

Characterising a unique recreational hunting method: hound hunting of sambar deer (Cervus unicolor) in Victoria, Australia
Jordan O. Hampton A B * , Andrew J. Bengsen C D , Sebastien Comte C , Jason S. Flesch E , Simon D. Toop E , Christopher Davies F H and David M. Forsyth C G
A B * , Andrew J. Bengsen C D , Sebastien Comte C , Jason S. Flesch E , Simon D. Toop E , Christopher Davies F H and David M. Forsyth C G
A Faculty of Science, University of Melbourne, Parkville, Vic. 3052, Australia.
B Harry Butler Institute, Murdoch University, 90 South Street, Murdoch, WA 6150, Australia.
C Vertebrate Pest Research Unit, New South Wales Department of Primary Industries, 1447 Forest Road, Orange, NSW 2800, Australia.
D Biosphere Environmental Consultants, South Lismore, NSW 2480, Australia.
E Game Management Authority, Level 2, 535 Bourke Street, Melbourne, Vic. 3000, Australia.
F Australian Deer Association, PO Box 299, Warragul, Vic. 3820, Australia.
G School of Biological, Earth and Environmental Sciences, University of New South Wales, Sydney, NSW 2052, Australia.
H Present address: Parks Victoria, 65 Church Street, Morwell, Vic. 3840, Australia.
Wildlife Research 50(9) 657-668 https://doi.org/10.1071/WR22117
Submitted: 29 June 2022 Accepted: 3 November 2022 Published: 9 December 2022
© 2023 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Context: Scent-trailing dogs (‘hounds’) are used to hunt introduced sambar deer (Cervus unicolor) in south-eastern Australia, but little is known about this recreational hunting technique.
Aim: The aim of this study was to characterise hound hunting of sambar deer in Victoria, Australia. Our study had three objectives as follows: (1) to report the hunting success of the technique; (2) to estimate pursuit times and distances; and (3) to investigate the landscape features associated with kill sites.
Methods: Data were collected from four hound-hunting crews during 2020–2021. GPS data from collars fitted to hounds were used to quantify hunt duration (min) and hunt distance (m). Logistic regression was used to estimate the effects of deer sex and hound pack size on the probability of hunting success. We used a Bayesian multinomial regression resource selection function (RSF) to identify the characteristics of sites where deer were killed (elevation, aspect, and distances to water and roads).
Key results: The four hunting teams ranged in size from 2 to 10 people and from one to eight hounds. Of 136 hunts, most (88%) harvested a deer, with pursued deer escaping on the other 12% of occasions. Pursuit times and distances were highly variable, with pursuit time >60 min for 46% of hunts and pursuit distance >5 km for 30% of hunts. The probability of killing a pursued male and female deer were similar, and there was not a positive relationship between pack size and hunt success. The RSF showed that both male and female deer were more likely to be killed on steeper slopes and closer to roads.
Conclusions: Successful hound hunting of sambar deer involves pursuits of considerable duration and requires a network of roads, with geographical features associated with deer kills sites being related to hunter access.
Implications: Hound hunting may be a useful wildlife management tool for land managers, but further studies are needed to assess its efficacy for achieving management goals.
Keywords: animal welfare, anthropogenic impacts, domestic animals, human dimensions, introduced species, population control, stress, wildlife management.
Introduction
There is an increasing need to manage the undesirable impacts of wild deer (family Cervidae) in Australia (Davis et al. 2016; Forsyth et al. 2017). Currently, there are few methods available for deer impact mitigation in Australia, and most programs rely on lethal control, with shooting being the most common method of killing. Deer are subject to several forms of population control, all being based on lethal shooting. These include commercial harvesting on agricultural land (Watter et al. 2020), aerial shooting (Bengsen et al. 2022; Hampton et al. 2022a), professional ground-based culling (Comte et al. 2022a), and recreational hunting (Moloney et al. 2022). Recreational hunters kill large numbers of deer annually in Australia (Moloney et al. 2022) and New Zealand (Kerr and Abell 2014). For example, approximately 40 000 licensed hunters reported killing approximately 174 000 deer in the state of Victoria during the 2019 calendar year, the most recent calendar year not severely affected by COVID-19 restrictions (Moloney and Hampton 2020). The deer species most commonly harvested by recreational hunters in Victoria is the sambar deer (Cervus unicolor), accounting for approximately 131 000 (75%) of the deer harvested in 2019 (Moloney and Hampton 2020).
The sambar is Australia’s largest deer species (mean live mass of 140 kg for females, 220 kg for males; Harrison 2010; Leslie 2011). It is native to the Indian subcontinent but has established non-native populations in Australia and New Zealand (Long 2003). A large and increasing population of sambar deer occurs in eastern Victoria (Forsyth et al. 2018; Watter et al. 2020; Moloney et al. 2022), primarily inhabiting native Eucalyptus forests (Gormley et al. 2011; Comte et al. 2022b; Forsyth et al. in press).
Two main methods are used to recreationally hunt sambar deer in Victoria. Stalking, as for deer species globally (Aebischer et al. 2014), involves hunting of deer by hunters on foot. This method accounted for 81% of the 2019 Victorian sambar deer harvest (Moloney and Hampton 2020). The other recreational hunting method involves the use of a pack of purpose-trained domestic dogs (Canis familiaris) (‘hounds’) to facilitate hunting (Game Management Authority 2014). ‘Hound hunting’ of sambar deer relies on dogs to scent-trail, follow and bail sambar deer to allow strategically positioned hunters to shoot the deer (Game Management Authority 2014). In 2019, 19% of the recreational sambar deer harvest (i.e. 25 000) was taken by this method (Moloney and Hampton 2020). Hound hunting is allowed only for sambar deer and only in the state of Victoria (Game Management Authority 2014), where it has been practiced for more than a century (Mason 2008). Hound hunting is thought to be more effective than stalking in heavily timbered and steep forest, particularly in those with a dense understorey (Bentley 1957).
Hound hunting of sambar deer in Victoria is contentious. Pursuits of deer on public land may be visible and audible to other public land users, and hounds sometimes stray into prohibited areas. Animal welfare concerns, as for other hunting methods relying on the use of dogs (Orr et al. 2019), include those affecting the hounds, the deer they pursue, and any other non-target wildlife they chase or attack. Previous accounts of hound hunting indicated that the pursuit (‘chase’) phase of hunts can last up to 2.5 h (Bentley 1957; Mason 2008; Harrison 2010). Beyond these anecdotal accounts, we are not aware of any empirical data characterising this hunting method.
