

Catch me if you can: personality drives technique-specific biases during live-capture trapping
Kyla Chloe Johnstone A B C , Clare McArthur A and Peter Bruce Banks A
A B C , Clare McArthur A and Peter Bruce Banks A
A School of Life and Environmental Sciences, The University of Sydney, Sydney, NSW 2006, Australia.
B Manaaki Whenua – Landcare Research, PO Box 69040, Lincoln 7640, New Zealand.
C Corresponding author. Email: johnstonek@landcareresearch.co.nz
Wildlife Research 48(8) 713-721 https://doi.org/10.1071/WR20121
Submitted: 27 July 2020 Accepted: 15 May 2021 Published: 21 June 2021
Journal Compilation © CSIRO 2021 Open Access CC BY-NC-ND
Abstract
Context: Wildlife surveys often rely on a single live-capture technique to sample animal populations. Yet, animal personality (e.g. bold vs shy) can drive heterogeneity in capture probability, leading to biased sampling during live-capture trapping surveys.
Aims: We tested whether a personality-related capture bias is similar between two live-capture techniques, or whether techniques with different capture mechanisms are biased towards certain spectrums of personality.
Methods: We compared two live-capture techniques commonly used for surveying lizards, namely, noosing and pitfall traps. Techniques were deployed several days apart to survey populations of a desert-dwelling agamid, the military dragon, and we used outdoor open-field arenas to test for personality traits relating to boldness, activity and exploration.
Key results: We found that noosing and pitfall traps sampled distinctly different spectrums of personality, with no individuals being captured by both techniques. Unexpectedly, noosing, which involved people approaching dragons to capture them, was biased towards shyer individuals that stayed close to shelter. In contrast, pitfall traps, which were generally set in open areas, were biased towards capturing individuals that moved further from shelter.
Conclusions: We demonstrated that different live-capture techniques can be biased towards different spectrums of personality. We attribute the relationship between personality and technique to an interaction between the capture mechanisms of each technique and individual perceptions of risk and microhabitat use.
Implications: To overcome biased and selective sampling and target individuals along a broad spectrum of personality, surveys should use complementary techniques that vary in their capture mechanisms.
Keywords: applied ecology, behaviour, ethology, mark–recapture, wildlife management.
Introduction
Obtaining representative population data is a core objective of much wildlife management and research. For example, live-capture techniques (i.e. capture and release) are a mainstay of wildlife surveys and are essential for collecting population data, even though only a portion of a population will be sampled (Skalski 1994). Data (e.g. physical measurements, genetic samples and parasite loads) from captured individuals are used to draw inferences about the health and status of the greater population, and provide estimates on other population measures including density (Efford 2004), abundance (Davis et al. 2003), reproduction (Greenville and Dickman 2005), movement (Broome 2001), microhabitat use (Schirmer et al. 2019) and behaviour (Nersesian et al. 2011). There exists a long-standing drive to develop ‘best-practice’ approaches for wildlife surveys by using the most effective sampling technique (e.g. Norbury et al. 2002), namely, one that maximises capture rates, reduces survey costs and effort (Garden et al. 2007) and accounts for ethical considerations. Although the best practice approach is often the technique that outperforms others (i.e. higher capture rates; O’Farrell et al. 1994; Rolfe and McKenzie 2000), different live-capture techniques can sample different groups of individuals in a population (Beacham and Krebs 1980; Harrison et al. 2002; Michael et al. 2012). This outcome suggests that despite best-practice approaches, capture probability may vary among individuals, leading to biased population samples.
Most live-capture techniques have some degree of bias (e.g. Herremans 1989; Rodda et al. 2007), often in relation to where a technique can be deployed. For example, small-mammal capture rates often increase with vegetation density (Cox et al. 2000), seine nets are effective for surveying fish in shallow (wade-able) open water but are difficult to manipulate in deeper water or along riverbanks (Pierce et al. 1990), and pitfall traps, which are popular for sampling small mammals and reptiles (Bury and Corn 1987; Moseby and Read 2001), must be buried and cannot be deployed in rocky areas or in dense vegetation. Although each technique may be effective at targeting the ‘average’ animal in a population, individuals vary in microhabitat use (Montiglio et al. 2013; Schirmer et al. 2019) and devices that target only part of a species’ habitat are unlikely to provide representative samples.
