

Integrating western and Indigenous knowledge to identify habitat suitability and survey for the white-throated grasswren (Amytornis woodwardi) in the Arnhem Plateau, Northern Territory, Australia
Kelly M. Dixon A B * , Brenton von Takach C , Brittany Hayward-Brown A , Terrah Guymala D ,
A B * , Brenton von Takach C , Brittany Hayward-Brown A , Terrah Guymala D , A
B
C
D
E
F
G
H
I
Abstract
Many Australian threatened species occur on Indigenous-owned and/or managed lands, often in vast, remote areas that are difficult and expensive to access. One such species is the white-throated grasswren (WTGW, Amytornis woodwardi), a rare ground-dwelling bird found in rocky spinifex-covered escarpment habitats of northern Australia. To make surveying rare species more tractable, we can predict habitat suitability by associating occurrence points with environmental covariates that may influence the species’ distribution.
Here, we combine western and Indigenous knowledge and approaches to better quantify the habitat associations and distribution of the WTGW.
We modelled habitat suitability across the region using historical occurrence records and applicable environmental variables with input from Traditional ecological knowledge. We then used this habitat-suitability map as a visual tool for participatory mapping and planning sessions with Traditional Custodians to select on-ground survey sites. Collaborative surveys were then undertaken to target WTGWs at 39 sites across the Arnhem Plateau by using several methods, including bioacoustic audio recorders (BARs), call-playback (CPB) surveys, and motion-detection cameras.
Collaboration between Traditional Custodians and scientists at all stages helped make this project a success. Our model suggests that WTGWs typically occupy habitat patches that have lower distance-to-unburnt (fire extent) values, lower proportion-of-area-burnt values, lower vegetation-cover values, and higher time-since-fire values. On-ground surveys detected WTGWs at six sites with BARs and at one of these six sites with CPB and camera-trapping, suggesting that BARs were the most effective detection method.
Our results provided key ecological information for use by land managers in the region and highlighted the importance of effective fire management for the persistence of WTGW populations. The success of the cross-cultural collaboration across several Indigenous organisations relied on the expertise of Traditional Custodians and Indigenous rangers. With Traditional Custodians and Indigenous rangers leading the fieldwork, co-benefits of the program included connecting people with Country and supporting the transfer of intergenerational knowledge surrounding the WTGW.
Whereas fire management in the region over the past decade has led to broadscale reductions in the frequency, extent and intensity of fires, strategic imposition of fire regimes that retain sufficient unburnt refugia at habitat scales appears necessary for viable populations of species such as the WTGW to persist.
Keywords: Amytornis, applied ecology, Arnhem Land, conservation management, cross-cultural, ecological niche, grasswren, indigenous knowledge, modelling.
| Short summary and translation |
| Here, we integrate western and Indigenous knowledge to improve our understanding of white-throated grasswren (WTGW) distribution and habitat requirements through habitat suitability modelling and on-ground surveys. We detected WTGWs at 6 of 39 sites. Indigenous-led fieldwork helped connect people with Country, supported knowledge exchange and active discussions around fire management to protect the species. |
| Bininj kunwok: Ngarridjare ngarrinan kore baleh nawu mayhmayh, kangeyyo yilingkirrkkirr, kabirrini dja kabirriyo, mak kore baleh kabirriredmarnbun. Ngad Bininj dja Balanda wanjh karrirawon kunmayali ba bu karriwernhburrbun kore baleh kunred nawu yilingkirrkkirr mayhmayh kabirridi dja kabirriyo. |
| Ngad rangers ngarridjare ngarriborlbme ba ngarriwernhburrbun bu ngarrinahnan nahni mayhmayh yilingkirrkkirr ba kabirridarrkiddi munguyh. Ngarri-awani nawu yilingkirrkkirr mayhmayh. Wanjh ngarringalkeng kore six kunred kabirrihdi. |
| Wanjh bolkkime ngarriburrbun nawu yilingkirrkkirr mayhmayh kabirrihdi kore minj karralkrung munguyh. Bu mahni ngarridurrkmirri, wanjh kamak ba bu ngarridurndeng kore kunred ngadberre, karringurlkmarren kunred dja mayhmayh mak mayh. Dja mak Balanda dja ngad nawu Bininj karrimarneborlbmerren kore karridjarrkdurrkmirri. Dja wanjh karriwokdi karrikarremarnbun bu baleh karriyimuwon bu kunak karriwurlhke, ba bu minj mayh karribun dja kabirridjaldarrkiddi munguyh. |
| The summary spoken in Bininj kunwok is available here. |
Introduction
Indigenous peoples and their traditional knowledge and governance systems play a crucial role in biodiversity conservation globally (Dawson et al. 2021). In Australia, ~39% of the continent is owned and/or managed by Indigenous peoples (Jacobsen et al. 2020). Almost three quarters of federally listed threatened species are likely to occur on these lands (Renwick et al. 2017) and the entire range of 14 imperilled bird taxa exist solely on these lands (Lilleyman et al. 2024). Conservation practices are shifting to recognise Indigenous stewardship and governance to care for biocultural values (Austin et al. 2018; Garnett et al. 2018) and the critical role Indigenous peoples’ involvement has in solving the biodiversity crisis (UNEP 2022). In Australia, collaborations between Traditional Custodians, local Indigenous rangers, and conservation scientists have grown over the past decade to identify and monitor threatened species and threatening processes, and better guide management practices (McKemey et al. 2021; Ens and Turpin 2022; Skroblin et al. 2022). Support and respect for Traditional knowledge has been demonstrated to have a direct impact on the overall success of cross-cultural collaborations and, particularly, the efficacy of threatened species projects (Ward-Fear and Shine 2019; Wysong et al. 2022).
Species of conservation concern can be difficult to survey and monitor adequately, because their ranges are restricted, remotely located, or have low occupancy across vast landscapes. Often, it can be a combination of all the above. To make this problem more tractable, we can predict species habitat suitability or probability of occurrence across large spatial areas, on the basis of small numbers (e.g. dozens) of accurate occurrence points (Phillips et al. 2006; Gomes et al. 2018). By identifying similar environmental conditions throughout the potential distribution of a species, it is possible to accelerate the discovery of unknown populations, particularly in remote and historically under-surveyed regions, such as the Top End (the 112,295 km2 northernmost region of the Northern Territory, Australia). Ecological niche modelling (ENM) has accelerated as a tool to (1) quantify range size, (2) identify unsuitable habitat characteristics that separate subpopulations (Bertola et al. 2018), and (3) highlight localities that (a) are potentially occupied by the species, (b) can be targeted for surveys or monitoring, or (c) would derive the greatest benefit from conservation resources (von Takach et al. 2020). Quantifying the range of environmental variables that represent suitable habitat for a species also allows us to identify conditions under which the species is less likely to persist. For example, patchy, low-intensity fires may allow populations to persist more readily than do larger, high-intensity fires (Radford et al. 2020).
The distribution of many threatened species in northern Australia partly or fully overlaps highly rugged areas of the landscape. Despite being largely undeveloped, the native ecosystems of northern Australia have been degraded by a variety of factors, including feral herbivores and pastoralism (Woinarski and Ash 2002; Legge et al. 2011), feral cats (Fisher et al. 2014; Frank et al. 2014), and frequent high-intensity fires (Woinarski et al. 2011). One species about which we know very little is the white-throated grasswren (WTGW, Amytornis woodwardi), a ground-dwelling bird with a restricted distribution (Noske 1992). As is the case for much of the mammalian fauna of the region (Woinarski et al. 2001, 2010), this species appears to have declined even in areas with protected status, such as the World Heritage-listed Kakadu National Park, where many other species are also declining (Woinarski et al. 2012). Because of the species’ reliance on mature spinifex patches, which requires specific fire-history attributes, and the legacy of unmanaged Country from the early 1940s to 2000s because of the disruption of traditional fire-management practices resulting from colonisation (Ritchie 2009; Russell-Smith et al. 2013), the decline and fragmentation of suitable habitat are likely to have had adverse impacts on WTGW populations (NESP TSR Hub 2019). This has led to the species being listed as Vulnerable under the Federal Environment Protection and Biodiversity Conservation Act (EPBC) 1999 and Endangered under the IUCN Red List (IUCN 2006).
The WTGW is the largest of the Amytornis grasswrens at 20–22 cm (Menkhorst et al. 2017) and is endemic to the rugged escarpment and outcrops of the Arnhem Plateau sandstone shrubland complex, itself an endangered ecological community EPBC Act. The Arnhem Plateau bioregion extends across an area of approximately 32,000 km2, with the western portion in Kakadu National Park and Nitmiluk National Park, and the eastern area within several current or planned Indigenous Protected areas (Warddeken, Mimal, and Djelk) and Aboriginal Land Trusts.
The WTGW is known locally as Yirlinkirrkkirr or Djirnidjirnirrjken in Bininj kunwok, and Nyirrnyirr in Jawoyn. The Dalabon name for the species has been lost, although the few elderly remaining speakers of the severely endangered language (Cutfield 2016) remember sighting the species in central southern Arnhem Land. The species is considered culturally important and is connected with songlines in the region:
The Yirlinkirrkkirr they have a connection with us, with the land, with the story, the songline, with the dance. All that comes together. [Terrah Guymala]
The WTGW is typically found in small groups of two to four individuals (likely being made up of a breeding pair and offspring from the most recent brood), and nests exclusively close to the ground in mature hummocks of perennial Triodia spp. (Schodde and Mason 1975) that occur in rocky gorges, outcrops and the crevices among large boulders often associated with dense shrubby gullies and boulder-strewn creek lines. The species is often found with the sympatric purple-backed fairywren (PBFW; Malurus assimilis dulcis), a more common and widespread species. Bininj Nawarddeken people think of the PBFW, locally known as the Dalkkidalkken, as an indicator species of WTGW presence, and it is sometimes affectionately referred to as a cousin (T. Guymala, pers. comm., 2022). Because of this, previous surveys for WTGW led by Warddeken and Djelk rangers also searched for and recorded the PBFW (Stevens et al. 2012). WTGWs tend to breed in the northern wet season, typically between December and April (extending to May or June, possibly depending on conditions), sometimes with a second brood in the same season (Schodde and Mason 1975; Noske 1992). Pairs with their offspring occupy territories of about 7–10 ha (Noske 1992). The WTGW feeds both on invertebrates (e.g. beetles, ants, grasshoppers) and plant material (e.g. grass seeds and leaf and stem fragments), with shifts in diet composition likely occurring as resource availability changes throughout the year (Noske 1992).
The importance of safeguarding the species from fire has long been known to Bininj Nawarddeken because grasswrens are not strong flyers (Noske 1992) and are likely to have a limited dispersal capacity. Fire is likely to affect WTGWs by removing food resources (plants and insects on living plants), depleting older spinifex necessary for nesting, and thereby reducing the ability to produce a brood if the fire occurs prior to breeding season, and large fires may directly kill some WTGWs (Noske 1992; Garnett et al. 2012). In more recent years, there has also been growing concern for adequate man-karrandalk (Triodia spp.) and shrubs to provide protection to the ground-dwelling species from suspected predation by feral cats.
