

The role of jasmonate signalling in quinolizidine alkaloid biosynthesis, wounding and aphid predation response in narrow-leafed lupin
Karen M. Frick A B C E , Rhonda C. Foley B , Kadambot H. M. Siddique C , Karam B. Singh B C D and Lars G. Kamphuis B C D F
C , Karam B. Singh B C D and Lars G. Kamphuis B C D F
A UWA School of Agriculture and Environment, The University of Western Australia, LB 5005, Perth, WA 6001, Australia.
B CSIRO Agriculture and Food, 147 Underwood Avenue, Floreat, WA 6014, Australia.
C The UWA Institute of Agriculture, The University of Western Australia, LB 5005, Perth, WA 6001, Australia.
D Centre for Crop and Disease Management, Curtin University, Bentley, WA 6102, Australia.
E Present address: Section for Plant Biochemistry, Department of Plant and Environmental Sciences, Faculty of Science, University of Copenhagen, 1871 Frederiksberg C, Denmark.
F Corresponding author. Email: lars.kamphuis@csiro.au
Functional Plant Biology 46(5) 443-454 https://doi.org/10.1071/FP18278
Submitted: 22 October 2018 Accepted: 17 January 2019 Published: 25 February 2019
Journal Compilation © CSIRO 2019 Open Access CC BY-NC-ND
Abstract
Quinolizidine alkaloids (QAs) are toxic secondary metabolites produced in lupin species that protect the plant against insects. They form in vegetative tissues and accumulate to a different extent in the grains: high levels in ‘bitter’ narrow-leafed lupin (NLL) and low levels in ‘sweet’ NLL. Grain QA levels vary considerably, and sometimes exceed the industry limit for food and feed purposes. We hypothesised that jasmonates regulate QA biosynthesis in response to environmental stresses such as wounding and aphid predation, which may explain non-genetic variability in grain QA levels. Methyl jasmonate (MeJA)-inducible genes were identified and verified in NLL. Exogenous MeJA application-induced expression of QA biosynthetic genes and QA levels for bitter, but not sweet NLL. Although MeJA-inducible genes responded to wounding, the expression of QA biosynthetic genes was not induced for bitter and sweet NLL. We assessed the effect of aphid predation on QA production for two cultivars – one moderately resistant and one susceptible to aphid predation. Although MeJA-inducible genes responded to aphid predation, no change in QA levels was found for either cultivar. These findings offer insights into the regulation of QA biosynthesis in bitter and sweet NLL and concludes that aphids are not a concern for increasing grain QAs in NLL cultivars.
Additional keywords: biotic stress, grain quality, green-peach aphid, Myzus persicae, plant defence, plant defense, secondary metabolism.
Introduction
Plants respond to biotic stress such as insect predation, by the induction of plant defence signalling pathways which can result in the accumulation of protective secondary metabolites (Zhao et al. 2005; Bari and Jones 2009). Narrow-leafed lupin (NLL; Lupinus angustifolius L.) is a major grain legume crop that produces quinolizidine alkaloids (QAs) – toxic secondary metabolites that are produced in aerial tissues and accumulate in the grain (Wink 1987; Lee et al. 2007). Levels of QAs in the grain must remain below 0.02% to be acceptable for food and feed purposes (Allen 1998); however, they often exceed this threshold and the environmental conditions that cause elevated QA levels are not well understood (Cowling and Tarr 2004). Although studies have investigated the role of abiotic factors on grain QA content in NLL, such as drought, temperature, soil pH and nutrient availability (Christiansen et al. 1997; Gremigni et al. 2003; Jansen et al. 2009, 2012; Frick et al. 2018), there are fewer reports on the impact of biotic stresses (Pilar Vilariño et al. 2005; Chludil et al. 2009, 2013). This is an important consideration, as QAs protect the plant from insect pests (Wink 1992) and, therefore, their biosynthesis may be induced in response to insect herbivory, possibly explaining why grain QA content is highly unpredictable when only abiotic factors are assessed (Cowling and Tarr 2004).
The literature surrounding the response of QA levels in lupins to biotic stress provides no clear picture, and the response may vary depending on the species or their intrinsic QA levels. Mechanical damage increases QA levels in aerial tissues of Lupinus polyphyllus Lindl. and Lupinus succulentus Dougl. ex K. Koch (Wink 1983; Johnson et al. 1989) and both low- and high-QA NLL (Chludil et al. 2009, 2013). However, when fed on by caterpillars, leaf QA levels did not increase in either low- or high-QA NLL, but increased in low- and high-QA Lupinus albus L. (Pilar Vilariño et al. 2005). Any changes in the expression levels of key QA biosynthetic genes in response to biotic stresses still needs to be assessed.
The jasmonic acid (JA) plant defence signalling pathway plays a major role in regulating defence responses to biotic stressors and JA can induce the accumulation of a wide variety of secondary metabolites in plants (Zhao et al. 2005; Wasternack and Hause 2013). Jasmonic acid pathway-associated responses to stress signals involve the rapid synthesis and accumulation of JA, which induces the expression of key transcription factors that modulate distinct JA-dependent functions (Wasternack and Hause 2013). The application of exogenous JA or methyl jasmonate (MeJA) to plants can induce JA signalling; genes that are typically upregulated in response to JA are those involved in its biosynthesis (e.g. LIPOXYGENASE; LOX gene family members) (Wasternack 2007), as well as downstream genes involved in plant defence, such as PROTEINASE INHIBITORS (PI) (Farmer and Ryan 1990). Jasmonic acid accumulates in response to wounding caused by chewing insects or mechanical damage (Howe and Jander 2008), and is involved in responses to phloem-feeding insects such as aphids (Ellis et al. 2002; Gao et al. 2007).
Although no studies have previously assessed plant responses to JA in NLL, JA is a well-known activator of the biosynthesis of alkaloids produced in other plant species, such as pyridine alkaloids (Nicotiana spp.) and the monoterpenoid indole alkaloids (Catharanthus roseus (L.) G.Don) (Aerts et al. 1994; Baldwin 1996; Huang and Kutchan 2000; Ryan et al. 2015). The application of exogenous JA or MeJA increases the levels of these alkaloids, as well as the expression of biosynthetic genes, transcription factors that regulate the pathway, and genes involved in alkaloid transport throughout the plant (Memelink et al. 2001; Shoji et al. 2009; De Boer et al. 2011; Kato et al. 2015). Mechanical damage causes a JA-associated increase in nicotine levels of Nicotiana spp. leaves (Baldwin et al. 1994) and both mechanical and herbivore damage leads to increased alkaloid levels in Atropa acuminata Royle ex Lindl. leaves (Khan and Harborne 1990).
The main insect pest affecting NLL crops are aphids, which cause major yield losses in susceptible cultivars by feeding damage and the transmission of plant viruses (Berlandier and Sweetingham 2003). Aphid resistance has been linked to QA levels in leaf and grain tissues (Adhikari et al. 2012; Philippi et al. 2015) and ‘sweet’ NLL cultivars, harbouring the low-QA domestication locus iucundus (iuc) (von Sengbusch 1942) are much more susceptible to aphid predation than ‘bitter’ wild NLL accessions that produce high QA levels (Philippi et al. 2015). The response of QA biosynthesis to predation by phloem feeding insects such as aphids has not yet been assessed. This response may differ to that of wounding or chewing insects such as caterpillars as phloem-feeding insects probe plant tissue intercellularly, minimising wounding of the plant and resulting in distinct transcriptional responses (Appel et al. 2014).
