

Accuracies and biases of ageing white-tailed deer in semiarid environments
Aaron M. Foley A C , John S. Lewis A , Oscar Cortez B , Mickey W. Hellickson B , David G. Hewitt A , Randy W. DeYoung A , Charles A. DeYoung A and Matthew J. Schnupp B
A C , John S. Lewis A , Oscar Cortez B , Mickey W. Hellickson B , David G. Hewitt A , Randy W. DeYoung A , Charles A. DeYoung A and Matthew J. Schnupp B
A Caesar Kleberg Wildlife Research Institute, Texas A&M University-Kingsville, 700 University Boulevard, MSC 218 Kingsville, Texas 78363, USA.
B King Ranch, Inc., 2205 Highway 141 West, Kingsville, TX 78363, USA.
C Corresponding author. Email: aaron.foley@tamuk.edu
Wildlife Research 49(3) 237-249 https://doi.org/10.1071/WR21050
Submitted: 9 March 2021 Accepted: 4 August 2021 Published: 23 December 2021
Journal Compilation © CSIRO 2022 Open Access CC BY-NC-ND
Abstract
Context: The ability to accurately estimate age of animals is important for both research and management. The two methods for age estimation in ungulates are tooth replacement and wear (TRW) and cementum annuli (CA). Errors in estimated TRW ages are commonly attributed to environmental conditions; however, the influence of environmental variables on tooth wear has not been quantified. Further, the performance of CA in environments with weak seasonality has not been thoroughly evaluated.
Aims: The study had the following three goals: identify environmental and morphological factors that influenced estimated ages, quantify accuracy of TRW and CA, and develop TRW ageing criteria that minimise error.
Methods: We used data from harvested (n = 5117) and free-ranging, known-age white-tailed deer (n = 134) collected in southern Texas, USA, to quantify environmental and morphological influences on estimated TRW ages, and assess biases in both methods.
Key results: We observed substantial variation in age estimates for both TRW and CA. Soil, drought and supplemental nutrition had minor effects on tooth wear, insufficient to alter age estimates by ≥1 year. Body mass and antler size influenced age estimates for TRW only for extreme outliers. Both methods were biased and tended to under-estimate ages of adult deer, especially TRW. Wear on the first molar was most correlated with the known age (r2 = 0.78) and allowed biologists to correctly place known-age deer into age classes of 2, 3–5, and ≥6 years old 72%, 73% and 68% of the time, an improvement compared with the 79%, 48% and 28% accuracy from pooled TRW.
Conclusions: We observed substantial inter- and intra-individual variation in tooth-wear patterns that became more pronounced in older deer. Individual variation had a greater influence on TRW ages than did environmental covariates, whereas CA ages appeared unaffected by environment. Although variable, age estimates were ±1 year of the true age 87% and 93% of the time for TRW and CA respectively.
Implications: Managers, ecologists and epidemiologists often incorporate ages into population models. The high inter-individual variation in estimated ages, the tendency to underestimate ages of older deer, and the ageing method need to be considered.
Keywords: ageing, cementum annuli, management, Odocoileus virginianus, Severinghaus, soils, supplemental nutrition, tooth replacement and wear, white-tailed deer.
Introduction
Reliable estimates of age are critical to the management of long-lived animals, such as ungulates. For instance, population dynamics of ungulates are often a function of age structures, given that growth, survival, reproduction, and behaviour are age-dependent (Gaillard et al. 1998). Disease prevalence or contact rates may be sex- and age-dependent (Miller and Conner 2005). Furthermore, most populations of ungulates are managed via recreational harvest, and wildlife agencies use age structures of harvested ungulates to estimate population parameters and direct future management plans (Roseberry and Woolf 1991). Management by selective harvest (Jacobson et al. 2011) has become increasingly widespread, and relies on scientifically sound criteria for different age and phenotypic classes (Demarais and Strickland 2011). Accurate ageing methods also allow managers to design harvest criteria that target different antler or horn characteristics (Solberg and Sæther 1994; Hewitt et al. 2014). If an animal cannot be placed accurately into age classes, then choosing an age at which harvest is acceptable becomes a subjective exercise.
Given the widespread reliance on age estimates of ungulates, the accuracy of ageing methods is an important consideration. The two primary methods for estimating ages of large herbivores are tooth replacement and wear (TRW; Severinghaus 1949) and cementum annuli (CA; Gilbert 1966). Estimating ages via TRW is based on replacement of the juvenile dentition for young animals and the amount of wear on the permanent dentition for older animals that accumulates from mastication of plant matter. Ageing via CA requires extraction of a tooth root, usually an incisor, which is sectioned, stained and viewed under a microscope where the annuli are counted. Development of annuli typically occurs during periods of slow somatic growth (e.g. winter); thus, each annulus represents 1 year of age.
Estimating ages of white-tailed deer (Odocoileus virginianus) has received the most attention in both scientific and popular media. Scientists have described biases, limitations and recommendations for use of TRW (Erickson et al. 1970; Jacobson and Reiner 1989; Hamlin et al. 2000; Gee et al. 2002; Meares et al. 2006). Overall, most studies on the accuracy of TRW have been hampered by small sample sizes (Cook and Hart 1979; Hamlin et al. 2000) or lack of known-age animals (Gilbert and Stolt 1970; Storm et al. 2014). A synthesis of the literature suggests the TRW method is inversely biased with age; there is a broad tendency to overestimate the age of younger deer and underestimate the age of older deer (DeYoung 1989; Hamlin et al. 2000). Finally, the TRW method is susceptible to bias based on deer morphometrics. For instance, body mass or antler size may subconsciously influence an observer’s estimated age (e.g. Gee et al. 2014).
Accuracy of TRW ages is affected by variation among individual deer and regional populations in patterns of tooth wear. These observations have prompted many untested hypotheses about variation in tooth-wear patterns, and their correlation with environmental covariates (McCullough 1996; Hewison et al. 1999; Hall et al. 2012; Storm et al. 2014). For instance, sandy soils may accelerate tooth wear if grit from soil or wind-blown dust is incidentally consumed while foraging (Kaiser et al. 2013; Schulz et al. 2013). Provision of supplemental nutrition in the form of pelleted rations (Kozicky 1997) is a common management technique in some areas (Jacobson et al. 2011). Access to pelleted feed or other supplemental nutrition may reduce tooth wear compared with a forage diet because pellets require less mastication than do natural forages (Severinghaus and Cheatum 1956; Kaiser et al. 2009). In semiarid environments, frequent droughts have a strong effect on the diet of white-tailed deer (Folks et al. 2014; DeYoung et al. 2019). Specifically, during drought years, proportion of shrubs increased in deer diets, whereas forbs declined. Thus, fibrous diets available during dry years may hasten tooth wear compared with wet years, when abundant non-fibrous vegetation is available (Kaiser et al. 2013).
Accuracy of CA for white-tailed deer can be 70–100% (Roseberry 1980; McCaffery 2001), often greater than TRW ages (85% CA vs 43% TRW; Hamlin et al. 2000). Annuli counts may vary regionally because seasonality becomes less pronounced as latitude decreases. Thus, accuracy of CA may vary geographically. In areas with mild winters, faint annuli may result in underestimated ages (McCullough 1996). In areas where multiple nutritional stress periods occur, such as summer droughts and harsh winters, multiple annuli may develop (Jacobson and Reiner 1989). As with TRW, factors that influence the accuracy of CA have been difficult to quantify because of small sample sizes (Cook and Hart 1979; DeYoung 1989) or lack of known-aged individuals (Asmus and Weckerly 2011).
