

Ground-based counting methods underestimate true numbers of a threatened colonial mammal: an evaluation using drone-based thermal surveys as a reference
Eliane D. McCarthy A B * , John M. Martin C , Matthias M. Boer A and Justin A. Welbergen A
A B * , John M. Martin C , Matthias M. Boer A and Justin A. Welbergen A
A The Hawkesbury Institute for the Environment, Western Sydney University, Richmond, NSW 2753, Australia.
B Present address: School of Life and Environmental Sciences, The University of Sydney, Camperdown, NSW 2006, Australia.
C Institute of Science and Learning, Taronga Conservation Society Australia, Mosman, NSW 2088, Australia.
Wildlife Research 50(6) 484-493 https://doi.org/10.1071/WR21120
Submitted: 11 August 2021 Accepted: 5 June 2022 Published: 4 August 2022
© 2023 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Context: Accurate and precise monitoring practises are key for effective wildlife conservation management; providing reliable estimates of spatiotemporal changes in species abundance on which sound decision-making can be based. Advancements in drone and satellite technology are providing new standards for survey accuracy and precision and have great potential for enhancing population monitoring of numerous difficult to survey species. Flying-foxes (Pteropus spp.) are large bats that roost in groups of a few hundred to many thousands in the canopies of trees, where they are difficult to census accurately and precisely by human observers. Globally, 35 of the 64 flying-fox species are listed as threatened under the IUCN Red List of Threatened Species, and reliable monitoring methods are needed for the effective management of this ecologically important group.
Aims: Recently, we showed that drone-acquired thermal imagery can be used to count flying-foxes in their roost with high accuracy and precision. In the present study, we aimed to assess the accuracy and precision of whole colony counts derived from ground-based counting methods against reference counts derived from drone-acquired thermal imagery.
Methods: We evaluated the relationship between ground-based counts by two groups of human observers to highly accurate and precise counts derived from drone-acquired thermal orthomosaics for 25 counts conducted across seven flying-fox roosts throughout the Greater Sydney region, Australia.
Key results: We found that ground-based counts by human observers were positively correlated with those obtained from concurrent drone-acquired thermal imagery. However, drone-acquired estimates of colony size were 2.05 and 1.92 times higher than ground-based counts by the experimenter and Australian government counters respectively. When compared against drone-acquired reference counts, the precision (coefficient of variation) of ground-based counts was 26.3% when conducted by a single counter and 55.1% when conducted by multiple counters.
Conclusions: Our research indicates that ground-based counting methods underestimate true population sizes by substantial margins and have limited precision. Drone-based monitoring provides highly accurate and precise population estimates, and thus is expected to yield more reliable information on flying-fox abundance and allow for trends to be established over shorter timescales.
Implications: Using ground counting methods alone, population trends can only be established with significance after protracted periods of monitoring. Incorporating the use of thermal drones into current monitoring practises would enhance the capacity to detect population trends earlier and more accurately, so that conservation management can more effectively respond.
Keywords: bat, Chiroptera, colonial animal, drone, infrared imagery, RPAS, survey error, thermal, wildlife surveys.
Introduction
Knowledge of the distribution, abundance and density of animal populations is key for conservation and management (Marsh and Trenham 2008). Critically, monitoring species over time can elucidate trends and alert researchers and policy makers to species declines, so that conservation measures can be taken (Goldsmith 1991; Sutherland 2006; Westcott et al. 2012; Woinarski 2018). However, population survey methods are subject to many sources of error, including but not limited to site accessibility (Wagner 1981; Reddy and Dávalos 2003), animal detectability (Otto and Pollock 1990), and between-observer variability (Erwin 1982). A fundamental limitation of imprecise counting methods is the potential failure to detect changes in species population trajectory, potentially with detrimental implications for species conservation and management (Elphick 2008; McKelvey et al. 2008; Martin et al. 2012).
With populations of known size, researchers have previously been able to test the accuracy and precision of ground-based observer methods (Elphick 2008). For example, using a physical model of a wading bird colony it was found that observer counts were 29% lower than true counts on average, and undercounting occurred for 81% of estimates, with high variation between and within individual counters, irrespective of experience (Frederick et al. 2003). Conversely, distance sampling methods have been shown to produce large positive biases in predicted population density as a result of counting errors (Alldredge et al. 2008). Therefore, emerging technologies are increasingly being employed with the aim to collect more accurate and precise wildlife monitoring data (Fretwell et al. 2012; Cilulko et al. 2013; Wich and Koh 2018).
Thermal cameras detect radiation emitted by animals in the thermal infrared region of the electromagnetic spectrum, and infrared technology has been used for remotely censusing animals for more than half a century (Croon et al. 1968). Advances in infrared technology and improvements in the resolution and portability of thermal cameras allow for surveillance of increasingly smaller animals and for surveys to be conducted further from target species, minimising disturbance (Cilulko et al. 2013; Chabot and Bird 2015). Infrared population surveys have now been conducted for many species, such as elk (Cervus elaphus) (Dunn et al. 2002), using thermal sensor equipped manned aircraft, and the greater glider (Petauroides volans) (Vinson et al. 2020) and Brazilian free-tailed bats (Tadarida brasiliensis) (Betke et al. 2008) using ground mounted thermal sensors.
Remotely piloted aircraft systems (RPAS; hereafter ‘drones’) equipped with thermal sensors enable researchers to collect high resolution imagery closer to target animals than is possible by piloted aircraft (Anderson and Gaston 2013). Population counts conducted from drone-acquired imagery in the visible spectrum have been shown to be more accurate and precise than ground counts of target species (Hodgson et al. 2018; Witt et al. 2020). Drones equipped with thermal sensors have been used to monitor mammal species such as grey seals (Halichoerus grypus) (Seymour et al. 2017) and common hippopotami (Hippopotamus amphibius). In Australia, drones have been used to monitor the arboreal koala (Phascolarctos cinereus) (Witt et al. 2020; Beranek et al. 2021). Importantly, these studies show that drone-based thermal surveys can improve the detection and quantification of less conspicuous and inaccessible animal populations.
The family Pteropodidae comprises approximately 200 Old-World fruit bat species, distributed throughout Africa, the eastern Mediterranean, south and southeast Asia, Australia and the Pacific Islands (Simmons 2020). Within these regions, species in this family perform key roles as pollinators and plant propagators (Fujita and Tuttle 1991; Aziz et al. 2021). One hundred and nine of the species in the Pteropodidae family are considered threatened or data deficient according to the IUCN Red List of Threatened Species (International Union for Conservation of Nature and Natural Resources (IUCN) 2020). Threats are wide ranging and include overhunting (Mildenstein et al. 2016), climate change (Welbergen et al. 2008; Dey et al. 2015) and habitat loss (Mohd-Azlan et al. 2001). Within the Pteropodidae, flying-foxes of the genus Pteropus tend to rest by day in arboreal roosts that may contain colonies of many thousands of individuals (Hall and Richards 2000) with complex social dynamics and architecture (Welbergen 2005; Klose et al. 2009a, 2009b). However, due to the physical structure and inaccessibility of their roosts, flying-fox colonies are difficult to survey with accuracy and precision (Westcott et al. 2015). This is important as 35 of the 64 Pteropus species globally are listed as threatened with extinction (International Union for Conservation of Nature and Natural Resources (IUCN) 2020).