In the present study we assessed the response of the JA signalling pathway and QA biosynthesis to exogenous MeJA, mechanical damage and aphid predation (Myzus persicae Sulzer), to better understand how biotic stressors affect QA levels in NLL. We assessed the response of the JA pathway in NLL to these treatments by identifying and measuring the expression of genes homologous to those related to JA signalling and induced by MeJA and aphid predation in the model legume Medicago truncatula Gaertn. (MtLOX4, MtLOX5, and MtPI), which served as marker genes (Gao et al. 2007). We then measured the expression of four key QA biosynthetic genes in response to these treatments and compared this expression to the observed induction of the JA pathway. Although the QA biosynthetic pathway is only partly elucidated (Frick et al. 2017), genes that have been molecularly characterised as having a role in QA biosynthesis include L. angustifolius lysine/ornithine decarboxylase (LaL/ODC), L. angustifolius copper amine oxidase (LaCAO) and L. albus tigloyl-CoA:(–)-13α-hydroxymultiflorine/(+)-13α-hydroxylupanine O-tigloyltransferase (LaHMT/HLT) (Okada et al. 2005; Bunsupa et al. 2012; Yang et al. 2017a). L. angustifolius acyltransferase (LaAT) has also been suggested as being involved in QA biosynthesis (Bunsupa et al. 2011). Finally, we measured QA levels to assess how the expression of these key QA biosynthetic genes correspond to QA levels in leaves, and in the case of aphid predation, also in grain.
Materials and methods
NLL varieties
High (bitter) and low (sweet) QA producing NLL (accession P27255 and cultivar Tanjil, respectively) were used to assess the impact of exogenous MeJA on QA production, as these accessions have also been used to create genomic and transcriptomic resources for the crop (Kamphuis et al. 2015; Hane et al. 2017). Tanjil also shows moderate insect resistance (Edwards et al. 2003). As only specialised aphids use bitter NLL as host plants which are not found in Australia (Wink and Witte 1991; Philippi et al. 2015), cultivars Kalya and Tallerack were used to assess the impact of aphid feeding in sweet NLL; Kalya is moderately resistant to aphids, whereas Tallerack is susceptible (Berlandier and Sweetingham 2003; Edwards et al. 2003).
Growth conditions
Seeds of cultivars Kalya and Tallerack were treated with P-Pickle T fungicide treatment and two seeds were planted in pots (115 mm diameter, 120 mm height) filled with steam-sterilised sandy soil, treated with Rhizobium group G and beneficial soil microbes (GOGO juice). Plants were grown at 20/16°C day/night cycle and 12 h of light and watered twice a week. At 14 days after sowing (DAS), plants were trimmed to one plant per pot. At 31 DAS, plants were watered with half-strength Thrive liquid fertiliser. Plants at this stage were used to assess the responses in leaves to aphid predation, while growth was continued for those used to assess the response in grain.
For bitter accession P27255 and cultivar Tanjil, growth was carried out as outlined above, except seeds were scarified using sharp pliers before sowing, and sown in square pots (85 mm width, 180 mm height) and grown with 14 h of light.
Aphids
The green-peach aphid (Myzus persicae Sulzer) is one of three common aphid species found on NLL (Sandow 1987). A colony of green-peach aphid clone C61 was maintained on radish (Raphanus sativus L.) plants. The green-peach aphids were transferred to the experimental plants with a fine paintbrush.
Experimental conditions
Assessing leaf responses to MeJA treatment
At 28 DAS, eight pots of accession P27255 or cultivar Tanjil were placed in 30 L sealed containers with cotton buds soaked in 150 µL ethanol or 150 µL of ethanol with MeJA (Sigma Aldrich) for a final concentration of 0.9 µM MeJA L–1 air. All leaf tissue from each plant was collected after 6, 12 and 36 h, frozen immediately in liquid nitrogen, and stored at −80°C until further analysis. Leaf tissue was collected at the same time each day (~1200 hours).
Assessing leaf responses to mechanical wounding
At 21 DAS, plants of accession P27255 or cultivar Tanjil were either undamaged or wounded by rolling a fabric pattern wheel over leaves. Leaf tissue was collected at 5, 24, 72 and 120 h after wounding, frozen immediately in liquid nitrogen and stored at −80°C until further analysis. For the mechanical wounding treatment, a single leaf (including all leaflets) was damaged on each plant, and the entire leaf was collected. Single leaves at a similar developmental stage to the wounded leaves were collected from separate plants for the control treatment. All leaf material was collected at the same time each day (1400–1600 hours).
Assessing leaf responses to aphid predation
At 33 DAS, control plants of cultivars Kalya and Tallerack were enclosed in a plastic bottle with mesh sides to allow air flow. Aphid-infested plants were enclosed in the same type of bottles each with 60 green-peach aphids. Aphids were allowed to feed for 24, 48 or 72 h, and all leaf material was collected – taking care to remove aphids – frozen immediately in liquid nitrogen and stored at −80°C until further analysis. All leaf material was collected at the same time each day (1400–1600 hours).
Assessing grain responses to aphid attack
At 54 DAS, plants were placed in large mesh cages in a temperature-controlled glasshouse with natural lighting and treated again with half-strength Thrive and beneficial soil microbes (GOGO juice). Plants for control treatments were placed in one cage, and Kalya and Tallerack plants for aphid infestations were placed in two separate cages. Once plants had begun to set pods on the main stem (73 DAS) four green-peach aphids were placed on each plant, with an additional 30 aphids placed on each plant a week later. One month after the initial aphid infestation (101 DAS), all Tallerack plants were treated with insecticide (Confidor, Yates), as plants were heavily infested, to ensure continued seed development. At 115 DAS, the main stem pods of both Kalya and Tallerack were mature and plants were left to dry an additional week before collecting seed, which was stored at room temperature until further analysis. For the above experiments, one biological replicate represents material from an individual plant.
QA quantification in NLL grain and leaf tissues
Mature grain and leaf material was ground to a fine powder under liquid nitrogen using a mortar and pestle. Tissue samples were submitted to the ChemCentre for QA analysis by capillary gas chromatography using 0.5 g of material and in-house method ORG180 (Priddis 1983; Harris and Wilson 1988). The limit of quantification for each QA is 0.001%.
RNA extraction and cDNA synthesis
Leaf tissue was ground to a fine powder under liquid nitrogen using a chilled pestle. RNA extractions were conducted using ~100 mg of tissue (from the uppermost leaves of aphid-infested plants or homogenised leaf material from MeJA-treated plants). RNA extraction and cDNA synthesis were carried out as described by Frick et al. (2018).
Primer design and quantitative real-time –PCR
Primers used in this study for quantitative real-time –PCR have been described previously (Frick et al. 2018). These primers are used to amplify the quantitative real-time PCR reference gene L. angustifolius UBC9-like (LaUBC9-like; Lup018255.1), QA biosynthetic genes LaL/ODC (Lup009726.1) (Bunsupa et al. 2012), LaCAO (Lup000530.1) (Yang et al. 2017a) and LaHMT/HLT (Lup022251.1) (Okada et al. 2005) and candidate biosynthetic gene LaAT (Lup021586.1) (Bunsupa et al. 2011).