Although many have studied the TRW and CA ageing techniques, the role of individual and environmental factors in ageing accuracy remains obscure; many hypotheses about the causal factors of variation in ageing accuracy are untested. The older age classes (≥2 years old) have the most variation in tooth-wear patterns; one proposed solution would be to place deer in three age classes, namely, fawn, yearling and ≥2 years old (Gee et al. 2002). Unfortunately, this approach has limitations for animals that may live for ≥10 years. The ability to separate adults into prime-age and post-prime age classes is critical for insight into population demographics (DelGiudice et al. 2007). We used large-scale and long-term datasets to quantify factors that affect accuracy of ageing by TRW and CA in semiarid environments. Our specific objectives were to (1) determine the influence of environmental and morphometric variables on tooth wear as indexed by the difference between TRW and CA ages of harvested deer, (2) assemble a large sample of known-age, free-ranging deer to understand biases of the TRW and CA techniques, and (3) evaluate a modification of the TRW criteria on the basis of information from Objectives 1 and 2 to improve the accuracy of age estimates for adult deer.
Materials and methods
Environmental and visual variables
We evaluated effects of environmental and visual variables on TRW ages (Objective 1) by using a large sample of deer harvested on the King Ranch, located in Brooks, Jim Wells, Kenedy, Kleberg, Nueces and Willacy counties, Texas, USA. Annual temperature and rainfall averaged 27.7°C and 44.7 cm respectively, during 1971–2000 (Parent et al. 2016); rainfall was variable (>20% coefficient of variation, Norwine and John 2007). The King Ranch is divided into four non-contiguous divisions: Laureles (796.3 km2), Norias (971.0 km2), Encino (423.6 km2) and Santa Gertrudis (816.6 km2, Fig. 1). The Laureles Division consists of dense, dark-clay fertile soils, with sandy soils in coastal pastures. The Norias Division consists of sand to sandy loam soils and is dominated by live oak (Quercus virginiana) in its central portions with grasslands and rolling sand dunes near the coastline. The Encino Division consists of sandy to sandy loam soils with mixed brush species such as mesquite–granjeño thornscrub (Prosopis glandulosa–Celtis pallida). The Santa Gertrudis Division consists of clay, sandy to sandy loam soils, with mixed brush containing mesquite, huisache (Acacia farnesiana), brasil (Condalia hookeri) and granjeño. Within the four divisions, there are 125 wildlife management units (WMU) ranging from 30 to 93 km2 in size. Each WMU had specific wildlife management programs; ~80% of the WMUs were leased for recreational hunting. On King Ranch, many hunting leases choose to implement supplemental nutrition programs because of the positive influence of nutrition on individual and population attributes in the semiarid climate (DeYoung et al. 2019); density of feed stations in some WMUs were adjusted as deer management program objectives were modified.

|
For Objective 1, we evaluated the influence of soil composition, supplemental nutrition, harvest date, antler size and eviscerated body mass on TRW age estimates by using data from male white-tailed deer harvested on King Ranch during 2000–2015. The King Ranch hunting program involved both leases for hunting access and guided hunting opportunities led by ranch personnel. Each leased WMU was required to employ a biologist approved by King Ranch to ensure that biological data were collected properly. For every male deer harvested on King Ranch during 2000–2015, biologists recorded location and date of harvest, age by TRW, eviscerated body mass, and antler measurements according to the Boone and Crockett scoring system (Nesbitt and Wright 1981). An I1 incisor (Lyons et al. 2012) was extracted from each male with the aid of a dental elevator and forceps, keeping the tooth root intact. Incisors were stored in paper envelopes and allowed to air-dry, then sent to a commercial laboratory for CA analysis (Matson’s Laboratory, Manhattan, MT, USA).
To evaluate the hypothesis that soil composition influences tooth wear, we identified soil characteristics on the study sites. We overlaid WMU boundaries over the Texas Natural Resources Information System soil map (TNRIS 2019) with ArcGIS (ESRI, Redlands, CA, USA). We obtained the % sand composition for each soil class within each WMU, then calculated weighted % sand composition on the basis of area of each soil class within each WMU (Fig. S1, available as Supplementary material to this paper).
To evaluate the hypothesis that enhanced nutrition from pelleted feed influences TRW ages, we collected information on density of feed stations annually during 2000–2015 from each WMU wildlife biologist. At each feed station, there were 1–4 independent feeders; most were timed to release pellets twice daily. The number of feed stations within some WMU were unknown for some years; harvest records associated with these sites were excluded from analyses.
Finally, visual cues may unintentionally cause bias of estimated TRW ages. For instance, antler size increases with age (Hewitt et al. 2014), where most males reach full antler potential at 5–7 years of age. However, antler size also varies within an age class. Therefore, males with antlers much larger or smaller than average for a given age might be more likely to be over- or under-aged respectively. Similarly, body mass correlates with age as males achieve skeletal maturity; thus, one may over-age a young male with an unusually high body mass and vice versa. Further, male white-tailed deer may lose ≥20–30% of body mass during the breeding season (Foley et al. 2018). Therefore, if body mass is important as a visual cue, estimated TRW ages may vary temporally, because deer hunting seasons generally coincide with the breeding season.
Our response variable was the TRW age minus CA age for each individual. Matson’s laboratory specifies a score for each assigned CA age on the basis of their ability to distinguish annuli, where an ‘A’ score indicates a high confidence level of age assignment. We used only records with an ‘A’ score in our model. We excluded fawns and 1-year-old deer aged via TRW from the environmental analysis because these age classes can be determined unambiguously by tooth replacement and few males in these age classes were harvested. We also excluded males aged by TRW as 8 years old from our dataset because King Ranch biologists placed all deer ≥8 years old in a single age class. This is because tooth wear becomes so pronounced that it becomes difficult to apply the Severinghaus (1949) technique for deer >8 years old.
We tested hypotheses about environmental variables by using a linear model with antler size (gross Boone and Crockett score in centimetres), intensity of enhanced nutrition (feed stations km−2), eviscerated body mass (kg), sand composition of soils (%) and harvest date (day) as explanatory variables. Preliminary analyses showed heteroscedasticity issues when age classes were pooled because young males had relatively smaller morphometric measures than did older males, so we performed separate analyses for each TRW age class (2–7 years old). We standardised the explanatory variables before running each age-specific model.
Given the large sample size of harvested deer and effects of sample size on parametric tests (Johnson 1999), we anticipated that small effect sizes may be statistically significant but not biologically important (Nakagawa and Cuthill 2007). Most ungulates are long-lived and thus adults are aged at 1-year increments (such as 1, 2, 3 years, and so on). Therefore, the effect of a given covariate must be large enough to influence estimated ages by ≥1 year to be biologically meaningful. Further, the effect needed to influence estimated age by ≥1 year must be within the range of observed values to be plausible. For instance, if sand (%) was a statistically significant predictor but the effect needed to change estimated TRW age by a full year was >100% sand, then the sand effect on tooth wear is probably not biologically meaningful. For each age-specific model, we calculated the effect of each covariate that would be needed to change the value of the intercept by 1 year, which we defined as a minimum biologically meaningful increment for the categorical variable age class. We compared statistically significant age-specific predicted effects with age-specific empirical ranges of observed values to determine whether the environmental or visual effects were large enough to be biologically meaningful. Because some of the empirical ranges may contain extreme values that were infrequently observed, we also compared the predicted effect with the range of 1 s.d. (68% of the population) to determine whether the predicted effect would influence estimated ages of ‘representative’ deer. We also calculated % agreement between TRW and CA for each age class.
Our use of TRW age minus CA age for the response variable assumes that only TRW ages are affected by environmental variables or visual cues. Previous studies of age estimation in white-tailed deer have found that CA ages are variable but less biased than are TRW; the use of CA ages produced more realistic population age distributions than did TRW ages (DeYoung 1989). For CA ages, although males were not of a known age, laboratory technicians viewed only the extracted incisor and would not be biased by the deer’s physical appearance or by the wear patterns of molars. If tooth wear differs according to environmental variables or observers tend to give biased age estimates, the distribution of TRW ages should depart from the distribution of CA ages for a given age class or physical variable. As long as the CA ages are unaffected by environment, we should be able to detect meaningful departures in the distribution of TRW ages unless CA ages are highly variable. The discrepancy in TRW age distributions should be more pronounced for older deer, because they have had a longer exposure to environmental variables.
