et al. (2018) demonstrated that selecting heifers on the basis of rib-fat EBVs clearly led to cows that were fatter across the annual production cycle. The difference between high- and low-fat Angus cow ‘lines’ in rib fat EBV was 2.5 mm and the difference in rib fat depth at first, second and third parity was 1.7, 1.2 and 2.4 mm respectively. Herein, it was estimated that the relationship between adult condition and young breeding values was stronger for fat than muscle (Fig. 1). The relationship matches the differences reported by Accioly et al. (2018) in Angus cattle and demonstrates that the current producer practice of selecting on the basis of young measures does work and is all that is currently available.
Management of ewes on the basis of condition score changes throughout the production cycle has been shown to improve productivity and be economically profitable (Edwards et al. 2011; Young et al. 2011). Lifetimewool and the subsequent Lifetime Ewe Management programs have been very successful in leading to practice change (Trompf et al. 2011). The primary principle is to train producers in assessing BCS and then managing ewes to achieve BCS targets for joining and lambing. In addition to improved management, while it is difficult to quantify, during the past 10–15 years, there have been large premiums paid for bulls and rams with greater muscle and fat EBVs, with the premise that their female offspring will be more resilient or at least require less supplementary feed. The calculations herein support this, but it is likely that far greater premiums are being paid than the possible return on investment. Thus, the aim of this study was to quantify not only the genetic variation in condition associated with young composition traits, but also the amount of genetic variation which is independent, to demonstrate the need for better tools for producers.
All of the data sets herein were large and should be representative of the populations they contain. Generally, the results were remarkably consistent across breed types and species in that the majority (about ¾) of the genetic variation in adult body condition is independent of variation in young composition traits (WT, muscle and fat, Table 3, Fig. 2). It is likely that some of this genetic variation in BCS is associated with milk production, although preliminary analyses (Pitchford WS and Lee SJ, unpubl. data) do not support a large proportion.
Body condition is more important than sufficiency for reproduction. Byrne et al. (2013) reported that, in dairy cattle, it defines the feeding management required to return the cow to adequate pre-calving condition score targets to support good production and fertility in the following lactation. Producers following Lifetime Ewe Management or beef production best-practice guidelines regularly use condition to guide supplementary feeding strategies to maximise subsequent productivity. Pitchford et al. (2018b) demonstrated that the cows in better condition are not more biologically efficient, but are likely to have eaten more during spring when feed was available.
With the exception of grass-finished cattle and purebred Merino lambs, over-fat carcasses continue to be a significant wastage cost for both producers and processors. This is exacerbated if producers select for increased young fat breeding values to improve resilience of breeding cows. Thus, better tools are required for industry to be able to make ongoing improvement in carcass quality of young stock and maternal productivity.
Conclusions
Regardless of the biological cause, cows and ewes in low condition during times of feed shortage have a significant cost in supplementary feed or lost subsequent production. Thus, condition score has a significant economic value and should be included in the breeding objective for cattle and sheep breeding programs. Given that such a large proportion of variation is independent of young muscle and fat traits, it is important that BCS be included as a selection criterion, which means being recorded in seedstock herds and flocks to then have breeding values reported for BCS. Furthermore, this would facilitate selection for increased BCS without reducing carcass meat yield. This has begun, but there is still work to do or this to be routine and for commercial producers to utilise the trait directly, instead of placing too much emphasis on yearling traits.
Data availability
All data used herein are from published papers that have been referred to.
Conflicts of interest
The author declares not conflicts of interest.
Declaration of funding
This work was supported by funding from the JS Davies Bequest to The University of Adelaide through the Davies Livestock Research Centre.
References
Accioly JM, Copping KJ, Deland MPB, Hebart ML, Herd RM, Lee SJ, Jones FM, Laurence M, Speijers EJ, Walmsley BJ, Pitchford WS (2018) Divergent breeding values for fatness or residual feed intake in Angus cattle. 4. Fat EBVs’ influence on fatness fluctuation and supplementary feeding requirements. Animal Production Science 58, 67–79.| Divergent breeding values for fatness or residual feed intake in Angus cattle. 4. Fat EBVs’ influence on fatness fluctuation and supplementary feeding requirements.Crossref | GoogleScholarGoogle Scholar |
Barwick SA, Johnston DJ, Burrow HM, Holroyd RG, Fordyce G, Wolcott ML, Sim WD, Sullivan MT (2009) Genetics of heifer performance in ‘wet’ and ‘dry’ seasons and their relationships with steer performance in two tropical beef genotypes. Animal Production Science 49, 367–382.
| Genetics of heifer performance in ‘wet’ and ‘dry’ seasons and their relationships with steer performance in two tropical beef genotypes.Crossref | GoogleScholarGoogle Scholar |
Brown DJ, Swan AA (2016) Genetic importance of fat and eye muscle depth in Merino breeding programs. Animal Production Science 56, 690–697.
| Genetic importance of fat and eye muscle depth in Merino breeding programs.Crossref | GoogleScholarGoogle Scholar |
Byrne TJ, Santos B, Amer PR, Bryant JR (2013) The economic value of body condition score in New Zealand seasonal dairying systems. In ‘Proceedings of the twentieth conference of the association for the advancement of animal breeding and genetics, translating science into action, 20th−23rd October 2013, Napier, New Zealand’. pp. 479–482. (Association for the Advancement of Animal Breeding and Genetics)
Donoghue KA, Lee SJ, Parnell PF, Pitchford WS (2018) Maternal body composition in seedstock herds. 4. Genetic parameters for body composition of Angus and Hereford cows. Animal Production Science 58, 145–155.