Candidate MeJA-inducible genes in NLL were chosen based on annotated genes (Hane et al. 2017) with the highest BLAST-sequence homology to M. truncatula MeJA-inducible genes M. truncatula lipoxygenase 4 (MtLOX4; TC100162), M. truncatula lipoxygenase 5 (MtLOX5; TC100513) and M. truncatula proteinase inhibitor (MtPI; TC100490) (Gao et al. 2007). These genes are Lup026769.1 (LaLOX4-like), Lup003340.1 (LaLOX5-like) and Lup009615.1 (LaPI-like). Primers were designed as outlined in Frick et al. (2018). Primer sequences are listed in Table S1, available as Supplementary Material to this paper.
The quantitative real-time PCR was conducted according to Frick et al. (2018), performed in 10 µL reactions with 5 µL SsoFast Evagreen Supermix, 4 µL 1 : 50 cDNA reaction dilution and 1 µL primer mix containing 5 µM each of forward and reverse primers, including a reference gene and non-template control on each plate. Relative gene expression was calculated using the 2–ΔΔCT method (Livak and Schmittgen 2001) to normalise the average Ct values of technical replicates to the reference gene LaUBC9-like.
Statistical analyses
Statistical analyses were conducted using Genstat (18th edn). Data for gene expression analyses are presented as the mean and standard error (s.e.) of three biological replicates. A one-sided two sample t-test was used to assess whether gene expression of MeJA-inducible genes or QA biosynthetic genes increased under certain conditions compared with control plants at a certain time point. Three-way analysis of variance (ANOVA) was used to assess the effect of aphid treatment, cultivar, time and their interactions on QA biosynthetic gene expression in leaf tissue. Data for leaf QA content assessing the impact of MeJA is presented as the mean and s.e. of five biological replicates. Two-way ANOVA was used to assess the effect of MeJA treatment, time, and MeJA treatment × time interaction on QA content in leaf tissue. Data for leaf and grain QA content assessing the impact of aphid predation is presented as the mean and s.e. of six biological replicates. Two-way ANOVA was used to assess the effect of NLL cultivar, aphid infestation, and cultivar × aphid infestation interaction on QA content in leaf and grain tissue. Means were compared at the P ≤ 0.05 level of significance.
Results
Identification and verification of NLL MeJA-inducible genes
To identify genes that may be used as markers for JA induction in NLL, genes were identified from the NLL gene annotation (Hane et al. 2017) that have the highest sequence similarity to those related to JA signalling that are highly upregulated in M. truncatula following treatment with exogenous MeJA and aphid predation (Gao et al. 2007). These include two genes encoding LOX, putatively involved in JA biosynthesis (LaLOX4-like; Lup026769.1 and LaLOX5-like; Lup003340.1), and one encoding PI (LaPI-like; Lup009615.1), a downstream JA-responsive gene. To assess whether these genes were indeed MeJA-inducible, the expression of these genes was investigated in leaves of bitter accession P27255 and sweet cultivar Tanjil following exogenous MeJA treatment. For bitter accession P27255, the expression of LaLOX4-like was induced at 12 and 36 h after treatment with MeJA (P ≤ 0.05), LaLOX5-like was induced at 6 and 12 h after MeJA treatment (P ≤ 0.05), and LaPI-like was induced at 36 h after MeJA treatment (P ≤ 0.05) (Fig. 1). For sweet cultivar Tanjil, LaLOX4-like was not induced following MeJA treatment, whereas LaLOX5-like was induced after 36 h. Although LaPI-like had higher transcript levels in Tanjil MeJA-treated leaves, these did not significantly differ from the control by one-sided two-sample t-test (P ≤ 0.05). In conclusion, the three genes were MeJA inducible in the bitter accession P27255, but not all genes were induced in the sweet cultivar Tanjil. As these genes were demonstrated as MeJA inducible, at least in bitter accession P27255, these were selected for use as gene markers to indicate induction of the JA pathway in subsequent experiments and are hereafter referred to as ‘MeJA-inducible’ genes.

|
Response of MeJA-inducible genes to mechanical wounding and aphid predation
The response of NLL MeJA-inducible genes was assessed following treatment of two conditions that can induce the JA plant signalling pathway: mechanical wounding and aphid predation (Wasternack and Hause 2013). For both bitter accession P27255 and sweet cultivar Tanjil, LaLOX4-like was induced at 24 h after wounding (P ≤ 0.05), whereas LaPI-like was induced after 24 h for P25255, but a significant induction was not observed for Tanjil (Fig. 2). In contrast to LaLOX4-like, LaLOX5-like expression was not strongly-induced with wounding for either accession. For both LaLOX4-like and LaPI-like, the strongest increase in transcript abundance was observed at 24 h post wounding in both bitter and sweet NLL.

|
After observing a response of MeJA-inducible genes following MeJA treatment and mechanical wounding in both bitter and sweet NLL, how these genes respond to aphid predation was assessed as the JA plant signalling pathway is known to play a role in the defence against phloem-feeding insects (Ellis et al. 2002; Gao et al. 2007). As aphid species are generally unable to infest bitter varieties of NLL (Wink and Witte 1991; Philippi et al. 2015), the response of sweet cultivars Kalya and Tallerack was investigated because these cultivars have varied responses to aphids. Kalya is a moderately aphid-resistant cultivar while Tallerack is susceptible to aphid predation (Berlandier and Sweetingham 2003; Edwards et al. 2003). For resistant cultivar Kalya, the expression of LaLOX4-like and LaPI-like was induced at 24, 48 and 72 h after aphid infestation (P ≤ 0.05), whereas for susceptible cultivar Tallerack, LaLOX4-like was only induced at 72 h after aphid infestation (P ≤ 0.05), and LaPI-like was not induced with aphid predation (Fig. 3a, c). LaLOX5-like did not increase expression in aphid infested samples compared with controls in either cultivar, although expression was higher for Kalya than Tallerack (P ≤ 0.05) (Fig. 3b). In conclusion, two of the three candidate NLL MeJA-inducible genes were induced to a greater extent for the NLL moderately aphid-resistant cultivar Kalya than the aphid-susceptible cultivar Tallerack upon aphid predation.

|
Response of QA biosynthetic genes to MeJA
Having indicated how the JA signalling pathway in NLL was responding to exogenous MeJA application, wounding and aphid predation, how this relates to the expression of genes involved in QA biosynthesis was then assessed, as alkaloids produced in other plant species are regulated by the JA signalling pathway (Aerts et al. 1994; Baldwin 1996; Huang and Kutchan 2000; Ryan et al. 2015). Following MeJA treatment, three of the four QA biosynthetic genes (LaL/ODC, LaCAO and LaAT) displayed similar expression patterns and were induced in leaf tissue of bitter accession P27255 at 12 h and/or 36 h after MeJA treatment (P ≤ 0.05) but were not induced in sweet cultivar Tanjil (Fig. 4a–c). The expression of LaHMT/HLT was induced at 6 and 36 h after MeJA treatment for bitter accession P27255 (P ≤ 0.05). Although a small significant increase in transcript abundance of LaHMT/HLT was observed 36 h after MeJA treatment compared with the control for cultivar Tanjil (P ≤ 0.05), this abundance was far lower than the levels observed in the P27255 MeJA-treated and control leaf samples at 36 h (Fig. 4d). In conclusion, for bitter accession P27255, all four QA biosynthetic genes were upregulated following MeJA treatment, whereas no strong induction was observed in the sweet cultivar Tanjil.