Our long-term dataset encompassed two distinct climatic patterns, which gave us the opportunity to evaluate whether the climate affects deer diet, which in turn affected tooth wear or CA. During 2000–2005, the southern–central Texas climatic region was free of exceptional droughts (≤ –5.0 Palmer drought severity index), whereas during 2006–2013, the region experienced exceptional drought conditions during February 2006 – January 2007, June 2008 – November 2009, and March 2011 – July 2013 (United States Drought Monitor 2019). To investigate whether drought conditions influenced tooth-wear patterns, we used Welch’s t-test, because of unequal variances (Welch 1951), to determine whether the means in age differences between TRW and CA for males aged as 5 years old via TRW were different between the wetter period (2005) and the drought period (2012). We limited the sample to males aged 5 years old because we wanted to focus on males that were alive during the entirety of the drought or wet years, but not both. For the two periods, we also compared the frequency of incisors from males estimated to be 5 years old via TRW that were scored ‘A’ by Matson’s Laboratory to test whether prevalence of distinct annuli differed between the wetter period and the drought period. All statistical analyses were performed in R (R Core Team 2013).
Accuracy evaluation
We captured and marked deer from six sites in southern Texas during 1998–2007 to obtain free-ranging, known-age deer for the evaluation of TRW and CA (Objectives 2, 3; Fig. 1). Study sites included four sites in Webb County, one in Brooks County, and one in Kleberg County, Texas, USA. The Webb County sites (numbered Webb 1–4, hereafter) ranged in size from 49.4 to 101.27 km2 of mesquite-dominated shrubland located north-east and east of Laredo, Texas. The Brooks site (144 km2) was within the Encino Division of the King Ranch and consisted of mesquite savannas and mixed-brush rangeland. Deer in Kleberg County were captured on a 77.3 km2 site on the Laureles Division of King Ranch, 25 km east of Kingsville, Texas, in the eastern Rio Grande Plains ecoregion. The Kleberg County site consisted of mesquite- and huisache (Acacia farnesiana)-dominated mixed-shrub rangeland. Free water was available on all six sites through ephemeral streams, livestock troughs and earthen water catchments. Supplemental nutrition was provided in the form of pelleted feed at rates of one feed station per 1.22, 1.37, 0.77 and 1.41 km2 for Webb 1–4 respectively. Site 3 in Webb County also used additional feeders that dispensed shelled corn near hunting blinds (1 feed station per 1.74 km2) during hunting season and provided whole cotton seed as an additional supplement at all feed sites. The density of pelleted feed stations was consistent throughout the study. No pelleted supplement was provided on the Brooks and Kleberg County sites, but shelled corn was available as bait during the hunting season.
Capture and handling
To establish a sample of free-ranging, known-age deer for Objectives 2 and 3, we captured deer during 1998–2008 in Webb and Kleberg Counties using a net-gun fired from a helicopter (DeYoung 1988). Male deer of ≥1 year of age were captured as encountered by the helicopter pilot, without selection for age or antler size. In some years, also fawns were captured. Ground crews physically subdued each captured deer, removed the net, and blindfolded deer to reduce stress. Age of deer was estimated using the TRW method; only fawns and yearlings (1 year old) of both sexes were retained for analysis (see Data collection section). Passive integrated transponder tags (Avid Identification Systems, Norco, California, USA) were implanted in Webb and Kleberg County deer for future identification, and numbered, colour-coded ear tags were placed on deer at the Webb County sites. All deer were released at their capture site. In Brooks County, fawns were captured by hand shortly after birth from 1982 through to 2007, marked with a unique α-numeric code using a freeze brand (Newsom and Sullivan 1968), and released at their capture site. Capture and handling procedures were approved by the Texas A&M University-Kingsville Institutional Animal Care and Use Committee (Authorisation No. 3-98-09, 99-5-2, 2003-5-14). Fawn captures and branding were approved by Texas Parks and Wildlife Department (Permit # SPR-0790-200).
Data collection
Mandibles were collected from tagged and freeze-branded deer harvested as part of normal hunting activity on each site. All mandibles from deer first captured as a fawn or yearling were considered to be of a known age because these two age classes can be unambiguously aged by replacement of juvenile teeth. Mandibles were sectioned between the canine and first premolar, and unique, numbered metal tags were wired to each section for identification. Each incisor arcade was stored in a labelled manila envelope. The premolar and molar section was cleaned by boiling, and teeth were glued to the bone to prevent loss. Following cleaning, the identification number matching the metal tag was written on the mandible in permanent ink.
TRW method
Six experienced observers, holding at least a Master of Science degree in wildlife biology, aged known-age mandibles from deer that were ≥2 years old. We provided TRW reference literature during tests to ensure that our evaluation did not confound biologists’ memory of the technique with bias in the technique. Biologists had access to Severinghaus’ (1949) manuscript and a visual ageing guide (Ramsey et al. 1993) and aged each mandible independently. Estimated ages were capped at ≥8 years old. We calculated % of mandibles that were correctly aged for each age class and % of paired mandibles (e.g. right and left mandible from an individual deer) that had identical ages by each observer for each age class. In the event that the observers assigned different ages for the paired mandibles, we determined whether accuracy would be improved by using the younger or older estimated age.
CA method
We sent incisors from known-age deer to a commercial laboratory (Matson’s Laboratory) for sectioning and ageing. Matson’s laboratory staff was not given access to the deer’s known age.
Modified TRW technique
We used the known-age mandibles to develop new criteria based on tooth wear that more accurately reflected the known ages or age intervals. Specifically, we ranked wear on each of the three molars on a scale 1–5, where 1 is little or no wear and 5 is pronounced wear. Molars with enamel width greater than dentine width on the lingual crest were ranked 1, molars with dentine width equal to enamel width were ranked 2, and molars with dentine width greater than enamel width were ranked 3. Molars ranked 4 had dentine width greater than enamel width, and the infundibulum was worn to a smooth ridge. Molars that had the infundibulum completely worn away and were of a concave appearance were ranked 5 (Table 1). We ranked each mandible separately to quantify inter-individual variation and recorded the number of mandibles with specific molar-wear combinations. Our criteria for an ideal modification to existing ageing criteria were that the methods are simple, easy to interpret, and able to categorise age classes or intervals with greater accuracy than our evaluation of the Severinghaus (1949) method. We conducted linear regressions to determine which of the three molars had wear patterns that best fit known ages. We evaluated these new criteria by using an independent set of known-age mandibles that were collected during the last 3 years of the study. Twenty Texas Parks and Wildlife Department biologists with experience using the TRW method were given visual guides with the new criteria (Fig. 2) and asked to age mandibles to three age classes: 2, 3–5 and ≥6 years old. We calculated % of mandibles that had correct age estimates. We also compared methods by contrasting the % correct classifications by using the modified criteria versus pooling TRW estimates into the same age categories. We did not evaluate whether accuracy was influenced by environmental conditions as was undertaken in the Environmental and visual variables section (see above) because some covariates were not recorded, and small sample sizes would result in over-parametrisation in modelling efforts.
|

|
Results
Environmental and visual effects
Soil composition values within WMU on King Ranch ranged from 21% to 97% sand; intensity of enhanced nutrition ranged from 0 to 1.16 feed stations km−2. Our original dataset consisted of 7134 harvest records. After removing harvest records with missing covariates, incisor samples from Matson’s laboratory lacking discrete age estimates (i.e. range 5–7 years old) or with B or C scores, and age differences of >5 years (for normal distribution, n = 14), our final dataset consisted of 5117 males. Age-specific TRW sample sizes were as follows: 2 (n = 49), 3 (n = 305), 4 (n = 761), 5 (n = 1700), 6 (n = 1383) and 7 (n = 919).