In an adaptation of methods used in targeted surveys in Kakadu (Mahney et al. 2011), three bushwalk surveys were conducted across the Warddeken IPA in 2012 and six separate groups of WTGWs were found across the three locations (Stevens et al. 2012; Warddeken Land Management 2020). Regular monitoring in the Warddeken IPA by using motion-activated cameras has detected 10 new WTGW occurrences since 2017 (Warddeken Land Management in press). Nonetheless, across the region of the Arnhem Plateau, the few targeted surveys that have taken place for the WTGW do not have ideal temporal and spatial replication to enable robust inferences regarding population trends.
Under Phase 2 of the Australian Government’s National Landcare Program (2018–2023), Territory Natural Resource Management (TNRM) was contracted to facilitate and lead a 22-month project collaborating with groups across the Arnhem Plateau, to undertake targeted surveys to improve knowledge of WTGW locations and support strategic fire management to protect the species habitat. The project specified that survey locations were to be informed by predictive modelling, with input by expert and local knowledge.
Here, we aimed to characterise the habitat conditions under which WTGWs persist, as well as map these conditions across the species distribution in the Arnhem Plateau region. This was initially undertaken by constructing an ecological niche model that treats fire as a dynamic, rather than static, variable (as per Stoetzel et al. 2020, Amytornis dorotheae). We then integrated knowledge of Indigenous and non-Indigenous stakeholders through a combination of participatory planning, face-to-face consultations, and key Indigenous knowledge holder involvement for site-selection to test the habitat suitability predictions in the field. We aimed to test the most effective method for surveying WTGW presence by trialling bioacoustic audio recorders, call playback surveys, and motion-activated cameras at each site, and working with local Indigenous ranger groups to contextualise the ENM model and cultivate place-based conservation knowledge of the species. Improving our understanding of the drivers of WGTW occurrence, and particularly the influence of fire on habitat, will enable more appropriate fire management to improve conservation management of remnant populations.
Materials and methods
Occurrence records and cleaning
Occurrence (presence) points for the WTGW were collated from the Atlas of Living Australia, eBird, NRMaps, museum collections, and the Warddeken Indigenous Protected Area (IPA) records database. Occurrence points were then evaluated individually and any duplicates (including any from the same location in the same year) or points suspected of having inaccurate coordinates were removed. Coordinate uncertainty was provided in the metadata for many occurrence points, and for others, the accuracy was assessed on the basis of the locality description. In many cases, particularly for the eBird occurrences, the latitude and longitude that were supplied with the occurrence were not accurate enough for use with fine-scale topographic and fire data. As the earliest producible fire covariates were for the year 2000, any occurrences prior to the year 2000 were removed, retaining a total of 62 accurate records over a period of 20 years (Fig. 1). Across this period, there were 8 years (2005, 2006, 2007, 2008, 2010, 2013, 2015, 2018) with no high-quality records. The small number of records is predominantly due to the remoteness and inaccessibility of the region, the vast majority of which is off limits to the public and requires multiple permits and helicopter flights or overnight treks to conduct bird surveys. Of the 62 high-quality occurrence records, 29 observations included a count of the number of individual birds. The mean number of individuals observed was 2.3, with a minimum of one and a maximum of five.
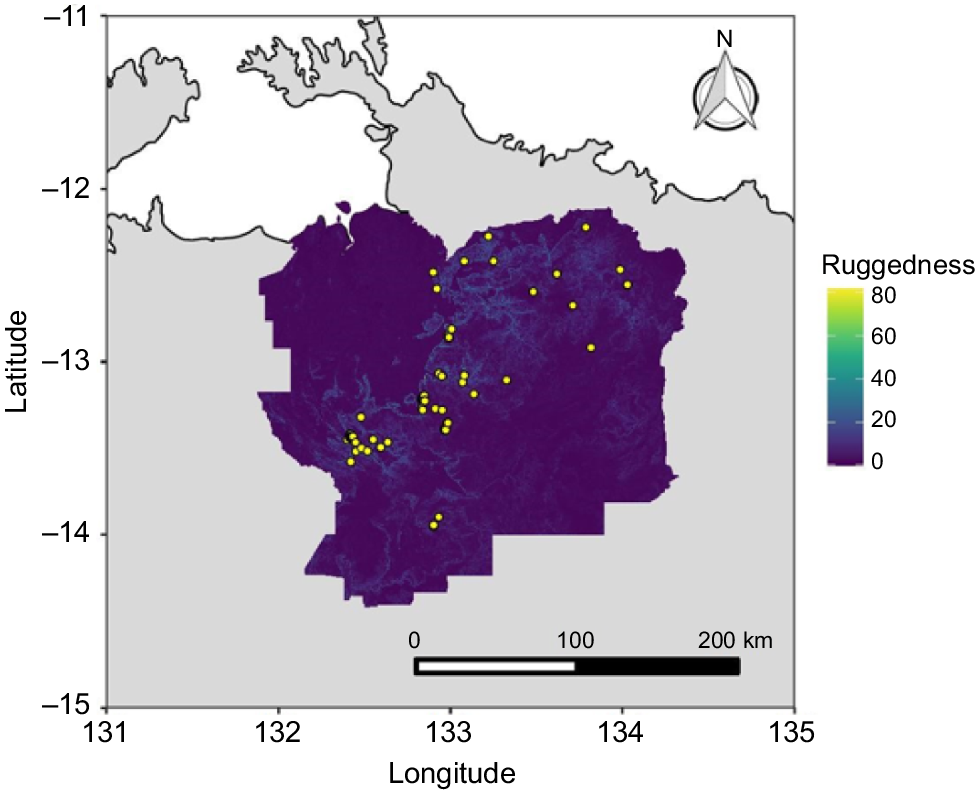
Study area and background sampling
A spatially appropriate study area is needed to (1) obtain adequate representation of the range of environmental variation available to the WTGW and (2) prevent models from overfitting to the bias in occurrence points (Anderson and Raza 2010). The ~32,000 km2 region that incorporates the entire known distribution (and potential habitat outside of the known distribution) of the WTGW is managed or co-managed by a range of organisations, including Warddeken Land Management, Jawoyn Association Aboriginal Corporation, Mimal Land Management, Nitmuluk National Park, Bawinanga Aboriginal Corporation, Kakadu National Park, Gundjeihmi Aboriginal Corporation, and Njanjma Aboriginal Corporation. To obtain a suitable study area for model fitting within this region, we used the historical extent of occurrence (EOO) of the WTGW (as per Moore et al. 2019; von Takach et al. 2020). To define the EOO, we circumscribed all filtered presence points by using α-hulls, a standard IUCN measure for delineating the geographic distribution of a species (IUCN 2006). Once the model was built, it was then used to predict habitat suitability to the whole study area.
To obtain background (pseudo-absence) points with which to contrast the occurrence points, we extracted 10,000 points from all covariate (predictor) rasters. We compensated for sampling bias in the occurrence data by incorporating a probability function into the background sampling that corresponded to the bias in the occurrence data. This was derived from a Gaussian kernel-density grid of all bird species records (68,085 occurrences) across the study area, via the kde2d function of the ‘MASS’ R package (ver. 7.3-53, https://CRAN.R-project.org/package=MASS) (Venables and Ripley 2002), with the default normal reference bandwidth calculation. This method of target-group sampling is widely used to account for spatial bias in sampling effort (Phillips et al. 2009; Syfert et al. 2013; Molloy et al. 2017).
Environmental covariates
The environmental covariates selected for use in the ecological niche model were those either known to be relevant for Amytornis grasswrens or were considered by the authors, including Traditional Custodians, to be of potential ecological importance for the WTGW. Between 2018 and 2021, TNRM facilitated a series of workshops with Traditional Custodians and Indigenous ranger groups under a separate National Landcare Program-funded project that spanned the greater West Arnhem region. In these workshops, the extent of fires in WTGW habitat was highlighted as a threat and some people confirmed current and previous occurrence records.
Three fire-related variables were calculated from the years 2000–2019 by using a regional fire-history archive derived primarily from Landsat and Sentinel-2 (20 m × 20 m resolution) satellite sources (Table 1). These consisted of the 5 year averages of (1) the proportion of area burnt (PAR) around each site (400 m radius), (2) the distance to nearest unburnt (DUB) vegetation of all pixels in a 400 m radius, and (3) the proportion of vegetation in a ≥5-year-old category within a 400 m radius (time since fire, TSF). The radius of 400 m was selected because this represents an area approximately five times larger than the typical home-range size of the WTGW (Noske 1992), thus representing the local patch, while also accounting for (1) small (e.g. 100–200 m) inaccuracies in occurrence location and (2) movements of individuals around the precise occurrence point. The proportion of vegetation that had not experienced fire in ≥5 years was selected because grasswrens have the specialist habit of nesting in mature spinifex clumps, with the sister species, the Carpentarian grasswren, requiring at least 3 years for spinifex to mature before it is occupied (Perry et al. 2011; Harrington and Murphy 2016). Three additional environmental covariates were selected, including (1) a measure of potential soil moisture/water availability at a 90 m × 90 m resolution (topographic wetness index, TWI), (2) a measure of topographic heterogeneity at a 90 m × 90 m resolution (topographic ruggedness index, TRI), and (3) a measure of vegetation cover (mean long-term vegetation fractional cover, at 500 m × 500 m resolution, VEG) (Table 1). All variables in the model were raster layers unified to a resolution of 100 m × 100 m as well as the same extent and projection.
| Acronym | Name | Dynamic or static | Description and source | |
|---|---|---|---|---|
| PAB | Proportion of area burnt | Dynamic | Proportion of area burnt around each site using a search radius of 0.4 km for the preceding calendar year and averaged over 5 years. Includes all fires (i.e. no seasonality) (Lawes et al. 2015). Source data 20 m × 20 m resolution. | |
| DUB | Distance to unburnt (m) | Dynamic | A measure of fire extent. Average distance to nearest unburnt vegetation from all pixels within a search radius of 0.4 km around the survey point at the end of preceding calendar year and averaged over preceding 5 years. Includes all fires (i.e. no seasonality) (Lawes et al. 2015). Source data 20 m × 20 m resolution | |
| TSF | Time since fire | Dynamic | Proportion unburnt within a search radius of 0.4 km at the end of each calendar year for the preceding calendar year and averaged over preceding 5 years for post-fire age class of >5 years. Includes all fires (i.e. no seasonality) (Radford et al. 2015). Source data 20 m × 20 m resolution | |
| TWI | Topographic wetness index | Static | Estimates the relative wetness within a catchment. Calculated as log_e (specific catchment area/slope). Obtained from the CSIRO data collection (Gallant and Austin 2012; Gallant et al. 2012). Source data ~90 m × 90 m resolution | |
| TRI | Topographic ruggedness index | Static | A measure of topographic heterogeneity based on elevation differences between adjacent pixels. Made in R by using the terrain function of the ‘raster’R package (ver. 3.0.2, https://cran.r-project.org/web/packages/dismo/index.html) and a 3-s digital elevation model (von Takach et al. 2020). Source data ~90 m × 90 m resolution. | |
| VEG | Mean vegetation fractional cover (%) | Static | The percentage of photosynthetic vegetation, calculated by averaging the vegetation fractional cover identified by MODIS imagery from 2000 to 2018 using the month of September (after annual grass has died off). Source data 500 m × 500 m resolution. Obtained from TERN Auscover (http://www.auscover.org.au/) (Guerschman et al. 2009; Einoder et al. 2018; von Takach et al. 2020). |
We treated all fire variables as dynamic covariates (as per Stoetzel et al. 2020). To do this, each fire variable was calculated for each year, values for each covariate were extracted for all WTGW occurrences, and the date of each occurrence was used to identify the relevant time-sliced fire data. The fire history pertaining to each occurrence date is thus much more relevant to the ecology of the target species than would be a static longer-term average, and allows for investigation of the change in habitat suitability across the landscape from 2000 to 2019. Background points for the fire covariates were randomly sampled from the same set of years as for the occurrence data. All raw data for each variable was scaled and centred to allow for comparison and then plotted as boxplots to visualise differences between values at presence and background points.