|
MeJA induces QA levels in bitter NLL but not sweet NLL
After observing a differential response of both MeJA-inducible and key QA biosynthetic genes to exogenous MeJA application in bitter and sweet NLL, with the expression of key QA biosynthetic genes being induced for bitter but not sweet NLL, we investigated the effect of MeJA application on leaf QA levels. For bitter accession P27255, total QA levels increased by 22% following MeJA treatment, independent of whether the plants were treated with MeJA for 12 or 36 h (P ≤ 0.05) (Fig. 5). This was largely due to a 42% increase in lupanine levels after MeJA treatment (P ≤ 0.05). Angustifoline and 13-hydroxylupanine levels increased for P27255 after MeJA treatment (P ≤ 0.05) (16 and 18% respectively), and 13-hydroxylupanine also increased over time (P ≤ 0.05). Levels of α-isolupanine increased for P27255 over time (P ≤ 0.05) but did not increase with MeJA treatment. The QAs in sweet NLL cultivar Tanjil were at levels below the limit of quantification at 36 h after MeJA treatment. However, 13-hydroxylupanine was detectable in both MeJA-treated and control plants (Table S2). Overall, leaf QA levels increased following MeJA treatment for bitter accession P27255, while no significant changes were observed for Tanjil.
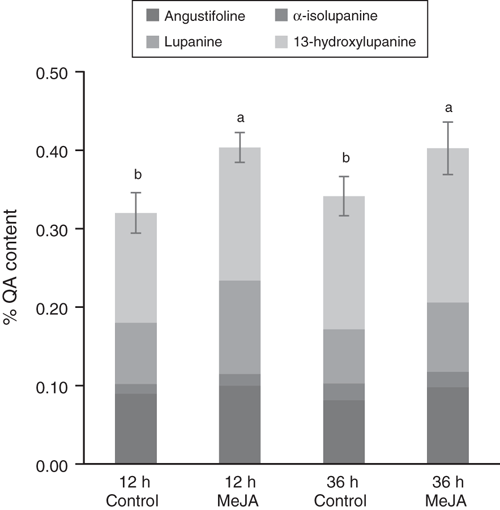
|
Wounding and aphid predation does not affect the expression of QA biosynthetic genes
Following mechanical wounding, none of the four QA biosynthetic genes was induced in either bitter accession P27255 or sweet cultivar Tanjil, although LaL/ODC, LaCAO, and LaAT displayed higher basal expression levels in P27255 compared with Tanjil (see Fig. S1, available as Supplementary Material to this paper). This was consistent with the higher expression levels of these genes in P27255 than Tanjil observed in the MeJA treatments (Fig. 4).
Following aphid predation, the expression of QA biosynthetic genes LaL/ODC, LaCAO and LaHMT/HLT did not change in leaf tissue of both moderately aphid-resistant cultivar Kalya and aphid-susceptible cultivar Tallerack (Fig. S2). The expression of LaL/ODC and LaCAO was also similar between cultivars, and LaHMT/HLT expression was higher in Kalya (P ≤ 0.05) than Tallerack. LaAT expression was barely detected or absent in all samples (data not shown). In conclusion, genes involved in QA biosynthesis appear to be responsive to MeJA treatment in bitter NLL material but not in sweet cultivars. Furthermore, both wounding and aphid predation does not induce the expression of QA biosynthetic genes in sweet cultivars.
Aphid predation does not induce QA levels in moderately aphid-resistant or aphid-susceptible sweet NLL cultivars
While no response of QA biosynthetic gene expression was observed following aphid predation for both moderately aphid-resistant cultivar Kalya and aphid-susceptible cultivar Tallerack (Fig. S2), QA levels in leaf and grain were assessed because QA levels may be affected by factors other than the expression of key QA biosynthetic genes. The impact of aphid feeding on leaf QA levels was assessed at 72 h after infestation, however, no effect of aphid feeding for either cultivar was found. Leaf QA levels for Kalya were significantly higher than for Tallerack (P ≤ 0.05), with QA levels in Tallerack below the limit of quantification and not detected for most samples (Tables 1, S3). For cultivar Kalya, only 13-hydroxylupanine was present at levels above the limit of quantification (Table S3).

|
We then assessed the impact of aphid predation on grain QA content in resistant and susceptible NLL cultivars subjected to sustained aphid predation over pod set. Although no changes in QA biosynthesis or QA content were identified with aphid predation in leaves, the accumulation of QAs in the grain may be affected. However, no change in total QA levels, or levels of individual QAs (13-hydroxylupanine, lupanine, angustifoline and α-isolupanine) was found in either cultivar under aphid predation. Cultivar Kalya accumulated more total QAs than cultivar Tallerack, as well as levels of 13-hydroxylupanine and angustifoline (P ≤ 0.05) (Fig. 6). A significant interaction between NLL cultivar and aphid infestation was found for α-isolupanine (P ≤ 0.05), such that Kalya aphid-infested grain had higher levels than Tallerack aphid-infested grain.
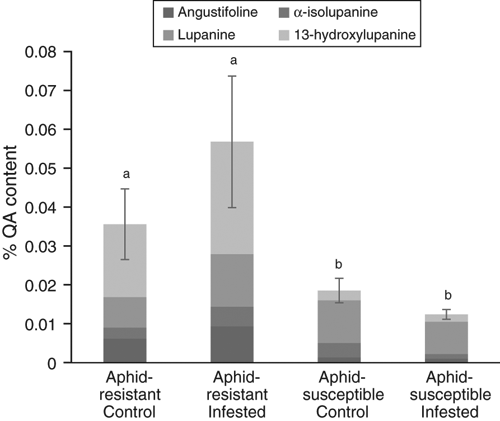
|
Discussion
Quinolizidine alkaloids are the main anti-nutritional factor in NLL grain and pose a problem as the grain cannot be used for food and feed purposes if levels exceed the industry threshold (0.02%) (Allen 1998). However, grain QA levels are highly unpredictable when only abiotic factors are assessed (Cowling and Tarr 2004). We hypothesised that the JA plant signalling pathway plays a role in the regulation of QA biosynthesis in NLL in response to biotic stresses, which may explain some environmental variability found in grain QA content in NLL cultivars. Jasmonic acid is involved in plant defence responses to mechanical wounding and insect predation (Wasternack and Hause 2013) and is known to regulate the biosynthesis of alkaloids found in other plant species (e.g. Nicotiana spp. and Catharanthus roseus) (Aerts et al. 1994; Baldwin 1996; Huang and Kutchan 2000; Ryan et al. 2015). In the present study we assessed the response of the JA plant signalling pathway and QA biosynthesis to exogenous MeJA treatment and mechanical wounding, which provides new insights in the responses of both bitter and sweet NLL to these treatments. We then assessed the response of NLL cultivars to the green-peach aphid, a major insect pest of NLL, to identify whether this increases QA levels.
The exogenous MeJA treatment induced the expression of three candidate MeJA-inducible genes in bitter NLL, which were identified based on sequence homology to M. truncatula MeJA-inducible genes (Gao et al. 2007), demonstrating that the JA signalling pathway was induced (Fig. 1). Three QA biosynthetic genes (LaL/ODC, LaCAO and LaAT) were similarly induced in bitter NLL following exogenous MeJA treatment (Fig. 4), which corresponded to an increase in leaf QA levels (Fig. 5). These results demonstrate a direct role of the JA signalling pathway in the regulation of QA biosynthesis in bitter NLL. This will assist efforts to uncover other genes involved in QA biosynthesis, as this pathway is largely unknown (Frick et al. 2017). For example, transcriptome profiling of MeJA-induced bitter NLL tissues could be undertaken to identify QA biosynthetic genes or regulatory genes regulated in a similar manner to LaL/ODC, LaCAO and LaAT, a method previously used to uncover genes associated with alkaloid biosynthesis in Papaver somniferum and Nicotiana tabacum (Gurkok et al. 2015; Yang et al. 2017b).