With the exception of sand, all covariates were statistically significant at α = 0.05 for ≥1 age class; antler size and body mass were often statistically significant for an age class (Table 2). Although antler size and body mass could have influenced estimated TRW age by ≥1 year, most of those effects would be limited to the few individuals with extreme outliers, given that most of the predicted effects were larger than the 1 s.d. ranges of the age-specific observed values (Table 3). On the basis of 1 s.d. ranges, the influence of antler size in 2-year-old males, our smallest sample size, is the only covariate that could have biased TRW estimates in representative males. Harvest date, another indicator of changes in body characteristics, had a significant effect on estimated ages for males 6 and 7 years old; for every ~30 days into the hunting season, estimated TRW ages were 0.18 and 0.27 days older than were estimated CA ages respectively, but would not influence the estimated age by 1 year for a representative deer (Table 3). Our models explained less variation in age differences as the estimated TRW age of males increased (Table 2).

|

|
Despite removal of deer TRW aged as ≥8 years old, there were numerous older deer within most age classes. For instance, 35% of males aged as 7 on the basis of TRW had CA ages ranging from 8 to 12 years old. Further, 32% of males aged as 6 by TRW had CA ages ranging from 7 to 16 years old. Agreement for TRW and CA ages peaked at 4 years old (Fig. S2). The TRW and CA agreement of unknown-age males was lowest for 2-year-old males (24%), ranging from 31% to 37% for 3–5-year-old males, then declining as males became older (Fig. S2). Overall, TRW and CA were ±1 year 74% of the time, but accuracy declined with age. For 2-year-old males, 83% of TRW estimates were ±1 year of CA estimates, then gradually declined to ±1 year 63% of the time at 7 years old.
Mean difference between TRW and CA ages for males aged 5 years old by TRW were not different during drought and wet years (t309.5 = –1.18, P = 0.24). During the drought years, age differences between TRW and CA averaged –0.21, versus –0.40 during wet years. All incisors were scored as ‘A’ during the wet and drought years.
Accuracy evaluation
TRW method
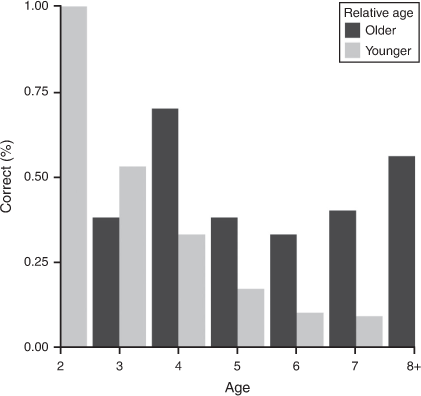
Six observers estimated ages of 264 known-age mandibles from 134 deer ≥2 years old. Estimated ages by TRW of mandibles resulted in accuracies ranging 43–51% among observers. All observers agreed on the same age for only 19% of mandibles; average agreement was 4.1 observers per mandible. Mandible-specific age estimates were more accurate for younger deer and declined with age (Table 4). In general, observers tended to underestimate the age of deer ≥4 years old, whether the deer was male or female (Fig. S3). Although mandibles from the same deer were not presented side by side during the evaluation, observers had the same ages for both mandibles 67% (511/768) of the time; agreement was greatest for 2-year-old deer (84%) and declined to 52–69% for ≥3-year-old deer. When an observer had different ages for mandibles from the same deer, using the younger age would increase accuracy for 2-year-old deer and vice versa for 4–8-year-old deer (Fig. 2), but only by a few percentile points (2–9%). The TRW estimates were within ±1 year for 87% of known-age mandibles.

|
CA method
Accuracy of CA ranged from 60% to 62% (n = 2 observers). There was a trend for accuracy to gradually decline up to 6 years old; 7- and ≥8-year-old individuals were correctly aged ~60–65% of the time (Table 4). Deer ≥3 years old that were misclassified were more likely to be underestimated than overestimated. Cementum annuli estimates were within ±1 year for 93% of incisors. Relative to TRW, variances of CA-estimated ages for most known-age classes were lower (Table 4).
Modified TRW technique
We categorised mandibles on the basis of overlap in wear characteristics among age classes and to minimise misclassification. Known ages were most correlated with tooth-wear ranks of the M1 (r2 = 0.78), relative to M2 (r2 = 0.74) and M3 (r2 = 0.54). Although the correlations between M1 and M2 were similar, M1 had lower and consistent standard errors among age classes ( = 0.18, range = 0.15–0.20) than M2 (
= 0.18, range = 0.15–0.20) than M2 ( = 0.28, range = 0.17–0.51, Fig. S4). Therefore, the bicuspid first molars (M1) appeared to have the most distinct characteristics that allowed for separation of age classes (Fig. 3). Deer of 2 years of age had M1 with no visible wear; 83% of mandibles fitting this description were known to be 2 years old. Deer of 3–5 years of age largely had dentine width ≥enamel width and retained a stair-step appearance between the lingual and buccal crests of the M1; 87% of mandibles fitting this description were known to be 3–5 years old. Jaws with M1 in which dentine >enamel and no stair-step appearance (i.e. infundibulum was only a small ridge or was absent) were primarily from deer ≥6 years of age; 94% of mandibles fitting this description were known to be ≥6 years old. We used these groupings to develop a modified ageing guide for deer ≥2 years old (Fig. 3).
= 0.28, range = 0.17–0.51, Fig. S4). Therefore, the bicuspid first molars (M1) appeared to have the most distinct characteristics that allowed for separation of age classes (Fig. 3). Deer of 2 years of age had M1 with no visible wear; 83% of mandibles fitting this description were known to be 2 years old. Deer of 3–5 years of age largely had dentine width ≥enamel width and retained a stair-step appearance between the lingual and buccal crests of the M1; 87% of mandibles fitting this description were known to be 3–5 years old. Jaws with M1 in which dentine >enamel and no stair-step appearance (i.e. infundibulum was only a small ridge or was absent) were primarily from deer ≥6 years of age; 94% of mandibles fitting this description were known to be ≥6 years old. We used these groupings to develop a modified ageing guide for deer ≥2 years old (Fig. 3).

|
We obtained an additional 62 mandibles from 32 known-age deer (2 (n = 4), 3–5 (n = 47) and ≥6 (n = 11) years old) to evaluate the modified TRW technique. Our modified TRW technique of dividing adults into three age classes yielded an overall accuracy of 72%. Individual biologists correctly categorised 57–82% of mandibles, an improvement over the previous evaluation. Average correct classifications were 73% for the 3–5-year-old age class and 68% for the ≥6-year-old age class (Table 5). Pooling the traditional TRW age estimates from the earlier six observers in our study for 3–5-year-old deer (n = 124) and ≥6-year-old deer (n = 79) resulted in accuracies of 48% and 28% respectively.

|
Discussion
Despite the many evaluations of TRW and CA methods for estimation of age in ungulates (DeYoung 1989; Hamlin et al. 2000; Gee et al. 2002), there is often little consensus beyond the detection of unmodelled variation in age estimates. In part, the ambiguity is due to limitations in sample size and failure to quantify factors that may influence estimated ages. The present study amassed one of the most extensive datasets of paired TRW and CA ages for harvested adult white-tailed deer as well as the largest published dataset of known-age adult white-tailed deer. Our results showed important insights into the sources of variation and limitations of both ageing methods.
Environmental and visual effects
We observed statistically significant variation in tooth wear attributable to several environmental and visual cues; however, the effect sizes were too small to be biologically meaningful. This is because the predicted effect of a given covariate needed to change estimated TRW ages by 1 full year exceeded the range of observed values. In some cases, the predicted effect was within empirical ranges, but larger than the 1 s.d. range, indicating that the influences on estimated TRW ages were mostly limited to extreme outliers of visual characteristics. For instance, individuals with abnormally small antler sizes or body mass for a given age class may influence age estimates. The lack of an effect of physical traits on estimated TRW ages suggests that observers were not consistently influenced by morphometrics. The effect of harvest date was significant for TRW ages 6 and 7, but the effect was too small to cause a ≥1-year change during the hunting season.