In mainland Australia, the grey-headed flying-fox (Pteropus poliocephalus) and the spectacled flying-fox (Pteropus conspicillatus) are listed respectively as ‘Vulnerable’ and ‘Endangered’ on the IUCN Red List and under the Australian Environment Protection and Biodiviersity Conservation Act 1999 (Commonwealth of Australia Department of Environment and Energy 2019a; Roberts et al. 2020; Eby et al. 2021). Like some other Pteropus species, the grey-headed and spectacled flying-fox can now increasingly be found in urban and peri-urban habitats (Tait et al. 2014; Meade et al. 2021; Timmiss et al. 2021; Yabsley et al. 2021), which exposes them to a range of novel threats including human-wildlife conflict, electrocution on power lines and entanglement in fruit tree netting in gardens (Mo et al. 2021). Australian flying-foxes are also susceptible to extreme heat stress events, notably, an extreme heat event in November 2018 that killed ~23 000 spectacled flying-foxes, representing approximately one-third of the species’ population in Australia at the time (Mao 2019). Given the threats to and status of Pteropus species in Australia and elsewhere, rigorous long-term population monitoring is warranted.
It is well established that the various ground based flying-fox counting methods have their limitations. By comparing live fly-out counts to reference counts from slowed down recorded footage of fly-outs, research has shown that fly-out counter error decreases with observer experience, and increases with increasing width, speed and number of animals in fly out streams (Westcott and McKeown 2004; Forsyth et al. 2006). Furthermore, live fly-out counts can both under and overestimate the true number of animals in a fly-out stream (Forsyth et al. 2006). To date, there has been less focus on evaluating the sources of error for ground counting, especially considering no methods are currently available for determining the true abundances of flying-foxes in their roosts. Prior work acknowledges that ground counts can be impractical if the colony is inaccessible or situated in dense vegetation (Eby et al. 1999). Furthermore, counters passing through the colony conducting ground counts are known to cause a high degree of disturbance to flying-foxes, which would also affect ground count accuracy (Westcott et al. 2011). Despite this, ground and fly-out counts remain widely used for flying-fox population monitoring.
Australia’s National Flying-Fox Monitoring Program (NFFMP), is a government-run programme that aims to survey all known flying-fox roosts quarterly to generate population estimates, by either ground or fly-out counts, to establish long term population trends, particularly of the grey-headed and spectacled flying-fox (Westcott et al. 2015). While data collected quarterly under the NFFMP do not provide accurate estimates of the absolute national population sizes of the species, they are meaningful indicators of long-term population trends. Nevertheless, limited precision, combined with the low quarterly survey rates and overall inconsistency in survey efforts, means that population trends can only be established with significance after many years of monitoring (e.g. Westcott et al. 2012) limiting the capacity for conservation management to respond quickly and proactively to threats.
Recently, a method was developed for quantifying the abundance of flying-fox in their colonies, through counting individuals in drone-acquired thermal orthomosaics (McCarthy et al. 2021). The method was shown to yield highly accurate and precise estimates of flying-fox abundance at roosts, and without significant effects of between-observer variability (McCarthy et al. 2021), thus potentially greatly enhancing the capacity for monitoring to detect significant population trends over shorter timeframes. In the present study we use colony size estimates obtained from drone-acquired thermal orthomosaics as a reference to assess the accuracy and precision of ground counts conducted by the experimenter as well as by NFFMP counters over 13 months. By directly comparing paired sets of ground counts to highly accurate and precise reference counts derived from drone-acquired thermal orthomosaics, we measure the error associated with ground counts.
Materials and methods
Study area
This study was conducted within the Greater Sydney Region in south-eastern Australia (Fig. 1). The remnant vegetation within this urban/peri-urban area consisted mainly of Eucalypt open forests and woodlands, with an understory of grass, shrubs, ferns, and herbs (Geoscience Australia 2001). This region comprises approximately 30 known flying-fox roosts (Commonwealth of Australia Department of Environment and Energy 2019b) that were continually or periodically occupied by the grey-headed flying-fox, the black flying-fox (P. alecto) and the little-red flying-fox (P. scapulatus) (E. McCarthy, Western Sydney University, personal observation). Within the study area, the grey-headed flying-fox is the most common species by far but black flying-foxes (Pteropus alecto) and little red flying-foxes (Pteropus scapulatus) can be present as well.

|
Drone survey procedure
A DJI Inspire 1 ver. 2.0 drone (DJI, Shenzhen, China) equipped with Zenmuse XT 19 mm radiometric thermal camera (DJI, Shenzhen, China) was used to conduct all surveys. The drone was fitted with one lithium polymer battery (TB48, 130.0 Wh), giving a maximum flight duration of approximately 15 min. The FLIR longwave infrared thermal sensor in the Zenmuse XT has a sensitivity of 50 mK @nf/1.0 at a resolution of 640 × 520 pixels.
For this study, flying-fox roosts were selected based on the airspace regulations for flying drones, their accessibility for safe flights (the drone needed to be visible at all times while in flight, in keeping with Australian regulations), and access for ground counting. Colony-wide drone surveys were conducted between October 2019 and August 2020 at seven roosts including: Camellia Gardens (n = 3 surveys), Campbelltown (n = 5), Emu Plains (n = 2), Kareela (n = 5), Macquarie Fields (n = 2), Warriewood (n = 1), and Yarramundi (n = 7) (shown in Fig. 1; exact locations given in Supplementary material Table S1). These roosts were predominantly inhabited by grey-headed flying-foxes but have been known to have lower numbers of black flying-foxes and little red flying-foxes (Commonwealth of Australia Department of Environment and Energy 2019b).