All covariates were checked for multicollinearity. Although our method of habitat suitability modelling is robust to correlations among variables (Elith et al. 2011), we investigated the potential for any bias using variance inflation factors (VIFs). We used the vif function of the R package ‘usdm’ (ver. 1.1-18, https://CRAN.R-project.org/package=usdm) (Naimi et al. 2014) to calculate VIFs based on the square of the multiple correlation coefficient resulting from regressing a predictor variable against all other predictor variables. No variable had a VIF of ≥2, indicating that collinearity was not a problem.
Ecological niche modelling
We built an ecological niche model of habitat suitability by using the Maxent algorithm (Phillips et al. 2006; Phillips et al. 2017). This method is effective at accounting for spatial bias in sampling, using large numbers of background (pseudo-absence) points from across the landscape to contrast with occurrence points (Elith et al. 2011; Syfert et al. 2013). The default output of Maxent is a habitat-suitability value that has undergone a logistic post-transformation, in an attempt to obtain a value that is as close to the probability of occurrence as possible, given the environment (Elith et al. 2011; Calabrese et al. 2014).
The model was run via the Maxent function of the ‘dismo’ R package (ver. 1.1.4, https://CRAN.R-project.org/package=dismo), by using five cross-validation runs, a betamultiplier of 1.5 and linear, quadratic and product feature classes. Average values across all cross-validation runs for two metrics were extracted from the model output. These metrics included (a) the area under the curve (AUC) of a receiver operating characteristic, which estimates overall model fit via a threshold-independent measure of predictivity, and (b) permutation importance, a measure of the relevance of each predictor variable to the response variable. To calculate permutation importance, the values of a predictor variable are first randomly permuted among the occurrence and background points, the model is re-evaluated, and the resulting drop in the AUC value is calculated and normalised to a percentage (Searcy and Shaffer 2016; Phillips et al. 2017).
We then overlaid the rasters for each year from 2005 to 2019 and summed the habitat-suitability values across all layers, producing a long-term habitat-suitability model. The values of this model were compared with those of a short-term habitat-suitability model (2017–2019), via Pearson’s product–moment correlation. Although short-term suitability may be useful for inferring current habitat conditions, we consider the long-term suitability more appropriate for targeted survey design because the long-term model provides information on consistency of suitability. White-throated grasswrens are unlikely to be strong dispersers, owing to their small home ranges and tendency to avoid flying. This means that sites that have been consistently suitable for the species are more likely to be occupied than are sites that have recently become suitable, as recolonisation of historically unsuitable patches may not have occurred.
Site selection with Traditional Custodians
The project area occurs across many Aboriginal clan estates and the management areas of several Indigenous ranger groups and a national park. As such, we wanted to ensure (1) that there was free, prior and informed consent with Traditional Custodians for the project to occur on their land, (2) that we conducted a collaborative approach to site selection where Traditional Custodians chose the specific sites where surveys would take place, and (3) that people had the opportunity to provide their knowledge of the species, including current or previously occupied areas, on the basis of their understanding of WTGW habitat requirements and suitable management actions.
The first and third authors and their colleagues facilitated face-to-face collaborative site-selection workshops by using the ENM map as a visual basis for planning in Katherine, Pine Creek, Jabiru, Mirrar estate, and Bulman with Jawoyn, Djurrubu, and Mimal Traditional Custodians and rangers respectively. Warddeken held face-to-face consultations by the last author in Jabiru, Kabulwarnamyo, Manmoyi, Mamadawerre and Marlkawo with Dawarro and Djungkay (Traditional Custodians). Through the participatory workshops and consultations, ecologists and Indigenous participants outlined their understanding of the species, current threats, habitat requirements, discussed potential survey methods and whether people would be happy to use them, and Traditional Custodians provided their knowledge of areas of historical WTGW occurrence and of local importance within or nearby varying values of habitat suitability as predicted by the model. This approach was aimed at testing the model’s predictive accuracy in the field. Traditional Custodians also indicated areas that were predicted to have suitable habitat but were culturally inappropriate to access and thus were marked as ‘exclusion zones’ and avoided in further discussions regardless of habitat suitability. Because access was restricted via helicopter, logistical considerations that influenced site selection included (1) distance to transport personnel from outstations/communities to sites, (2) feasibility of landing sites, and (3) distance to fuel-drop locations. We then mapped out the sites and plots in chosen areas to upload to devices for use in the field and printed in hard copy. At the onset of each field trip, a Traditional Custodian or Indigenous ranger for each area visited each site by helicopter with ecologists for visual approval, to assess fine-scale habitat attributes of the site, identify evidence of any recent fire that may have occurred since the data within the model and ground truth, and identify suitable landing and equipment-installation areas. Owing to the size of each site (each length was 560 m long) and the rugged nature of the landscape, sometimes several of the 2 ha plots within a site were unavoidably mapped over less suitable habitat which meant that predicted areas of suitable habitat within a plot were relatively small compared to other plots. Therefore, when undertaking the initial aerial site surveillance, these 2 ha plots would be crossed off a hard copy map and excluded from ground survey.
Survey methodology
In total, 39 sites were surveyed between August 2021 and May 2023. Each site comprised a 32 ha area containing 16 × 2 ha plots (modified from NT DEPWS Flora and Fauna protocols developed in 2016 (L. Einoder, pers. comm., 2021), e.g. Fig. 2) for methods regionally comparable to Kakadu National Park. The following three methods were used to target WTGWs: (1) bioacoustic audio recorders (BARs); (2) call-playback (CPB) surveys; and (3) motion-detection camera-traps. All sites were surveyed with BARs; however, owing to the inability to safely traverse on foot around some of the most rugged sites, only a subset of sites were also surveyed using CPB (n = 26) and camera-trapping (n = 32) (Table 2).
Clockwise from top left: the five sites on Mirrar estate demonstrating the 16 × 2 ha plots contained within each site; tracks walked in six plots within one site during CPB surveys on Jawoyn country; Djurrubu rangers setting a BAR on Mirrar estate; Jawoyn ranger conducting CPB survey in Nitmiluk National Park; Mimal rangers setting speaker to prepare for CPB survey on Dalabon Country; Warddeken daluk rangers setting a camera-trap in Warddeken IPA; white-throated grasswrens detected during CPB survey on Warddeken IPA. Photos: Kelly Dixon, Brittany Hayward-Brown, Cara Penton. Maps base layer: Google.

| Year | Site | Group/s | Number of BAR sites | Number of camera sites | Number of call-playback sites | Number of call-playback plots (2 ha) | |
|---|---|---|---|---|---|---|---|
| 2021 | Manyallaluk ALT | Jawoyn | 9 | 9 | 6 | 37 | |
| 2022 | Warddeken IPA | Warddeken | 9 | 9 | 5 | 28 | |
| 2022 | Mirrar estate | Djurrubu | 5 | 5 | 4 | 22 | |
| 2023 | Nitmiluk NP | Jawoyn and NT Government Parks & Wildlife rangers | 5 | 5 | 4 | 23 | |
| 2023 | Dalabon Country (proposed Mimal IPA) | Mimal | 4 | 4 | 4 | 17 | |
| 2023 | Warddeken IPA | Warddeken | 7 | 0 | 3 | 18 | |
| Total | 39 | 32 | 26 | 145 |
Automated (passive) recording devices, i.e. bioacoustic audio recorders placed in a study area to record vocalisations and detect the presence of species, have been increasingly used in recent years for wildlife surveys and are especially useful for monitoring cryptic and threatened species (Blumstein et al. 2011). During the collaborative planning sessions, many ranger groups expressed a desire to build their capacity by learning new survey methods, including the use of BARs. In this study, we used BARs with a solar panel (‘solar BARs’; Frontier Labs, Qld, Australia) that continually charges the battery. While undertaking the initial site visit with appropriate Traditional Custodians, a suitable location was chosen in which to deploy the BAR and marked in a GPS app. The location was chosen on the basis of height relative to surrounding habitat to increase capture of sound, closeness to potentially suitable WTGW habitat patch, and helicopter-landing accessibility. Each solar BAR was attached with its bracket to a 1.8 m star-post and faced north 10° to ensure maximum sun capture. The devices were mostly programmed to capture sound continuously from 5 am until 8 pm (the devices were programmed for 24 h in the Jawoyn 2021 surveys), with each file recording for 1 h. BARs were left in situ for 4–6 weeks. No efficient automated system to detect the WTGW call existed and commercially available bioacoustic software programs proved ineffective for detecting and extracting the complex call. Therefore, a semi-automated bioacoustic call-recognition system was created for this project, which was successful at detecting the WTGW call (K. Armstrong, K. Dixon, R. Morgans, unpubl. data).
Call-playback (CPB) involves the call of the target species being played to elicit a call or visual response. WTGWs are very responsive to call playback, and some ranger groups had previously used CPB when searching for the species (Stevens et al. 2012); therefore, it was chosen as a survey method. CPB surveys were undertaken in 145 plots within 26 of the 39 sites in the early morning or late afternoon when birds are more active. Two teams of observers comprised of ecologists and Indigenous rangers and/or Traditional Custodians (a mixture of older and younger people) undertook surveys concurrently in plots at opposite ends of the site. Within each 2 ha plot, a 30 min survey was undertaken. WTGW calls from the Pizzey and Knight Birds of Australia Digital Edition (ver. 1.9) mobile app (http://play.google.com/store; Gibbon Multimedia) were broadcast from an Ultimate Ears (UE) Boom 3 portable Bluetooth speaker. Observers set the speaker on a rock and crouched behind a spinifex clump, rock, or shrub and played the call for 30 s. Observers then waited for another minute for a return call or a sighting of the bird then played the call again for a further 10 s, if no response was detected. In each 30 min survey, the call was broadcast six times, while walking around the 2 ha plot, with no less than 5 min between each call. While undertaking the surveys, any other bird species seen and/or heard within the plot were recorded. Of particular interest were detections of the purple-backed fairywren, a species with similar habitat requirements to WTGW.