We have demonstrated that JA is involved in the regulation of QA biosynthesis in bitter NLL, although QA biosynthetic genes were not induced following mechanical wounding (Fig. S1) despite an induction in MeJA-inducible genes (Fig. 2). The observed induction of QA biosynthesis in bitter NLL following exogenous MeJA treatment, but not wounding, may due to cross-talk of the JA signalling pathway with other plant signalling pathways that are induced after wounding, such as abscisic acid and ethylene pathways (León et al. 2001). Ethylene suppresses JA-mediated alkaloid biosynthetic gene induction in Nicotiana spp., and attack by the specialist herbivore Manduca sexta releases ethylene, suppressing nicotine accumulation that results from wounding alone (Shoji et al. 2000; Winz and Baldwin 2001). The role of other plant signalling pathways, such as ethylene, and the cross-talk between ethylene and JA pathways in NLL and for QA biosynthesis remains to be investigated. In summary, for bitter NLL, both wounding and MeJA application recruit JA- responsive genes (LaLOX4-like, LaLOX5-like and LaPI-like). Furthermore, wounding did not upregulate QA biosynthetic genes whereas MeJA application did (Figs 4, S1).
Additionally, although we have demonstrated an involvement of JA in QA biosynthesis for bitter NLL, this was not the case for sweet NLL as neither expression of QA biosynthetic genes (Fig. 4), nor leaf QA levels (Table S2) were induced following exogenous MeJA treatment. Moreover, MeJA-inducible genes were also not induced, or induced to a lesser extent for sweet NLL than bitter NLL (Fig. 1). Following mechanical wounding, sweet NLL displayed a comparative induction of MeJA-inducible genes to bitter NLL (Fig. 2), indicating that JA biosynthesis is not impaired for sweet NLL. This suggests that either the response of MeJA-inducible genes in sweet NLL was not captured by the time points sampled, or that sweet NLL is less responsive to exogenous MeJA treatment than bitter NLL.
After demonstrating that JA is involved in the regulation of QA biosynthesis in bitter NLL, we investigated the effect of aphid predation in NLL cultivars as aphids are the major insect pest affecting NLL crops (Berlandier and Sweetingham 2003) and JA is associated with resistance to some phloem-feeding insects (Ellis et al. 2002; Gao et al. 2007). Only specialised aphids use bitter NLL as host plants and these are not found in Australia (Wink and Witte 1991; Philippi et al. 2015), therefore we assessed the effect of aphid predation in two sweet NLL cultivars – one moderately resistant (cultivar Kalya) and one susceptible (cultivar Tallerack) to aphid predation. Following aphid predation, the induction of MeJA-inducible genes differed between the two cultivars, with a higher response for cultivar Kalya than Tallerack (Fig. 3). Kalya demonstrated a greater JA response to aphid predation than Tallerack and produced higher intrinsic QA levels in leaves (Table 1), although no differences in QA biosynthetic gene expression were observed, both following aphid predation and between the two cultivars (Fig. S2).
We assessed the effect of sustained aphid predation during pod set on grain QA levels in the two cultivars, as it is possible that aphid predation increases grain QA levels through increased transport to the grain, rather than increased biosynthesis in the leaves. Surprisingly, no response in grain QA levels was found following aphid predation, although Kalya had higher intrinsic QA levels than Tallerack (Fig. 6). Therefore, we conclude that aphid predation by Myzus persicae does not increase grain QA levels in sweet NLL cultivars. However, changes in response to other aphid species found on NLL, such as blue-green aphid (Acyrthosiphon kondoi) and cowpea aphid (Aphis craccivora) (Sandow 1987), require further investigation. The aphid resistance of Kalya may be conferred by intrinsic QA levels that do not increase with aphid predation or by other defence mechanisms that are induced to a greater extent than for Tallerack as induction of MeJA-inducible genes differed between the two cultivars (Fig. 3), suggesting differential induction of the JA signalling pathway. If the genetic mechanism underlying aphid resistance for Kalya is, in part, unrelated to QA levels, this would be of interest to NLL breeders.
Finally, our findings from this study, combined with our previous findings examining QA biosynthetic gene expression following drought and temperature stresses in NLL cultivars (Frick et al. 2018), reinforces our conclusion that QA levels in NLL cultivars are not controlled at the gene regulation level of the key QA biosynthetic genes. The expression of LaL/ODC and LaCAO does not differ between NLL cultivars producing different leaf and grain QA levels (Figs 6, S2; Table S3) (Frick et al. 2018). Since a clear induction of QA biosynthetic gene expression occurs in bitter NLL following exogenous MeJA treatment, but no differences are evident in the expression of QA biosynthetic genes between several treatments for NLL cultivars (Fig. S2; Frick et al. 2018), we suggest that the gene controlling the low-QA locus iuc, harboured by all modern NLL cultivars, is a major QA regulatory gene. The low-QA iuc phenotype may be caused by the loss-of-function of this regulatory gene, resulting in a low basal expression of QA biosynthetic genes that do not respond to environmental conditions without the action of this regulatory gene. Low alkaloid loci for Nicotiana tabacum (nic1 and nic2) positively regulate several genes of the pyridine alkaloid biosynthetic pathway (Hibi et al. 1994; Cane et al. 2005), and suppression of one of the genes deleted in nic2 (NtERF189) suppresses MeJA-induced nicotine biosynthesis (Shoji et al. 2010). We suggest that the NLL iuc locus functions in a similarly to these N. tabacum low alkaloid loci and may explain why QA biosynthesis is not induced after aphid predation in NLL cultivars.
Conclusion
The present study is the first to assess responses of the JA plant signalling pathway in NLL and how this pathway regulates QA biosynthesis. Our results have demonstrated an involvement of JA in the regulation of QA biosynthesis for bitter NLL, but this was not observed for sweet NLL. Additionally, aphid predation did not induce QA biosynthetic gene expression in sweet NLL cultivars that were susceptible or moderately resistant to an aphid pest, M. persicae, resulting in no differences in leaf nor mature grain levels following aphid predation. One possibility is that the gene controlling the low-QA locus iuc is a major QA regulatory gene and the low-QA phenotype of iuc-harbouring cultivars results in QA biosynthetic genes that do not respond to environmental conditions or JA signalling.
Conflicts of interest
The authors declare no conflicts of interest.
Acknowledgements
We thank Ling-Ling Gao and Kathleen DeBoer for their advice on this project. We thank Jens Berger and Louise Thatcher for critical reading of the manuscript. KMF was supported by a Grains Industry Research Scholarship from the Grains Research and Development Corporation (GRS10935) and a University Postgraduate Award from The University of Western Australia.