Individual behaviour can also strongly influence susceptibility to capture. Animal movement (Read and Moseby 2001), demographics (Kay et al. 2000; Rodda et al. 2007), state-dependent factors (Laves and Loeb 2006; Bisi et al. 2011) and individual personality (Biro and Dingemanse 2009) can influence encounter and capture rates. Live-capture techniques rely on animal decision-making for success (Garvey et al. 2021); however, if techniques are perceived to present danger or risk, capture success may vary. For example, entering a trap can be associated with a degree of risk by animals (Stokes 2013; K. Johnstone, C. McArthur, and P. Banks, unpubl. data) and risk sensitive individuals may actively avoid capture. Active search techniques, such as noosing and hand-catching, which search for and target individuals also likely to present a degree of danger (Cooper Jr 2003), and are likely to select for risk-taking individuals with short flight-initiation distances (the distance at which an animal flees an approaching threat, as discussed by Carter et al. 2012). In contrast, techniques that are less reliant on decision-making by animals may have a lower chance for biased sampling. Pitfall traps, for example, are a passive technique (i.e. set and left in the environment) and may randomly capture animals as they move through the landscape (NSW Department of Primary Industries 2020), while netting (seine nets for fish, cannon nets for birds, or flushing mammals towards nets) is assumed to target all animals in the immediate area (e.g. Noss 1999; Walsh et al. 2002).
Variation in risk-taking among individuals is a common component of animal personality (consistent among-individual differences in behaviour; Réale et al. 2007). Personality traits, including boldness, activity, exploration and aggression, can influence individual fitness (see table 1 in Biro and Stamps 2008) and are often associated with risk-taking behaviours. During population sampling, risk-taking individuals are often trapped more easily and more frequently than risk-averse individuals (Biro and Post 2008; Boon et al. 2008 Garamszegi et al. 2009; Wilson et al. 2011; Carter et al. 2012). However, a personality-related bias is not always an inevitable outcome of population sampling, such as, for instance, if a technique targets animals indiscriminately (Biro and Dingemanse 2009) or does not present as a novel situation (Michelangeli et al. 2016).
Personality-related sampling biases are typically studied using a single method of live-capture trapping (Biro and Post 2008; Boon et al. 2008; Garamszegi et al. 2009; Carter et al. 2012). In examining the interaction between personality and capture technique, one study (Wilson et al. 2011) demonstrated that different techniques (angling and seine nets) captured different spectrums of personality in bluegill sunfish (Lepomis macrochirus). However, another study (Michelangeli et al. 2016) found no relationship between skink (Lampropholis delicata) personality and capture technique (mealworm fishing, hand-capture or pitfall trap). Importantly, both studies used techniques concurrently, and so, individuals captured by one technique were not able to be captured by another. Thus, it remains unclear whether distinctive personality-related biases may arise among techniques as a result of the interaction between individual personality and the different mechanisms of live-capture techniques.
To test whether different techniques sample different spectrums of personality, we used two live-capture techniques deployed several days apart and compared the personality of captured individuals. We used noosing and pitfall traps, two common techniques for surveying lizards (García-Muñoz and Sillero 2010; Greenville et al. 2016). Both techniques have limitations and may select for certain individuals. Noosing, like other active techniques, searches for and targets detected individuals, but may select for risk-taking individuals with short flight-initiation distances (Ward‐Fear et al. 2019). In contrast, pitfall trapping is a passive technique that is assumed to present little to no risk, and should have a lower potential for biased sampling (Bury and Corn 1987). However, pitfall traps may be limited to open exposed microhabitats where only some individuals may be active.