All flights were mapped in the Pix4D Capture iOS application (Pix4D, 2017) prior to flight. Flight plan mapping was conducted on location, after a visual inspection of the extent of the flying-fox colony. Drone surveys commenced in the early morning, between 0 and 2 h after sunrise, with air temperature ranging between 10.3 and 24.3°C (median air temperature was sourced from nearest weather station; Bureau of Meteorology 2020), in low wind conditions (<10 km/h; data sourced from nearest weather station). The drone was launched from a cleared area, at a minimum of 20 m from the colony, which caused no visible disturbance to the roosting flying-foxes. During flight, the drone flew at a speed of 2 m/s and captured images with a forward and lateral overlap of 90% in a lawnmower pattern. For larger colonies where the battery life of the drone was not sufficient to complete the entire survey on a single battery, the drone was returned to base and the batteries changed midway through the survey (up to five battery changes were required for colonies with a large area (up to ~1.5 ha), with changes taking <5 min each) after which the drone resumed the survey from the point at which the last thermal image was taken. Following drone surveys, all survey images were saved as radiometric JPEGs, with embedded EXIF data providing latitude, longitude, and altitude. Agisoft Metashape Professional ver. 1.5 was used to generate orthomosaics (Supplementary material Table S2; orthomosaics are viewable in Supplementary material 2, available at https://figshare.com/s/00084845b1e3c628f9d4) (LLC Agisoft 2019). Flying-foxes in orthomosaics were then manually counted by the experimenter using Fiji 1.8.0_172 (Schneider et al. 2012; McCarthy et al. 2021). Previously, there was shown to be a high concordance between counts derived from drone-acquired imagery and exhaustive visual counts of flying-foxes in single trees, demonstrating the high accuracy of counts derived from drone-acquired imagery (McCarthy et al. 2021). Furthermore, the precision of counts from drone-acquired thermal orthomosaics was very high (6.8 ± 6.0%; McCarthy et al. 2021). Therefore, for the purposes of the present study, the counts derived from thermal orthomosaics were assumed to provide ‘true’ counts of the number of flying-foxes present in colonies, and counts derived from thermal orthomosaics were used as a reference to assess the accuracy and precision of ground counts. In addition, counts derived from thermal orthomosaics have been shown to be highly precise and not subject to observer bias (McCarthy et al. 2021); therefore, in the present study, a single trained observer conducted all counts of the number of flying-foxes in thermal orthomosaics.
Animal ethics considerations
In this study, the drone was flown at a height at least 30 m above roosting flying-foxes over a period of up to 2 h for each survey, and our visual observations indicated that the presence of the drone did not disturb the roosting flying-foxes. Prior to commencing surveys, we established a disturbance threshold, whereby if more than 20 individuals took flight for more than 1 min, we would end a survey. However, this threshold was never reached. During the surveys, flying-foxes remained where they were roosting while the drone flew above them, and did not seek to leave the roost or to relocate within the roost.
Ground count procedure
In Australia, daytime ground counts were used to conduct 94% the NFFMP quarterly counts between February 2018 and February 2020 (A. McKeown, Commonwealth Scientific and Industrial Research Organisation, pers. comm.). Hence, the focus of our study was on assessing the performance of this method against counts from drone surveys.
In each of the seven roosts, the experimenter conducted a ground count estimate of the total number of flying-foxes on the same day as each drone survey, replicating the procedures routinely performed by NFFMP counters. Ground-based counts are necessarily not exhaustive, as roosts are made up of 10–100 of trees over a large area (ranging from 0.13 to 1.48 ha in this study; Supplementary material Table S1) with many roosting individuals obscured by foliage; and exhaustive, single tree based counts as conducted by McCarthy et al. (2021), are generally not an option for NFFMP counters due to time and accessibility constraints and the risk of disturbing roosting individuals. Therefore, ground based counts rely on counters extrapolating an estimate for the total number of individuals in a colony, as described below. For the Camellia Gardens colony, where flying-foxes exhibited a high tolerance to human presence, the experimenter slowly walked through the accessible areas of the colony counting all visible individuals, recording each lot of 10 sighted individuals using a hand tally counter. All other roosts were either difficult to access or entering would have caused a high degree of disturbance to the colony of flying-foxes. Therefore, for these roosts, whole or part of the colony boundary was traversed, and counts were performed from positions where animals were visible. Following this, the proportion of the colony area surveyed was estimated, and a figure for the total number of flying-foxes in the roost extrapolated based on the partial count for the colony and the area which was not visible. Note, this is one of the formal counting methods used routinely as part of the NFFMP (Westcott et al. 2011).
For 16 of the 25 surveys, within 7 days (average 2 days) of each drone survey and accompanying ground count by the experimenter, another ground count was conducted as part of the NFFMP quarterly counts (NFFMP surveyed colonies listed in Supplementary material Table S1; Westcott et al. 2015). Of the 16 NFFMP counts which contributed to this study, five counters conducted to these counts.
Statistical analyses
All statistical analyses were two-tailed, employed an α value of 0.05, and were conducted in R (R Core Team 2018) interfaced via RStudio Desktop (v. 1.2.5042). General linear mixed models (GLMMs) were constructed using the nlme package (Pinheiro et al. 2022). All models had a Gaussian distribution and were fitted with an identity link function. We standardised response and predictor variables using the standardise function in the arm package (Gelman 2008; Gelman et al. 2021). First, GLMMs were fit to determine the relationship between colony size estimates from drone-acquired orthomosaics (reference counts) and ground counts conducted by the experimenter and NFFMP, to determine the accuracy of ground counts. Two GLMMs were fit with (1) the experimenter ground count, and (2) NFFMP ground count as the response variable in each model, and thermal image point count as the fixed effect. Both models had roost ID (colony location) as a random effect. For each model, the effect of the response variable was tested through a likelihood ratio test (ANOVA). The significance of the random effect was tested through parametric bootstrapping with one thousand iterations. For GLMM 2), we conducted the analysis both with and without an extreme outlier, the Emu Plains NFFMP ground count from 24 February 2020, where the drone-acquired thermal orthomosaic count was 21.4 times higher than the corresponding NFFMP ground count. To provide a measure of precision for the ground counts, the coefficient of variation (CV) of the response variable was calculated for each model, using the sjstats package (Lüdecke 2019).
Then, we assessed the relationship between ground counts by the experimenter and ground counts by NFFMP counters, by fitting a GLMM with NFFMP ground count as the response variable, and experimenter ground count as a fixed effect, with roost ID as a random effect.
This research was approved by Western Sydney University Animal Research Authority no. A12217 and NPWS scientific licence SL102047.
Results
The numbers of flying-foxes counted roosting in colonies ranged between 550 and 7290 individuals for those counted by the experimenter, and between 530 and 7670 for those counted by NFFMP counters. The numbers of flying-foxes counted roosting in these colonies from drone-acquired thermal orthomosaics ranged between 1115 and 12131 (Supplementary material Table S1, see Fig. S1 for an example of a thermal orthomosaic output).
Whole colony ground counts by the experimenter were positively correlated with counts from drone-acquired thermal orthomosaics (GLMM: F1,17 = 50.61, P < 0.001, marginal R2 = 0.60, n = 25; Fig. 2a). Counts from drone-acquired thermal orthomosaics were higher than ground counts conducted by the experimenter in 22 of 25 comparative counts for an average 2.05 times higher than ground counts conducted by the experimenter. The random effect, roost ID, explained significant variation in counts from drone-acquired thermal orthomosaics (P = 0.009). The CV (precision) of the experimenter’s ground counts as assessed against counts from drone-acquired thermal orthomosaics was 26.3%.