Tracks were recorded and KMLs of the boundaries of the 2 ha plots were displayed on the Geotracker app (https://geo-tracker.org) to ensure as much of the 2 ha plot was traversed as possible, while not crossing into the neighbouring plot. Within each 2 ha plot, habitat characteristics were recorded at the completion of each 30 min survey. Variables recorded included the landscape position, habitat type, evidence of fire, and the approximate percentage cover of trees, shrubs, grasses and bare rocky areas.
At 32 of the 39 sites (all sites except the Warddeken 2023 surveys), an array of five motion-detection camera-traps (HP2W, Reconyx, Wisconsin, USA) were set in a diamond configuration, i.e. a camera in each of four directions 50 m from a centre camera, nearby the BAR in areas that would be likely to detect WTGWs and feral cats, a probable key threat to the species. Unbaited cameras were set to take photos day and night, on high sensitivity, with three photos and a 5 s video per trigger. Cameras were left in situ for 4–6 weeks and images were tagged using the Timelapse software (ver. 2.3.2) (https://saul.cpsc.ucalgary.ca/timelapse/). Reports, maps, and data were provided back to each group after analyses and included a list of all species detected and photos of animals that were known to be culturally important or of ecological significance, such as echidnas, black wallaroos, Arnhem rock-rats, cats, and other feral animals.
Results
Ecological niche modelling
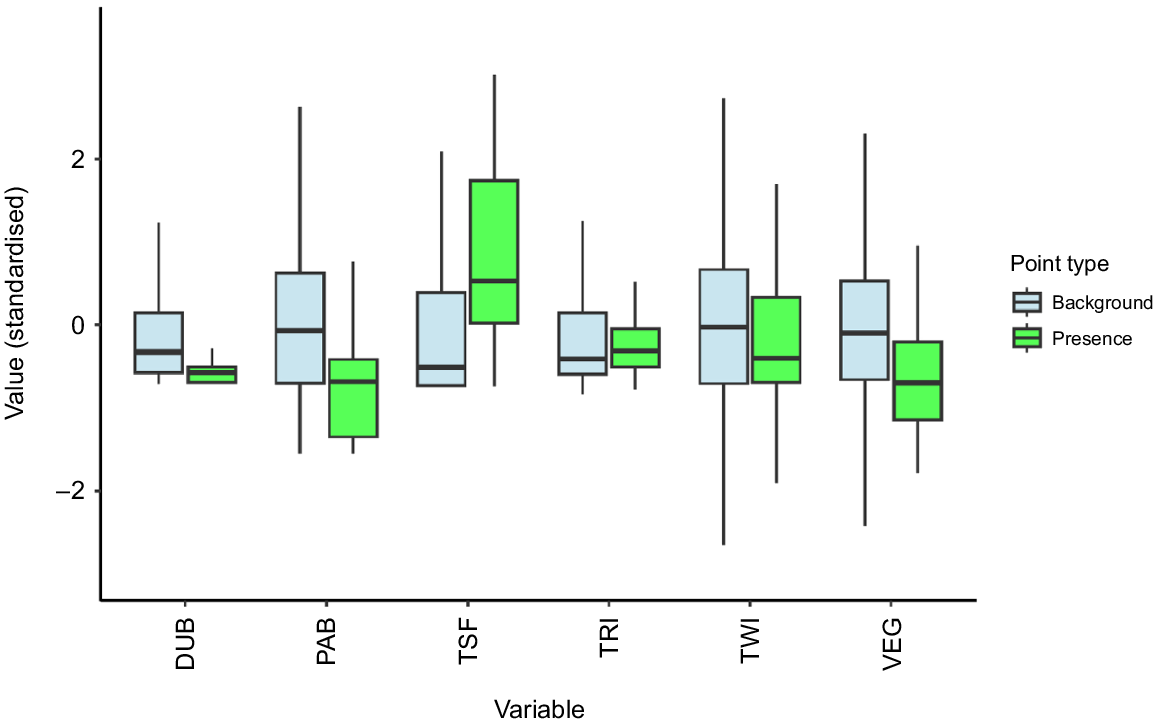
Visualising the covariate data for the presence points and background points shows some clear differences in the median values of environmental variables (Fig. 3). WTGWs are typically found in locations that have lower DUB, PAB, and VEG values, and higher TSF values, than the average background values across the landscape.
Visualising data used in the ecological niche model for the WTGW. Values for each variable were scaled and centred to allow for comparison. Light blue boxplots represent data at 10,000 randomly sampled background locations across the landscape, whereas green boxplots represent data at the 62 locations where white-throated grasswrens were observed between 2000 and 2019.
The ecological niche model was a robust predictor of WTGW occurrence, with a mean training AUC value of 0.87 and mean testing AUC of 0.85. The covariates that contributed most strongly to the predictive power of the model, as identified using the permutation-importance values (Table 3), were mean vegetation fractional cover (32.3%) and distance to unburnt (55.6%). Although the amount of long-unburnt vegetation (‘time since fire’) around a point had a reasonably large (23.5%) contribution to the heuristic process involved in fitting an optimal model, the lower permutation-importance value (8.7%) suggested that the final model does not heavily rely on this covariate. Overall, our model suggested that WTGWs are strongly associated with areas that have a history of vegetation being retained in the immediate vicinity of a survey point, as well as areas that tend towards lower vegetation cover (e.g. spinifex-dominated sites with areas of bare rock). In this context, the model was not biased towards areas of Allosyncarpia forest, despite such areas having a strong association with long-unburnt vegetation. There was a highly significant, moderate correlation (r = 0.68, P < 2.2–16) between the long-term (2005–2019) habitat-suitability and the short-term (2017–2019) habitat-suitability layers, suggesting that there is at least some regular turnover in habitat suitability across the landscape through time.
| Variable | Percentage contribution | Permutation importance | |
|---|---|---|---|
| VEG | 37.8 | 32.3 | |
| DUB | 36.8 | 55.6 | |
| TSF | 23.5 | 8.7 | |
| PAB | 1.2 | 2 | |
| TRI | 0.5 | 0.6 | |
| TWI | 0.3 | 0.9 |
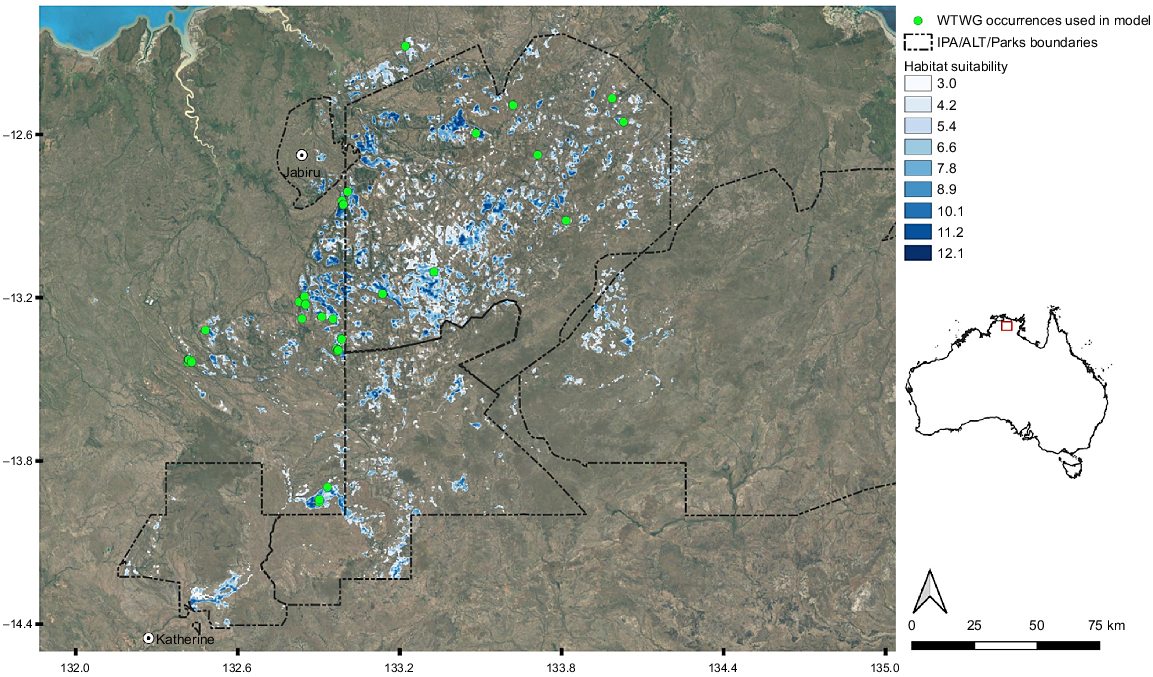
The total extent of occurrence (EOO) of the WTGW using all accurate records from 2000 onwards was calculated to be 18,638 km2. However, the area of occupancy within this EOO is far less than this, owing to the highly fragmented nature of potential habitat across the region. Summing all cells with suitability scores of >6 from our long-term (2005–2019) habitat suitability-model prediction, we calculated an area of 1421 km2 of habitat. Reducing this threshold to scores of >3 (Fig. 4), or increasing it to scores of >9, results in areas of 2631 km2 and 536 km2 respectively. Thirty-nine sites were chosen by Traditional Custodians within this area (Fig. 5).
Habitat-suitability mapping for the WTGW. All suitability values of <3 have been removed from the map to highlight areas of higher suitability across the landscape. WTGW occurrences include only confirmed records between 2000 and 2019. Base map layer: Google.
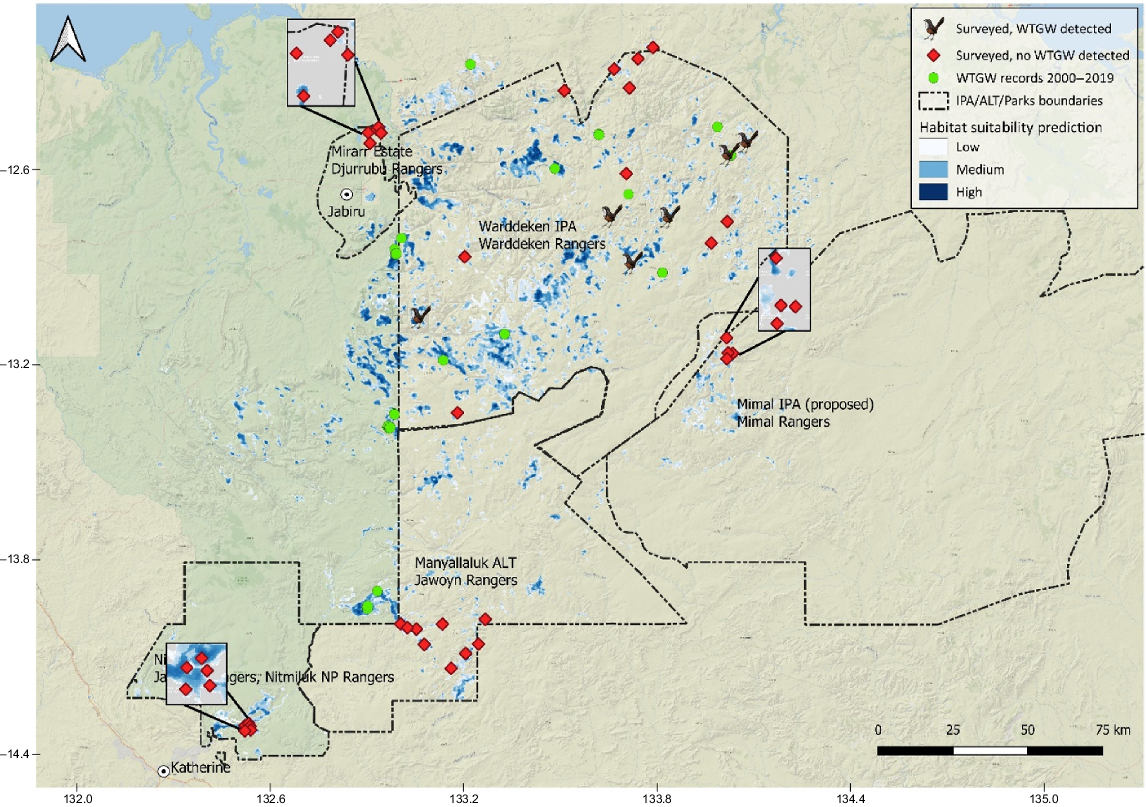
Survey area indicating six survey sites that detected grasswrens (grasswren icon), 33 sites that did not (red) and previous occurrence records from 2000 to 2019 (green). Three survey areas have magnified inset boxes to better display the locations of sites. Base map layer: ESRI.