Management of recreational hunting requires understanding of the impacts of hunting for individual animals (Hampton and Hyndman 2019; Græsli et al. 2020). Accordingly, for most recreational hunting activities, published studies typically report basic metrics characterising the hunting methods used and their outcomes. Such metrics include the frequency of hunts that result in a deer being killed (Bradshaw and Bateson 2000), and the landscape features associated with successful hunts (Rowland et al. 2021). Furthermore, many studies have quantified metrics related to the fates of individual hunted deer, including average duration and distance of pursuit (Cederlund and Kjellander 1991).
Here, we provide the first evaluation of using hounds to hunt sambar deer in Victoria, Australia. Our preliminary study had three objectives as follows: (1) to report the hunting success of the technique; (2) to estimate the pursuit times and distances; and (3) to investigate the landscape features associated with kill sites. Further, the findings were intended to provide initial evidence regarding the strengths and weaknesses of hound hunting as a management tool for deer in Australia.
Materials and methods
Study areas and timing
Our study was conducted at four areas in eastern Victoria (Fig. 1) during April–November, the legal hound hunting season (Game Management Authority 2014), in 2020 and 2021. The four areas include the Highlands–Southern Falls and Highland–Northern Falls bioregions and contain a wide variety of floristic communities. Wet sclerophyll forests, characterised by Eucalyptus spp., Acacia dealbata and Pomaderris aspera with an understorey of Comprosma quadrifida, Correa lawrencia and Pittosporum bicolor, dominate the region (Costermans 1981). Several species of introduced ungulates are sympatric with sambar deer in eastern Victoria, including fallow deer (Dama dama), red deer (Cervus elaphus) and feral goats (Capra hircus). However, none of those species is as widespread or abundant as is sambar deer (Forsyth et al. 2015). Dingoes/wild dogs (Canis familiaris) commonly eat sambar deer (Forsyth et al. 2018), but the extent to which this is by scavenging carcasses (Forsyth et al. 2014) or preying on calves and juveniles (Forsyth et al. 2019) is unknown. The climate is temperate with winter-dominant, but variable, average annual rainfall between 400 mm and 1500 mm across the region (Bureau of Meteorology 2021). Mean summer temperatures (December to February) vary from 23°C to 27°C and mean winter temperatures (June to August) vary from 13°C to 16°C (Bureau of Meteorology 2021).

|
Hunting regulations
Several procedural documents govern hound hunting in Victoria (Game Management Authority 2014). Current regulations stipulate that hounds be of the following three recognised breeds: beagles, bloodhounds, and harriers (Game Management Authority 2014). The laws stipulate a maximum pack size of five hounds but allow the use of up to three additional pups (i.e. dogs under 12 months of age) in training. A maximum of 10 human hunters is allowed with each hound pack. Participants are also required to pass a hound-hunting test, which assesses hunters’ knowledge of relevant legislation (Game Management Authority 2014). To minimise exhaustion of pursued deer, it is not permitted to use a second team of scent-trailing hounds on a deer’s trail if the first team fails to locate the deer or loses the deer’s trail (Bureau of Animal Welfare 2017).
The same minimum rifle calibre and bullet mass prescriptions apply as for other forms of sambar deer hunting in Victoria, i.e. a rifle with a minimum calibre of 0.270 (6.85 mm) and a minimum projectile mass of 130 grains (8.45 g) (Hampton et al. 2022b). Although not being legal requirements, hunters are encouraged to harvest and carry out as much meat as possible from the carcasses of deer killed via hound hunting, and to not leave carcasses in waterways (Game Management Authority 2014). All contributing hunting crews were assumed to have complied with these prescriptions after being explicitly asked whether they were aware of, and would comply with them, with the awareness that the data that they provided could show violations if they had occurred.
Hunting methods
Hound hunting consists of the following three components: (1) finding fresh deer scent trails by driving four-wheel drive (4WD) tracks, hereafter ‘roads’, with hounds being restrained on the back of vehicles; (2) scent-trailing of deer by hounds, which we term the ‘chase’ component; followed by (3) shooting of bailed or pursued deer by hunters (Mason 2008). The composition of the recreational hunters in each crew on any given hunt is flexible, depending on availability. The hounds used in a given hunt are generally owned by one or more of the hunters present (Game Management Authority 2014). Scent trails are found by driving roads with hounds in vehicles and stopping when hounds vocalise, indicating that they have detected the scent of a deer. The hounds are then released from their vehicle, their GPS collars are activated, and they are released from restraint. Hunters are deployed according to the terrain and track network; typically hunters will follow the hounds on foot or drive to a strategic location where a game trail intersects a road. All hunters carry a hand-held GPS unit linked to the GPS collars worn by their hounds, to enable them to visualise the location of all hounds and to move to strategic shooting locations.
Hound-hunting crew recruitment
We used our professional and private networks to identify hound hunters likely to be willing to be involved in our study. We disproportionally targeted hunters who were members of one organised recreational hunting association, the Australian Deer Association. We selected which crews to approach on the basis of geographic separation in their hunting areas. We approached four crews and all agreed to participate in the study. The four areas hunted by the four crews were Omeo (Crew 1), Jamieson (Crew 2), Buchan (Crew 3) and Dargo (Crew 4; Fig. 1).
Field data collection
Participating crews collected the following data from each hunt on paper data sheets: (a) date of hunt; (b) number of hunters; (c) number of hounds (adults and pups); (d) outcome of hunt (deer escaped without wounding, wounded (shot but escaped) or killed; Hampton et al. 2022c); (e) shot distance (the straight-line distance between the shooter and the deer at the time of shooting, measured by GPS or range finder; Hampton et al. 2022b); (f) number of shots; (g) sex of deer killed (from external genitalia and from the presence of antlers); (h) estimated age of deer killed (adult or juvenile based on mass and, for males, antler characteristics); and (i) the latitude and longitude of the site at which deer were killed (‘kill site’), recorded from the crews’ hand-held GPS units.
Hound telemetry devices
We also collected data from a telemetry collar worn by the ‘lead hound’ (the dominant dog in each pack as designated by the hunters) in each hunt. Hunters used Garmin Astro GPS collars (Fig. 2) and hand-held GPS units (Sepúlveda et al. 2015) as part of a ‘handheld tracking system for sporting dogs’ (Garmin, KS, USA). Collars weighed 260 g (1–3% of the body weight of a typical adult hound). Information collected from tracking collars was restricted to factory-standard ‘hunt metrics’ supplied by the Garmin Astro system. Two metrics were recorded from the ‘lead hound’ for each hunt. The first was ‘hunt distance’, derived from a track log generated by the GPS collar recording a location every 5 s, which we designated ‘chase distance’ (i.e. the total distance the dog travelled during a hunt, km). The second was ‘hunt time’, which we designated ‘chase time’ (CT, min). This was the interval between the start of a hunt, the onset of the second phase of the hunt when hounds are released from a vehicle and begin to scent-trail a deer, and that deer being killed or the hunt being abandoned (Cederlund and Kjellander 1991; Vaughan and Inman 2002).