References
Adhikari KN, Edwards OR, Wang S, Ridsdill-Smith TJ, Buirchell B (2012) The role of alkaloids in conferring aphid resistance in yellow lupin (Lupinus luteus L.). Crop and Pasture Science 63, 444–451.| The role of alkaloids in conferring aphid resistance in yellow lupin (Lupinus luteus L.).Crossref | GoogleScholarGoogle Scholar |
Aerts RJ, Gisi D, Carolis E, Luca V, Baumann TW (1994) Methyl jasmonate vapor increases the developmentally controlled synthesis of alkaloids in Catharanthus and Cinchona seedlings. The Plant Journal 5, 635–643.
| Methyl jasmonate vapor increases the developmentally controlled synthesis of alkaloids in Catharanthus and Cinchona seedlings.Crossref | GoogleScholarGoogle Scholar |
Allen JG (1998) Toxins and lupinosis. In ‘Lupins as crop plants: biology, production and utilization’. (Eds JS Gladstone, CA Atkins, J Hamblin) pp. 411–428. (CAB International: Cambridge, UK)
Appel HM, Fescemyer H, Ehlting J, Weston D, Rehrig E, Joshi T, Xu D, Bohlmann J, Schultz J (2014) Transcriptional responses of Arabidopsis thaliana to chewing and sucking insect herbivores. Frontiers of Plant Science 5, 565
| Transcriptional responses of Arabidopsis thaliana to chewing and sucking insect herbivores.Crossref | GoogleScholarGoogle Scholar |
Baldwin IT (1996) Methyl jasmonate‐induced nicotine production in Nicotiana attenuata: inducing defenses in the field without wounding. Entomologia Experimentalis et Applicata 80, 213–220.
| Methyl jasmonate‐induced nicotine production in Nicotiana attenuata: inducing defenses in the field without wounding.Crossref | GoogleScholarGoogle Scholar |
Baldwin IT, Schmelz EA, Ohnmeiss TE (1994) Wound-induced changes in root and shoot jasmonic acid pools correlate with induced nicotine synthesis in Nicotiana sylvestris spegazzini and comes. Journal of Chemical Ecology 20, 2139–2157.
| Wound-induced changes in root and shoot jasmonic acid pools correlate with induced nicotine synthesis in Nicotiana sylvestris spegazzini and comes.Crossref | GoogleScholarGoogle Scholar | 24242736PubMed |
Bari R, Jones JD (2009) Role of plant hormones in plant defence responses. Plant Molecular Biology 69, 473–488.
| Role of plant hormones in plant defence responses.Crossref | GoogleScholarGoogle Scholar | 19083153PubMed |
Berlandier FA, Sweetingham MW (2003) Aphid feeding damage causes large losses in susceptible lupin cultivars. Animal Production Science 43, 1357–1362.
| Aphid feeding damage causes large losses in susceptible lupin cultivars.Crossref | GoogleScholarGoogle Scholar |
Bunsupa S, Okada T, Saito K, Yamazaki M (2011) An acyltransferase-like gene obtained by differential gene expression profiles of quinolizidine alkaloid-producing and nonproducing cultivars of Lupinus angustifolius. Plant Biotechnology 28, 89–94.
| An acyltransferase-like gene obtained by differential gene expression profiles of quinolizidine alkaloid-producing and nonproducing cultivars of Lupinus angustifolius.Crossref | GoogleScholarGoogle Scholar |
Bunsupa S, Katayama K, Ikeura E, Oikawa A, Toyooka K, Saito K, Yamazaki M (2012) Lysine decarboxylase catalyzes the first step of quinolizidine alkaloid biosynthesis and coevolved with alkaloid production in Leguminosae. The Plant Cell 24, 1202–1216.
| Lysine decarboxylase catalyzes the first step of quinolizidine alkaloid biosynthesis and coevolved with alkaloid production in Leguminosae.Crossref | GoogleScholarGoogle Scholar | 22415272PubMed |
Cane KA, Mayer M, Lidgett AJ, Michael AJ, Hamill JD (2005) Molecular analysis of alkaloid metabolism in AABB v. aabb genotype Nicotiana tabacum in response to wounding of aerial tissues and methyl jasmonate treatment of cultured roots. Functional Plant Biology 32, 305–320.
| Molecular analysis of alkaloid metabolism in AABB v. aabb genotype Nicotiana tabacum in response to wounding of aerial tissues and methyl jasmonate treatment of cultured roots.Crossref | GoogleScholarGoogle Scholar |
Chludil HD, Vilariño MDP, Franco ML, Leicach SR (2009) Changes in Lupinus albus and Lupinus angustifolius alkaloid profiles in response to mechanical damage. Journal of Agricultural and Food Chemistry 57, 6107–6113.
| Changes in Lupinus albus and Lupinus angustifolius alkaloid profiles in response to mechanical damage.Crossref | GoogleScholarGoogle Scholar | 19537792PubMed |
Chludil HD, Leicach SR, Corbino GB, Barriga LG, Vilariño MdP (2013) Genistin and quinolizidine alkaloid induction in L. angustifolius aerial parts in response to mechanical damage. Journal of Plant Interactions 8, 117–124.
| Genistin and quinolizidine alkaloid induction in L. angustifolius aerial parts in response to mechanical damage.Crossref | GoogleScholarGoogle Scholar |
Christiansen JL, Jørnsgård B, Buskov S, Olsen CE (1997) Effect of drought stress on content and composition of seed alkaloids in narrow-leafed lupin, Lupinus angustifolius L. European Journal of Agronomy 7, 307–314.
| Effect of drought stress on content and composition of seed alkaloids in narrow-leafed lupin, Lupinus angustifolius L.Crossref | GoogleScholarGoogle Scholar |
Cowling WA, Tarr A (2004) Effect of genotype and environment on seed quality in sweet narrow-leafed lupin (Lupinus angustifolius L.). Australian Journal of Agricultural Research 55, 745–751.
| Effect of genotype and environment on seed quality in sweet narrow-leafed lupin (Lupinus angustifolius L.).Crossref | GoogleScholarGoogle Scholar |
De Boer K, Tilleman S, Pauwels L, Vanden Bossche R, De Sutter V, Vanderhaeghen R, Hilson P, Hamill JD, Goossens A (2011) APETALA2/ETHYLENE RESPONSE FACTOR and basic helix–loop–helix tobacco transcription factors cooperatively mediate jasmonate‐elicited nicotine biosynthesis. The Plant Journal 66, 1053–1065.
| APETALA2/ETHYLENE RESPONSE FACTOR and basic helix–loop–helix tobacco transcription factors cooperatively mediate jasmonate‐elicited nicotine biosynthesis.Crossref | GoogleScholarGoogle Scholar | 21418355PubMed |
Edwards O, Ridsdill-Smith T, Berlandier F (2003) Aphids do not avoid resistance in Australian lupin (Lupinus angustifolius, L. luteus) varieties. Bulletin of Entomological Research 93, 403–411.
| Aphids do not avoid resistance in Australian lupin (Lupinus angustifolius, L. luteus) varieties.Crossref | GoogleScholarGoogle Scholar | 14641979PubMed |
Ellis C, Karafyllidis I, Turner JG (2002) Constitutive activation of jasmonate signaling in an Arabidopsis mutant correlates with enhanced resistance to Erysiphe cichoracearum, Pseudomonas syringae and Myzus persicae. Molecular Plant-Microbe Interactions 15, 1025–1030.
| Constitutive activation of jasmonate signaling in an Arabidopsis mutant correlates with enhanced resistance to Erysiphe cichoracearum, Pseudomonas syringae and Myzus persicae.Crossref | GoogleScholarGoogle Scholar | 12437300PubMed |
Farmer EE, Ryan CA (1990) Interplant communication: airborne methyl jasmonate induces synthesis of proteinase inhibitors in plant leaves. Proceedings of the National Academy of Sciences of the United States of America 87, 7713–7716.
| Interplant communication: airborne methyl jasmonate induces synthesis of proteinase inhibitors in plant leaves.Crossref | GoogleScholarGoogle Scholar | 11607107PubMed |
Frick KM, Kamphuis LG, Siddique KHM, Singh KB, Foley RC (2017) Quinolizidine alkaloid biosynthesis in lupins and prospects for grain quality improvement. Frontiers of Plant Science 8, 87
| Quinolizidine alkaloid biosynthesis in lupins and prospects for grain quality improvement.Crossref | GoogleScholarGoogle Scholar |
Frick KM, Foley RC, Kamphuis LG, Siddique KH, Garg G, Singh KB (2018) Characterisation of the genetic factors affecting quinolizidine alkaloid biosynthesis and its response to abiotic stress in narrow‐leafed lupin (Lupinus angustifolius L.) Plant, Cell & Environment 41, 2155–2168.