The fine-scale fire data used in the ENM were available only until 2019; therefore, we anticipated that some of the areas predicted to be suitable by the model were likely to have had fire occur since 2019. Prior to surveys, we mapped any fire in the survey area by using NAFI data to identify whether fire had occurred since the data in the model. In some instances, a small amount of a NAFI pixel from one of the previous 2 years overlapped a partial part of some of the plots within a site. As these pixels are relatively coarse compared with the fine-scale data included in the model, we did not immediately omit these areas for survey, and visited them for ground-truthing. We did not identify any sites with evidence of recent fire across more than 3 of the 16 plots. Fire was actively burning in the valley below one of the Mirrar sites at the time of surveys. At this site, a BAR and cameras were set but CPB was not undertaken. While undertaking CPB surveys, some sites within Jawoyn and Warddeken Country had small patches of evidence of fire (e.g. old charring on logs), whereas the spinifex hummocks were large, and Traditional Custodians explained that the fire had most likely occurred several years prior.
Survey results
We detected WTGWs at 6 of the 39 sites, all within the Warddeken IPA (Fig. 5). WTGWs were detected at all six sites on BARs. BAR data totalled 1488 days, 25,767 h, and 6.5 terabytes (TB) of recordings, and analysis produced a total of 3246 manually validated ‘song sequences’ (K. Armstrong, K. Dixon, R. Morgans, unpubl. data). BAR results indicated that the majority of the species’ calls were around dawn and dusk, with the most prolonged songs in the 2 h before sunset, with a few outliers after sunset and in the middle of the day. At one of these six sites, WTGWs were also detected on one camera and by CPB surveys (in two plots at the one site) (Table 4).
| Year | Location | Number of sites WTGW detected by method | Number of sites PBFW detected by CPB | Number of sites feral cat detected by CT | |||
|---|---|---|---|---|---|---|---|
| BAR | CPB | CT | |||||
| 2021 | Manyallaluk ALT | 0 | 0 | 0 | 2 | 3 | |
| 2022 | Warddeken IPA | 4 | 1 | 1 | 4 | 3 | |
| 2022 | Mirrar estate | 0 | 0 | 0 | 2 | 1 | |
| 2023 | Nitmiluk NP | 0 | 0 | 0 | 3 | 2 | |
| 2023 | Dalabon Country (proposed Mimal IPA) | 0 | 0 | 0 | 0 | 0 | |
| 2023 | Warddeken IPA | 2 | 0 | NA | 2 | NA | |
The sites in which four of the six detections occurred were within the model prediction area. Two were slightly outside; one site was 320 m and one site was 630 m from the edge of the nearest pixel. The closest detection during these surveys (Warddeken 2022 survey) to a previous occurrence record (from 2014) was 672 m. The next-closest detection to a previous record was 7 km, with the furthest detection being 58.5 km from a previous record. Thirteen of the sites were slightly outside the predictive area, owing to the rugged nature of the terrain and the need to sometimes move the sites slightly when ground-truthing from what had been mapped. Most of these sites were within 500 m from the edge of the nearest pixel, with the furthest being 630 m from the nearest pixel (Fig. 5, Supplementary Table S1). Because these distances are within about one pixel of resolution of the most important predictor covariate (Tables 1, 3), we consider these sites as within the model prediction area.
Traditional ecological knowledge (‘local knowledge’) was fundamental throughout this project, including prior to the creation of the model, for site selection, while ground-truthing and undertaking surveys, in addition to subsequent fire management planning and burning activities. Local knowledge by Bininj Nawarddeken people such as senior knowledge women Mary Naborlhborlh, who has since passed away, and Mary Kolkkiwarra commonly reported sightings of the WTGW near outstations in the Warddeken IPA including Manmoyi, Kamarrkawarn, and Kabulwarnmyo in place where ‘karri kore kanjdjikandji, ankebkakebkali’, translating to ‘the river by the edge of the rocks where they meet the savanna’ (Bininj Kunwok Language Centre 2012).
Corresponding with traditional knowledge of the WTGW, the ENM highlighted the critical importance of long-unburnt habitat, as follows:
He hides in the Man-karrandalk [spinifex], that one. [Stuart Guymala]
All WTGW detections in this project were within the Warddeken IPA, directly corresponding to Nawarddeken Bininj knowledge of suitable habitat in the kuwarddewardde Country for the WTGW, as stated in the following:
He is saying who he is, for sure you will be able to find one, the white-throated grasswren… Just try and have a look around. I’m only assuming you will find him there because it’s the right kind of habitat in the rock country there. [Mary Kolkiwarra Nadjamerrek]
This project involved collaboration with six Indigenous land management organisations, engaging 86 Indigenous experts and practitioners in collaborative site selection and data collection. Specifically, 78 individuals participated in on-Country surveys across the Arnhem Plateau, comprising 65 Traditional Custodians or Indigenous rangers and 13 non-Indigenous practitioners. Additionally, 21 Traditional Custodians contributed to the participatory mapping and site selection though they did not partake in fieldwork. The project cost over A$220,000 and more than A$50,000 was negotiated for fee-for-service work with organisations, allowing them to propose suitable time frames for completing work and noting the right people to be involved. Overall, 32 days of fieldwork facilitated helicopter access to reconnect with Country across the Arnhem Plateau, most of which is highly inaccessible.
Discussion
Our research employed a multiple evidence-based approach, leading to the identification of new presence records for the WTGW within the Warddeken IPA. By combining our ecological niche model, with the input of local knowledge and verified on-ground observations, our study employs both scientific and Traditional ecological knowledge to evaluate all available information on the species to efficiently and effectively guide a survey program. This collaborative approach brings together not only scientific and land management practitioners, but also two different worldviews, conservation science and Indigenous, to strengthen our collective knowledge of the distribution of a threatened species in a highly remote and largely inaccessible landscape. This effort has provided these practitioners with a valuable resource for making well-informed, strategic decisions regarding the conservation and management of the WTGW (Verdon et al. 2019). Importantly, this approach recognises the importance of considering not only scientific data, which can be scarce in remote northern Australia, but also the rich historical, cultural and spiritual knowledge held by Indigenous communities in the region in threatened-species conservation projects (Campbell et al. 2022; Ghanbari and Turvey 2022; Russell et al. 2023).
Indigenous communities possess invaluable knowledge, especially when it comes to their deep relational understanding of animals, plants, and the landscape at large (Tynan 2021). In the context of this research, Traditional Custodians played a pivotal role in authenticating and refining the accuracy of the scientific model.
Occupancy of the available habitat across the distribution of the WTGW has long been a topic of some debate. For example, Noske (1992) estimated that there was about 14,000 km2 of habitat available for the species. Woinarski (1992) responded to this by suggesting that the occupancy within this area of habitat was low, with less than 3000 km2 of suitable habitat actually available at the time. Using rough high and low thresholds of suitability scores, our model predicted that there is likely to be somewhere between 536 km2 and 2631 km2 of suitable habitat for the species. This indicates either (1) that there has been a decline in the amount of habitat since those original calculations were made (e.g. owing to an inappropriate fire regime), or (2) that the area of habitat across the landscape is perhaps smaller than was originally estimated. We consider our estimates to be slightly optimistic, because our model was used to predict areas slightly outside the known distribution of the species, including some locations that are isolated from the core of the distribution by unsuitable lowland woodland habitat.
The ENM found that the species’ relationship with vegetation cover broadly agrees with the conventional and traditional belief that the species prefers sandstone heath vegetation located among large areas of bare rock, the inference being that these areas provide refuge from fire (Noske 1992). If this is true, it may be a result of the realised niche (i.e. the range of occupied habitats) contracting towards such areas through time because of unfavourable fire regimes. Interestingly, in the model, the influence of fire interval (higher average time since fire values) is not as relevant for population persistence as is the requirement for the local retention of at least some unburnt vegetation from year to year (lower average distance to unburnt values). Although our model does not incorporate effects of fire seasonality or the presence of unburnt patches within a fire perimeter, on the Arnhem Plateau fires are much smaller (Yates et al. 2008) and the retention of unburnt patches is far greater in areas where fires occur during the cooler months of the early dry season, especially in areas where bare rock physically separates patches of flammable spinifex-dominated vegetation (Price et al. 2003). Nonetheless, challenges remain for managers tasked with mitigating severe fire, while at the same time retaining functional habitat given that spinifex-dominated vegetation in the region can support intense fire after ~3 years of being unburnt (Russell-Smith et al. 1998).
This species appears to have declined from previously known sites because of the impacts of fires on its preferred habitat. Fire regimes in the Arnhem Land sandstone plateau over much of the past century have been characterised by frequent, high-intensity fires (Russell-Smith et al. 1998). Many plant species in the sandstone heath vegetation are obligate seeders, and require particular intervals of time between fires to successfully develop viable seed crops; therefore, such fire regimes were likely to be detrimental to both heath vegetation and WTGW population growth.
Following Bininj Nawarddeken people returning to ancestral lands as part of the homelands movement (House of Representatives Standing Committee on Aboriginal Affairs (HRSCAA) 1987), and in recent decades the advent of carbon market-based resourcing, the opportunity for fire management by Traditional Custodians and rangers in the region has steadily increased, resulting in marked improvements to the fire regime (Russell-Smith et al. 2015; Evans and Russell-Smith 2019; Ansell and Evans 2020; Edwards et al. 2021). Although some anecdotal evidence of localised WTGW recovery exists (e.g. observed recolonising of areas near Manmoyi after fire has occurred where unburnt patches have persisted nearby (T. Guymala, pers comm., May 2022), there has been no long-term monitoring of WTGW population trajectories.