|
Data analysis
Expected values for key variables including chase distance, shot distance, number of hunters in a crew, number of hounds in a pack, and number of deer harvested were described using arithmetic means and standard deviations. We used Bayesian logistic regression to estimate the probability of successfully killing a deer across all hunts. We also used Bayesian logistic regression to estimate the effects of deer sex and hound pack size on the probability of success. For this model, we specified separate intercepts for deer sex and a common slope for the effect of pack size. Pack size was centred on the minimum pack size of one hound to aid interpretation of the slope parameter because a pack size of zero hounds has no inherent meaning. Ten cases (7.2%) with missing covariate data were discarded. Logistic regression models were fitted using JAGS (Plummer 2003) via the runjags package (ver. 2.0.4-6) (Denwood 2016) in the R statistical environment (R Core Team 2020). Parameters were estimated using four chains of 10 000 Markov-chain Monte Carlo (MCMC) draws after discarding 5000 burn-in draws.
We used a Kaplan–Meier cumulative hazard model (Kaplan and Meier 1958) to estimate CT for successful and unsuccessful hunts after discarding two cases (1.4%) with missing covariate data. Estimates and confidence intervals were calculated using the survival package (Therneau 2020) in the R statistical environment (R Core Team 2020). We used logistic regression to estimate the proportion of chases that lasted <60 min or <5 km.
Kill-site characteristics
We used a Bayesian multinomial regression resource-selection function (RSF; Diefenbach et al. 2005) to identify the characteristics of kill sites for the two hunting crews for which we had >15 kill sites (i.e. Omeo and Dargo). For each area, we used a Bayesian multinomial regression with male and female kill sites as two independent used resources. We then defined the available resources by generating random points (three times the number of kill sites) within a buffer around each known kill site (Northrup et al. 2013) to achieve representative coverage of the available landscape (Fig. 3). In the absence of predefined boundaries for the hound hunting activity, we selected the buffer size for the available resources as the maximum nearest neighbour distance among all kill sites in each area (i.e. 1802 m in Omeo and 2324 m in Dargo).

|
We characterised the resources (used and available) in both areas by using the following six environmental covariates (spatial resolution 100 × 100 m): elevation (m above sea level); slope (degrees); woody vegetation cover (1–100%); distance to road (m); distance to watercourse (m); sin(aspect) → west–east, and; cos(aspect) → south–north. Further details on these environmental covariates, including their sources, are provided in Supplementary material Table S1. The 1-ha spatial resolution was chosen because it approximated typical ‘flight distances’ between where deer of the size of sambar are shot and where they fall (Stokke et al. 2018; Hampton et al. 2022b). Pearson correlation coefficients indicated that none of these six covariates was significantly correlated at either the Omeo or Dargo areas (Table S2). Hence, we used all six covariates in our analyses. For each of the two models (Omeo and Dargo), we ran three MCMC chains with 5000 adaptation runs and 5000 burn-in runs for a total sample size of 50 000. We assessed the mixing of the MCMC chains visually and with the Gelman–Rubin diagnostic (R; Gelman and Rubin 1992).
Results
Data scope
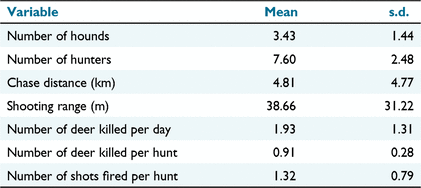
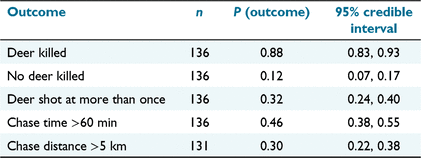
The four hunting crews collected data from 136 chases on 60 hunting days. Crew sizes varied from 2 to 10 hunters, and hound pack sizes varied from one to eight (including pups). Mean crew size was 7.60 (s.d. = 2.48) for hunters and 3.43 (s.d. = 1.44) for hounds (Table 1). In total, 122 deer were killed but two deer were shot when opportunistically encountered in the forest rather than having been pursued by hounds, and were excluded from the analysis. Hence, the mean number of sambar deer harvested per crew per hunting day was 1.93 (s.d. = 1.31, range 0–5). There were no instances in which more than one deer was harvested by the one hunting crew from a single chase. The majority (72%) of killed deer were adults, with a male:female sex ratio of 51:49. Mean shooting distance was 39 m (s.d. = 31 m), with the majority of shots from <50 m (Fig. 4). The frequency of more than one shot being fired at a deer was 0.32 (95% credible interval = 0.24, 0.40; Table 2), with the mean number of shots fired at each deer chased being 1.32 (s.d. = 0.79; Table 1).
|

|
|
Hunting success
The overall probability of success across all 136 hunts was 0.88 (95% credible interval [CrI] = 0.83, 0.93; Table 2). Twelve hunts (8.8%) concluded with deer escaping without shots being fired at them when pursuits were abandoned or hounds lost their trail. No deer were reported as having escaped wounded after shooting. Four deer (2.9%) were shot at, missed and escaped (i.e. escaped unwounded, see Hampton et al. 2022b). Mean hound hunter efficiency was 0.35 (s.d. = 0.34) deer harvested per hunter per hunting day. Logistic regression showed no evidence of a difference in the probability of hunting success between female (0.92, 95% CrI = 0.79, 0.99) and male (0.88, 95% CrI = 0.73, 0.97) deer for a chase, using a single hound. The probability of a positive relationship between hunting success and the number of hounds used (i.e. β > 0) was only 0.54. Hence, each additional hound, up to a maximum of seven hounds (including pups), had no discernible effect on the odds that a chase would be successful (odds ratio = 1.04, 95% CrI = 0.65, 1.58).
Chases
Tracking data from GPS collars on lead hounds indicated that most chases (70%, 95% CrI = 62, 78%) were <5 km in length, although some were >20 km (Fig. 5). Logistic regression indicated that a small majority of chases (54%, 95% CrI = 45, 62%) lasted less than 60 min (Table 2 and Fig. 6). The Kaplan–Meier survival model showed that unsuccessful hunts lasted longer, on average (median chase time = 108 min, 95% CI = 90, 330 min), than did successful hunts (median chase time = 45 min, 95% CI = 40, 65 min; Fig. 6).