Gao L-L, Anderson JP, Klingler JP, Nair RM, Edwards OR, Singh KB (2007) Involvement of the octadecanoid pathway in bluegreen aphid resistance in Medicago truncatula. Molecular Plant-Microbe Interactions 20, 82–93.
| Involvement of the octadecanoid pathway in bluegreen aphid resistance in Medicago truncatula.Crossref | GoogleScholarGoogle Scholar | 17249425PubMed |
Gremigni P, Hamblin J, Harris D, Cowling W (2003) The interaction of phosphorus and potassium with seed alkaloid concentrations, yield and mineral content in narrow-leafed lupin (Lupinus angustifolius L.). Plant and Soil 253, 413–427.
| The interaction of phosphorus and potassium with seed alkaloid concentrations, yield and mineral content in narrow-leafed lupin (Lupinus angustifolius L.).Crossref | GoogleScholarGoogle Scholar |
Gurkok T, Turktas M, Parmaksiz I, Unver T (2015) Transcriptome profiling of alkaloid biosynthesis in elicitor induced opium poppy. Plant Molecular Biology Reporter 33, 673–688.
| Transcriptome profiling of alkaloid biosynthesis in elicitor induced opium poppy.Crossref | GoogleScholarGoogle Scholar |
Hane JK, Ming Y, Kamphuis LG, Nelson MN, Garg G, Atkins CA, Bayer PE, Bravo A, Bringans S, Cannon S, et al. (2017) A comprehensive draft genome sequence for lupin (Lupinus angustifolius), an emerging health food: insights into plant–microbe interactions and legume evolution. Plant Biotechnology Journal 15, 318–330.
| A comprehensive draft genome sequence for lupin (Lupinus angustifolius), an emerging health food: insights into plant–microbe interactions and legume evolution.Crossref | GoogleScholarGoogle Scholar | 27557478PubMed |
Harris DJ, Wilson PE (1988) A rapid manual method of lupin alkaloid analysis. In ‘Proceedings of the 5th international lupin conference’. (International Lupin Association: Poznan, Poland)
Hibi N, Higashiguchi S, Hashimoto T, Yamada Y (1994) Gene expression in tobacco low-nicotine mutants. The Plant Cell 6, 723–735.
| Gene expression in tobacco low-nicotine mutants.Crossref | GoogleScholarGoogle Scholar | 8038607PubMed |
Howe GA, Jander G (2008) Plant immunity to insect herbivores. Annual Review of Plant Biology 59, 41–66.
| Plant immunity to insect herbivores.Crossref | GoogleScholarGoogle Scholar | 18031220PubMed |
Huang F-C, Kutchan TM (2000) Distribution of morphinan and benzo[c]phenanthridine alkaloid gene transcript accumulation in Papaver somniferum. Phytochemistry 53, 555–564.
| Distribution of morphinan and benzo[c]phenanthridine alkaloid gene transcript accumulation in Papaver somniferum.Crossref | GoogleScholarGoogle Scholar | 10724180PubMed |
Jansen G, Jürgens HU, Ordon F (2009) Effects of temperature on the alkaloid content of seeds of Lupinus angustifolius cultivars. Journal Agronomy & Crop Science 195, 172–177.
| Effects of temperature on the alkaloid content of seeds of Lupinus angustifolius cultivars.Crossref | GoogleScholarGoogle Scholar |
Jansen G, Jürgens H-U, Schliephake E, Ordon F (2012) Effect of the soil pH on the alkaloid content of Lupinus angustifolius. International Journal of Agronomy 2012, 269878
| Effect of the soil pH on the alkaloid content of Lupinus angustifolius.Crossref | GoogleScholarGoogle Scholar |
Johnson ND, Rigney LP, Bentley BL (1989) Short-term induction of alkaloid production in lupines. Differences between N2-fixing and nitrogen-limited plants. Journal of Chemical Ecology 15, 2425–2434.
| Short-term induction of alkaloid production in lupines. Differences between N2-fixing and nitrogen-limited plants.Crossref | GoogleScholarGoogle Scholar | 24271539PubMed |
Kamphuis LG, Hane JK, Nelson MN, Gao L, Atkins CA, Singh KB (2015) Transcriptome sequencing of different narrow‐leafed lupin tissue types provides a comprehensive uni‐gene assembly and extensive gene‐based molecular markers. Plant Biotechnology Journal 13, 14–25.
| Transcriptome sequencing of different narrow‐leafed lupin tissue types provides a comprehensive uni‐gene assembly and extensive gene‐based molecular markers.Crossref | GoogleScholarGoogle Scholar | 25060816PubMed |
Kato K, Shitan N, Shoji T, Hashimoto T (2015) Tobacco NUP1 transports both tobacco alkaloids and vitamin B6. Phytochemistry 113, 33–40.
| Tobacco NUP1 transports both tobacco alkaloids and vitamin B6.Crossref | GoogleScholarGoogle Scholar | 24947336PubMed |
Khan MB, Harborne JB (1990) Induced alkaloid defence in Atropa acuminata in response to mechanical and herbivore leaf damage. Chemoecology 1, 77–80.
| Induced alkaloid defence in Atropa acuminata in response to mechanical and herbivore leaf damage.Crossref | GoogleScholarGoogle Scholar |
Lee MJ, Pate JS, Harris DJ, Atkins CA (2007) Synthesis, transport and accumulation of quinolizidine alkaloids in Lupinus albus L. and L. angustifolius L. Journal of Experimental Botany 58, 935–946.
| Synthesis, transport and accumulation of quinolizidine alkaloids in Lupinus albus L. and L. angustifolius L.Crossref | GoogleScholarGoogle Scholar | 17189595PubMed |
León J, Rojo E, Sánchez‐Serrano JJ (2001) Wound signalling in plants. Journal of Experimental Botany 52, 1–9.
| Wound signalling in plants.Crossref | GoogleScholarGoogle Scholar | 11181708PubMed |
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 25, 402–408.
| Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method.Crossref | GoogleScholarGoogle Scholar | 11846609PubMed |
Memelink J, Verpoorte R, Kijne JW (2001) ORCAnization of jasmonate-responsive gene expression in alkaloid metabolism. Trends in Plant Science 6, 212–219.
| ORCAnization of jasmonate-responsive gene expression in alkaloid metabolism.Crossref | GoogleScholarGoogle Scholar | 11335174PubMed |
Okada T, Hirai MY, Suzuki H, Yamazaki M, Saito K (2005) Molecular characterization of a novel quinolizidine alkaloid O-Tigloyltransferase: cDNA cloning, catalytic activity of recombinant protein and expression analysis in Lupinus plants. Plant & Cell Physiology 46, 233–244.