We focussed on the military dragon (Ctenophorus isolepis), a conspicuous lizard common in arid areas of Australia. The military dragon (hereafter dragon) is a small (snout–vent length = 75 mm) terrestrial agamid with an annual lifecycle (Dickman et al. 1999). From observations, a typical home-range size is ~20 m2 (C. Dickman, pers. comm.) and population density correlates with spinifex grass (Triodia basedowii) density (Dickman et al. 1999). In open, exposed microhabitats, dragons are at an increased risk of predation (Daly et al. 2008), mostly by raptors and corvids (Pianka 1971; Daly et al. 2008). The Military dragon has been used to investigate lizard thermoregulation and antipredator responses (Losos 1987), microhabitat selection (Downey and Dickman 1993), physiology (Thompson and Withers 2005) and the influence of rainfall (Dickman et al. 1999) and fire (Masters 1996) on population dynamics. If personality influences responses to the perceived risks associated with different live-capture techniques, we predicted that noosing would capture a narrow spectrum of bolder individuals that allow surveyors to approach within noosing distance, whereas pitfall traps will capture a broader personality spectrum through random sampling.
Materials and methods
Study area
We worked on Ethabuka Station in the Simpson Desert, south-western Queensland (23°46′S, 138°28′E) in September 2015. Parallel sand dunes up to 10 m high are separated by swales spanning from 0.6 km to 1 km (Purdie 1984). Vegetation in the swales is dominated by spinifex grass (>50% ground cover), although vegetation diversity increases slightly from the dune swale to crest (Greenville and Dickman 2005). During our study, mean vegetation cover at sites (measured following Glen et al. 2010) was 20–50%, a density that falls within the habitat requirements to support a stable dragon population (Dickman et al. 1999). Hourly air temperatures ranged from 6°C to 36°C and no rain fell, although local and regional rains can occur year-round (Greenville and Dickman 2005).
Experimental design
We sampled dragon populations at five 1 ha sites that encompassed pre-existing pitfall-trap grids established for long-term research (Downey and Dickman 1993). Pitfall traps are closed between surveys, but the trapping infrastructure (traps with lids and mesh fencing) remain in situ year-round. Sites were separated by 500 m; 25 times greater than the expected home-range length of a dragon (C. Dickman, pers. comm). To expose all animals to both live-capture techniques, we used noosing and pitfall traps several days apart. To minimise avoidance behaviours by dragons, we conducted noosing first, because noosing requires the handler to approach within noosing distance. We considered it unlikely that the noosing or handling experience would affect pitfall-trap captures, because, in captures, pitfall traps are largely random and incidental (NSW Department of Primary Industries 2020). Although post-capture behaviour may have been influenced by capture technique, there is no obvious reason why this would occur, because (i) animals were left undisturbed for 24 h following capture (except to provide food) and (ii) any influence of capture technique is likely to be far-outweighed by the influence of handling, which can have a considerable impact on animal behaviour (Prout and King 2006). We kept handling to a minimum and handled all animals similarly.
Surveying populations
First, sites were surveyed by noosing between 0900 hours and 1600 hours for three consecutive days (~2.5 h per site, per day). Two surveyors separated by 20 m walked parallel 1200-m serpentine transects from the dune swale to dune crest and back (Fig. 1). This method ensured that the entire site was surveyed, including the area around each (closed) pitfall trap. Using a fishing rod and noose (made from fishing line or dental floss), surveyors attempted to catch any dragon sighted within a 10 m radius. Because lizards are more likely to flee when approached quickly and directly (Cooper Jr 2009), surveyors approached slowly, either side-on or crouching. Encounters where dragon escaped unmarked were also recorded.

|
Captured dragons were uniquely marked for temporary identification by using a black marker that persisted for the duration of our study. We also recorded the weight, sex, snout–vent length, and breeding condition of individuals. Dragons with a snout–vent length of >55 mm were considered to be adults, and all others, to be juveniles (Dickman et al. 1999). We housed dragons (3–5 nights) individually in aquariums with a sand substrate and plastic hide. Water was provided ad libitum and food (mealworms) was provided twice daily. After testing for potential personality traits (described below) we released all dragons at their point of capture. All dragons were released 18–48 h before the pitfall traps were opened.