|

|
Spatial distribution of kill sites
We performed RSF for deer killed in Omeo, n = 19 (hound-hunting Crew 1 in Fig. 1) and Dargo, n = 89 (hound-hunting Crew 4 in Fig. 1). The spatial distribution of kill sites in our study varied both between sites and between sexes. In Omeo, hound hunters killed 8 male and 11 female sambar deer. There, hunters were more likely to kill male deer on steep westerly slopes with high levels of woody vegetation (Table 3). Male deer were also more likely to be killed near roads. Opposingly, given the same available landscape as for males, there was no evidence of habitat associated with kill sites of female deer. In Dargo, hunters killed 50 male deer and 43 female deer and kill sites for both sexes showed a similar association with habitat. Sambar deer were more likely to be killed on steep slopes and near roads (Table 4).

|

|
Discussion
This is the first study quantifying recreational hound hunting of sambar deer in Victoria, Australia. We found that deer were harvested in the majority of hunts. Chase times and distances were highly variable, and deer were more likely to be killed on steep slopes, in areas with woody vegetation cover, and close to roads.
Relative to other published methods relying on the use of dogs to pursue hunted deer, sambar deer hound hunting was relatively efficient, with 88% of hunts resulting in the harvest of a deer. This was markedly higher than the 46% (n = 170) observed for hound hunting of red deer (Cervus elpahus) in the United Kingdom (Bradshaw and Bateson 2000). However, this measure of efficiency reflects only the success of each hunting crew, not individual hunters. The mean hound hunter success rate in our study (0.35 deer harvested per hunter per hunting day) was less than that estimated for all hound hunters in Victoria (0.45) in 2019 (Moloney et al. 2022), and was considerably lower than the efficiency of all deer hunting (including stalking; 0.50) estimated for the same year (Moloney and Hampton 2020). Caley and Ottley (1995) reported identical hunting success (88%) for use of hunting ‘pig’ dogs to capture feral pigs (Sus scrofa) in Northern Territory, Australia. That study reported a significantly male-biased harvest, but analysed data from one-dog team (Caley and Ottley 1995), whereas we examined data from four independent teams.
Sambar deer were more likely to be killed on steep slopes, near roads and in woody cover. The non-random distribution of deer kill sites is likely to be a characteristic of the hunting method rather than of sambar deer distribution. Conversations with the four hunting crews indicated that the proximity to roads is likely to reflect the use of roads by hunters to approach and shoot deer that are bailed by hounds. Finding a positive association between kill sites and the presence of roads is consistent with hunters using vehicles to position themselves for strategic shots along deer escape routes (Steyaert et al. 2016). It may also reflect the use of roads as a (presumably) faster way to find deer-scent trails than is looking for scent trails on foot. Accessibility has been associated with increased hunting success for recreational deer hunters in the United States (Lebel et al. 2012; Rowland et al. 2021). The positive association of steep slopes with the location of kill sites may be due to sambar deer moving more slowly up steep hills and therefore representing an easier target for the hunters (Lone et al. 2014). Steep slopes and dense woody vegetation could also represent an escape strategy for deer trying to outpace the chasing hounds. Another reason could be that deer feel safer on steeper slopes and in dense vegetation when chased by hounds (Bongi et al. 2008). A common escape strategy used by sambar deer is to cross rivers and streams (Fig. 2), apparently in an attempt to throw the hounds off their scent trail (Mason 2008), but there was no evidence of association between watercourses and kill sites in our study. This is most likely due to us analysing only kill-site location data and not the lead hounds’ complete track log. Hunting prescriptions encourage the removal of meat from carcasses, and discourage hunters from leaving carcasses in waterways (Game Management Authority 2014), and this could also have influenced where hunters shot deer.
Our results suggest that there is minimal sex bias in sambar deer harvested by recreational hound hunters in Victoria. This conclusion is supported by historical records. Bentley (1957) described the activities of one Victorian hound hunting pack from 1954 to 1956 and reported that a total of 134 deer was killed, with a slightly male-biased sex ratio of 56:44. In contrast, sambar deer killed by other shooting methods in Victoria (i.e. culling, commercial harvesting and recreational stalking) had a slightly female-biased sex ratio (55:45, n = 144; Watter et al. 2020). We are unaware of any data on the sex ratio of live (as opposed to killed) sambar deer populations in Australia. If the eastern Victoria sambar deer population is female-biased, as some populations in the native range are (Ramesh et al. 2012), then hound hunting may involve the selective harvest of males.
Mean shooting distance was only 39 m for hound hunting, which is markedly lower than the average of 111 m recorded for ground-based shooting of the same species without the aid of hounds (Hampton et al. 2022b). This result suggests that hound hunting allows shooters to get much closer to deer than is possible through diurnal stalking or nocturnal culling (Comte et al. 2022a). The relatively high frequency of repeat shooting (>1 shot fired at a deer) of 32% for hound hunting (compared with 7–14% for stalking and culling; Hampton et al. 2022b) is likely to reflect hunters shooting at moving deer and a greater ability to take follow-up shots at deer when pursued or bailed by hounds.
Average sambar deer pursuits were of considerable length as measured by distance (>4 km) and duration (>90 min). The longest of these were >20 km (Fig. 5) and >7 h (Fig. 6) respectively. It is possible that multiple deer were involved in some hunts, allowing some deer to be chased for only a proportion of our reported chase times before escaping, whereas other deer may have been chased multiple times. Unfortunately, the data we collected could not be used to quantify these outcomes. Chase distances were lower than for other deer species studied overseas. For example, red deer hunted with hounds in the United Kingdom in the 1990s ran an average of 19 km before being killed (Bateson and Bradshaw 1997). However, the duration of stress imposed by hound hunting is considerably longer than that associated with other deer killing methods used in Australia, notably professional nocturnal vehicle-based culling of rusa deer (Cervus timorensis) (Hampton et al. 2022c) and aerial shooting of fallow (Dama dama) and chital deer (Axis axis) (Hampton et al. 2022a).
For most species, physiological exertion imposed over timeframes of hours raises animal-welfare concerns (Le Grand et al. 2019). We did not collect biological samples from sambar deer pursued by hounds, but it is likely that the physiological effects of pursuit (as measured by the concentrations of markers of exertion in blood) would be similar to those documented in red deer (Bradshaw and Bateson 2000; Gentsch et al. 2018) and moose (Alces alces) (Græsli et al. 2020) pursued by scent-trailing hounds.