|
Whole colony ground counts by the NFFMP counters were also positively correlated with counts from drone-acquired thermal orthomosaics (GLMM: F1,9 = 14.51, P = 0.002, marginal R2 = 0.50, n = 15; Fig. 2b). Counts from drone-acquired thermal orthomosaics were higher than NFFMP ground counts in 13 of 16 comparative counts for an average 1.92 times higher than NFFMP counts overall (excluding the 24 February 2020 Emu Plains outlier). The random effect, roost ID, did not explain significant variation in counts from drone-acquired thermal orthomosaics (P = 0.895). The CV of NFFMP ground counts as assessed against counts from drone-acquired thermal orthomosaics was 55.1%. When we included the outlying NFFMP count of the Emu Plains colony on the 24 February 2020 (see Methods), NFFMP ground counts were no longer significantly correlated with counts from drone-acquired thermal orthomosaics (F1,9 = 3.20, P = 0.073 marginal R2 = 0.18, n = 16; Supplementary material Fig. S2) and the CV of NFFMP ground counts was 78.5%.
Whole colony ground counts by the experimenter and by NFFMP counters were positively correlated (GLMM: F1,9 = 9.46, P = 0.004, marginal R2 = 0.39, n = 16; Supplementary material Fig. S3). The random effect, roost ID, did not significantly explain variation in counts from drone-acquired thermal orthomosaics (P = 0.999).
Discussion
In this study we compared accurate and precise flying-fox colony size estimates derived from drone-acquired thermal imagery to results from ground-based counts by human observers to assess the accuracy and precision of routinely performed ground-based counts. We found that drone-acquired estimates of colony size were approximately twice as large as those from ground-based counts, indicating that ground-based counting methods tend to severely underestimate true population sizes. In addition, ground-based estimates had limited precision, and more so when they involved multiple human counters, implying that with ground counting methods, population trends can only be established with significance after protracted periods of monitoring. Our findings are important as the sound management and conservation of the world’s threatened pteropodid species are critically dependent on rapid and reliable information about trends in populations.
Daytime ground counts are currently recommended as one of the preferred methods for monitoring and quantifying flying-fox colony size in Australia (Westcott et al. 2015) and globally (Worthington et al. 2001; Cousins and Compton 2005; Rahaingodrahety et al. 2008; Chaiyes et al. 2017; Manandhar et al. 2018; Sharma and Rai 2020). However, the accuracy of ground counts was hitherto unknown (Westcott et al. 2015) because no methods were available for determining the true abundances of flying-foxes in their roosts to assess the accuracy of the counts against. Recently, McCarthy et al. (2021) showed that the number of flying-foxes observed in drone-acquired thermal imagery provides highly accurate quantification of the true abundance of flying-foxes in single roost trees, and by extension, roost sites as a whole. In our present study, we found that while ground counts by both the experimenter and NFFMP counters were positively related to drone-based counts, when assessed against drone-based counts, neither gave accurate estimates of the number of individuals present (Fig. 2a, b). We found that colony size estimates from thermal orthomosaics averaged approximately two times higher than ground counts conducted by the experimenter and NFFMP counters. Considering colony counts derived from drone-acquired thermal orthomosaics as ‘true’ counts, this indicates that population estimates based on ground counts at roosts underestimate the true size of the species population by substantial margins.
The most recently available population estimate of the grey-headed flying-fox in Australia is 467 000 individuals (November 2019), estimated mainly through ground counts (Commonwealth Scientific and Industrial Research Organisation (CSIRO) 2019). In contrast, when using the parameter estimates obtained from our models (Fig. 2a, b), we predict the abundance of the species to be between 684 812 and 1 205 524 (±95% CI; 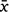 = 873 860) from experimenter counts, and between 643 685 and 2 012 248 (±95% CI;
= 873 860) from experimenter counts, and between 643 685 and 2 012 248 (±95% CI; 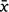 = 972 321) from NFFMP counts (see Supplementary material Item S1). Importantly, the grey-headed flying-fox is listed nationally (Australian Government Department of Agriculture, Water and the Environment 2020) and internationally (Eby et al. 2021) as ‘vulnerable to extinction’ because of ‘continuing population decline’. While our study indicates that ground-based counting methods underestimate the true size of the grey-headed flying-fox population, colony counts derived from drone-acquired thermal imagery are still positively correlated with ground counts, meaning the ground counting method is still able to capture population trends, Therefore, any conclusions about the species’ continuing population decline remain unaffected. Thus, care should be taken in interpreting our findings in the context of the conservation status of flying-foxes.
= 972 321) from NFFMP counts (see Supplementary material Item S1). Importantly, the grey-headed flying-fox is listed nationally (Australian Government Department of Agriculture, Water and the Environment 2020) and internationally (Eby et al. 2021) as ‘vulnerable to extinction’ because of ‘continuing population decline’. While our study indicates that ground-based counting methods underestimate the true size of the grey-headed flying-fox population, colony counts derived from drone-acquired thermal imagery are still positively correlated with ground counts, meaning the ground counting method is still able to capture population trends, Therefore, any conclusions about the species’ continuing population decline remain unaffected. Thus, care should be taken in interpreting our findings in the context of the conservation status of flying-foxes.
Lack of precision in animal counting due to between-observer variability and other factors is well documented (Harris and Lloyd 1977; Erwin 1982; Short and Bayliss 1985; Forsyth et al. 2006; Hanger et al. 2017), but is problematic because it hampers the ability for conservation management to quickly detect and respond to population declines (Elphick 2008). In this study, precision (CV) of ground counts as assessed against the counts from drone-acquired thermal orthomosaics was 26.3% for single-observer ground counts and 55.1% (78.5% with outlier) for multiple-observer NFFMP counts. Due to limited precision of the NFFMP ground count methodology, the time taken to reach 80% statistical power in detecting rates of population declines that would warrant listing under Australia’s Environment Protection and Biodiversity Conservation Act 1999 and IUCN’s Red List Criteria, has previously been modelled to be ~13.5 years (Westcott et al. 2012). However, here the authors assumed a very high baseline precision (CV) for ground counts of 10%. Clearly, the much lower precision for ground counts reported in our study would substantially extend the time by which declines could be detected with significance. Subsequent to their study, Westcott et al. (2015) reported a precision of ground counts (defined as the absolute difference between counts expressed as a percentage of the mean) of 28 ± 21% (mean ± s.d.). In contrast, the CV of counts from drone-acquired thermal orthomosaics has previously been shown to be much lower at 6.8 ± 6.0%, meaning they have higher precision (McCarthy et al. 2021), and hence the drone-based method can detect more subtle fluctuations in true colony size than ground count methods allowing conservation practitioners to detect population trends with significance over shorter timeframes, and so respond with greater immediacy to threats.