Pitfall traps were open for five nights (one site was open three nights), checked each morning at sunrise and left open during the day when dragons are active. Traps consisted of a PVC pipe (16 cm diameter × 60 cm deep) buried flush to the ground with aluminium fly-wire mesh fixed to the base to prevent animals from digging to escape. Mesh drift fence (5 m long, 30 cm high) ran either side of each trap to increase trap success (Friend et al. 1989). Pitfall traps were arranged in a 6 × 6 grid of 36 traps (Fig. 1), spanning 100 m from dune swale to dune crest (as described in Dickman et al. 1999). Traps were separated by 20 m, exposing all resident dragons within the pitfall trap grid to at least one trap. Dragons caught by pitfall traps underwent the same protocols as noosed dragons.
Testing for personality traits
We tested personality traits under semi-wild conditions using an open-field test (with repeated measures, n = 3). The open-field test is a standard method for measuring traits including boldness, activity and exploration (Walsh and Cummin 1976; Carter et al. 2013). We used 12 outdoor yards (5 m × 5 m) made of mesh wire (60 cm high) covered with plastic (buried to 10 cm, to prevent escape). The sand substrate was smoothed before each test, so that dragon tracks could be mapped to record movement. Dragons were tested once a day between 0900 hours and 1700 hours over three or four consecutive days. To reduce the potential influence of time, temperature and habituation on behaviour, dragons were assigned a random order and random yard for testing.
Before each test, a dragon was placed in partial sun for 15 min for thermoregulation, then released into a plastic hide (15 cm × 15 cm × 9 cm) in a corner of the yard and left to roam undisturbed for 60 min. We then recaptured the dragon and mapped the tracks. The wind made some tracks difficult to record, so we used a reliability scoring system of 0–2: ‘0’ = unreliable, missing tracks; ‘1’ = mostly reliable, some missing tracks; and ‘2’ = reliable, all tracks clear. Tests with a score of ‘0’ (n = 5) were excluded from analyses. One dragon had two tests excluded, reducing our sample of individuals with repeated tests from 31 to 30. Hourly air temperatures were downloaded from the local Ethabuka camp weather station.
Quantifying behaviours
To quantify behaviours from the open-field test, yards were divided into 16 grids (12 perimeter grids, 4 interior grids). Thigmotaxis is prevalent in species, including dragons, where predation risk increases in exposed areas (Choleris et al. 2001; Daly et al. 2008; Carlson and Langkilde 2013). Thigmotaxis is associated with the bold–shy trait (Burns 2008). Therefore, we considered that dragons perceived the perimeter grids near the tarp to be safer, sheltered areas, and the interior grids furthest from the tarp to be riskier, exposed area; hence, the use of the interior grids as a measure of boldness. We calculated the following three behavioural metrics: index of exposed grids entered, number of grids entered (of a maximum of 16) and cumulative movements between grids.
Statistical analysis
Quantifying personality traits
Because there was a difference in hourly air temperatures between the open-field tests of animals caught by noosing and those of animals caught with pitfall traps (mean hourly air temperature: 22.9°C, 29.7°C respectively; d.f. = 1, χ2 = 13.47, P = 0.0002), we incorporated hourly air temperature into our model when testing for personality traits. Quantified behaviours from the open-field test were analysed separately as potential personality traits using the GLIMMIX procedure in SAS (Dingemanse and Dochtermann 2013). The Shapiro–Wilk test confirmed that distributions were normal. We compared two reduced models; Model 1 had significant fixed-effects only (test order and hourly air temperature, the latter accounting for temperature during each open-field test for each animal), whereas Model 2 also included individual identity as a random factor (following Dingemanse and Dochtermann 2013). A significant difference in the –2 log-likelihood (–2 LL) between the models indicated that a behaviour was a significant personality trait. As a secondary method of validation, we used the Akaike information criterion (AIC), and a difference of >2 between the two models indicated an improvement in explanatory power (Burnham and Anderson 2002). We measured trait repeatability (following Dingemanse and Dochtermann 2013) and tested for trait plasticity by comparing Model 2 with Model 3, which included the interaction between individual identity and test order as a random factor. We calculated trait indices using a fourth model that included test order and hourly air temperature from the open-field tests, again to account for any effects of ambient air temperature on the behaviour of this ectotherm during testing.