Animal-welfare implications for hound hunting include those affecting the deer (Bradshaw and Bateson 2000), the hounds (Orr et al. 2019), and any non-target wildlife that may be disturbed or harassed (Allen et al. 2019). These animal-welfare concerns are common to any recreational hunting methods that use dogs to pursue animals (Hampton and Hyndman 2019). Our study did not attempt to elucidate outcomes for hunting dogs or non-target wildlife species, but did allow some inference regarding impacts on deer. There were no reports of non-fatal wounding in this study, likely because shooting distances were small relative to other those of deer hunting methods, and the presence of hounds permitted hunters to track wounded deer to allow repeat shooting (Gentsch et al. 2018). However, the presence of some ‘gundogs’ has been associated with an increase in the frequency of non-fatal wounding for recreational hunting of other deer species (Godwin et al. 2013). Non-fatal wounding does not appear to be a central animal-welfare concern for hound hunting as it is for other deer management methods, such as nocturnal shooting from vehicles (Hampton et al. 2022c). However, pursuit is likely to cause animal-welfare impacts.
Other authors have speculated that the capacity for deer to cope physiologically with being pursued by hounds is shaped by their evolutionary or individual histories (Bateson and Bradshaw 1997). In this context, sambar deer have evolved with a suite of large mammalian predators, such as, for example, tigers (Panthera tigris) in southern Asia (Hayward et al. 2012) and are likely to be harrassed or attacked occasionally by dingoes/wild dogs in contemporary south-eastern Australia (Forsyth et al. 2019). Although the stress experienced prior to death is likely to be significant for sambar deer hunted with hounds, the impact of hunting on chronic stress of surviving deer is unknown (Sauerwein et al. 2004). Bradshaw and Bateson (2000) questioned what proportion of red deer that are hunted but escape die from severe physiological injuries (i.e. exertional myopathy and hyperthermia) sustained during a hunt. Our study was unable to address this question for sambar deer. To better understand animal-welfare impacts, future studies of hound hunting could collect blood samples to measure cortisol concentrations (or other physiological stress markers; Bradshaw and Bateson 2000). Alternatively, captured sambar deer could be equipped with ruminal temperature loggers, subcutaneous heart rate loggers and GPS collars with accelerometers, as was undertaken for moose in Sweden (Græsli et al. 2020).
There is interest in increasing the role of Australian recreational hunters in programs aiming to reduce the undesirable impacts of sambar deer (Australian Senate 2021), but only stalkers have been involved in management programs (Comte et al. 2022a). Hound hunting could prove useful for removing sambar deer surviving after other techniques such as stalking and aerial shooting have been used. However, our results suggest that hound hunting is most likely to be useful in areas with a developed road network. Land-use types such as timber (forestry) plantations (Davis et al. 2017) might be amenable. However, hound hunting may be problematic to employ on public land close to adjoining private land because of the risk of hounds (and pursued deer) crossing fences into livestock grazing properties (Game Management Authority 2014). Dog-assisted hunting has been widely used to eradicate feral pigs on islands (Cruz et al. 2005), and was used in the final stage of red deer eradication on Secretary Island, New Zealand (Macdonald et al. 2019). If hound hunting was to be used to control or eradicate sambar deer in remote areas, then a track network would need to be established to facilitate the rapid movement of hunters, as was done on Secretary Island (Macdonald et al. 2019).
Our study had several limitations. First, our sample size was relatively small (n < 150), so our ability to robustly estimate the frequency of infrequent events such as non-fatal wounding (no occurrences were reported) was limited (Hampton et al. 2019). Second, we relied on data collected by hunters rather than by independent observers (e.g. Bradshaw and Bateson 2000). This could have biased our results towards underestimation of ambiguous events with negative connotations (e.g. non-fatal wounding). Third, selection of hunting crews was non-random, and it is possible that the outcomes achieved by the crews that volunteered to participate in this study may have been better than in the general hunter population. Fourth, the hunters estimated (rather than measured) the ages of deer. Last, we did not have access to the complete track logs of lead hounds, limiting our analyses to the total time and distance of the hunt and, for successful hunts, to the kill site. Hence, our inferences relating to landscape features reflect sites where hunters prefer to take shots at deer, rather than the habitat deer use during a hunt (Lebel et al. 2012; Comte et al. 2022a).
Conclusions
Successful hound hunting requires a network of roads, potentially limiting the usefulness of this method for controlling deer in remote areas. Hound hunting chase times and distances are long relative to those of other deer hunting methods, but non-fatal wounding was not reported.
Supplementary material
Supplementary material is available online.
Data availability
The data that support this study are not publicly shared due to privacy reasons but may be shared upon reasonable request to the corresponding author if appropriate.
Conflicts of interest
David Forsyth is a guest Associate Editor of Wildlife Research but was blinded from the peer-review process for this paper. Three authors are employees of government agencies involved with the management of deer hunting activities in the state of Victoria, Australia: Jason Flesch and Simon Toop work for the Victorian Game Management Authority, and Chris Davies works for Parks Victoria. Their employment could not reasonably have interfered with the full and objective presentation of this research.
Declaration of funding
This project was funded by the Victorian Game Management Authority and the McKenzie Fellowship Program of the University of Melbourne.
Acknowledgements
This was an observational study of legally compliant recreational hunting activities and did not require Animal Ethics Committee (AEC) approval. We thank Danny Edebohls, Chris Eaton, Corey Lewis, Craig Hill and their colleagues for collecting data. We thank Zachary Powell, Barry Howlett, Bronwyn Orr and Jon Arnemo for initial discussions that led to the conception of this project. The Australian Deer Association assisted with outreach to hunters.