While drone-acquired thermal imagery provides highly accurate and precise estimates of flying-fox numbers at a roost, there is likely to remain a need for ground-based counts in the foreseeable future. Importantly, more research is needed to understand the sensitivity of flying-foxes to drones. While in our study flying-foxes did not visibly react to the drone, responses may be species-specific, and/or dependent on other external factors such as overall levels of human disturbance, including from hunting. Also, the accuracy of drone-based counts has yet to be assessed particularly in more tropical locations where higher temperatures and vegetation densities may adversely affect the detectability of animals in drone-derived thermal imagery (Kays et al. 2019; McCarthy et al. 2021), so the effectiveness of drone-based counts under such conditions remains unproven. In addition, drone-acquired imagery does not yet allow the identification of flying-fox species (or sex or age). Mixed species roosting occurred in 62% of the 430 flying-fox roosts monitored as part of the NFFMP (Timmiss et al. 2021), and the relative presence of different species can be dynamic (Welbergen 2005; Westcott et al. 2015). Thus, ground truthing via visual surveys remains essential to ensure that population estimates are species specific. Finally, given the extreme mobility of flying-foxes among roosts (Welbergen et al. 2020), to obtain meaningful population estimates, roosts must be surveyed within a short period (days) across the species range, requiring a fleet of drones and skilled drone pilots to be deployed at a landscape scale, which may be prohibitively expensive, especially in developing countries where most of the world’s flying-foxes occur. Alternatively, drone-acquired colony size estimates could be used to periodically calibrate ground count accuracy, as has been suggested for other species, such as turtles (Dunstan et al. 2020). Drone-acquired colony size estimates could also be used to benchmark the accuracy of automated counts obtained from weather radar data, which could then in principle generate accurate daily estimates of the numbers of flying-foxes in roosts (Meade et al. 2019). In summary, drone based-monitoring practises have their limitations and can only supplement rather than supplant current monitoring practises.
More than half of the members of the Pteropodidae family and of the Pteropus genus are listed as threatened, and reliable monitoring of their population trends is essential for facilitating sound conservation actions (International Union for Conservation of Nature and Natural Resources (IUCN) 2020). Drone-based monitoring can yield both more accurate and precise colony size estimates than ground-based counting methods, and as such, incorporating the use of drones into current monitoring practises would enable conservation practitioners to respond with greater immediacy and so more effectively to population declines. Thus, while drones currently cannot fully replace traditional monitoring practises, we anticipate that they will soon help set a new standard for evidence based conservation and management of these ecologically important threatened species.
Supplementary material
Supplementary material is available online.
Data availability
The data that support this study are available in the article and accompanying online supplementary material, as well as on Figshare at https://figshare.com/s/00084845b1e3c628f9d4.
Conflicts of interest
The authors declare no conflicts of interest.
Declaration of funding
This research was supported by a Paddy Pallin Foundation-sponsored Australasian Bat Society grant to EDM, and an ARC Discovery Grant (DP170104272) to JAW.
Author contributions
All authors conceived the study. EDM performed the drone surveys, statistical analyses, and led writing of the manuscript. All authors contributed to drafts and gave final approval for publication.
Acknowledgements
The authors thank David Westcott for his constructive feedback on an earlier draft of the manuscript, and Sebastian Pfautsch for generously loaning us the drone and thermal camera. We are also very grateful to the staff at Camellia Gardens, Bec Williams and Jaynia Sladek of Sutherland Shire Council, Andrew Jennings of Northern Beaches Council, Michael Ellison and Mitchell Clark of Campbelltown City Council, Amara Glynn of Centennial Parklands and Matthew Mo at NSW Department of Planning, Industry and Environment for assisting us with site access.
References
Alldredge, MW, Pacifici, K, Simons, TR, and Pollock, KH (2008). A novel field evaluation of the effectiveness of distance and independent observer sampling to estimate aural avian detection probabilities. Journal of Applied Ecology 45, 1349–1356.| A novel field evaluation of the effectiveness of distance and independent observer sampling to estimate aural avian detection probabilities.Crossref | GoogleScholarGoogle Scholar |
Anderson, K, and Gaston, KJ (2013). Lightweight unmanned aerial vehicles will revolutionize spatial ecology. Frontiers in Ecology and the Environment 11, 138–146.
| Lightweight unmanned aerial vehicles will revolutionize spatial ecology.Crossref | GoogleScholarGoogle Scholar |
Australian Government Department of Agriculture, Water and the Environment (2020) Species profile and threats database: Pteropus poliocephalus — grey-headed flying-fox. Available at http://www.environment.gov.au/cgi-bin/sprat/public/publicspecies.pl?taxon_id=186 [Accessed 7 October 2020]
Aziz, SA, McConkey, KR, Tanalgo, K, Sritongchuay, T, Low, M-R, Yong, JY, Mildenstein, TL, Nuevo-Diego, CE, Lim, V-C, and Racey, PA (2021). The critical importance of old world fruit bats for healthy ecosystems and economies. Frontiers in Ecology and Evolution 9, 641411.
| The critical importance of old world fruit bats for healthy ecosystems and economies.Crossref | GoogleScholarGoogle Scholar |
Beranek, CT, Roff, A, Denholm, B, Howell, LG, and Witt, RR (2021). Trialling a real-time drone detection and validation protocol for the koala (Phascolarctos cinereus). Australian Mammalogy 43, 260–264.
| Trialling a real-time drone detection and validation protocol for the koala (Phascolarctos cinereus).Crossref | GoogleScholarGoogle Scholar |
Betke, M, Hirsh, DE, Makris, NC, McCracken, GF, Procopio, M, Hristov, NI, Tang, S, Bagchi, A, Reichard, JD, Horn, JW, Crampton, S, Cleveland, CJ, and Kunz, TH (2008). Thermal imaging reveals significantly smaller Brazilian free-tailed bat colonies than previously estimated. Journal of Mammalogy 89, 18–24.
| Thermal imaging reveals significantly smaller Brazilian free-tailed bat colonies than previously estimated.Crossref | GoogleScholarGoogle Scholar |
Bureau of Meteorology (2020) Latest weather observations for the Sydney area. Available at http://www.bom.gov.au/nsw/observations/sydney.shtml?ref=hdr [Accessed 10 October 2020]
Chabot, D, and Bird, DM (2015). Wildlife research and management methods in the 21st century: where do unmanned aircraft fit in? Journal of Unmanned Vehicle Systems 3, 137–155.
| Wildlife research and management methods in the 21st century: where do unmanned aircraft fit in?Crossref | GoogleScholarGoogle Scholar |
Chaiyes, A, Duengkae, P, Wacharapluesadee, S, Pongpattananurak, N, Olival, KJ, and Hemachudha, T (2017). Assessing the distribution, roosting site characteristics, and population of Pteropus lylei in Thailand. Raffles Bulletin of Zoology 65, 670–680.