Population recovery of declined species may not occur as rapidly as fire-regime change. In the case of the WTGW, areas that are now becoming suitable may not have been available for occupation for most of the previous few decades. The recolonisation rate of previously unsuitable patches is likely to be an important driver of WTGW occupancy across the landscape. If the species is a poor disperser, as predicted, it will be less able to recolonise patches from which it was previously extirpated, implying lower levels of occupancy. Developing our understanding of the species distribution, habitat associations, and the relationships between habitat connectivity and recolonisation ability over longer time scales will greatly assist with conservation management of WTGWs in the Northern Territory.
Survey methodologies
Aspects of some methods were adapted slightly among some groups, on the basis of cultural priorities and ranger skills/preferences. For example, BARs were set to record overnight as requested by Jawoyn Traditional Custodians, and Warddeken rangers recorded fieldwork metadata in the Cybertracker app (Ansell and Koenig 2011; Ens 2012). The use of multiple digital tools provided the opportunity for upskilling and trialling new methods, with some partners integrating into their own Indigenous community-based monitoring (ICBM) (Warddeken Land Management, unpubl. data). This approach of adapting to the preferences and consent of Traditional Custodians ensures a more inclusive and respectful collaborative environment (Paltridge et al. 2020).
The ongoing collaborative process, characterised by its flexibility, plays a crucial role in facilitating greater participation by Indigenous rangers, accommodating various levels of consent for different types of fieldwork (Robinson et al. 2023). For example, there were some preferences for passive sampling methods such as BARs or cameras, as opposed to CPB surveys, because some people felt they were less intrusive, as stated in the following:
That was last year when they went to Mokmek area and looking for Yirlinkkirrkirr. If you walking towards him, he can hear from long way. And run inside the spinifex grass. I can hear. [Deborah Nabarlambarl]
In our project, BARs were the best method at detecting WTGWs (when analysing the enormous amount of data with the semi-automated system developed to identify their call (K. Armstrong, K. Dixon, R. Morgans, unpubl. data). BARs have the capacity to monitor bird species as well as, or better than, higher-effort methods (Darras et al. 2019); however, they are unable to collect certain information such as the number and sex of animals. Nonetheless, to identify whether target species such as the WTGW are present in an area, BARs provide a more cost-effective option (Darras et al. 2019) and could be used as an initial method, to then follow up with more intensive visual surveys if the species is detected. Bioacoustics are a novel technology for many land-management groups and hold significant future potential, especially considering the adaptability of these datasets with emerging artificial intelligence (AI) technology (Sharma et al. 2020). Gaining research skills and training to increase the capacity of rangers on Country is considered a priority in research collaborations by many groups (Robinson et al. 2023). During collaborative planning meetings, many people were excited at the opportunity to learn novel methods for monitoring, stating the following, for example:
I’m really glad we using that [BAR] differently because we want the method to focus on just the grasswren. [Terrah Guymala]
The integration of bioacoustic data and AI can lead to more sophisticated and efficient methods for monitoring and understanding diverse ecosystems (Kwok 2019; Wearn et al. 2019); however, this integration needs to be appropriately governed, especially in the context of Indigenous community-based monitoring (Kukutai 2024).
We deployed one BAR at each of the 39 sites within the six survey areas over 2 years. There was a minimum distance of 963 m and a maximum distance of 34.86 km from a BAR to the nearest BAR, regardless of deployment time (Fig. 5). Although it was beyond the scope of this project to identify the maximum distance that these devices can detect a WTGW call, we assumed that bird calls would not be detected further than 50–100 m away (Darras et al. 2018) and expected that climatic and geographic variables would influence the distance a call would carry. The maximum number of BARs that were deployed on any one survey was nine. We chose the location of equipment deployment on the basis of visual inspection of the site via helicopter with a Traditional Custodian (as described in Materials and methods). Other areas within the larger site may have been more likely to detect WTGWs than were the areas where BARs were placed; however, the limited number of BARs available and the ruggedness, and therefore inability to land the helicopter in many suitable looking areas, dictated where the BAR was deployed. It is likely that if more BARs were able to be deployed across the larger area, more WTGW detections would occur.
Call-playback (CPB) surveys offered the opportunity for substantial time on Country and the possibility of sighting the WTGW; however, the 2 ha plot methodology proved to be cumbersome in rugged terrains and did not necessarily align with the way people would navigate the landscape using landform and considering safety. This challenge highlighted the need for adapting methodologies to suit the varied and often demanding physical landscapes of the survey areas, taking into consideration the preferences of Traditional Custodians. Previous sightings of WTGW in the Warddeken IPA have occurred during Bininj Manborlh (bush walks) (Stevens et al. 2012), involving multiple generations of Bininj Nawarddeken camping on-Country during a multi-day walk. The plot method was used in these surveys to ensure consistency across the WTGW’s distribution and to standardise surveys across all sites for reporting to the funder. However, a transect-style method may prove more suitable for future presence/non-detection surveys that allow for the on-Country time and intergenerational knowledge transfer that is often the primary goal of Traditional Custodians. Warddeken Land Management has started trialling combining BARs and bushwalking.
Motion cameras, frequently employed by Indigenous ranger groups, have been shown to be a valuable tool (Robinson et al. 2022). Their use not only aligns with the preferences of these groups, but also provides additional data on other native and feral species, contributing significantly to the understanding of wildlife in an under-surveyed region (Stobo-Wilson et al. 2020). The WTGW was detected on only one camera during this project. Nonetheless, other species of importance to Aboriginal peoples were detected and it is beneficial having these images available to provide to Traditional Custodians and their families. The main expense with the equipment (BAR and camera) surveys apart from the initial cost of purchase is the helicopter cost to access sites (>A$1000 per hour) for deployment and retrieval. Therefore, if cameras are available for use, the small additional amount of human hours to deploy a camera array nearby a BAR and for subsequent image analysis is worth undertaking.
Co-benefits
Weaving knowledge co-production into the validation of large niche modelling and species distribution models was integral to the success of this project and could be more readily adapted in Australia (Campbell et al. 2022; Russell et al. 2023). This integration of Traditional ecological knowledge with scientific methods enriches the accuracy and relevance of these models, particularly in diverse Australian ecosystems. The regional engagement and adaptive approach can enable Traditional Custodians to devise co-benefits, such as revitalising traditional ecological knowledge, species-specific practices, and language. Fieldwork teams comprising younger rangers and more senior experienced rangers undertaking the CBP surveys facilitated a direct opportunity for knowledge transfer. This included identifying bird species from both a western science and local Traditional knowledge in language, and identifying the micro-habitat of the target species. Indirectly related to the project, several groups led their own learning on Country activities focused on the collaborative WTGW research, which fostered intergenerational knowledge transfer by uniting senior knowledge holders with younger rangers and children (Hancock 2023).
One of the most profound co-benefits, led by Traditional Custodians, was reconnecting with the land. In post-colonial Arnhem Plateau, challenges such as ‘empty country’ or ‘lonely country’ highlight the difficulty of re-establishing this connection without facilitated access through ecological and cultural programs (Warddeken Land Management 2021). Interactions with Country have personal health and well-being benefits (Sithole et al. 2008) and this connection is crucial for environmental health, as stated in the following:
When we visit Country it recognises our sweat, those mayh [animals or birds] recognise that Country and they come back. [Terrah Guymala]
The actualisation of these co-benefits is vitally dependent on the engagement and leadership of senior Indigenous experts and practitioners, whose profound knowledge and intrinsic connection to their ancestral lands and customs provide a depth of understanding that non-Indigenous practitioners are unable to fully emulate (Campion et al. 2023a, 2023b).
Learning from both western side of knowledge and from Bininj way. We don’t just see them as an animal, we see them deep inside. We see them, the whole being of this bird. They are really important, they play a big role in our life and that’s what we mean we really happy to see them back on this landscape and back where they belong because the connection is there. That means the connection with us, with the land, with the story, the songline, with the dance. All that comes together. [Terrah Guymala]
Conclusions
In this project, we have developed a robust habitat-suitability model for the WTGW, underscoring the inherent value in incorporating knowledge and practices from both science and Traditional perspectives. The inclusivity of meaningful engagement and stakeholder input in validating this model is integral, promoting equitable involvement for all stakeholders. This process also demystified how the model was generated and how it can be used, mitigating the potential for differing interpretations and fostering wider adoption into land management practices. The outcomes of this project have made significant contributions to the biocultural conservation efforts of the WTGW, highlighting the importance of fire management strategies that utilise the imposition of cooler, patchy fires to mitigate extensive severe fire, and through collaboration with Indigenous land-management organisations, preserving the cultural significance of this species.
Data availability
The authors confirm that the traditional and cultural knowledge of Bininj Nawarddeken people supporting the findings of this study is available within the article and can be publicly shared. The ecological data that support findings of this study will be made available upon request to the corresponding author.
Declaration of funding
This project was supported by Territory Natural Resource Management with funding from the Australian Government’s National Landcare program (RLPMU03P1, RLPMU03P10).
Acknowledgements
This project worked across a vast area that encompassed the lands of many clan groups. We thank all the Traditional Owners, Custodians, and community members who were involved in any aspect of the project. We also thank the many other ecological staff who assisted with facilitating planning sessions and on field surveys. The work in this project was conducted under several permits: CDU AEC #A21014; NTG permit to interfere with wildlife # 69265; NLC permits to undertake work on Aboriginal Land #91779, #106932, and #120687, and Kakadu research permit (for Mirrar estate) #RK960.
References
Anderson RP, Raza A (2010) The effect of the extent of the study region on GIS models of species geographic distributions and estimates of niche evolution: preliminary tests with montane rodents (genus Nephelomys) in Venezuela. Journal of Biogeography 37, 1378-1393.
| Crossref | Google Scholar |
Ansell J, Evans J (2020) Contemporary Aboriginal savanna burning projects in Arnhem Land: a regional description and analysis of the fire management aspirations of Traditional Owners. International Journal of Wildland Fire 29, 371-385.
| Crossref | Google Scholar |
Ansell S, Koenig J (2011) CyberTracker: an integral management tool used by rangers in the Djelk Indigenous Protected Area, central Arnhem Land, Australia. Ecological Management & Restoration 12(1), 13-25.
| Crossref | Google Scholar |
Austin BJ, Robinson CJ, Fitzsimons JA, Sandford M, Ens EJ, Macdonald JM, Hockings M, Hinchley DG, McDonald FB, Corrigan C, Kennett R, Hunter-Xenie H, Garnett ST (2018) Integrated measures of Indigenous land and sea management effectiveness: challenges and opportunities for improved conservation partnerships in Australia. Conservation and Society 16(3), 372-384.
| Crossref | Google Scholar |
Bertola LV, Higgie M, Hoskin CJ (2018) Resolving distribution and population fragmentation in two leaf-tailed gecko species of north-east Australia: key steps in the conservation of microendemic species. Australian Journal of Zoology 66, 152-166.