Testing for potential sampling biases
Each individual was caught either by noosing or in pitfall traps (i.e. never by both methods, see Results). Therefore, we ran several generalised linear models (JMP Pro 13) with binomial distributions to test the probability of capture by noosing (rather than in pitfall traps) as a function of individual personality, sex and age, and we included max temperature on day of capture in all models to account for any influence of differences in temperature when each technique was used. Low captures in pitfall traps meant that we could not include interactions among all main effects in a single model. So, we ran three models testing two-way interactions between personality, sex and/or age before reducing to a final best model of significant effect(s) only. We included the Firth adjustment to decrease bias and standard error.
Results
Across the five sites, we conducted ~38 search hours during noosing and 828 trap-nights for pitfall traps. We captured 31 individual dragons, including 23 by noosing and eight in pitfall traps. Of these, 17 were male (4 adults, 13 juveniles), 12 were female (5 adults, 7 juveniles) and two were juveniles of indeterminate sex. No individual was captured by both techniques, even though all individuals would have been exposed to both noosing and pitfall traps. During noosing, we recorded 17 encounters (of a total 40 encounters) in which a dragon escaped capture, unmarked, although these animals may have been caught later. It was unknown how often dragons encountered, but avoided, pitfall traps.
The 30 dragons with quantified behaviours displayed significant and repeatable differences in the index of exposed grids entered (significant personality trait, P = 0.02, r = 0.28; Table 1) accounting for the significant positive influence of hourly air temperature on index of exposed grids entered during the open-field tests (F1,48 = 29.28, P < 0.0001). No other behaviours were significant personality traits (Table 1).

|
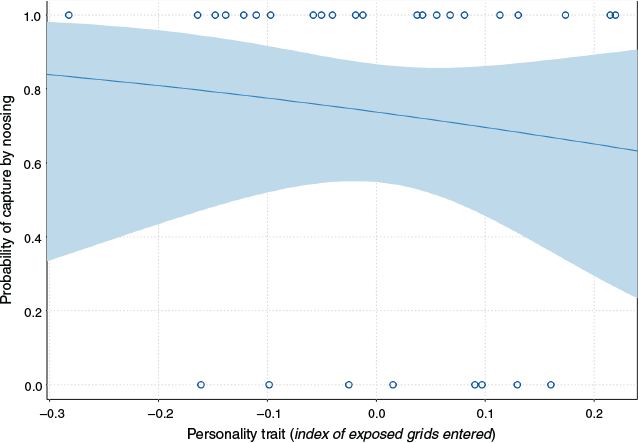
The probability of capture by noosing was significantly influenced by personality (index of exposed grids entered; likelihood-ratio Chi-squared test ((LRχ2) = 8.23, P = 0.004; Fig. 2) and by temperature (max temperature on day of capture; LRχ2 = 34.43, P < 0.0001). Shyer animals (lower index of exposed grids entered) were more likely to be noosed than were bolder animals (higher index of exposed grids entered), and vice versa for captures in pitfall traps. The relationship between probability of capturing animals by noosing and temperature was negative, reflecting the cooler temperature during noosing surveys than during pitfall trapping. Capture probability was not significantly (P ≥ 0.99) affected by sex, age or any interaction.
|
We used the minimum number known alive (MNKA; Krebs 1966) to estimate population size at each of the five sites by using each technique. Overall, pitfall traps yielded a lower total and mean population estimate with a smaller variance (total MNKA across sites, n = 8, mean 1.6 ± variance 1.8) than did noosing (total MNKA across sites, n = 22, mean 4.6 ± variance 6.3), although both techniques yielded lower population estimates than when combined (total MNKA across sites, n = 31, mean 6.2 ± variance 13.2).
Discussion
We found that a personality-related sampling bias varied between the two live-capture techniques. All sites were surveyed using both techniques and all individuals were available for capture by noosing and by pitfall traps. However, noosing captured what we consider to be shyer individuals (lower index of exposed grids entered) and pitfall traps captured what we consider to be bolder individuals (higher index of exposed grids entered). Our findings showed that a personality bias can occur when using active and passive live-capture techniques. But the nature of the bias depends on the technique.