Cilulko, J, Janiszewski, P, Bogdaszewski, M, and Szczygielska, E (2013). Infrared thermal imaging in studies of wild animals. European Journal of Wildlife Research 59, 17–23.
| Infrared thermal imaging in studies of wild animals.Crossref | GoogleScholarGoogle Scholar |
Commonwealth of Australia Department of Environment and Energy (2019a) Flying-foxes and national environment law. Available at http://www.environment.gov.au/biodiversity/threatened/species/flying-fox-law [Accessed 23 June 2019]
Commonwealth of Australia Department of Environment and Energy (2019b) National Flying-Fox Monitoring viewer. Available at http://www.environment.gov.au/webgis-framework/apps/ffc-wide/ffc-wide.jsf [Accessed 23 June 2019]
Commonwealth Scientific and Industrial Research Organisation (CSIRO) (2019) The National Flying-fox Monitoring Program – report on the November 2019 survey. Available at https://www.environment.gov.au/system/files/pages/391f5fed-e287-4dd3-85ac-640037926ef5/files/flying-fox-nov2019-count-report.pdf [Accessed 5 June 2020]
Cousins, JA, and Compton, SG (2005). The Tongan flying fox Pteropus tonganus: status, public attitudes and conservation in the Cook Islands. Oryx 39, 196–203.
| The Tongan flying fox Pteropus tonganus: status, public attitudes and conservation in the Cook Islands.Crossref | GoogleScholarGoogle Scholar |
Croon, GW, McCullough, DR, Olson, CE, and Queal, LM (1968). Infrared scanning techniques for big game censusing. The Journal of Wildlife Management 32, 751–759.
| Infrared scanning techniques for big game censusing.Crossref | GoogleScholarGoogle Scholar |
Dey, S, Roy, US, and Chattopadhyay, S (2015). Effect of heat wave on the Indian flying fox Pteropus giganteus (Mammalia: Chiroptera: Pteropodidae) population from Purulia District of West Bengal, India. Journal of Threatened Taxa 7, 7029–7033.
| Effect of heat wave on the Indian flying fox Pteropus giganteus (Mammalia: Chiroptera: Pteropodidae) population from Purulia District of West Bengal, India.Crossref | GoogleScholarGoogle Scholar |
Dunn, WC, Donnelly, JP, and Krausmann, WJ (2002). Using thermal infrared sensing to count elk in the southwestern United States. Wildlife Society Bulletin 30, 963–967.
Dunstan, A, Robertson, K, Fitzpatrick, R, Pickford, J, and Meager, J (2020). Use of unmanned aerial vehicles (UAVs) for mark-resight nesting population estimation of adult female green sea turtles at Raine Island. PLoS ONE 15, e0228524.
| Use of unmanned aerial vehicles (UAVs) for mark-resight nesting population estimation of adult female green sea turtles at Raine Island.Crossref | GoogleScholarGoogle Scholar |
Eby, P, Richards, G, Collins, L, and Parry-Jones, K (1999). The distribution, abundance and vulnerability to population reduction of a nomadic nectarivore, the grey-headed flying-fox Pteropus poliocephalus in New South Wales, during a period of resource concentration. Australian Zoologist 31, 240–253.
| The distribution, abundance and vulnerability to population reduction of a nomadic nectarivore, the grey-headed flying-fox Pteropus poliocephalus in New South Wales, during a period of resource concentration.Crossref | GoogleScholarGoogle Scholar |
Eby, P, Roberts, B, Pennay, M, and Welbergen, JA (2021). Pteropus poliocephalus. The IUCN Red List of Threatened Species 2021, e.T18751A22085511.
| Pteropus poliocephalus.Crossref | GoogleScholarGoogle Scholar |
Elphick, CS (2008). How you count counts: the importance of methods research in applied ecology. Journal of Applied Ecology 45, 1313–1320.
| How you count counts: the importance of methods research in applied ecology.Crossref | GoogleScholarGoogle Scholar |
Erwin, RM (1982). Observer variability in estimating numbers: an experiment. Journal of Field Ornithology 53, 159–167.
Forsyth, DM, Scroggie, MP, and McDonald-Madden, E (2006). Accuracy and precision of grey-headed flying-fox (Pteropus poliocephalus) flyout counts. Wildlife Research 33, 57–65.
| Accuracy and precision of grey-headed flying-fox (Pteropus poliocephalus) flyout counts.Crossref | GoogleScholarGoogle Scholar |
Frederick, PC, Hylton, B, Heath, JA, and Ruane, M (2003). Accuracy and variation in estimates of large numbers of birds by individual observers using an aerial survey simulator. Journal of Field Ornithology 74, 281–287.
| Accuracy and variation in estimates of large numbers of birds by individual observers using an aerial survey simulator.Crossref | GoogleScholarGoogle Scholar |
Fretwell, PT, LaRue, MA, Morin, P, Kooyman, GL, Wienecke, B, Ratcliffe, N, Fox, AJ, Fleming, AH, Porter, C, and Trathan, PN (2012). An emperor penguin population estimate: the first global, synoptic survey of a species from space. PLoS ONE 7, e33751.
| An emperor penguin population estimate: the first global, synoptic survey of a species from space.Crossref | GoogleScholarGoogle Scholar |
Fujita, MS, and Tuttle, MD (1991). Flying foxes (Chiroptera: Pteropodidae): threatened animals of key ecological and economic importance. Conservation Biology 5, 455–463.
| Flying foxes (Chiroptera: Pteropodidae): threatened animals of key ecological and economic importance.Crossref | GoogleScholarGoogle Scholar |
Gelman, A (2008). Scaling regression inputs by dividing by two standard deviations. Statistics in Medicine 27, 2865–2873.
| Scaling regression inputs by dividing by two standard deviations.Crossref | GoogleScholarGoogle Scholar |
Gelman A, Su Y, Yajima M, Hill J, Pittau M, Kerman J, Zheng T (2021) arm: data analysis using regression and multilevel/hierarchical models (version 1.2-9.01). Available at https://CRAN.R-project.org/package=arm [Accessed 10 October 2021]
Geoscience Australia (2001) Geoscience Australia GEODATA TOPO series – 1:1 million to 1:10 million scale. bioregional assessment source dataset. Available at http://data.bioregionalassessments.gov.au/dataset/310c5d07-5a56-4cf7-a5c8-63bdb001cd1a [Accessed 23 April 2020]
Goldsmith B (1991) ‘Monitoring for conservation and ecology.’ (Chapman & Hall: London, UK)
Hall LS, Richards G (2000) ‘Flying foxes: fruit and blossom bats of Australia.’ (UNSW Press: Sydney, Australia)
Hanger J, de Villiers D, Forbes N, Nottidge B, Beyer H, Loader J, Timms P (2017) Moreton Bay rail koala management program: final technical report for Queensland Department of Transport and Main Roads. Endeavour Veterinary Ecology, Toorbul, Australia.
Harris, MP, and Lloyd, CS (1977). Variations in counts of seabirds from photographs. British Birds 70, 200–205.
