

An assessment of dingo ancestry in camp dogs in Western Australia
T. L. Kreplins A * , D. Stephens B , N. Laing C and C. Oskam D
A * , D. Stephens B , N. Laing C and C. Oskam D
A
B
C
D
Abstract
Hybridisation between Australian dingoes and domestic dogs is a controversial area of interest and research. An ongoing canine sterilisation programme in rural and remote Western Australia provided an opportunity to assess the dingo ancestry of camp dogs and opportunities for hybridisation. Blood samples were collected from 345 individual community dogs at 21 locations. Dogs were screened using 23 microsatellite loci and ancestry percentage assigned using an iterative Bayesian assignment algorithm. A single individual was a dingo, 96% were domestic dogs and 3.5% were hybrids. Camp dog and dingo hybridisation in these areas is of little concern in terms of conserving dingo purity.
Keywords: Australian dingoes (Canis familiaris), camp dog, dingo, domestic dog, hybridisation, Western Australia.
Introduction
Dogs are one of the most widespread domesticated species and are found on all continents (Balme and O’Connor 2016) except Antarctica. Australian dingoes (Canis familiaris) (Jackson et al. 2017) are adaptable wild-living canids that are found across the breadth of Australian landscapes, from deserts to alpine, forests to peri-urban. Dingoes have been present in Australian landscapes for approximately 4500 years (Balme et al. 2018; Zhang et al. 2020). Surveys of dingoes by DNA across the continent have revealed that few domestic dogs live away from urban settlements, and even dingo–dog hybrids with low dingo ancestry seem rare (Stephens et al. 2015). Despite being a single species, dingoes and domestic dogs differ in a few phenotypic characteristics. The proposed explanations for this finding are that behavioural and/or social factors make domestic dogs unsuited to life without humans, either through an inability to survive directly, or inhibition/hostility from dingoes (Wilton et al. 1998). Dingoes are valued culturally (Hytten 2009) and conservation of their purity is valued in some parts of the Australian community (van Eeden et al. 2017). There is an alternate view that dingoes are their own species (Smith et al. 2019), but this has recently been questioned (see Supplementary material; Jackson et al. 2021).
A major concern of dingo conservation is the hybridisation with domestic dogs (Stephens et al. 2015; Cairns et al. 2020). ‘Camp-dogs’ are populations of domestic but often free-living dogs residing in and around remote or regional human settlements. These camp dogs exploit human resources and have close associations with the human residents (Smith and Litchfield 2005; Newsome et al. 2014). In previous studies where camp dogs have been included in surveys of dingo ancestry (Newsome et al. 2013; Bombara et al. 2017), the camp dogs have shown low percentages of dingo ancestry, both within individuals and in the population as a whole.
Understanding the relationship between dingoes and free-roaming dogs will provide knowledge around the potential animal and human health risks. Free-roaming dogs residing in and around human settlement not only pose a genetic risk to dingoes but can also introduce parasite and disease risks (Barker et al. 2012; King et al. 2012). Many of the free-roaming dogs have poor health due to lack of veterinary support and resources in remote areas (Sparkes et al. 2014). This pathogen risk can also be transferred to livestock, native species, and even zoonotic risks to humans.
Here we surveyed 21 sites with 1–70 individuals per site (345 total individuals) where there is coexistence between humans and semiowned camp dogs. We aimed to identify the percentage of free-roaming camp dogs with dingo ancestry or identify pure dingoes at the sites.