References
Aebischer, NJ, Wheatley, CJ, and Rose, HR (2014). Factors associated with shooting accuracy and wounding rate of four managed wild deer species in the UK, based on anonymous field records from deer stalkers. PLoS ONE 9, e109698.| Factors associated with shooting accuracy and wounding rate of four managed wild deer species in the UK, based on anonymous field records from deer stalkers.Crossref | GoogleScholarGoogle Scholar |
Allen, BL, Allen, LR, Ballard, G, Drouilly, M, Fleming, PJS, Hampton, JO, Hayward, MW, Kerley, GIH, Meek, PD, Minnie, L, O’Riain, MJ, Parker, DM, and Somers, MJ (2019). Animal welfare considerations for using large carnivores and guardian dogs as vertebrate biocontrol tools against other animals. Biological Conservation 232, 258–270.
| Animal welfare considerations for using large carnivores and guardian dogs as vertebrate biocontrol tools against other animals.Crossref | GoogleScholarGoogle Scholar |
Australian Senate (2021) ‘Impact of feral deer, pigs and goats in Australia.’ (Australian Government: Canberra, ACT, Australia)
Bateson, P, and Bradshaw, EL (1997). Physiological effects of hunting red deer (Cervus elaphus). Proceedings of the Royal Society of London B: Biological Sciences 264, 1707–1714.
| Physiological effects of hunting red deer (Cervus elaphus).Crossref | GoogleScholarGoogle Scholar |
Bengsen, AJ, Forsyth, DM, Pople, A, Brennan, M, Amos, M, Leeson, M, Cox, TE, Gray, B, Orgill, O, Hampton, JO, and Crittle, T (2022). Effectiveness and costs of helicopter-based shooting of deer. Wildlife Research , .
| Effectiveness and costs of helicopter-based shooting of deer.Crossref | GoogleScholarGoogle Scholar |
Bentley, A (1957). A brief account of the deer in Australia. The Journal of Wildlife Management 21, 221–225.
| A brief account of the deer in Australia.Crossref | GoogleScholarGoogle Scholar |
Bongi, P, Ciuti, S, Grignolio, S, Del Frate, M, Simi, S, Gandelli, D, and Apollonio, M (2008). Anti-predator behaviour, space use and habitat selection in female roe deer during the fawning season in a wolf area. Journal of Zoology 276, 242–251.
| Anti-predator behaviour, space use and habitat selection in female roe deer during the fawning season in a wolf area.Crossref | GoogleScholarGoogle Scholar |
Bradshaw, EL, and Bateson, P (2000). Welfare implications of culling red deer (Cervus elaphus). Animal Welfare 9, 3–24.
Bureau of Animal Welfare (2017) ‘Code of practice for the welfare of animals in hunting.’ (Department of Jobs, Precincts and Regions: Melbourne, Vic., Australia)
Bureau of Meteorology (2021) ‘Average annual, seasonal and monthly rainfall.’ (Bureau of Meteorology: Melbourne, Vic., Australia)
Caley, P, and Ottley, B (1995). The effectiveness of hunting dogs for removing feral pigs (Sus scrofa). Wildlife Research 22, 147–154.
| The effectiveness of hunting dogs for removing feral pigs (Sus scrofa).Crossref | GoogleScholarGoogle Scholar |
Cederlund G, Kjellander P (1991) Effects of chasing by hunting dogs on roe deer. In ‘Transactions of the 20th Congress of the International Union of Game Biologists’ (Eds S Csanyi, J Emhaft) pp. 363–370. (University of Agricultural Science: Gödöllö, Hungary)
Comte, S, Thomas, E, Bengsen, AJ, Bennett, A, Davis, NE, Brown, D, and Forsyth, DM (2022a). Cost-effectiveness of volunteer and contract ground-based shooting of sambar deer in Australia. Wildlife Research , .
| Cost-effectiveness of volunteer and contract ground-based shooting of sambar deer in Australia.Crossref | GoogleScholarGoogle Scholar |
Comte, S, Thomas, E, Bengsen, AJ, Bennett, A, Davis, NE, Freney, S, Jackson, SM, White, M, Forsyth, DM, and Brown, D (2022b). Seasonal and daily activity of non-native sambar deer in and around high-elevation peatlands, south-eastern Australia. Wildlife Research 49, 659–672.
| Seasonal and daily activity of non-native sambar deer in and around high-elevation peatlands, south-eastern Australia.Crossref | GoogleScholarGoogle Scholar |
Costermans LF (1981) ‘Native trees and shrubs of south-eastern Australia.’ (Rigby Publishers: London, UK)
Cruz, F, Donlan, CJ, Campbell, K, and Carrion, V (2005). Conservation action in the Galapagos: feral pig (Sus scrofa) eradication from Santiago Island. Biological Conservation 121, 473–478.
| Conservation action in the Galapagos: feral pig (Sus scrofa) eradication from Santiago Island.Crossref | GoogleScholarGoogle Scholar |
Davis, NE, Bennett, A, Forsyth, DM, Bowman, DMJS, Lefroy, EC, Wood, SW, Woolnough, AP, West, P, Hampton, JO, and Johnson, CN (2016). A systematic review of the impacts and management of introduced deer (family Cervidae) in Australia. Wildlife Research 43, 515–532.
| A systematic review of the impacts and management of introduced deer (family Cervidae) in Australia.Crossref | GoogleScholarGoogle Scholar |
Davis, PB, Munn, IA, Henderson, JE, and Strickland, BK (2017). Economic tradeoffs of managing for timber production or wildlife habitat. The Journal of Wildlife Management 81, 1363–1371.
Denwood, MJ (2016). Runjags: An R package providing interface utilities, model templates, parallel computing methods and additional distributions for MCMC models in JAGS. Journal of Statistical Software 71, 1–25.
| Runjags: An R package providing interface utilities, model templates, parallel computing methods and additional distributions for MCMC models in JAGS.Crossref | GoogleScholarGoogle Scholar |
Diefenbach, DR, Finley, JC, Luloff, AE, Stedman, R, Swope, CB, Zinn, HC, and San Julian, GJ (2005). Bear and deer hunter density and distribution on public land in Pennsylvania. Human Dimensions of Wildlife 10, 201–212.
| Bear and deer hunter density and distribution on public land in Pennsylvania.Crossref | GoogleScholarGoogle Scholar |
Forsyth DM, Bilney RJ, Bennett A (in press) Sambar deer. In ‘Strahan’s Mammals of Australia’. (Eds A Baker, I Gynther) (New Holland Publishers: Sydney, NSW, Australia)
Forsyth, DM, Woodford, L, Moloney, PD, Hampton, JO, Woolnough, AP, and Tucker, M (2014). How does a carnivore guild utilise a substantial but unpredictable anthropogenic food source? Scavenging on hunter-shot ungulate carcasses by wild dogs/dingoes, red foxes and feral cats in south-eastern Australia revealed by camera traps. PLoS ONE 9, e97937.
| How does a carnivore guild utilise a substantial but unpredictable anthropogenic food source? Scavenging on hunter-shot ungulate carcasses by wild dogs/dingoes, red foxes and feral cats in south-eastern Australia revealed by camera traps.Crossref | GoogleScholarGoogle Scholar |
Forsyth DM, Stamation K, Woodford L (2015) ‘Distributions of sambar deer, rusa deer and sika deer in Victoria.’ (Arthur Rylah Institute: Melbourne, Vic., Australia)
Forsyth DM, Pople T, Page B, Moriarty A, Ramsey D, Parkes J, Wiebkin A, Lane C (Eds) (2017) ‘2016 National Wild Deer Management Workshop Proceedings’, 17–18 November 2016, Adelaide, SA, Australia. (Invasive Animals Cooperative Research Centre: Canberra, ACT, Australia)
Forsyth, DM, Caley, P, Davis, NE, Latham, ADM, Woolnough, AP, Woodford, LP, Stamation, KA, Moloney, PD, and Pascoe, C (2018). Functional responses of an apex predator and a mesopredator to an invading ungulate: dingoes, red foxes and sambar deer in south-east Australia. Austral Ecology 43, 375–384.