Hodgson, JC, Mott, R, Baylis, SM, Pham, TT, Wotherspoon, S, Kilpatrick, AD, Raja Segaran, R, Reid, I, Terauds, A, and Koh, LP (2018). Drones count wildlife more accurately and precisely than humans. Methods in Ecology and Evolution 9, 1160–1167.
| Drones count wildlife more accurately and precisely than humans.Crossref | GoogleScholarGoogle Scholar |
International Union for Conservation of Nature and Natural Resources (IUCN) (2020) The IUCN red list of threatened species. Available at http://www.iucnredlist.org/ [Accessed 22 May 2020]
Kays, R, Sheppard, J, Mclean, K, Welch, C, Paunescu, C, Wang, V, Kravit, G, and Crofoot, M (2019). Hot monkey, cold reality: surveying rainforest canopy mammals using drone-mounted thermal infrared sensors. International Journal of Remote Sensing 40, 407–419.
| Hot monkey, cold reality: surveying rainforest canopy mammals using drone-mounted thermal infrared sensors.Crossref | GoogleScholarGoogle Scholar |
Klose, SM, Welbergen, JA, Goldizen, AW, and Kalko, EKV (2009a). Spatio-temporal vigilance architecture of an Australian flying-fox colony. Behavioral Ecology and Sociobiology 63, 371–380.
| Spatio-temporal vigilance architecture of an Australian flying-fox colony.Crossref | GoogleScholarGoogle Scholar |
Klose, SM, Welbergen, JA, and Kalko, EKV (2009b). Testosterone is associated with harem maintenance ability in free-ranging grey-headed flying-foxes, Pteropus poliocephalus. Biology Letters 5, 758–761.
| Testosterone is associated with harem maintenance ability in free-ranging grey-headed flying-foxes, Pteropus poliocephalus.Crossref | GoogleScholarGoogle Scholar |
LLC Agisoft (2019) ‘Agisoft metashape user manual: professional edition, version 1.5.’ (LLC Agisoft: St. Petersburg, Russia)
Lüdecke D (2021) sjstats: statistical functions for regression models (version 0.18.1). Available at https://CRAN.R-project.org/package=sjstats [Accessed 25 March 2022]
Manandhar, S, Thapa, S, Shrestha, TK, Jyakhwo, R, Wright, W, and Aryal, A (2018). Population status and diurnal behaviour of the Indian flying fox Pteropus giganteus (Brünnich, 1782) in Kathmandu Valley, Nepal. Proceedings of the Zoological Society 71, 363–375.
| Population status and diurnal behaviour of the Indian flying fox Pteropus giganteus (Brünnich, 1782) in Kathmandu Valley, Nepal.Crossref | GoogleScholarGoogle Scholar |
Mao F (2019) How one heatwave killed ‘a third’ of a bat species in Australia. BBC News. Available at https://www.bbc.com/news/world-australia-46859000 [Accessed 19 May 2019]
Marsh, DM, and Trenham, PC (2008). Current trends in plant and animal population monitoring. Conservation Biology 22, 647–655.
| Current trends in plant and animal population monitoring.Crossref | GoogleScholarGoogle Scholar |
Martin, TG, Nally, S, Burbidge, AA, Arnall, S, Garnett, ST, Hayward, MW, Lumsden, LF, Menkhorst, P, McDonald-Madden, E, and Possingham, HP (2012). Acting fast helps avoid extinction. Conservation Letters 5, 274–280.
| Acting fast helps avoid extinction.Crossref | GoogleScholarGoogle Scholar |
McCarthy, ED, Martin, JM, Boer, MM, and Welbergen, JA (2021). Drone-based thermal remote sensing provides an effective new tool for monitoring the abundance of roosting fruit bats. Remote Sensing in Ecology and Conservation 7, .
| Drone-based thermal remote sensing provides an effective new tool for monitoring the abundance of roosting fruit bats.Crossref | GoogleScholarGoogle Scholar |
McKelvey, KS, Aubry, KB, and Schwartz, MK (2008). Using anecdotal occurrence data for rare or elusive species: the illusion of reality and a call for evidentiary standards. Bioscience 58, 549–555.
| Using anecdotal occurrence data for rare or elusive species: the illusion of reality and a call for evidentiary standards.Crossref | GoogleScholarGoogle Scholar |
Meade, J, van der Ree, R, Stepanian, PM, Westcott, DA, and Welbergen, JA (2019). Using weather radar to monitor the number, timing and directions of flying-foxes emerging from their roosts. Scientific Reports 9, 10222.
| Using weather radar to monitor the number, timing and directions of flying-foxes emerging from their roosts.Crossref | GoogleScholarGoogle Scholar |
Meade, J, Martin, JM, and Welbergen, JA (2021). Fast food in the city? Nomadic flying-foxes commute less and hang around for longer in urban areas. Behavioral Ecology 32, 1151–1162.
| Fast food in the city? Nomadic flying-foxes commute less and hang around for longer in urban areas.Crossref | GoogleScholarGoogle Scholar |
Mildenstein T, Tanshi I, Racey PA (2016) Exploitation of bats for bushmeat and medicine. In ‘Bats in the anthropocene: conservation of bats in a changing world’. (Eds CC Voigt, T Kingston) pp. 325–375. (Springer International Publishing: New York, NY, USA)
Mo, M, Roache, M, Haering, R, and Kwok, A (2021). Using wildlife carer records to identify patterns in flying-fox rescues: a case study in New South Wales, Australia. Pacific Conservation Biology 27, 61–69.
| Using wildlife carer records to identify patterns in flying-fox rescues: a case study in New South Wales, Australia.Crossref | GoogleScholarGoogle Scholar |
Mohd-Azlan, J, Zubaid, A, and Kunz, TH (2001). Distribution, relative abundance, and conservation status of the large flying fox, Pteropus vampyrus, in peninsular Malaysia: a preliminary assessment. Acta Chiropterologica 3, 149–162.
Otto, MC, and Pollock, KH (1990). Size bias in line transect sampling: a field test. Biometrics 46, 239–245.
| Size bias in line transect sampling: a field test.Crossref | GoogleScholarGoogle Scholar |
Pinheiro J, Bates D, DebRoy S, Sarkar D, Heisterkamp S, Van Willigen B, Maintainer R (2022) nlme: Linear and Nonlinear Mixed Effects Models (version 3.1-158). Available at https://CRAN.R-project.org/package=nlme [Accessed 10 October 2021]
R Core Team (2018) R: A Language and Environment For Statistical Computing (R foundation for Statistical Computing: Austria, Vienna) Available at https://www.R-project.org/ [Accessed 25 March 2022]
Rahaingodrahety, VN, Andriafidison, D, Ratsimbazafy, JH, Racey, PA, and Jenkins, RK (2008). Three flying fox (Pteropodidae: Pteropus rufus) roosts, three conservation challenges in southeastern Madagascar. Madagascar Conservation & Development 3, 17–21.