Methods
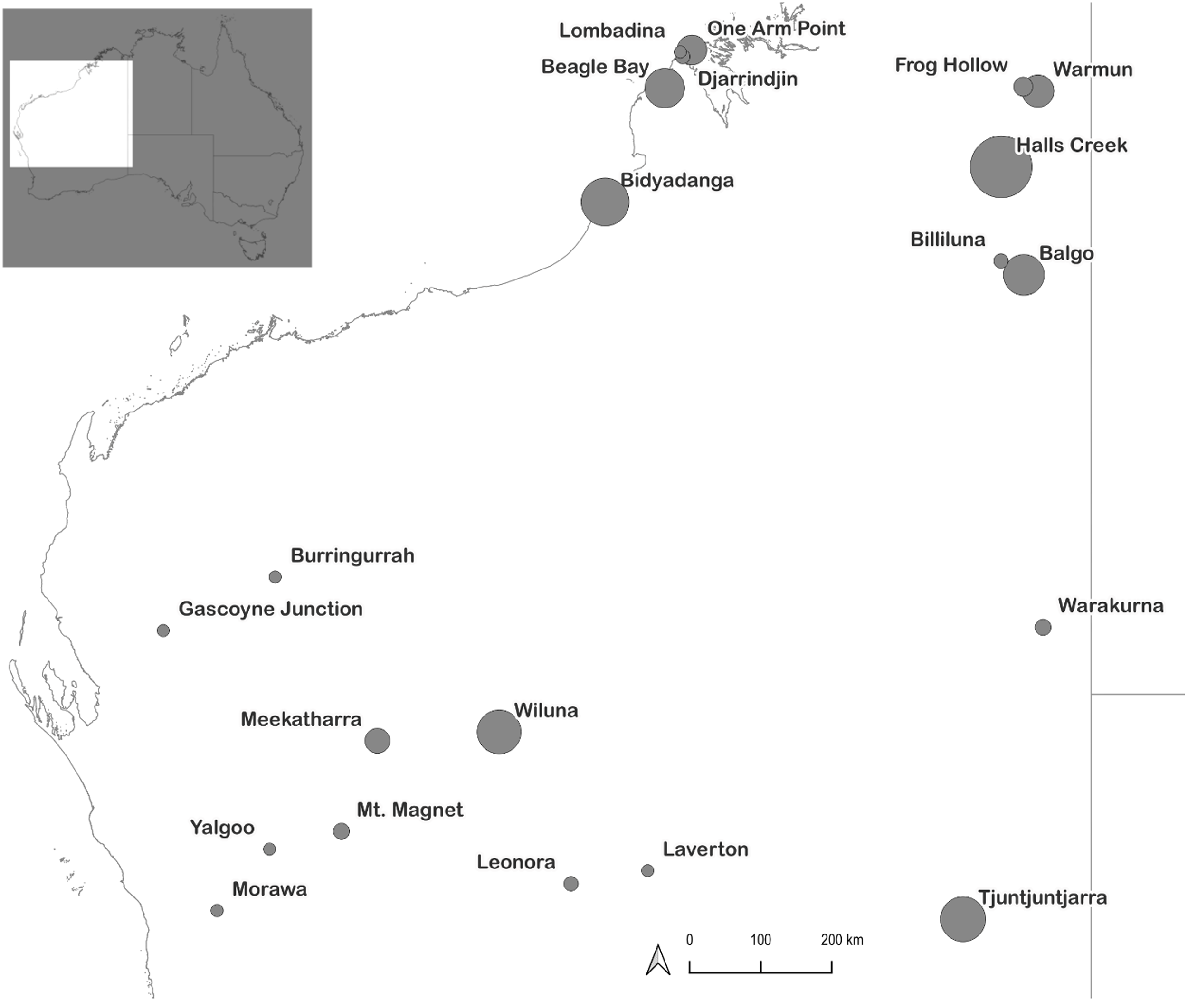
The camp dogs were part of a canine sterilisation programme in which they were health screened, sterilised and treated for illness by Murdoch University veterinarians in training at a range of communities across Western Australia (Fig. 1).
Locations where dogs were sampled for this study within Western Australia. Grey dots are scaled by the number of animals collected at each site. The inset shows location of the study area within Australia.