Forsyth, DM, Latham, ADM, Davis, NE, Caley, P, Letnic, M, Moloney, PD, Woodford, LP, and Woolnough, AP (2019). Interactions between dingoes and introduced wild ungulates in Australia: concepts, evidence and predictions. Australian Mammalogy 41, 12–26.
| Interactions between dingoes and introduced wild ungulates in Australia: concepts, evidence and predictions.Crossref | GoogleScholarGoogle Scholar |
Game Management Authority (2014) ‘A guide to the use of hounds for hunting sambar deer in Victoria.’ (Game Management Authority: Melbourne, Vic., Australia)
Gelman, A, and Rubin, DB (1992). Inference from iterative simulation using multiple sequences. Statistical Science 7, 457–472.
| Inference from iterative simulation using multiple sequences.Crossref | GoogleScholarGoogle Scholar |
Gentsch, RP, Kjellander, P, and Röken, BO (2018). Cortisol response of wild ungulates to trauma situations: hunting is not necessarily the worst stressor. European Journal of Wildlife Research 64, 11.
| Cortisol response of wild ungulates to trauma situations: hunting is not necessarily the worst stressor.Crossref | GoogleScholarGoogle Scholar |
Godwin, C, Schaefer, JA, Patterson, BR, and Pond, BA (2013). Contribution of dogs to white-tailed deer hunting success. The Journal of Wildlife Management 77, 290–296.
| Contribution of dogs to white-tailed deer hunting success.Crossref | GoogleScholarGoogle Scholar |
Gormley, AM, Forsyth, DM, Griffioen, P, Lindeman, M, Ramsey, DSL, Scroggie, MP, and Woodford, L (2011). Using presence-only and presence–absence data to estimate the current and potential distributions of established invasive species. Journal of Applied Ecology 48, 25–34.
| Using presence-only and presence–absence data to estimate the current and potential distributions of established invasive species.Crossref | GoogleScholarGoogle Scholar |
Græsli, AR, Le Grand, L, Thiel, A, Fuchs, B, Devineau, O, Stenbacka, F, Neumann, W, Ericsson, G, Singh, NJ, Laske, TG, Beumer, LT, Arnemo, JM, Evans, AL, and Cooke, S (2020). Physiological and behavioural responses of moose to hunting with dogs. Conservation Physiology 8, coaa122.
| Physiological and behavioural responses of moose to hunting with dogs.Crossref | GoogleScholarGoogle Scholar |
Hampton, JO, and Hyndman, TH (2019). Underaddressed animal-welfare issues in conservation. Conservation Biology 33, 803–811.
| Underaddressed animal-welfare issues in conservation.Crossref | GoogleScholarGoogle Scholar |
Hampton, JO, MacKenzie, DI, and Forsyth, DM (2019). How many to sample? Statistical guidelines for monitoring animal welfare outcomes. PLoS ONE 14, e0211417.
| How many to sample? Statistical guidelines for monitoring animal welfare outcomes.Crossref | GoogleScholarGoogle Scholar |
Hampton, JO, Bengsen, AJ, Pople, A, Brennan, M, Leeson, M, and Forsyth, DM (2022a). Animal welfare outcomes of helicopter-based shooting of deer in Australia. Wildlife Research 49, 264–273.
| Animal welfare outcomes of helicopter-based shooting of deer in Australia.Crossref | GoogleScholarGoogle Scholar |
Hampton, JO, Bengsen, AJ, Flesch, JS, Toop, SD, Davies, C, Forsyth, DM, Kanstrup, N, Stokke, S, and Arnemo, JM (2022b). A comparison of lead-based and lead-free bullets for shooting of sambar deer (Cervus unicolor) in Australia. Wildlife Research , .
| A comparison of lead-based and lead-free bullets for shooting of sambar deer (Cervus unicolor) in Australia.Crossref | GoogleScholarGoogle Scholar |
Hampton, JO, MacKenzie, DI, and Forsyth, DM (2022c). Animal welfare outcomes of professional vehicle-based shooting of peri-urban rusa deer in Australia. Wildlife Research , .
| Animal welfare outcomes of professional vehicle-based shooting of peri-urban rusa deer in Australia.Crossref | GoogleScholarGoogle Scholar |
Harrison M (2010) ‘Sambar: the magnificent deer.’ (The Australian Deer Research Foundation: Melbourne, Vic., Australia)
Hayward, MW, Jędrzejewski, W, and Jêdrzejewska, B (2012). Prey preferences of the tiger Panthera tigris. Journal of Zoology 286, 221–231.
| Prey preferences of the tiger Panthera tigris.Crossref | GoogleScholarGoogle Scholar |
Kaplan, EL, and Meier, P (1958). Nonparametric estimation from incomplete observations. Journal of the American Statistical Association 53, 457–481.
| Nonparametric estimation from incomplete observations.Crossref | GoogleScholarGoogle Scholar |
Kerr, GN, and Abell, W (2014). Big game hunting in New Zealand: per capita effort, harvest and expenditure in 2011–2012. New Zealand Journal of Zoology 41, 124–138.
| Big game hunting in New Zealand: per capita effort, harvest and expenditure in 2011–2012.Crossref | GoogleScholarGoogle Scholar |
Le Grand, L, Thorsen, NH, Fuchs, B, Evans, AL, Laske, TG, Arnemo, JM, Saebø, S, and Støen, O-G (2019). Behavioral and physiological responses of Scandinavian brown bears (Ursus arctos) to dog hunts and human encounters. Frontiers in Ecology and Evolution 7, 134.
| Behavioral and physiological responses of Scandinavian brown bears (Ursus arctos) to dog hunts and human encounters.Crossref | GoogleScholarGoogle Scholar |
Lebel, F, Dussault, C, Massé, A, and Côté, SD (2012). Influence of habitat features and hunter behavior on white-tailed deer harvest. The Journal of Wildlife Management 76, 1431–1440.