In contrast to our prediction, pitfall traps did not sample a broader personality spectrum than did noosing. If pitfall traps sampled individuals randomly, we would expect a broader personality spectrum than with noosing. Although captures were low in pitfall traps, individuals were bolder-type animals (Fig. 2). A personality-related capture bias has been demonstrated using baited traps that rely on lures or attractant (e.g. Boon et al. 2008; Carter et al. 2012). But this is the first demonstration of a personality bias associated with an active search technique (noosing) or with pitfall traps, and the relationship between capture technique and personality in reptiles would benefit from further investigation.
We suggest that the sampling bias associated with the live-capture techniques resulted from an interaction among technique, personality and microhabitat. Owing to the presence of humans, noosing is usually associated with a high degree of perceived risk for animals and should select for bolder individuals with short flight-initiation distances (Carter et al. 2012); however, we found the opposite to be the case, assuming that a lower index of exposed grids entered represented shyer individuals. It is possible that we interpreted the direction of the bold–shy axes the wrong way around. However, we consider this to be unlikely (as justified in the methods) and, irrespective of the of the direction of the personality bias, personality drove differences in capture probability by each technique. Proximity to vegetation (i.e. shelter) reduces real and perceived predation risk (Cooper Jr and Whiting 2007; Daly et al. 2008) and can reduce flight-initiation distance in lizards (Cooper Jr and Whiting 2007). Shyer individuals may therefore have been approached and noosed more easily than were bolder individuals, simply because of the personality-related microhabitat preferences, with shy animals sheltering adjacent to, or under, spinifex, and bold (as our personality trials showed) animals making greater use of exposed and open microhabitat. This interpretation is also consistent with the pitfall-trap results. The area surrounding pitfall traps (within a 2.5 m radius) during our study was predominantly (>75%; Drew 2016) bare ground, and our pitfall traps were more likely to capture bolder individuals. Interestingly, vegetation cover around pitfall traps has had negligible effects on captures of other terrestrial reptiles (Craig et al. 2009), although the authors acknowledged their results were likely to have been influenced by low sample sizes and limited knowledge on species-specific habitat preferences (Craig et al. 2009), and they did not consider personality. Our results suggest that targeting different spectrums of personality during surveys can greatly increase captures and provide details about species behaviour that may otherwise go overlooked.
We could not quantify the personality of individuals that evaded capture during noosing or that might have avoided capture by pitfall traps. It is possible that, due to observer bias, during noosing we missed individuals that remained motionless. For example, indigenous rangers are more skilled at spotting varanids sheltering under vegetation or in the shade than are western ecologists (Ward‐Fear et al. 2019). We detected most dragons through movement (foraging, moving between shade and sun), but several motionless dragons were also detected and captured. A pilot trial tested for surveyor bias and showed it was unlikely that a significant portion of motionless individuals was missed. Surveying grids using parallel transects also increased the opportunity for detecting dragons. Because any bias against motionless individuals (i.e. the shyest that remained motionless or under vegetation) went undetected in our study, we are likely to be presenting a conservative estimate of capture bias. Targeting the untrappable individuals presents a major challenge for wildlife research and management, particularly for wildlife conservation (Garvey et al. 2021).
Both noosing and pitfall traps underestimated the total known population abundance. Unexpectedly, pitfall traps, which have often been considered more effective than are other techniques (Williams and Braun 1983), appeared to miss a significant proportion of the dragon population, in terms of both personality and numbers, and provided the lowest abundance estimate (MNKA). Given these results, previous estimates relying solely on pitfall traps may underestimate the abundance of dragons, and other species. Differences in capture efficacy could be attributed to the mechanistic differences between noosing and pitfall traps. Noosing and other active live-capture techniques actively target animals and can have greater encounter probabilities than do passive techniques (such as pitfall traps) that mostly rely on random encounters (Michael et al. 2012). The presence of drift fences helps improve pitfall-trap captures (Moseby and Read 2001). However, individuals that perceive drift fences as novel, may avoid them out of neophobic (avoidance of novelty) responses. Neophobia was unlikely in our study because the whole pitfall-trap system (including drift fences) has remained in the field year-round for >20 years. Instead, some dragons may have learnt to circumvent the drift fence and trap, and long-term studies using permanent drift fences may inadvertently lower capture rates over time. Fences at some pitfall traps were not fully taunt and straight, and some dragons may also have avoided capture by climbing the fences rather than following them.