DNA was extracted from 200 μL of each blood sample using the Qiagen QIAMP mini blood kit. Samples were processed according to the recommended protocol, with 150 μL final elution. Extracted DNA was amplified by PCR using the following conditions: 15 min at 95°C, 35 cycles of 30 s at 94°C, 90 s at 55°C and 60 s at 72°C, then 30 min final extension at 60°C at 23 microsatellite loci in five multiplexes. PCR primers used were as described in Stephens (2015; table 1). PCR products were visualised using an Applied Biosystems 3730 at the Australian Genome Research Facility (AGRF, Adelaide, Australia). Data from the amplified fragments were analysed using Structure ver. 2.3.4 (Pritchard et al. 2000) along with 322 reference dingoes and 109 domestic dogs, with allele frequencies updating and initialising from these reference populations. The selection of these reference individuals is detailed in Stephens et al. (2015). The number of populations was fixed at K = 2 for 10 replicates to produce a percentage genotype for each individual assigned to either the dingo or domestic dog ancestral group. Replicates were combined and averaged using pophelper (Francis 2017; R Core Team 2023). Stephens et al. (2015, 2022, 2023) has published many times using the stricter dingo purity definitions; using an error of ±10% based on this previous work was used to assign individuals to categories of dingo (>90% dingo ancestry), hybrid (11–89% dingo ancestry) or domestic dog (<10% dingo ancestry) for comparison between sites. This margin of error also covers the mean difference between microsatellite and single nucleotide polymorphism (SNP) testing in the study area (9%) as demonstrated by Cairns et al. (2023: table 5). Principal coordinates analysis was performed for comparison with the Structure results, and was calculated using GenAlEx ver. 6.51b2 (Peakall and Smouse 2012).
Results
A genotype was obtained from all of the 345 animals tested, and only one was classified as a dingo, with 94% dingo ancestry (Table 1). There were 12 dingo–dog hybrids with mostly domestic dog ancestry, but the majority of individuals (96%) were domestic dogs. Of the hybrid animals, all had less than 50% dingo ancestry, and nine of the 12 were less than 25% dingo ancestry (range 11–43%).
| Location | Dingo | Hybrid | Dog | |
|---|---|---|---|---|
| Balgo | 0 | 0 | 32 | |
| Beagle Bay | 0 | 1 | 30 | |
| Bidyadanga | 0 | 0 | 44 | |
| Billiluna | 0 | 1 | 3 | |
| Burringurrah | 0 | 0 | 2 | |
| Djarrindjin | 0 | 1 | 3 | |
| Frog Hollow | 0 | 0 | 7 | |
| Gascoyne Junction | 0 | 0 | 1 | |
| Halls Creek | 0 | 3 | 70 | |
| Warmun | 0 | 0 | 20 | |
| Laverton | 0 | 0 | 1 | |
| Leonora | 0 | 0 | 4 | |
| Lombadina | 0 | 0 | 2 | |
| Meekatharra | 0 | 0 | 12 | |
| Morawa | 0 | 0 | 3 | |
| Mount Magnet | 0 | 1 | 4 | |
| One Arm Point | 1 | 0 | 17 | |
| Tjuntjuntjarra | 0 | 3 | 36 | |
| Warakurna | 0 | 0 | 5 | |
| Wiluna | 0 | 2 | 35 | |
| Yalgoo | 0 | 0 | 1 | |
| Total | 1 | 12 | 332 | |
| Percentage | 0.3 | 3.5 | 96.2 |
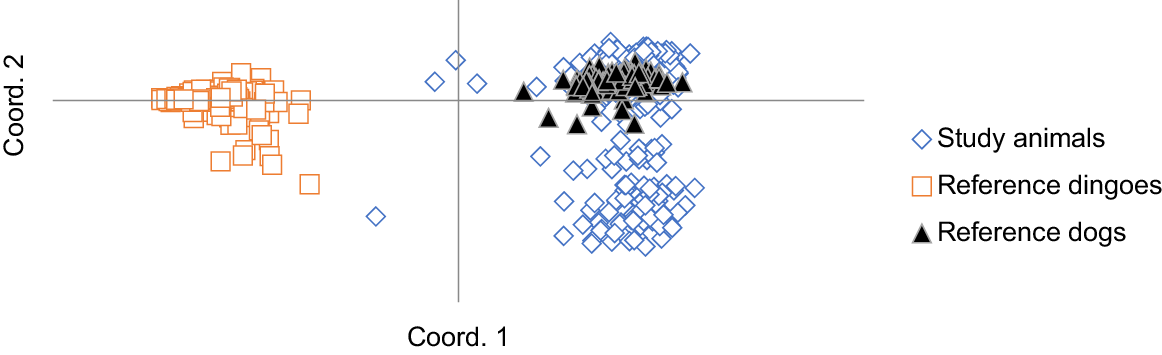
Separation of the study animals from the reference dingoes in the PCoA also indicates the lack of hybridisation between dingoes and community dogs (Fig. 2).