We observed a wide range in density of enhanced nutrition stations, which suggests that a measurable effect on tooth wear could have occurred. However, we observed no biologically meaningful effect of enhanced nutrition on estimated TRW ages. Previous studies have found that access to supplemental feed is not equal among individuals; only 23–56% of sampled male white-tailed deer consumed enhanced nutrition on three different sites in southern Texas (Bartoskewitz et al. 2003). Furthermore, dominant males are more likely to consume feed than are subdominant males (Bartoskewitz et al. 2003). We were unable to quantify the amount of feed consumed for each male; thus, detecting an effect of enhanced nutrition on tooth wear at the population level would have been difficult. Further, even with ad libitium access to enhanced nutrition, 28–61% of white-tailed deer diets may consist of natural vegetation (Darr et al. 2019). Nonetheless, given that we sampled wild deer with a presumably high variation in individual-specific feed-consumption rates, we cannot conclude that enhanced nutrition had no effect on an individual’s tooth wear, but rather had no consistent effect at the population level.
The soil represented a greater potential source of environmental effects on tooth wear because lifetime exposure to sandy soils should accumulate with age. Therefore, under this hypothesis, we expected an effect in the older age classes. Unlike other studies (Hewison et al. 1999; Hall et al. 2012), we found no statistically significant effect of soil. Hall et al. (2012) found that agreement between TRW and CA in fallow deer (Dama dama) was influenced by soil and whether animals primarily grazed or browsed; however, a single observer aged all deer. Thus, the lack of measurable effects in our environmental effects model may be because we did not use observer as a covariate. Observer was not used as a covariate because it would be difficult to determine who the wildlife biologist was for a given lease during the earlier years of data collection. This is an important consideration, given the variation in estimated TRW ages from mandibles among individual observers (Hewison et al. 1999; the present study). We were also unable to control for observers’ individual bias or perceptions, such as, for instance, the potential that some individuals consciously or unconsciously adjusted age estimates because of the widely held belief that environmental variables influence tooth wear.
Unmodelled variation is difficult to address for large-scale evaluations of estimated ages. However, recent research on dental micro-wear patterns in livestock have indicated that soil particles have minimal effect on tooth wear because plant matter is swallowed before rumination; thus, food is moistened and rinsed in the rumen before chewing (Merceron et al. 2016; Dittmann et al. 2017). Instead, the diet has a larger effect on tooth wear than do soil particles; the more siliceous phytoliths in plant matter, the greater the tooth wear (Merceron et al. 2016). White-tailed deer are small, browsing ruminants with a wide diet breadth; combined with seasonal and rainfall-induced variation in forage availability (Crider et al. 2015), it seems unlikely that diet would exert a consistent local effect on tooth wear. In fact, the variation in deer diet is the likely reason why we did not detect a meaningful drought effect on estimated ages or proportion of incisors with ‘A’ scores. Therefore, the hypothesis that soil particles influence tooth wear to the extent that age estimates are affected may be conceptually flawed.
Overall, we found no evidence that external factors influenced interpretation of tooth-wear patterns to the extent that age estimates were affected in a consistent manner. Some of our results differ from those of past studies that have found an environmental influence on tooth-wear patterns (Hewison et al. 1999; Hall et al. 2012), probably because there is a distinction between detectable tooth wear and biologically significant tooth wear (i.e. tooth wear influenced age by ≥1-year increment).
Accuracy evaluation
Our evaluation of ageing methods for southern free-ranging white-tailed deer was made possible by the collection of known-age mandibles (n = 264), including deer in the older age classes. Our sample size of jawbones from older known-age deer (≥6 years old, n = 41) was considerably larger than in other studies (n = 12, Gee et al. 2002; n = 10, Severinghaus 1949; n = 8, Hamlin et al. 2000). During our evaluation of the mandible ageing technique, we provided reference materials to aid in age determination, which tests the accuracy of the Severinghaus’ (1949) technique, not the ability of biologists to recall criteria within the paper. Ages of deer ≥2 years old in our study were correctly estimated only 49% of the time, being similar to the results of other studies (40%; Gee et al. 2002; 43%; Hamlin et al. 2000). Overall, there was a tendency to overestimate age of young deer and vice versa, being similar to findings of DeYoung (1989) and Hamlin et al. (2000). Further, our results agreed with previous studies, which indicated that accuracy of TRW ages declines for older deer because wear patterns vary among individuals within age classes (Hamlin et al. 2000; Gee et al. 2002). The variation in wear patterns among individuals was apparent and large enough to misclassify up to ±4 years. For instance, known-age 6-year-old deer fit nine different molar-wear categories (Table 1). Further, mandibles from the same deer often had different tooth-wear patterns, which suggests that some deer masticate more on one side of their mouth; there was no tendency to prefer the left or right side.
Clearly, the accuracy of Severinghaus’ (1949) method is limited by both inter- and intra-individual variation. The variation in tooth wear is apparent even with high-resolution measurements. Cooper et al. (2013) used calipers to measure characteristics of the third premolar tooth and found a high correlation between known age and dentine width (r2 = 0.73); however, only 48% of the ages estimated via a predictive equation were correct. Additionally, Meares et al. (2006) analysed three-dimensional measurements of molars and found that dentine:enamel ratios overlapped in each molar (M1–M3) from each of the known-age 2-, 3- and 4-year-old age classes. Nonetheless, given that TRW estimates were ±1 year of known ages 87% of the time, there is value in using this simple and rapid technique for research and management purposes.
Age estimates based on CA were less accurate for older deer in southern Texas than for deer from northern regions. Thomas and Bandy (1973) reported 100% accuracy (n = 32) for Columbian black-tailed deer (O. hemionus columbianus) aged ≥1 years old, and Hamlin et al. (2000) aged white-tailed deer from Montana with 85% accuracy (n = 74), which was >22 percentage points higher than our CA accuracy. Hamlin et al. (2000) correctly aged 8 of 12 (67%) white-tailed deer aged ≥5 years old, which was greater than the 53% (n = 88) of deer correctly aged in the present study. Annuli are a result of seasonal changes in animal physiology. Therefore, CA ages could be inaccurate in areas without strong seasonality (Low and Cowan 1963), such as southern Texas. Given that incorrect CA estimates were underestimates rather than overestimates for six of seven age classes, our results corroborated the suggestion that faint or non-existent annuli may cause an under-ageing bias in older deer in non-seasonal regions. However, CA were ±1 year of known ages 93% of the time, similar to moose (Alces alces; Rolandsen et al. 2008); thus, there is value in using incisors for CA to assign southern deer into broader age classes. Unlike the TRW method, the CA estimates can be used for deer ≥6 years old. In fact, the CA estimates showed that ~32% of the harvested deer aged as ≥6 years old via TRW were much older than expected. This has important implications for population dynamics and modelling, such as Foley et al. (2016), who observed that models of population growth did not parallel observed trends in population density unless the proportion of deer ≥6 years old was increased.