We considered three additional but unlikely factors that may have influenced our results. First, the noosing and handling experience may have influenced later responses to pitfall traps and some species may be able to detect the pitfall trap ‘pit’ and avoid it (e.g. Enge 2001). However, this behaviour has not been recorded in lizards. Our pitfall-trap success (1%) was comparable to that (1.2%) of a long-term (13–22 years) study in the area (Greenville et al. 2016) conducted across all seasons. Therefore, noosing before pitfall traps is likely to have had little impact on our results.
Second, ambient air temperature during our study was below the species’ optimal range (Melville and Schulte 2001), and particularly so, during the noosing survey. Temperature may have affected our results in two ways, namely, by varying (1) when dragons were caught by noosing versus in pitfall traps (lower in the former), and (2) among trials while testing for personality in the yards. By accounting for these effects statistically, we showed that despite any influence of temperature, personality remains a significant factor that can influence capture probability by different techniques.
More generally, cooler temperatures (minimum 8°C, mean 21°C, maximum 36°C) may have limited pitfall-trap efficacy. In the arid zone, reptile-capture success in pitfall traps often increases with temperature (Read and Moseby 2001). For Ctenophorus spp., captures increase when daily minimum and maximum temperatures exceed 24°C and 34°C respectively (Read and Moseby 2001). Because our pitfall-trap success was comparable to previous work in the area conducted across a gradient of temperatures (Greenville et al. 2016), cooler weather did not unduly influence captures.
The specific direction in the relationship between the personality trait and the impact on capture probability by one or the other technique shows that the direction of a personality bias is not easy to predict. In sampling different spectrums of personality by using an active (noosing) and a passive (pitfall traps) technique, we showed that a personality-related sample bias is not just an issue for live-capture traps (e.g. cage traps), but applies broadly to other survey techniques. As any interaction between personality and technique can generate a bias, the use of a single technique, even those assumed to have little bias, is unlikely to provide representative population data.
Noosing might be considered a best-practice approach because it caught nearly three times more dragons than did pitfall traps. However, it did not catch the bolder individuals. Without pitfall traps, we could not have detected behavioural differences among the dragons, nor would there have been a detectable personality bias. Our results therefore highlighted a considerable challenge for researchers selecting or relying on ‘best-practice’ methods and the need to account for animal personality during surveys. To overcome a personality-related bias, we have provided additional evidence in support the argument presented by (Biro and Dingemanse 2009), namely that deploying complementary live-capture techniques, i.e. ones that target different personality profiles, will provide more robust and representative population samples. Similar approaches are used in mammal pest control to target animals that avoid the initial technique (Liang et al. 2013). Using complementary techniques concurrently can certainly be incorporated into most ecological surveys to overcome sample biases and is an important strategy to consider when addressing similar biases in wildlife management operations (see review by (Merrick and Koprowski 2017). To reduce the potential for a personality-related bias during surveys, techniques should differ in their capture mechanisms, operate across different microhabitats, different temperature gradients and vary in the degree of risk they present to the target species.
Data accessibility statement
Analyses reported in this article can be reproduced using the data provided in figshare.com (doi:10.6084/m9.figshare.12451406).
Conflicts of interest
The authors declare no conflicts of interest.
Declaration of funding
This work was supported by the Australian Research Council (DP 140104621 to C.D) and the Ecological Consultants Association of NSW.
Acknowledgements
Thanks go to Chris Dickman and the Desert Ecology Research Group for advice, assistance and access to the sites. A heartfelt thanks go to volunteers David Martin, Clare Bowry and Stephen Greig for their dedicated fieldwork assistance. All research was conducted with Animal Ethics (2014/423) and Scientific Licence (WISP15192414, WITK15192514) approval. All authors gave final approval for publication and the authors declare no conflict of interest. K. C. J. and P. B. B. conceptualised and designed the research, K. C. J. conducted the field research and collected data. All authors contributed to the statistical analysis. K. C. J. wrote the manuscript with critical contributions from P. B. B. and C. M.