Discussion
How ‘wild’ and genetically pure dingoes are, is under constant debate. Dingoes most likely arose in Asia around 5000–12,000 years ago (Filios and Tacon 2016) through either descent from Asian wolves or hybridisation with Asian wolves, although the literature is unclear on this point (Corbett 2001; Savolainen et al. 2004). After arriving with humans, dingoes quickly spread across Australia and their relationships with dogs throughout the continent is becoming better understood.
Dingo purity, and in turn their conservation, is only at risk if camp dogs and dingoes intermix freely (Elledge et al. 2008). A reduction in dingo purity is unlikely if the dingoes and camp dogs do not intermix. The pattern of domestic dogs being largely absent from wild landscapes seems to exist in the inverse in human settlements. Dingoes are almost completely absent from the settlements in this study (except Allen et al. 2013), indicating that either the dingoes are not favoured by the humans present, or the dingoes themselves do not choose to remain in the settlements. Domestic dogs, on the other hand, seem able to persist in these areas as long as there is some assistance from humans or human resources, even without direct ownership.
Transmission of disease or pathogens could potentially be very high between all canids. Contact rates between free-roaming dogs of Indigenous communities in Northern Australia (of unknown ancestry) were 8–147 direct contacts (i.e. licking, mouthing, playing) between dogs within 24 h. Often this occurred in urban settings but also in the bush and beach lands (Bombara et al. 2017). However, if there is minimal hybridisation or socialisation between camp dogs and dingoes, as recorded from the lack of hybridised individuals, transmission of disease and pathogens is low. Parasites are known to be common among free-roaming camp dogs of Indigenous communities in Australia, in particular ectoparasites such as ticks (Newsome et al. 2013; Bombara et al. 2017). However, anecdotally, dingoes are not parasitised by ticks (Koungoulos and Contos 2019).
Other studies in Australia have found variable results in relation to roaming dogs of a range of ancestry. For example, a study on hair samples found approximately one-third of dogs were classified as hybrids (Bombara et al. 2017). However, the percentage of the samples differed between the regions: 41% for Galinwin’ku and 6% for Northern Peninsula Area. These dogs were also tracked and were demonstrated to stray into nearby bushland and may potentially interact with dingoes. Another study comparing the ancestry of dogs of mine sites and neighbouring communities found that 89% of samples from the mine were dingoes and there was minimal dingo ancestry in the community dogs. Only one of the studied communities showed the potential for interactions between community and dingoes (Newsome et al. 2013).
The next step in this research is to understand why there is a lack of hybridisation. Is the lack of intermixing between camp dogs and dingoes by choice or because camp dogs never encounter dingoes? Is there anything occurring differently in the Kimberely region compared to the Nullarbor region leading to a few more hybrids? Or is this an artefact of sampling (i.e. dogs that can be handled for sterilisation)? In other words, do the community dogs remain within the community bounds or do they enter the bushland where dingoes reside and choose to not interact? Studies that have investigated urban roaming domestic dogs indicate that each individual dog has a preference for roaming or not roaming (Meek 1999). However, domestic dogs that did roam made similar choices to dingoes and were active along waterways that had high prey activity (i.e. kangaroos). Is the ‘roaming’ dog moving further because of food availability regardless of its ancestry? Furthermore, is the persecution of dingoes in Western Australia for their predation of livestock creating a learned aversion to human settlements (Stephens et al. 2023)?
While dingoes are of high value to some Australians, camp dogs themselves are of high cultural importance to community members (Hytten 2009; Balme and O’Connor 2016). Although the companionship between dogs and humans covers 30,000 years, with dogs being in Australia for only approximately 4500 years, their significance is cherished by the entire Australian community, regardless of their ancestry (Filios and Tacon 2016). This study adds to previous research showing a lack of hybridisation occurring, which indicates that dingo purity is not of conservation concern at the present time in Western Australia and possible disease transmission pathways are minimal.
Declaration of funding
This work was funded by the Western Australia Wild Dog Action Plan 2016–2021 and 2021–2025.