| Influence of habitat features and hunter behavior on white-tailed deer harvest.Crossref | GoogleScholarGoogle Scholar |
Leslie, DM (2011). Rusa unicolor (Artiodactyla: Cervidae). Mammalian Species 43, 1–30.
| Rusa unicolor (Artiodactyla: Cervidae).Crossref | GoogleScholarGoogle Scholar |
Lone, K, Loe, LE, Gobakken, T, Linnell, JDC, Odden, J, Remmen, J, and Mysterud, A (2014). Living and dying in a multi-predator landscape of fear: roe deer are squeezed by contrasting pattern of predation risk imposed by lynx and humans. Oikos 123, 641–651.
| Living and dying in a multi-predator landscape of fear: roe deer are squeezed by contrasting pattern of predation risk imposed by lynx and humans.Crossref | GoogleScholarGoogle Scholar |
Long J (2003) ‘Introduced Mammals of the World.’ (CSIRO Publishing: Melbourne, Vic., Australia)
Macdonald N, Nugent G, Edge K-A, Parkes JP (2019) Eradication of red deer from Secretary Island, New Zealand: changing tactics to achieve success. In ‘Island invasives: scaling up to meet the challenge’. (Eds CR Veitch, MN Clout, AR Martin, JC Russell, CJ West) pp. 256–260. (IUCN: Gland, Switzerland)
Mason E (2008) ‘Secrets of the Sambar.’ (Shikari Press: Bairnsdale, Vic., Australia)
Moloney PD, Hampton JO (2020) ‘Estimates of the 2019 deer harvest in Victoria: results from surveys of Victorian Game Licence holders in 2019.’ (Game Management Authority: Melbourne, Vic., Australia)
Moloney, PD, Gormley, AM, Toop, SD, Flesch, JS, Forsyth, DM, Ramsey, DSL, and Hampton, JO (2022). Bayesian modelling reveals differences in long-term trends in the harvest of native and introduced species by recreational hunters in Australia. Wildlife Research 49, 673–685.
| Bayesian modelling reveals differences in long-term trends in the harvest of native and introduced species by recreational hunters in Australia.Crossref | GoogleScholarGoogle Scholar |
Northrup, JM, Hooten, MB, Anderson, CR, and Wittemyer, G (2013). Practical guidance on characterizing availability in resource selection functions under a use–availability design. Ecology 94, 1456–1463.
| Practical guidance on characterizing availability in resource selection functions under a use–availability design.Crossref | GoogleScholarGoogle Scholar |
Orr, B, Westman, M, Malik, R, and Norris, J (2019). The welfare of pig-hunting dogs in Australia. Animals 9, 853.
| The welfare of pig-hunting dogs in Australia.Crossref | GoogleScholarGoogle Scholar |
Plummer M (2003) JAGS: a program for analysis of Bayesian graphical models using Gibbs sampling. In ‘Proceedings of the 3rd International Workshop on Distributed Statistical Computing (DSC 2003)’, 20–22 March 2003, Vienna, Austria. (Eds K Hornik, F Leisch, A Zeileis) pp. 1–8. (Technische Universität Wien: Vienna, Austria)
Ramesh, T, Sankar, K, Qureshi, Q, and Kalle, R (2012). Group size, sex and age composition of chital (Axis axis) and sambar (Rusa unicolor) in a deciduous habitat of Western Ghats. Mammalian Biology 77, 53–59.
| Group size, sex and age composition of chital (Axis axis) and sambar (Rusa unicolor) in a deciduous habitat of Western Ghats.Crossref | GoogleScholarGoogle Scholar |
R Core Team (2020) ‘R: A language and environment for statistical computing.’ (R Foundation for Statistical Computing: Vienna, Austria). Available at https://www.R-project.org/ [verified 3 September 2021]
Rowland, MM, Nielson, RM, Wisdom, MJ, Johnson, BK, Findholt, S, Clark, D, Didonato, GT, Hafer, JM, and Naylor, BJ (2021). Influence of landscape characteristics on hunter space use and success. The Journal of Wildlife Management 85, 1394–1409.
| Influence of landscape characteristics on hunter space use and success.Crossref | GoogleScholarGoogle Scholar |
Sauerwein, H, Müller, U, Brüssel, H, Lutz, W, and Mstl, E (2004). Establishing baseline values of parameters potentially indicative of chronic stress in red deer (Cervus elaphus) from different habitats in western Germany. European Journal of Wildlife Research 50, 168–172.
| Establishing baseline values of parameters potentially indicative of chronic stress in red deer (Cervus elaphus) from different habitats in western Germany.Crossref | GoogleScholarGoogle Scholar |
Sepúlveda, M, Pelican, K, Cross, P, Eguren, A, and Singer, R (2015). Fine-scale movements of rural free-ranging dogs in conservation areas in the temperate rainforest of the coastal range of southern Chile. Mammalian Biology 80, 290–297.
| Fine-scale movements of rural free-ranging dogs in conservation areas in the temperate rainforest of the coastal range of southern Chile.Crossref | GoogleScholarGoogle Scholar |
Steyaert, SMJG, Zedrosser, A, Elfström, M, Ordiz, A, Leclerc, M, Frank, SC, Kindberg, J, Støen, OG, Brunberg, S, and Swenson, JE (2016). Ecological implications from spatial patterns in human-caused brown bear mortality. Wildlife Biology 22, 144–152.
| Ecological implications from spatial patterns in human-caused brown bear mortality.Crossref | GoogleScholarGoogle Scholar |
Stokke, S, Arnemo, JM, Brainerd, S, Söderberg, A, Kraabøl, M, and Ytrehus, B (2018). Defining animal welfare standards in hunting: body mass determines thresholds for incapacitation time and flight distance. Scientific Reports 8, 13786.
| Defining animal welfare standards in hunting: body mass determines thresholds for incapacitation time and flight distance.Crossref | GoogleScholarGoogle Scholar |
Therneau T (2020) ‘A Package for Survival Analysis in R; R Package Version 3.2-7.’ Availabe at https://cran.r-project.org/web/packages/survival/index.html [verified 17 November 2022]
Vaughan, MR, and Inman, KLH (2002). Hunter effort and success rates of hunting bears with hounds in Virginia. Ursus 13, 223–230.
Watter, K, Thomas, E, White, N, Finch, N, and Murray, P (2020). Reproductive seasonality and rate of increase of wild sambar deer (Rusa unicolor) in a new environment, Victoria, Australia. Animal Reproduction Science 223, 106630.
| Reproductive seasonality and rate of increase of wild sambar deer (Rusa unicolor) in a new environment, Victoria, Australia.Crossref | GoogleScholarGoogle Scholar |