Modified TRW technique
Our modified TRW approach yielded an overall accuracy of 72%. Compared with pooled TRW estimates, the largest improvements came from the 3–5-year-old (73% vs 48% accuracy) and ≥6-year-old (68% vs 28%) age classes. Our modified TRW method improved on previous literature in three ways. First, we were able to place deer into age classes by using fewer criteria than in the traditional TRW method. This is beneficial because the more criteria employed, the more variation among individual deer and the more likely the criteria are to differ in the age they indicate. Our findings of intra- and inter-individual variation in tooth-wear patterns, in conjunction with subjective determination of tooth wear, suggests that fewer criteria to determine age would be beneficial. By using only the first molar (M1) to classify ages, biologists ageing live deer may be more accurate because this molar is more visible than the second and third molars, and is a simple and straightforward approach. Second, pooling into age classes is an explicit acknowledgement of the uncertainty in wear patterns. Classifying middle age (3–5) reduced bias towards under-aging deer, given that only 11% were underestimated versus 40–63% being underestimated with the original TRW method and the 34% being underestimated from the pooled TRW. Although all the incorrect estimates for ≥6-year-old deer were underestimates (32%), accuracy may be improved by using the age of the mandible with greater wear on M1 when both mandibles are available because the largest improvements in accuracies were in the 6-year-old (9%) and 7-year-old (8%) age classes. Finally, clear criteria for dividing deer mandibles into young (2), middle (3–5), and older (≥6) age classes aids research and management efforts. For instance, in southern Texas, males add body and antler mass until 6 years old (Hewitt et al. 2014) and fawn recruitment rates increase when females become ≥3 years old (Rice 2018). Using ≥2 years old as one continuous ‘adult’ category (Gee et al. 2002) would limit the resolution for population management. Further, greater resolution in ageing would be useful when studying dynamics of unhunted or lightly hunted deer populations because a greater proportion of mature deer would be expected.
Evaluation of assumptions
It is difficult to study long-term, large-scale effects on tooth-wear patterns in long-lived animals. Our set of known-age deer used in the accuracy evaluation is larger than any previously published, yet, it was not of sufficient geographic scale to test for effects of environmental variables. Given the substantial investment of time and effort to obtain known ages, the investment required to have a large sample of known-age deer to represent each environmental variable is prohibitive. Therefore, we were forced to make some assumptions about the distributions of TRW ages relative to CA ages in harvested deer. The main assumption for use of TRW minus CA ages as the response variable is that CA ages were not influenced by environmental variables. Fortunately, we were able to evaluate our assumptions on the basis of several different lines of evidence. First, comparison of animals exposed to different climactic conditions (extreme drought vs mesic conditions) showed no evidence for changes in the distribution of TRW ages and no indications that the visibility of CA was affected. Second, the evaluation of known-age deer confirmed that CA ages were both more accurate (greater % correct) and less variable (lower s.d.) than were TRW ages. Finally, the β coefficients for the effect of harvest date on TRW age were positive for 6–7-year-old deer, suggesting that we could indeed detect within-year differences in the amount of tooth wear. Therefore, we conclude that CA ages were not likely to be affected by environmental variables and our response variable was capable of detecting biologically meaningful changes in tooth-wear patterns.
Conclusions
Variation in tooth wear appears to be mainly attributable to inter- and intra-individual variation. The effect of environmental variables was insufficient to affect assigned ages in the presence of individual variation and the results of dental microwear studies suggest that the soil particle hypothesis may be conceptually flawed. Because we detected no meaningful environmental or visual-cue effect on estimating ages, no correction or adjustment was possible for the accuracy evaluation. To reduce errors based on TRW (42% accuracy) in populations with older age structures, it is recommended to group ages into ±1-year age classes when ageing via Severinghaus’ (1949) method (87% accuracy) or to use the modified TRW method, which is based on tooth wear of a single molar (M1, 72% accuracy). Errors in ages were mostly underestimates for older deer for TRW (55%) and to a lesser extent, CA (35%). When left and right mandibles indicate different ages, observers should use the older age.
Accuracy of CA (58%) was lower in southern latitudes than northern latitudes, possibly because of faint annuli resulting in underestimated ages. However, CA accuracy was much higher than TRW, within ±1 year of true age 93% of the time, and CA ages were less variable, as indicted by lower s.d. Therefore, CA is the better method for ageing ≥6-year-old deer. The known bias in under-ageing older white-tailed deer, roe deer (Hewison et al. 1999), and possibly other cervid species, is not well appreciated because the inherent bias in TRW results in a biased view of population demographics. For instance, in our TRW versus CA analysis, we found that ~30% of males TRW-aged as 6 and 7 years old were actually much older (8–16 years old). Given that we found that CA age of older deer were likely to be underestimated (Table 4), these 8–16-year-old deer may be even older. Knowing the true age structure would have implications for understanding disease dynamics, population modelling, and other survival-based analyses. The large proportion of old deer indicates the importance of adult survival in the maintenance of populations in the semiarid environment of South Texas, where inter-annual variation in recruitment is associated with precipitation patterns. Overall, observers should expect variation in tooth-wear patterns among individual deer; however, understanding biases for each ageing method can provide support for research and management decisions (Mysterud and Ostbye 2006).
Data availability
The data that support this study are available in FigShare at DOI 10.6084/m9.figshare.15044304.
Conflicts of interest
The authors have no conflicts of interest to declare.
Declaration of funding
A. R. Sanchez, Jr., Carl Rush, Carlos Y. Benavides, II, International Bank of Commerce, Joe Finley, King Ranch, Inc., and the Texas Parks and Wildlife Department funded this project.
Acknowledgements
We thank the cooperating landowners for access to their property. A. M. Foley was supported by the King Ranch, Inc., and the East Foundation during paper preparation. S. Hayslette, J. Baumgardt, F. Hernández and an anonymous reviewer provided valuable input on earlier drafts of the paper. B. Loflin assisted with macro-photography. This is publication number 20–112 from the Caesar Kleberg Wildlife Research Institute and 053 of the East Foundation.
References
Asmus, J., and Weckerly, F. W. (2011). Evaluating precision of cementum annuli analysis for aging mule deer from southern California. The Journal of Wildlife Management 75, 1194–1199.| Evaluating precision of cementum annuli analysis for aging mule deer from southern California.Crossref | GoogleScholarGoogle Scholar |
Bartoskewitz, M. L., Hewitt, D. G., Pitts, J. S., and Bryant, F. C. (2003). Supplemental feed use by free-ranging white-tailed deer in southern Texas. Wildlife Society Bulletin 31, 1218–1228.
Cook, R. L., and Hart, R. V. (1979). Ages assigned known-age Texas white-tailed deer: tooth wear versus cementum analysis. Proceedings of the Annual Conference of the Southeastern Association of Fish and Wildlife Agencies 33, 195–201.
Cooper, S. M., Sieckenius, S. S., and Silva, A. L. (2013). Dentine method: aging white-tailed deer by tooth measurements. Wildlife Society Bulletin 37, 451–457.
| Dentine method: aging white-tailed deer by tooth measurements.Crossref | GoogleScholarGoogle Scholar |
Crider, B. L., Fulbright, T. E., Hewitt, D. G., DeYoung, C. A., Grahmann, E. D., Priesmeyer, W. J., Wester, D. B., Echols, K. N., and Draeger, D. (2015). Influence of white-tailed deer population density on vegetation standing crop in a semiarid environment. The Journal of Wildlife Management 79, 413–424.
| Influence of white-tailed deer population density on vegetation standing crop in a semiarid environment.Crossref | GoogleScholarGoogle Scholar |
Darr, R. L., Williamson, K. M., Garver, L. W., Hewitt, D. G., DeYoung, C. A., Fulbright, T. E., Gann, K. R., and Draeger, D. A. (2019). Effects of enhanced nutrition on white-tailed deer foraging behavior. Wildlife Monographs 202, 27–34.
DelGiudice, G. D., Lenarz, M. S., and Powell, M. C. (2007). Age-specific fertility and fecundity in northern free-ranging white-tailed deer: evidence for reproductive senescence? Journal of Mammalogy 88, 427–435.
| Age-specific fertility and fecundity in northern free-ranging white-tailed deer: evidence for reproductive senescence?Crossref | GoogleScholarGoogle Scholar |
Demarais, S., and Strickland, B. K. (2011). Antlers. In ‘Biology and Management of White-tailed Deer’. (Ed. D. G. Hewitt.) pp. 107–145. (CRC Press: Boca Raton, FL, USA.)