Acknowledgements
Many thanks to the final year veterinarian students and staff from Murdoch University who collected blood samples during the outreach program for community dogs.
References
Allen BL, Goullet M, Allen LR, Lisle A, Leung LK-P (2013) Dingoes at the doorstep: preliminary data on the ecology of dingoes in urban areas. Landscape and Urban Planning 119, 131-135.
| Crossref | Google Scholar |
Balme J, O’Connor S (2016) Dingoes and Aboriginal social organization in Holocene Australia. Journal of Archaeological Science: Reports 7, 775-781.
| Crossref | Google Scholar |
Balme J, O’Connor S, Fallon S (2018) New dates on dingo bones from Madura Cave provide oldest firm evidence for arrival of the species in Australia. Nature Scientific Reports 8, 9933.
| Crossref | Google Scholar | PubMed |
Barker EN, Langton DA, Helps CR, Brown G, Malik R, Shaw SE, Tasker S (2012) Haemoparasites of free-roaming dogs associated with several remote Aboriginal communities in Australia. BMC Veterinary Research 8, 55.
| Crossref | Google Scholar | PubMed |
Bombara C, Durr S, Gongora J, Ward MP (2017) Roaming of dogs in remote Indigenous communities in northern Australia and potential interaction between community and wild dogs. Australian Veterinary Journal 95, 182-188.
| Crossref | Google Scholar | PubMed |
Cairns KM, Nesbitt BJ, Laffan SW, Letnic M, Crowther MS (2020) Geographic hot spots of dingo genetic ancestry in southeastern Australia despite hybridisation with domestic dogs. Conservation Genetics 21, 77-90.
| Crossref | Google Scholar |
Cairns KM, Crowther MS, Parker HG, Ostrander EA, Letnic M (2023) Genome-wide variant analyses reveal new patterns of admixture and population structure in Australian dingoes. Molecular Ecology 32, 4133-4150.
| Crossref | Google Scholar | PubMed |
Elledge AE, Allen LR, Carlsson B-L, Wilton AN, Leung LK-P (2008) An evaluation of genetic analyses, skull morphology and visual appearance for assessing dingo purity: implications for dingo conservation. Wildlife Research 35, 812-820.
| Crossref | Google Scholar |
Filios M, Tacon PSC (2016) Who let the dogs in? A review of the recent genetic evidence for the introduction of the dingo to Australia and implications for the movement of people. Journal of Archaeological Science: Reports 7, 782-792.
| Crossref | Google Scholar |
Francis RM (2017) pophelper: an R package and web app to analyse and visualize population structure. Molecular Ecology Resources 17, 27-32.
| Crossref | Google Scholar |
Hytten K (2009) Dingo dualisms: exploring the ambiguous identity of Australian dingoes. Australian Zoologist 35, 18-27.
| Crossref | Google Scholar |
Jackson SM, Groves CP, Fleming PJS, Aplin KP, Eldridge MDB, Gonzalez A, Helgen KM (2017) The Wayward Dog: is the Australian native dog or Dingo a distinct species? Zootaxa 4317, 201-224.
| Crossref | Google Scholar |
Jackson SM, Fleming PJ, Eldridge MD, Archer M, Ingleby S, Johnson RN, Helgen KM (2021) Taxonomy of the dingo: it’s an ancient dog. Australian Zoologist 41, 347-357.
| Crossref | Google Scholar |
King JS, Brown GK, Jenkins DJ, Ellis JT, Fleming PJS, Windsor PA, Slapeta J (2012) Oocysts and high seroprevalence of Neospora caninum in dogs living in remote Aboriginal communities and wild dogs in Australia. Veterinary Parasitology 187, 85-92.
| Crossref | Google Scholar | PubMed |
Koungoulos L, Contos P (2019) Global expansion of the Australian biting louse Heterodoxus spiniger facilitated by human transport of dog (Canis familiaris), and implications for prehistoric cultural interaction in Australasia. Environmental Archaeology 26, 281-294.
| Crossref | Google Scholar |
Meek PD (1999) The movement, roaming behaviour and home range of free-roaming domestic dogs, Canis lupus familiaris, in coastal New South Wales. Wildlife Research 26, 847-855.