| Crossref | Google Scholar |
Bininj Kunwok Language Centre (2012) Bininj Kunwok Project 2023, Yirlinkirrkkirr. Available at https://bininjkunwok.org.au/yirlinkirrkkirr/ [Accessed 5 October 2023]
Blumstein DT, Mennill DJ, Clemins P, Girod L, Yao K, Patricelli G, Deppe JL, Krakauer AH, Clark C, Cortopassi KA, Hanser SF, McCowan B, Ali AM, Kirschel ANG (2011) Acoustic monitoring in terrestrial environments using microphone arrays: applications, technological considerations and prospectus. Journal of Applied Ecology 48(3), 758-767.
| Crossref | Google Scholar |
Calabrese JM, Certain G, Kraan C, Dormann CF (2014) Stacking species distribution models and adjusting bias by linking them to macroecological models. Global Ecology and Biogeography 23, 99-112.
| Crossref | Google Scholar |
Campbell B, Yirralka Rangers, Yolnu Knowledge Holders, Gallagher RV, Ens EJ (2022) Expanding the biocultural benefits of species distribution modelling with indigenous collaborators: case study from northern Australia. Biological Conservation 274, 109656.
| Crossref | Google Scholar |
Campion OB, Djarrbal M, Ramandjarri C, Malibirr GW, Djigirr P, Dalparri M, Djanbadi D, Malibirr R, Malibirr M, Munuygu E, O’Ryan S, Biridjala F, Bunbuyŋu P, Gulaygulay N, Yalandhu J, Guwankil M, Campion C, Campion B, Bidingal D, Guyula P, Guyula S, Guyula M, Ngurrwuthun E, Truscott H, Bar-Lev Y, Degnian K, Ignjic E, Austin BJ, West S, Waṉambi G (2023a) Monitoring and evaluation of Indigenous Land and Sea Management: an Indigenous-led approach in the Arafura Swamp, northern Australia. Ecological Management & Restoration 24, 75-88.
| Crossref | Google Scholar |
Campion OB, West S, Degnian K, Djarrbal M, Ignjic E, Ramandjarri C, Malibirr GW, Guwankil M, Djigirr P, Biridjala F, O’Ryan S, Austin BJ (2023b) Balpara: a practical approach to working with ontological difference in indigenous land & sea management. Society & Natural Resources 37, 695-715.
| Crossref | Google Scholar |
Darras K, Furnas B, Fitriawan I, Mulyani Y, Tscharntke T (2018) Estimating bird detection distances in sound recordings for standardizing detection ranges and distance sampling. Methods in Ecology and Evolution 9(9), 1928-1938.
| Crossref | Google Scholar |
Darras K, Batáry P, Furnas BJ, Grass I, Mulyani YA, Tscharntke T (2019) Autonomous sound recording outperforms human observation for sampling birds: a systematic map and user guide. Ecological Applications 29(6), e01954.
| Crossref | Google Scholar | PubMed |
Dawson NM, Coolsaet B, Sterling EJ, Loveridge R, Gross-Camp ND, Wongbusarakum S, Sangha KK, Scherl LM, Phuong Phan H, Zafra-Calvo N, Lavey WG, Byakagaba P, Idrobo CJ, Chenet A, Bennett NJ, Mansourian S, Rosado-May FJ (2021) The role of Indigenous peoples and local communities in effective and equitable conservation. Ecology and Society, Resilience Alliance 26(3), 19.
| Crossref | Google Scholar |
Edwards A, Archer R, De Bruyn P, Evans J, Lewis B, Vigilante T, Whyte S, Russell-Smith J (2021) Transforming fire management in northern Australia through successful implementation of savanna burning emissions reductions projects. Journal of Environmental Management 290, 112568.
| Crossref | Google Scholar | PubMed |
Einoder LD, Southwell DM, Lahoz-Monfort JJ, Gillespie GR, Fisher A, Wintle BA (2018) Occupancy and detectability modelling of vertebrates in northern Australia using multiple sampling methods. PLoS ONE 13, e0203304.
| Crossref | Google Scholar | PubMed |
Elith J, Phillips SJ, Hastie T, Dudík M, Chee YE, Yates CJ (2011) A statistical explanation of MaxEnt for ecologists. Diversity and Distributions 17, 43-57.
| Crossref | Google Scholar |
Ens EJ (2012) Monitoring outcomes of environmental service provision in low socio-economic indigenous Australia using innovative CyberTracker Technology. Conservation and Society 10(1), 42-52.
| Crossref | Google Scholar |
Ens EJ, Turpin G (2022) Synthesis of Australian cross-cultural ecology featuring a decade of annual Indigenous ecological knowledge symposia at the Ecological Society of Australia conferences. Ecological Management and Restoration 23, 3-16.
| Crossref | Google Scholar |
Evans J, Russell-Smith J (2019) Delivering effective savanna fire management for defined biodiversity conservation outcomes: an Arnhem Land case study. International Journal of Wildland Fire 29, 386-400.
| Crossref | Google Scholar |
Fisher DO, Johnson CN, Lawes MJ, Fritz SA, McCallum H, Blomberg SP, VanDerWal J, Abbott B, Frank A, Legge S, Letnic M, Thomas CR, Fisher A, Gordon IJ, Kutt A (2014) The current decline of tropical marsupials in Australia: is history repeating? Global Ecology and Biogeography 23, 181-190.
| Crossref | Google Scholar |
Frank ASK, Johnson CN, Potts JM, Fisher A, Lawes MJ, Woinarski JCZ, Tuft K, Radford IJ, Gordon IJ, Collis M-A, Legge S (2014) Experimental evidence that feral cats cause local extirpation of small mammals in Australia’s tropical savannas. Journal of Applied Ecology 51, 1486-1493.
| Crossref | Google Scholar |
Gallant J, Austin J (2012) Topographic wetness index derived from 1" SRTM DEM-H. v2. Available at https://doi.org/10.4225/08/57590B59A4A08
Gallant J, Dowling T, Austin J (2012) Multi-resolution Valley Bottom Flatness (MrVBF) v.3. Available at https://doi.org/10.4225/08/5701C885AB4FE
Garnett ST, Burgess ND, Fa JE, Fernández-Llamazares Á, Molnár Z, Robinson CJ, Watson JEM, Zander KK, Austin B, Brondizio ES, Collier NF, Duncan T, Ellis E, Geyle H, Jackson MV, Jonas H, Malmer P, McGowan B, Sivongxay A, Leiper I (2018) A spatial overview of the global importance of Indigenous lands for conservation. Nature Sustainability 1(7), 369-374.
| Crossref | Google Scholar |
Ghanbari S, Turvey ST (2022) Local ecological knowledge provides novel evidence on threats and declines for the Caucasian grouse Lyrurus mlokosiewiczi in Arasbaran Biosphere Reserve, Iran. People and Nature 4(6), 1536-1546.
| Crossref | Google Scholar |
Gomes VHF, Ijff SD, Raes N, Amaral IL, Salomão RP, et al. (2018) Species distribution modelling: contrasting presence-only models with plot abundance data. Scientific Reports 8, 1003.
| Crossref | Google Scholar | PubMed |
Guerschman JP, Hill MJ, Renzullo LJ, Barrett DJ, Marks AS, Botha EJ (2009) Estimating fractional cover of photosynthetic vegetation, non-photosynthetic vegetation and bare soil in the Australian tropical savanna region upscaling the EO-1 Hyperion and MODIS sensors. Remote Sensing of Environment 113, 928-945.
| Crossref | Google Scholar |
Hancock D (2023) Yirlinkirrkirr baleh? Cosmos Magazine 101, 42-47 Available at https://cosmosmagazine.com/nature/birds/yirlinkirrkirr-baleh-grasswren/ [Accessed December 2023].
| Google Scholar |
Harrington GN, Murphy SA (2016) The distribution and conservation status of Carpentarian grasswrens (Amytornis dorotheae), with reference to prevailing fire patterns. Pacific Conservation Biology 21, 291-297.
| Crossref | Google Scholar |
Jacobsen R, Howell C, Read SM (2020) Australia’s Indigenous land and forest estate: separate reporting of Indigenous ownership, management and other special rights ABARES technical report. (ABARES: Canberra, ACT, Australia) Available at https://doi.org/10.25814/bqr0-4m20 [Accessed 12 September 2023]
Kukutai T (2024) How Indigenous communities in New Zealand are protecting their data. Science 384(6691), eado9298.
| Crossref | Google Scholar |
Kwok R (2019) AI empowers conservation biology. Nature 567(7746), 133-134.
| Crossref | Google Scholar | PubMed |
Lawes MJ, Murphy BP, Fisher A, Woinarski JCZ, Edwards AC, Russell-Smith J (2015) Small mammals decline with increasing fire extent in northern Australia: evidence from long-term monitoring in Kakadu National Park. International Journal of Wildland Fire 24, 712-722.
| Crossref | Google Scholar |
Legge S, Kennedy MS, Lloyd R, Murphy SA, Fisher A (2011) Rapid recovery of mammal fauna in the central Kimberley, northern Australia, following the removal of introduced herbivores. Austral Ecology 36, 791-799.
| Crossref | Google Scholar |
Lilleyman A, Pascoe J, Robinson CJ, Legge SM, Woinarski JCZ, Garnett ST (2024) Imperilled birds and First Peoples’ land and sea Country in Australia. Emu – Austral Ornithology 124(1), 108-122.
| Crossref | Google Scholar |
McKemey MB, Banbai Rangers, Ens EJ, Hunter JT, Ridges M, Costello O, Reid NCH (2021) Co-producing a fire and seasons calendar to support renewed Indigenous cultural fire management. Austral Ecology 46(7), 1011-1029.
| Crossref | Google Scholar |
Molloy SW, Davis RA, Dunlop JA, van Etten EJB (2017) Applying surrogate species presences to correct sample bias in species distribution models: a case study using the Pilbara population of the Northern Quoll. Nature Conservation 18, 25-46.
| Crossref | Google Scholar |
Moore HA, Dunlop JA, Valentine LE, Woinarski JCZ, Ritchie EG, Watson DM, Nimmo DG (2019) Topographic ruggedness and rainfall mediate geographic range contraction of a threatened marsupial predator. Diversity and Distributions 25, 1818-1831.
| Crossref | Google Scholar |
Naimi B, Hamm NAS, Groen TA, Skidmore AK, Toxopeus AG (2014) Where is positional uncertainty a problem for species distribution modelling? Ecography 37, 191-203.
| Crossref | Google Scholar |
NESP TSR Hub (2019) Threatened species strategy year 3 scorecard – white-throated grasswren. (Australian Government: Canberra, ACT, Australia) Available at http://www.environment.gov.au/biodiversity/threatened/species/20-birds-by-2020/whitethroated-grasswren [Accessed 10 August 2023]
Noske RA (1992) The status and ecology of the white-throated grasswren Amytornis woodwardi. Emu 92, 39-51.
| Crossref | Google Scholar |
Paltridge R, Ward NN, West JT, Crossing K (2020) Is cat hunting by Indigenous tracking experts an effective way to reduce cat impacts on threatened species? Wildlife Research 47(8), 709-719.
| Crossref | Google Scholar |
Perry J, Fisher A, Palmer C (2011) Status and habitat of the Carpentarian grasswren (Amytornis dorotheae) in the Northern Territory. Emu – Austral Ornithology 111, 155-161.