DeYoung, C. A. (1988). Comparison of net-gun and drive-net capture for white-tailed deer. Wildlife Society Bulletin 16, 318–320.
DeYoung, C. A. (1989). Aging live white-tailed deer on southern ranges. The Journal of Wildlife Management 53, 519–523.
| Aging live white-tailed deer on southern ranges.Crossref | GoogleScholarGoogle Scholar |
DeYoung, C. A., Fulbright, T. E., Hewitt, D. G., Wester, D. B., and Draeger, D. A. (2019). Linking white-tailed deer density, nutrition, and vegetation in a stochastic environment. Wildlife Monographs 202, 1–63.
| Linking white-tailed deer density, nutrition, and vegetation in a stochastic environment.Crossref | GoogleScholarGoogle Scholar |
Dittmann, M. T., Kreuzer, M., Runge, U., and Clauss, M. (2017). Ingestive mastication in horses resembles rumination but not ingestive mastication in cattle and camels. Ecological and Integrative Physiology 327, 98–109.
| Ingestive mastication in horses resembles rumination but not ingestive mastication in cattle and camels.Crossref | GoogleScholarGoogle Scholar | 29356397PubMed |
Erickson, J. A., Anderson, A. E., Medin, D. E., and Bowden, D. C. (1970). Estimating ages of mule deer: an evaluation of technique accuracy. The Journal of Wildlife Management 34, 523–531.
| Estimating ages of mule deer: an evaluation of technique accuracy.Crossref | GoogleScholarGoogle Scholar |
Foley, A. M., Hewitt, D. G., DeYoung, C. A., DeYoung, R. W., and Schnupp, M. J. (2016). Modeled impacts of chronic wasting disease on white-tailed deer in a semi-arid environment. PLoS One 11, e0163592.
| Modeled impacts of chronic wasting disease on white-tailed deer in a semi-arid environment.Crossref | GoogleScholarGoogle Scholar | 27711208PubMed |
Foley, A. M., Hewitt, D. G., DeYoung, R. W., Schnupp, M. J., Hellickson, M. W., and Lockwood, M. A. (2018). Reproductive effort and success of males in scramble-competition polygyny: evidence for trade-offs between foraging and mate search. Journal of Animal Ecology 87, 1600–1614.
| Reproductive effort and success of males in scramble-competition polygyny: evidence for trade-offs between foraging and mate search.Crossref | GoogleScholarGoogle Scholar |
Folks, D. J., Gann, K., Fulbright, T. E., Hewitt, D. G., DeYoung, C. A., Wester, D. B., Echols, K. N., and Draeger, D. A. (2014). Drought but not population density influences dietary niche breadth in white-tailed deer in a semiarid environment. Ecosphere 5, 1–15.
| Drought but not population density influences dietary niche breadth in white-tailed deer in a semiarid environment.Crossref | GoogleScholarGoogle Scholar |
Gaillard, J.-M., Festa-Bianchet, M., and Yoccoz, N. G. (1998). Population dynamics of large herbivores: variable recruitment with constant adult survival. Trends in Ecology & Evolution 13, 58–63.
| Population dynamics of large herbivores: variable recruitment with constant adult survival.Crossref | GoogleScholarGoogle Scholar |
Gee, K. L., Holman, J. H., Causey, K., Rossi, A. N., and Armstrong, J. B. (2002). Aging white-tailed deer by tooth replacement and wear: a critical evaluation of a time-honored technique. Wildlife Society Bulletin 30, 387–393.
Gee, K. L., Webb, S. L., and Holman, J. H. (2014). Accuracy and implications of visually estimating age of male white-tailed deer using physical characteristics from photographs. Wildlife Society Bulletin 38, 96–102.
| Accuracy and implications of visually estimating age of male white-tailed deer using physical characteristics from photographs.Crossref | GoogleScholarGoogle Scholar |
Gilbert, F. F. (1966). Aging white-tailed deer by annuli in the cementum of the first incisor. The Journal of Wildlife Management 30, 200–202.
| Aging white-tailed deer by annuli in the cementum of the first incisor.Crossref | GoogleScholarGoogle Scholar |
Gilbert, F. F., and Stolt, S. L. (1970). Variability in aging Maine white-tailed deer by tooth-wear characteristics. The Journal of Wildlife Management 34, 532–535.
| Variability in aging Maine white-tailed deer by tooth-wear characteristics.Crossref | GoogleScholarGoogle Scholar |
Hall, G. P., Murray, P. J., Byrne, M. J., and Lisle, A. T. (2012). Is tooth wear a reliable means of aging wild European fallow deer in Tasmania, Australia? Wildlife Society Bulletin 36, 124–129.
| Is tooth wear a reliable means of aging wild European fallow deer in Tasmania, Australia?Crossref | GoogleScholarGoogle Scholar |
Hamlin, K. L., Pac, D. F., Sime, C. A., DeSimone, R. M., and Dusek, G. L. (2000). Evaluating the accuracy of ages obtained by two methods for Montana ungulates. The Journal of Wildlife Management 64, 441–449.
| Evaluating the accuracy of ages obtained by two methods for Montana ungulates.Crossref | GoogleScholarGoogle Scholar |
Hewison, A. J. M., Vincent, J. P., Angibault, J. M., Delorme, D., Laere, G. V., and Gaillard, J. M. (1999). Tests of estimation of age from tooth wear on roe deer of known-age: variation within and among populations. Canadian Journal of Zoology 77, 58–67.
| Tests of estimation of age from tooth wear on roe deer of known-age: variation within and among populations.Crossref | GoogleScholarGoogle Scholar |
Hewitt, D. G., Hellickson, M. W., Lewis, J. S., Wester, D. B., and Bryant, F. C. (2014). Age-related patterns of antler development in free-ranging white-tailed deer. The Journal of Wildlife Management 78, 976–984.
| Age-related patterns of antler development in free-ranging white-tailed deer.Crossref | GoogleScholarGoogle Scholar |
Jacobson, H. A., and Reiner, R. J. (1989). Estimating age of white-tailed deer: tooth wear versus cementum annuli. Proceedings of the Annual Conference of the Southeastern Association of Fish and Wildlife Agencies 43, 286–291.
Jacobson, H. A., DeYoung, C. A., DeYoung, R. W., Fulbright, T. E., and Hewitt, D. G. (2011). Management on private property. In ‘Biology and Management of White-tailed Deer’. (Ed. D. G. Hewitt.) pp. 453–480. (CRC Press: Boca Raton, FL, USA.)
Johnson, D. H. (1999). The insignificance of statistical significance testing. The Journal of Wildlife Management 63, 763–772.
| The insignificance of statistical significance testing.Crossref | GoogleScholarGoogle Scholar |
Kaiser, T. M., Brasch, J., Castell, J. C., Schulz, E., and Clauss, M. (2009). Tooth wear in captive wild ruminant species differs from that of free-ranging conspecifics. Mammalian Biology 74, 425–437.
| Tooth wear in captive wild ruminant species differs from that of free-ranging conspecifics.Crossref | GoogleScholarGoogle Scholar |
Kaiser, T. M., Muller, D. W., Fortelius, M., Schulz, E., Codron, D., and Clauss, M. (2013). Hypsodonty and tooth facet development in relation to diet and habitat in herbivorous ungulates: implications for understanding tooth wear. Mammal Review 43, 34–46.
| Hypsodonty and tooth facet development in relation to diet and habitat in herbivorous ungulates: implications for understanding tooth wear.Crossref | GoogleScholarGoogle Scholar |
Kozicky, E. L. (1997). A protein pellet feed-delivery system for white-tailed deer. Management Bulletin 1, Caesar Kleberg Wildlife Research Institute, Texas A&M University-Kingsville, Kingsville, TX, USA.