| Crossref | Google Scholar |
Newsome TM, Stephens D, Ballard G-A, Dickman CR, Fleming PJS (2013) Genetic profile of dingoes (Canis lupus dingo) and free-roaming domestic dogs (C. l. familiaris) in the Tanami Desert, Australia. Wildlife Research 40, 196-206.
| Crossref | Google Scholar |
Newsome TM, Ballard G-A, Crowther MS, Fleming PJS, Dickman CR (2014) Dietary niche overlap of free-roaming dingoes and domestic dogs: the role of human-provided food. Journal of Mammalogy 95, 392-403.
| Crossref | Google Scholar |
Peakall R, Smouse PE (2012) GenAlEx 6.5: genetic analysis in Excel. Population genetic software for teaching and research – an update. Bioinformatics 28, 2537-2539.
| Crossref | Google Scholar | PubMed |
Pritchard JK, Stephens M, Donnelly P (2000) Inference of population structure using multilocus genotype data. Genetics 155, 945-959.
| Crossref | Google Scholar | PubMed |
R Core Team (2023) ‘R: a language and environment for statistical computing.’ (R Foundation for Statistical Computing: Vienna, Austria) Available at https://www.R-project.org/
Savolainen P, Leitner T, Wilton AN, Matisoo-Smith E, Lundeberg J (2004) A detailed picture of the origin of the Australian dingo, obtained from the study of mitochondrial DNA. Proceedings of the National Academy of Sciences of the United States of America 101, 12387-12390.
| Crossref | Google Scholar |
Smith BP, Litchfield CA (2005) A review of the relationship between indigenous Australians, dingoes (Canis dingo) and domestic dogs (Canis familiaris). Anthrozoos 22, 111-128.
| Crossref | Google Scholar |
Smith BP, Cairns KM, Adams JW, Newsome TM, Fillios M, Deaux EC, Parr WC, Letnic M, Van Eeden LM, Appleby RG, Bradshaw CJA, Savolainen P, Ritchie EG, Nimmo DG, Archer-Lean C, Greenville AC, Dickman CR, Watson L, Moseby KE, Doherty TS, Wallach AD, Morrant DS, Crowther MS (2019) Taxonomic status of the Australian dingo: the case for Canis dingo Meyer, 1793. Zootaxa 4564, 173-197.
| Crossref | Google Scholar |
Sparkes J, Fleming PJS, Ballard G, Scott-Orr H, Durr S, Ward MP (2014) Canine rabies in Australia: a review of preparedness and research needs. Zoonoses and Public Health 62, 237-253.
| Crossref | Google Scholar |
Stephens D, Wilton AN, Fleming PJS, Berry O (2015) Death by sex in an Australian icon: a continent-wide survey reveals extensive hybridization between dingoes and domestic dogs. Molecular Ecology 24, 5643-5656.
| Crossref | Google Scholar | PubMed |
Stephens D, Fleming PJS, Sawyers E, Mayr TP (2022) An isolated population reveals greater genetic structuring of the Australian dingo. Scientific Reports 12, 19105.
| Crossref | Google Scholar | PubMed |
Stephens D, Kennedy MS, Kreplins TL (2023) Stable dingo population structure and purity over 11 years of lethal management. Wildlife Research 50(12), 980-992.
| Crossref | Google Scholar |
van Eeden LM, Dickman CR, Ritchie EG, Newsome TM (2017) Shifting public values and what they mean for increasing democracy in wildlife management decisions. Biodiversity and Conservation 26, 2759-2763.
| Crossref | Google Scholar |
Wilton AN, Steward DJ, Zafiris K (1998) Microsatellite variation in the Australian dingo. The American Genetic Association 90, 108-111.
| Crossref | Google Scholar |
Zhang S-J, Wang G-D, Ma P, Zhang L-L, Yin T-T, Liu Y-H, Otecko NO, Wang M, Ma Y-P, Wang L, Mao B, Savolainen P, Zhang Y-P (2020) Genomic regions under selection in the feralization of the dingoes. Nature Communications 11, 671.
| Crossref | Google Scholar | PubMed |