| Molecular characterization of a novel quinolizidine alkaloid O-Tigloyltransferase: cDNA cloning, catalytic activity of recombinant protein and expression analysis in Lupinus plants.Crossref | GoogleScholarGoogle Scholar |
Philippi J, Schliephake E, Jürgens HU, Jansen G, Ordon F (2015) Feeding behavior of aphids on narrow‐leafed lupin (Lupinus angustifolius) genotypes varying in the content of quinolizidine alkaloids. Entomologia Experimentalis et Applicata 156, 37–51.
| Feeding behavior of aphids on narrow‐leafed lupin (Lupinus angustifolius) genotypes varying in the content of quinolizidine alkaloids.Crossref | GoogleScholarGoogle Scholar |
Pilar Vilariño M, Mareggiani G, Grass MY, Leicach S, Ravetta D (2005) Post‐damage alkaloid concentration in sweet and bitter lupin varieties and its effect on subsequent herbivory. Journal of Applied Entomology 129, 233–238.
| Post‐damage alkaloid concentration in sweet and bitter lupin varieties and its effect on subsequent herbivory.Crossref | GoogleScholarGoogle Scholar |
Priddis CR (1983) Capillary gas chromatography of lupin alkaloids. Journal of Chromatography. A 261, 95–101.
| Capillary gas chromatography of lupin alkaloids.Crossref | GoogleScholarGoogle Scholar |
Ryan SM, DeBoer KD, Hamill JD (2015) Alkaloid production and capacity for methyljasmonate induction by hairy roots of two species in Tribe Anthocercideae, family Solanaceae. Functional Plant Biology 42, 792–801.
| Alkaloid production and capacity for methyljasmonate induction by hairy roots of two species in Tribe Anthocercideae, family Solanaceae.Crossref | GoogleScholarGoogle Scholar |
Sandow J (1987) Lupins, aphids and viruses: a complex problem. Journal of the Department of Agriculture, Western Australia, Series 4 28, 5–59.
Shoji T, Nakajima K, Hashimoto T (2000) Ethylene suppresses jasmonate-induced gene expression in nicotine biosynthesis. Plant & Cell Physiology 41, 1072–1076.
| Ethylene suppresses jasmonate-induced gene expression in nicotine biosynthesis.Crossref | GoogleScholarGoogle Scholar |
Shoji T, Inai K, Yazaki Y, Sato Y, Takase H, Shitan N, Yazaki K, Goto Y, Toyooka K, Matsuoka K (2009) Multidrug and toxic compound extrusion-type transporters implicated in vacuolar sequestration of nicotine in tobacco roots. Plant Physiology 149, 708–718.
| Multidrug and toxic compound extrusion-type transporters implicated in vacuolar sequestration of nicotine in tobacco roots.Crossref | GoogleScholarGoogle Scholar | 19098091PubMed |
Shoji T, Kajikawa M, Hashimoto T (2010) Clustered transcription factor genes regulate nicotine biosynthesis in tobacco. The Plant Cell 22, 3390–3409.
| Clustered transcription factor genes regulate nicotine biosynthesis in tobacco.Crossref | GoogleScholarGoogle Scholar | 20959558PubMed |
von Sengbusch R (1942) Sweet lupins and oil lupins. The history of the origin of some new crop plants. Landwirtschaftliche Jahrbücher 91, 719–880.
Wasternack C (2007) Jasmonates: an update on biosynthesis, signal transduction and action in plant stress response, growth and development. Annals of Botany 100, 681–697.
| Jasmonates: an update on biosynthesis, signal transduction and action in plant stress response, growth and development.Crossref | GoogleScholarGoogle Scholar | 17513307PubMed |
Wasternack C, Hause B (2013) Jasmonates: biosynthesis, perception, signal transduction and action in plant stress response, growth and development. An update to the 2007 review in Annals of Botany. Annals of Botany 111, 1021–1058.
| Jasmonates: biosynthesis, perception, signal transduction and action in plant stress response, growth and development. An update to the 2007 review in Annals of Botany.Crossref | GoogleScholarGoogle Scholar | 23558912PubMed |
Wink M (1983) Wounding-induced increase of quinolizidine alkaloid accumulation in lupin leaves. Zeitschrift für Naturforschung. Section C. Biosciences 38, 905–909.
| Wounding-induced increase of quinolizidine alkaloid accumulation in lupin leaves.Crossref | GoogleScholarGoogle Scholar |
Wink M (1987) Site of lupanine and sparteine biosynthesis in intact plants and in vitro organ cultures. Zeitschrift für Naturforschung. Section C. Biosciences 42, 868–872.
| Site of lupanine and sparteine biosynthesis in intact plants and in vitro organ cultures.Crossref | GoogleScholarGoogle Scholar |
Wink M (1992) The role of quinolizidine alkaloids in plant-insect interactions. Insect-Plant Interactions 4, 131–166.
Wink M, Witte L (1991) Storage of quinolizidine alkaloids in Macrosiphum albifrons and Aphis genistae (Homoptera: Aphididae). Entomologia Generalis 15, 237–254.
| Storage of quinolizidine alkaloids in Macrosiphum albifrons and Aphis genistae (Homoptera: Aphididae).Crossref | GoogleScholarGoogle Scholar |
Winz RA, Baldwin IT (2001) Molecular interactions between the specialist herbivore Manduca sexta (Lepidoptera, Sphingidae) and its natural host Nicotiana attenuata. IV. Insect-induced ethylene reduces jasmonate-induced nicotine accumulation by regulating putrescine N-methyltransferase transcripts. Plant Physiology 125, 2189–2202.
| Molecular interactions between the specialist herbivore Manduca sexta (Lepidoptera, Sphingidae) and its natural host Nicotiana attenuata. IV. Insect-induced ethylene reduces jasmonate-induced nicotine accumulation by regulating putrescine N-methyltransferase transcripts.Crossref | GoogleScholarGoogle Scholar | 11299398PubMed |
Yang T, Nagy I, Mancinotti D, Otterbach SL, Andersen TB, Motawia MS, Asp T, Geu-Flores F (2017a) Transcript profiling of a bitter variety of narrow-leafed lupin to discover alkaloid biosynthetic genes. Journal of Experimental Botany 68, 5527–5537.
| Transcript profiling of a bitter variety of narrow-leafed lupin to discover alkaloid biosynthetic genes.Crossref | GoogleScholarGoogle Scholar | 29155974PubMed |
Yang Y, Yan P, Yi C, Li W, Chai Y, Fei L, Gao P, Zhao H, Wang Y, Timko MP (2017b) Transcriptome-wide analysis of jasmonate-treated BY-2 cells reveals new transcriptional regulators associated with alkaloid formation in tobacco. Journal of Plant Physiology 215, 1–10.
| Transcriptome-wide analysis of jasmonate-treated BY-2 cells reveals new transcriptional regulators associated with alkaloid formation in tobacco.Crossref | GoogleScholarGoogle Scholar | 28527333PubMed |
Zhao J, Davis LC, Verpoorte R (2005) Elicitor signal transduction leading to production of plant secondary metabolites. Biotechnology Advances 23, 283–333.
| Elicitor signal transduction leading to production of plant secondary metabolites.Crossref | GoogleScholarGoogle Scholar | 15848039PubMed |