Low, W. A., and Cowan, I. M. (1963). Age determination of deer by annular structure of dental cementum. The Journal of Wildlife Management 27, 466–470.
| Age determination of deer by annular structure of dental cementum.Crossref | GoogleScholarGoogle Scholar |
Lyons, E. K., Schroeder, M. A., and Robb, L. A. (2012). Criteria for determining sex and age of birds and mammals. In ‘The Wildlife Techniques Manual, Vol. 1: Research’, 7th edn. (Ed. N. J. Silvy.) pp. 207–229. (The Johns Hopkins University Press: Baltimore, MD, USA.)
McCaffery, K. R. (2001). Evaluation of population estimation methods. In ‘Sandhill Whitetails: Providing New Perspective for Deer Management’. (Eds J. F. Kubisiak, K. R. McCaffery, W. A. Creed, T. A. Heberlein, R. C. Bishop, and R. E. Rolley.) pp. 136–154. (Wisconsin Department of Natural Resources: Madison, WI, USA.)
McCullough, D. R. (1996). Failure of the tooth cementum aging technique with reduced population density of deer. Wildlife Society Bulletin 24, 722–724.
Meares, J. M., Murphy, B. P., Ruth, C. R., Osborn, D. A., Warren, R. J., and Miller, K. V. (2006). A quantitative evaluation of the Severinghaus technique for estimating age of white-tailed deer. Proceedings of the Annual Conference of the Southeastern Association of Fish and Wildlife Agencies 60, 89–93.
Merceron, G., Ramdarshan, A., Blondel, C., Boisserie, J. R., Brunetiere, N., Francisco, A., Gautier, D., Milhet, X., Novello, A., and Pret, D. (2016). Untangling the environmental from the dietary: dust does not matter. Proceedings of the Royal Society of London. Series B, Biological Sciences 283, 20161032.
| Untangling the environmental from the dietary: dust does not matter.Crossref | GoogleScholarGoogle Scholar |
Miller, M. W., and Conner, M. M. (2005). Epidemiology of chronic wasting disease in free-ranging mule deer: spatial, temporal, and demographic influences on observed prevalence patterns. Journal of Wildlife Diseases 41, 275–290.
| Epidemiology of chronic wasting disease in free-ranging mule deer: spatial, temporal, and demographic influences on observed prevalence patterns.Crossref | GoogleScholarGoogle Scholar | 16107661PubMed |
Mysterud, A., and Ostbye, E. (2006). Comparing simple methods for ageing roe deer Capreolus capreolus: are any of them useful for management? Wildlife Biology 12, 101–107.
| Comparing simple methods for ageing roe deer Capreolus capreolus: are any of them useful for management?Crossref | GoogleScholarGoogle Scholar |
Nakagawa, S., and Cuthill, I. C. (2007). Effect size, confidence interval and statistical significance: a practical guide for biologists. Biological Reviews of the Cambridge Philosophical Society 82, 591–605.
| Effect size, confidence interval and statistical significance: a practical guide for biologists.Crossref | GoogleScholarGoogle Scholar | 17944619PubMed |
Nesbitt, W. H., and Wright, P. L. (1981). ‘Records of North American big game.’ (Boone and Crockett Club: Alexandria, VA, USA.)
Newsom, J. D., and Sullivan, J. S. (1968). Cyro-branding: a marking technique for white-tailed deer. Proceedings of the Southeastern Association of Fish and Wildlife Agencies 22, 128–133.
Norwine, J., and John, K. (2007). ‘The changing climate of South Texas, 1900–2100: Problems and prospects, impacts and implications.’ (CREST-RESSACA, Texas A&M University-Kingsville.)
Parent, C. J., Hernandez, F., Brennan, L. A., Wester, D. B., Bryant, F. C., and Schnupp, M. J. (2016). Northern bobwhite abundance in relation to precipitation and landscape structure. The Journal of Wildlife Management 80, 7–18.
| Northern bobwhite abundance in relation to precipitation and landscape structure.Crossref | GoogleScholarGoogle Scholar |
R Core Team (2013). ‘R: a language and environment for statistical computing.’ (R Foundation for Statistical Computing: Vienna, Austria.)
Ramsey, C. W., Steinbach, D. W., and Rideout, D. W. (1993). ‘Determining the age of a deer.’ (Texas Agricultural Extension Service, College Station: USA.)
Rice, M. (2018). Population parameters and stress in white-tailed deer in south Texas. Thesis. Texas A&M University-Kingsville, Kingsville, TX, USA.
Rolandsen, C. M., Solberg, E. J., Heim, M., Holmstrom, F., Solem, M. I., and Saether, B. E. (2008). Accuracy and repeatability of moose (Alces alces) age as estimated from dental cement layers. European Journal of Wildlife Research 54, 6–14.
| Accuracy and repeatability of moose (Alces alces) age as estimated from dental cement layers.Crossref | GoogleScholarGoogle Scholar |
Roseberry, J. L. (1980). Age determination of white-tailed deer in the Midwest – methods and problems. In ‘Proceedings of the 1979 Symposium. White-tailed deer population management in the north central states’. (Eds R. L. Hine and S. Nehls.) pp. 73–82. (The North Central Section of the Wildlife Society: Urbana, IL, USA.)
Roseberry, J. L., and Woolf, A. (1991). A comparative evaluation of techniques for analysing white-tailed deer harvest data. Wildlife Monographs 117, 3–59.
Schulz, E., Fraas, S., Kaiser, T. M., Cunningham, P. L., Ismail, K., and Wronski, T. (2013). Food preferences and tooth wear in the sand gazelle (Gazella marica). Mammalian Biology 78, 55–62.
| Food preferences and tooth wear in the sand gazelle (Gazella marica).Crossref | GoogleScholarGoogle Scholar |
Severinghaus, C. W. (1949). Tooth development and wear as criteria of age in white-tailed deer. The Journal of Wildlife Management 13, 195–216.
| Tooth development and wear as criteria of age in white-tailed deer.Crossref | GoogleScholarGoogle Scholar |
Severinghaus, C. W., and Cheatum, E. L. (1956). Life and times of white-tailed deer. In ‘The Deer of North America’. (Ed. W. P. Taylor.) pp. 57–186. (Stackpole: Harrisburg, Pennsylvania, USA and The Wildlife Management Institute: Washington, DC, USA.)
Solberg, E. J., and Sæther, B. E. (1994). Male traits as life-history variables: annual variation in body mass and antler size in moose (Alces alces). Journal of Mammalogy 75, 1069–1079.
| Male traits as life-history variables: annual variation in body mass and antler size in moose (Alces alces).Crossref | GoogleScholarGoogle Scholar |
Storm, D. J., Samuel, M. D., Rolley, R. E., Beissel, T., Richards, B. J., and Van Deelen, T. R. (2014). Estimating ages of white-tailed deer: age and sex patterns of error using tooth wear-and-replacement and consistency of cementum annuli. Wildlife Society Bulletin 38, 849–856.
| Estimating ages of white-tailed deer: age and sex patterns of error using tooth wear-and-replacement and consistency of cementum annuli.Crossref | GoogleScholarGoogle Scholar |
Texas National Resources Information System (TNRIS) (2019). Soils homepage. Available at https://tnris.org/data-catalog/entry/soils/ [verified 15 June 2018].
Thomas, D. C., and Bandy, P. J. (1973). Age determination of wild black-tailed deer from dental annulations. The Journal of Wildlife Management 37, 232–235.
| Age determination of wild black-tailed deer from dental annulations.Crossref | GoogleScholarGoogle Scholar |
United States Drought Monitor (2019). Time series home page. Available at https://droughtmonitor.unl.edu/Data/Timeseries.aspx [verified 3 March 2019].
Welch, B. L. (1951). On the comparison of several mean values: an alternative approach. Biometrika 38, 330–336.
| On the comparison of several mean values: an alternative approach.Crossref | GoogleScholarGoogle Scholar |