| Crossref | Google Scholar |
Phillips SJ, Anderson RP, Schapire RE (2006) Maximum entropy modeling of species geographic distributions. Ecological Modelling 190, 231-259.
| Crossref | Google Scholar |
Phillips SJ, Dudík M, Elith J, Graham CH, Lehmann A, Leathwick J, Ferrier S (2009) Sample selection bias and presence-only distribution models: implications for background and pseudo-absence data. Ecological Applications 19, 181-197.
| Crossref | Google Scholar | PubMed |
Phillips SJ, Anderson RP, Dudík M, Schapire RE, Blair ME (2017) Opening the black box: an open-source release of Maxent. Ecography 40, 887-893.
| Crossref | Google Scholar |
Price O, Russell-Smith J, Edwards A (2003) Fine-scale patchiness of different fire intensities in sandstone heath vegetation in northern Australia. International Journal of Wildland Fire 12, 227-236.
| Crossref | Google Scholar |
Radford IJ, Gibson LA, Corey B, Carnes K, Fairman R (2015) Influence of fire mosaics, habitat characteristics and cattle disturbance on mammals in fire-prone savanna landscapes of the northern Kimberley. PLoS ONE 10, e0130721.
| Crossref | Google Scholar | PubMed |
Radford IJ, Woolley L-A, Corey B, Vigilante T, Wunambal Gaambera Aboriginal Corporation, Hatherley E, Fairman R, Carnes K, Start AN (2020) Prescribed burning benefits threatened mammals in northern Australia. Biodiversity and Conservation 29, 2985-3007.
| Crossref | Google Scholar |
Renwick AR, Robinson CJ, Garnett ST, Leiper I, Possingham HP, Carwardine J (2017) Mapping Indigenous land management for threatened species conservation: an Australian case-study. PLoS ONE 12(3), e0173876.
| Crossref | Google Scholar | PubMed |
Ritchie D (2009) Things fall apart: the end of an era of systematic Indigenous fire management (chapter 2). In ‘Culture, ecology and economy of fire management in North Australian Savannas: rekindling the Wurrk tradition’. (Eds J Russell-Smith, P Whitehead, P Cooke) pp. 23–40. (CSIRO Publishing: Melbourne, Vic, Australia).
Robinson CJ, Macdonald JM, Douglas M, Perry J, Setterfield S, Cooper D, Lee M, Nadji J, Nadji S, Nayinggul A, Nayinggul A, Mangiru K, Hunter F, Coleman B, Barrowei R, Markham J, Alderson J, Moyle F, May K, Bangalang N-G (2022) Using knowledge to care for country: indigenous-led evaluations of research to adaptively co-manage Kakadu National Park, Australia. Sustainability Science 17, 377-390.
| Crossref | Google Scholar |
Robinson CJ, Urzedo D, Macdonald JM, Ligtermoet E, Penton CE, Lourie H, Hoskins A (2023) Place-based data justice practices for collaborative conservation research: a critical review. Biological Conservation 288, 110346.
| Crossref | Google Scholar |
Russell SR, Sultana R, Ngukurr Yangbala Rangers, Ens EJ (2023) Mepimbat tedul proujek: Indigenous knowledge of culturally significant freshwater turtles addresses species knowledge gaps in northern Australia. Austral Ecology 48(7), 1306-1327.
| Crossref | Google Scholar |
Russell-Smith J, Ryan PG, Klessa D, Waight G, Harwood R (1998) Fire regimes, fire-sensitive vegetation and fire management of the sandstone Arnhem Plateau, monsoonal northern Australia. Journal of Applied Ecology 35, 829-846.
| Crossref | Google Scholar |
Russell-Smith J, Cook GD, Cooke PM, Edwards AC, Lendrum M, Meyer CP, Whitehead PJ (2013) Managing fire regimes in north Australian savannas: applying Aboriginal approaches to contemporary global problems. Frontiers in Ecology and the Environment 11, e55-e63.
| Crossref | Google Scholar |
Russell-Smith J, Yates CP, Edwards AC, Whitehead PJ, Murphy BP, Lawes MJ (2015) Deriving multiple benefits from carbon market-based savanna fire management: an Australian example. PLoS ONE 10, e0143426.
| Crossref | Google Scholar | PubMed |
Schodde R, Mason IJ (1975) Occurrence, nesting and affinities of the white-throated grass-wren Amytornis woodwardi and white-lined honeyeater Meliphaga albilineata. Emu – Austral Ornithology 75, 12-18.
| Crossref | Google Scholar |
Searcy CA, Shaffer HB (2016) Do ecological niche models accurately identify climatic determinants of species ranges? The American Naturalist 187, 423-435.
| Crossref | Google Scholar | PubMed |
Sharma K, Fiechter M, George T, Young J, Alexander JS, Bijoor A, Suryawanshi K, Mishra C (2020) Conservation and people: towards an ethical code of conduct for the use of camera traps in wildlife research. Ecological Solutions and Evidence 1(2), e12033.
| Crossref | Google Scholar |
Skroblin A, Carboon T, Bidu G, Taylor M, Bidu N, Taylor W, Taylor K, Miller M, Robinson L, Williams C, Chapman N, Marney M, Marney C, Biljabu J, Biljabu L, Jeffries P, Samson H, Charles P, Game ET, Wintle B (2022) Developing a two-way learning monitoring program for Mankarr (Greater Bilby) in the Western Desert, Western Australia. Ecological Management & Restoration 23, 129-138.
| Crossref | Google Scholar |
Stobo-Wilson AM, Stokeld D, Einoder LD, Davies HF, Fisher A, Hill BM, Mahney T, Murphy BP, Scroggie MP, Stevens A, Woinarski JCZ, Bawinanga Rangers, Warddeken Rangers, Gillespie GR (2020) Bottom-up and top-down processes influence contemporary patterns of mammal species richness in Australia’s monsoonal tropics. Biological Conservation 247, 108638.
| Crossref | Google Scholar |
Stoetzel HJ, Leseberg NP, Murphy SA, Andrew ME, Plant KJ, Harrington GN, Watson JEM (2020) Modelling the habitat of the endangered Carpentarian Grasswren (Amytornis dorotheae): the importance of spatio-temporal habitat availability in a fire prone landscape. Global Ecology and Conservation 24, e01341.
| Crossref | Google Scholar |
Syfert MM, Smith MJ, Coomes DA (2013) The effects of sampling bias and model complexity on the predictive performance of MaxEnt species distribution models. PLoS ONE 8, e55158.
| Crossref | Google Scholar | PubMed |
Tynan L (2021) What is relationality? Indigenous knowledges, practices and responsibilities with kin. Cultural Geographies 28(4), 597-610.
| Crossref | Google Scholar |
UNEP (2022) ‘Decision adopted by the conference of the parties to the convention on biological diversity: Kunming–Montreal Global Biodiversity Framework.’ (United Nations Environment Programme: Montreal, Canada) Available at https://www.cbd.int/doc/decisions/cop-15/cop-15-dec-04-en.pdf
Venables WN, Ripley BD (Eds) (2002) ‘Modern applied statistics with S.’ 4th edn. (Springer-Verlag: New York, NY, USA) https://doi.org/10.1007/978-0-387-21706-2
Verdon SJ, Watson SJ, Clarke MF (2019) Modeling variability in the fire response of an endangered bird to improve fire-management. Ecological Applications 29(8), e01980.
| Crossref | Google Scholar |
von Takach B, Scheele BC, Moore H, Murphy BP, Banks SC (2020) Patterns of niche contraction identify vital refuge areas for declining mammals. Diversity and Distributions 26, 1467-1482.
| Crossref | Google Scholar |
Warddeken Land Management (2020) Warddeken annual report 2019–20. Available at https://www.warddeken.org.au/_files/ugd/982814_5ecf6390c85d4a7dad781c763fc65940.pdf
Warddeken Land Management (2021) Plan of management 2021–2025. Available at https://www.warddeken.org.au/_files/ugd/982814_acc7190430c24f9e88b283518be2fc3e.pdf [Accessed 13 November 2023]
Warddeken Land Management (in press) Warddeken annual report 2022-2023. Available at https://www.warddeken.org.au/about [Accessed 13 November 2023]
Ward-Fear G, Shine R (2019) How Indigenous expertise improves science: the curious case of shy lizards and deadly cane toads. In The Conversation, 9 April 2019. Available at https://theconversation.com/how-indigenous-expertise-improves-science-the-curious-case-of-shy-lizards-and-deadly-cane-toads-113997
Wearn OR, Freeman R, Jacoby DMP (2019) Responsible AI for conservation. Nature Machine Intelligence 1(2), 72-73.
| Crossref | Google Scholar |
Woinarski J (1992) The conservation status of the white-throated grasswren Amytornis woodwardi, an example of problems in status designation. Northern Territory Naturalist 13, 1-5.
| Crossref | Google Scholar |
Woinarski JCZ, Ash AJ (2002) Responses of vertebrates to pastoralism, military land use and landscape position in an Australian tropical savanna. Austral Ecology 27, 311-323.
| Crossref | Google Scholar |
Woinarski JCZ, Milne DJ, Wanganeen G (2001) Changes in mammal populations in relatively intact landscapes of Kakadu National Park, Northern Territory, Australia. Austral Ecology 26, 360-370.
| Crossref | Google Scholar |
Woinarski JCZ, Armstrong M, Brennan K, Fisher A, Griffiths AD, Hill B, Milne DJ, Palmer C, Ward S, Watson M, Winderlich S, Young S (2010) Monitoring indicates rapid and severe decline of native small mammals in Kakadu National Park, northern Australia. Wildlife Research 37, 116.
| Crossref | Google Scholar |
Woinarski JCZ, Legge S, Fitzsimons JA, Traill BJ, Burbidge AA, Fisher A, Firth RSC, Gordon IJ, Griffiths AD, Johnson CN, McKenzie NL, Palmer C, Radford I, Rankmore B, Ritchie EG, Ward S, Ziembicki M (2011) The disappearing mammal fauna of northern Australia: context, cause, and response. Conservation Letters 4, 192-201.
| Crossref | Google Scholar |
Woinarski JCZ, Fisher A, Armstrong M, Brennan K, Griffiths AD, Hill B, Choy JL, Milne D, Stewart A, Young S, Ward S, Winderlich S, Ziembicki M (2012) Monitoring indicates greater resilience for birds than for mammals in Kakadu National Park, northern Australia. Wildlife Research 39, 397-407.
| Crossref | Google Scholar |
Wysong ML, Gregory P, Watson AWT, Woolley L-A, Parker CW, Yawuru Country Managers, Karajarri Rangers, Nyikina Mangala Rangers (2022) Cross-cultural collaboration leads to greater understanding of the rare Spectacled Hare-wallaby in the west Kimberley, Western Australia. Ecological Management & Restoration 23(S1), 139-149.
| Crossref | Google Scholar |
Yates CP, Edwards AC, Russell-Smith J (2008) Big fires and their ecological impacts in Australian savannas: size and frequency matters. International Journal of Wildland Fire 17, 768-781.
| Crossref | Google Scholar |