Reddy, S, and Dávalos, LM (2003). Geographical sampling bias and its implications for conservation priorities in Africa. Journal of Biogeography 30, 1719–1727.
| Geographical sampling bias and its implications for conservation priorities in Africa.Crossref | GoogleScholarGoogle Scholar |
Roberts, B, Eby, P, and Westcott, D (2020). Pteropus conspicillatus. The IUCN Red List of Threatened Species 2020, e.T18721A22080456.
| Pteropus conspicillatus.Crossref | GoogleScholarGoogle Scholar |
Schneider, CA, Rasband, WS, and Eliceiri, KW (2012). NIH Image to ImageJ: 25 years of image analysis. Nature Methods 9, 671–675.
| NIH Image to ImageJ: 25 years of image analysis.Crossref | GoogleScholarGoogle Scholar |
Seymour, AC, Dale, J, Hammill, M, Halpin, PN, and Johnston, DW (2017). Automated detection and enumeration of marine wildlife using unmanned aircraft systems (UAS) and thermal imagery. Scientific Reports 7, 45127.
| Automated detection and enumeration of marine wildlife using unmanned aircraft systems (UAS) and thermal imagery.Crossref | GoogleScholarGoogle Scholar |
Sharma, P, and Rai, M (2020). Population status of indian flying fox, Pteropus giganteus in Urban Guwahati, Assam, India: a case study. Applied Ecology and Environmental Sciences 8, 287–293.
| Population status of indian flying fox, Pteropus giganteus in Urban Guwahati, Assam, India: a case study.Crossref | GoogleScholarGoogle Scholar |
Short, J, and Bayliss, P (1985). Bias in aerial survey estimates of kangaroo density. Journal of Applied Ecology 22, 415–422.
| Bias in aerial survey estimates of kangaroo density.Crossref | GoogleScholarGoogle Scholar |
Simmons NB (2020) Bats of the world: a taxonomic and geographic database. Available at https://batnames.org/ [Accessed 26 January 2022]
Sutherland, WJ (2006). Predicting the ecological consequences of environmental change: a review of the methods. Journal of Applied Ecology 43, 599–616.
| Predicting the ecological consequences of environmental change: a review of the methods.Crossref | GoogleScholarGoogle Scholar |
Tait, J, Perotto-Baldivieso, HL, McKeown, A, and Westcott, DA (2014). Are flying-foxes coming to town? Urbanisation of the spectacled flying-fox (Pteropus conspicillatus) in Australia. PLoS ONE 9, e109810.
| Are flying-foxes coming to town? Urbanisation of the spectacled flying-fox (Pteropus conspicillatus) in Australia.Crossref | GoogleScholarGoogle Scholar |
Timmiss, LA, Martin, JM, Murray, NJ, Welbergen, JA, Westcott, D, McKeown, A, and Kingsford, RT (2021). Threatened but not conserved: flying-fox roosting and foraging habitat in Australia. Australian Journal of Zoology 68, 226–233.
| Threatened but not conserved: flying-fox roosting and foraging habitat in Australia.Crossref | GoogleScholarGoogle Scholar |
Vinson, SG, Johnson, AP, and Mikac, KM (2020). Thermal cameras as a survey method for Australian arboreal mammals: a focus on the greater glider. Australian Mammalogy 42, 367–374.
| Thermal cameras as a survey method for Australian arboreal mammals: a focus on the greater glider.Crossref | GoogleScholarGoogle Scholar |
Wagner, JL (1981). Visibility and bias in avian foraging data. The Condor 83, 263–264.
| Visibility and bias in avian foraging data.Crossref | GoogleScholarGoogle Scholar |
Welbergen JA (2005) The social organisation of the grey-headed flying-fox, Pteropus poliocephalus. Dissertation. University of Cambridge, Cambridge, UK
Welbergen, JA, Klose, SM, Markus, N, and Eby, P (2008). Climate change and the effects of temperature extremes on Australian flying-foxes. Proceedings of the Royal Society B: Biological Sciences 275, 419–425.
| Climate change and the effects of temperature extremes on Australian flying-foxes.Crossref | GoogleScholarGoogle Scholar |
Welbergen, JA, Meade, J, Field, HE, Edson, D, McMichael, L, Shoo, LP, Praszczalek, J, Smith, C, and Martin, JM (2020). Extreme mobility of the world’s largest flying mammals creates key challenges for management and conservation. BMC Biology 18, 101.
| Extreme mobility of the world’s largest flying mammals creates key challenges for management and conservation.Crossref | GoogleScholarGoogle Scholar |
Westcott, DA, and McKeown, A (2004). Observer error in exit counts of flying-foxes (Pteropus spp.). Wildlife Research 31, 551–558.
| Observer error in exit counts of flying-foxes (Pteropus spp.).Crossref | GoogleScholarGoogle Scholar |
Westcott DA, McKeown A, Murphy HT, Fletcher CS (2011) A monitoring method for the grey-headed flying-fox, Pteropus poliocephalus. (CSIRO) Available at https://www.environment.gov.au/biodiversity/threatened/species/flying-fox-monitoring [Accessed 3 October 2020]
Westcott, DA, Fletcher, CS, McKeown, A, and Murphy, HT (2012). Assessment of monitoring power for highly mobile vertebrates. Ecological Applications 22, 374–383.
| Assessment of monitoring power for highly mobile vertebrates.Crossref | GoogleScholarGoogle Scholar |
Westcott DA, McKeown A, Parry H, Parsons J, Jurdak R, Kusy B, Caley P (2015) Implementation of the National Flying-Fox Monitoring Program. (Australian Government, Rural Industries Research and Development Corporation) Available at https://publications.csiro.au/rpr/pub?pid=csiro:EP156844 [Accessed 3 September 2020]
Wich SA, Koh LP (2018) ‘Conservation drones: mapping and monitoring biodiversity.’ (Oxford University Press: Oxford, UK)
Witt, RR, Beranek, CT, Howell, LG, Ryan, SA, Clulow, J, Jordan, NR, Denholm, B, and Roff, A (2020). Real-time drone derived thermal imagery outperforms traditional survey methods for an arboreal forest mammal. PLoS ONE 15, e0242204.
| Real-time drone derived thermal imagery outperforms traditional survey methods for an arboreal forest mammal.Crossref | GoogleScholarGoogle Scholar |
Woinarski J (2018) ‘A bat’s end: the Christmas Island pipistrelle and extinction in Australia.’ (CSIRO Publishing: Melbourne, Vic., Australia)
Worthington, DJ, Marshall, AP, Wiles, GJ, and Kessler, CC (2001). Abundance and management of Mariana fruit bats and feral ungulates on Anatahan, Mariana Islands. Pacific Conservation Biology 7, 134–142.
| Abundance and management of Mariana fruit bats and feral ungulates on Anatahan, Mariana Islands.Crossref | GoogleScholarGoogle Scholar |
Yabsley, SH, Meade, J, Martin, JM, and Welbergen, JA (2021). Human-modified landscapes provide key foraging areas for a threatened flying mammal: the grey-headed flying-fox. PLoS ONE 16, e0259395.
| Human-modified landscapes provide key foraging areas for a threatened flying mammal: the grey-headed flying-fox.Crossref | GoogleScholarGoogle Scholar |