| Maternal body composition in seedstock herds. 4. Genetic parameters for body composition of Angus and Hereford cows.Crossref | GoogleScholarGoogle Scholar |
Edwards JEH, Copping KJ, Thompson AN (2011) Managing the nutrition of twin-bearing ewes during pregnancy using Lifetimewool recommendations increases production of twin lambs. Animal Production Science 51, 813–820.
| Managing the nutrition of twin-bearing ewes during pregnancy using Lifetimewool recommendations increases production of twin lambs.Crossref | GoogleScholarGoogle Scholar |
Hickson RE, Pitchford WS (2021) Selection strategies for beef cow size and condition. Animal Production Science 61, 1925–1931.
| Selection strategies for beef cow size and condition.Crossref | GoogleScholarGoogle Scholar |
Higham NJ (2002) Computing the nearest correlation matrix – a problem from finance. IMA Journal of Numerical Analysis 22, 329–343.
| Computing the nearest correlation matrix – a problem from finance.Crossref | GoogleScholarGoogle Scholar |
Lee SJ, Nuberg IK, Pitchford WS (2018) Maternal body composition in seedstock herds. 1. Grazing management strategy influences perspectives on optimal balance of production traits and maternal productivity. Animal Production Science 58, 117–124.
| Maternal body composition in seedstock herds. 1. Grazing management strategy influences perspectives on optimal balance of production traits and maternal productivity.Crossref | GoogleScholarGoogle Scholar |
Pitchford WS, Lee SJ (2022) Genetic variation in cow body composition is relatively independent of yearling composition. Animal Production Science 62, cvii
Pitchford WS, Accioly JM, Banks RG, Barnes AL, Barwick SA, Copping KJ, Deland MPB, Donoghue KA, Edwards N, Hebart ML, Herd RM, Jones FM, Laurence M, Lee SJ, McKiernan WA, Parnell PF, Speijers EJ, Tudor GD, Graham JF (2018a) Genesis, design and methods of the Beef CRC Maternal Productivity Project. Animal Production Science 58, 20–32.
| Genesis, design and methods of the Beef CRC Maternal Productivity Project.Crossref | GoogleScholarGoogle Scholar |
Pitchford WS, Lines DS, Wilkes MJ (2018b) Variation in residual feed intake depends on feed on offer. Animal Production Science 58, 1414–1422.
| Variation in residual feed intake depends on feed on offer.Crossref | GoogleScholarGoogle Scholar |
Trompf JP, Gordon DJ, Behrendt R, Curnow M, Kildey LC, Thompson AN (2011) Participation in Lifetime Ewe Management results in changes in stocking rate, ewe management and reproductive performance on commercial farms. Animal Production Science 51, 866–872.
| Participation in Lifetime Ewe Management results in changes in stocking rate, ewe management and reproductive performance on commercial farms.Crossref | GoogleScholarGoogle Scholar |
Walkom SF, Brown DJ (2017) Genetic evaluation of adult ewe bodyweight and condition: relationship with lamb growth, reproduction, carcass and wool production. Animal Production Science 57, 20–32.
| Genetic evaluation of adult ewe bodyweight and condition: relationship with lamb growth, reproduction, carcass and wool production.Crossref | GoogleScholarGoogle Scholar |
Walkom SF, Brien FD, Hebart ML, Pitchford WS (2016) The impact of selecting for increased ewe fat level on reproduction and its potential to reduce supplementary feeding in a commercial composite flock. Animal Production Science 56, 698–707.
| The impact of selecting for increased ewe fat level on reproduction and its potential to reduce supplementary feeding in a commercial composite flock.Crossref | GoogleScholarGoogle Scholar |
Wolcott ML, Johnston DJ, Barwick SA, Corbet NJ, Williams PJ (2014a) The genetics of cow growth and body composition at first calving in two tropical beef genotypes. Animal Production Science 54, 37–49.
| The genetics of cow growth and body composition at first calving in two tropical beef genotypes.Crossref | GoogleScholarGoogle Scholar |
Wolcott ML, Johnston DJ, Barwick SA (2014b) Genetic relationships of female reproduction with growth, body composition, maternal weaning weight and tropical adaptation in two tropical beef genotypes. Animal Production Science 54, 60–73.
| Genetic relationships of female reproduction with growth, body composition, maternal weaning weight and tropical adaptation in two tropical beef genotypes.Crossref | GoogleScholarGoogle Scholar |
Young JM, Thompson AN, Curnow M, Oldham CM (2011) Whole-farm profit and the optimum maternal liveweight profile of Merino ewe flocks lambing in winter and spring are influenced by the effects of ewe nutrition on the progeny’s survival and lifetime wool production. Animal Production Science 51, 821–833.
| Whole-farm profit and the optimum maternal liveweight profile of Merino ewe flocks lambing in winter and spring are influenced by the effects of ewe nutrition on the progeny’s survival and lifetime wool production.Crossref | GoogleScholarGoogle Scholar |