References
Beacham, T. D., and Krebs, C. J. (1980). Pitfall versus live-trap enumeration of fluctuating populations of Microtus townsendii. Journal of Mammalogy 61, 486–499.| Pitfall versus live-trap enumeration of fluctuating populations of Microtus townsendii.Crossref | GoogleScholarGoogle Scholar |
Biro, P. A., and Dingemanse, N. J. (2009). Sampling bias resulting from animal personality. Trends in Ecology & Evolution 24, 66–67.
| Sampling bias resulting from animal personality.Crossref | GoogleScholarGoogle Scholar |
Biro, P. A., and Post, J. R. (2008). Rapid depletion of genotypes with fast growth and bold personality traits from harvested fish populations. Proceedings of the National Academy of Sciences of the United States of America 105, 2919–2922.
| Rapid depletion of genotypes with fast growth and bold personality traits from harvested fish populations.Crossref | GoogleScholarGoogle Scholar | 18299567PubMed |
Biro, P. A., and Stamps, J. A. (2008). Are animal personality traits linked to life-history productivity? Trends in Ecology & Evolution 23, 361–368.
| Are animal personality traits linked to life-history productivity?Crossref | GoogleScholarGoogle Scholar |
Bisi, F., Newey, S., Nodari, M., Wauters, L. A., Harrison, A., Thirgood, S., and Martinoli, A. (2011). The strong and the hungry: bias in capture methods for mountain hares Lepus timidus. Wildlife Biology 17, 311–316.
| The strong and the hungry: bias in capture methods for mountain hares Lepus timidus.Crossref | GoogleScholarGoogle Scholar |
Boon, A. K., Réale, D., and Boutin, S. J. O. (2008). Personality, habitat use, and their consequences for survival in North American red squirrels Tamiasciurus hudsonicus. Oikos 117, 1321–1328.
| Personality, habitat use, and their consequences for survival in North American red squirrels Tamiasciurus hudsonicus.Crossref | GoogleScholarGoogle Scholar |
Broome, L. S. (2001). Density, home range, seasonal movements and habitat use of the mountain pygmy‐possum Burramys parvus (Marsupialia: Burramyidae) at Mount Blue Cow, Kosciuszko National Park. Austral Ecology 26, 275–292.
| Density, home range, seasonal movements and habitat use of the mountain pygmy‐possum Burramys parvus (Marsupialia: Burramyidae) at Mount Blue Cow, Kosciuszko National Park.Crossref | GoogleScholarGoogle Scholar |
Burnham, K., and Anderson, D. (2002). ‘Model selection and multimodel inference: a practical information-theoretic approach.’ 2nd edn. (Springer-Verlag: New York, NY, USA.)
Burns, J. G. (2008). The validity of three tests of temperament in guppies (Poecilia reticulata). Journal of Comparative Psychology 122, 344.
| The validity of three tests of temperament in guppies (Poecilia reticulata).Crossref | GoogleScholarGoogle Scholar | 19014258PubMed |
Bury, R. B., and Corn, P. S. (1987). Evaluation of pitfall trapping in northwestern forests: trap arrays with drift fences. The Journal of Wildlife Management 51, 112–119.
| Evaluation of pitfall trapping in northwestern forests: trap arrays with drift fences.Crossref | GoogleScholarGoogle Scholar |
Carlson, B. E., and Langkilde, T. (2013). Personality traits are expressed in bullfrog tadpoles during open-field trials. Journal of Herpetology 47, 378–383.
| Personality traits are expressed in bullfrog tadpoles during open-field trials.Crossref | GoogleScholarGoogle Scholar |
Carter, A. J., Heinsohn, R., Goldizen, A. W., and Biro, P. A. (2012). Boldness, trappability and sampling bias in wild lizards. Animal Behaviour 83, 1051–1058.
| Boldness, trappability and sampling bias in wild lizards.Crossref | GoogleScholarGoogle Scholar |
Carter, A. J., Feeney, W. E., Marshall, H. H., Cowlishaw, G., and Heinsohn, R. (2013). Animal personality: what are behavioural ecologists measuring? Biological Reviews of the Cambridge Philosophical Society 88, 465–475.
| Animal personality: what are behavioural ecologists measuring?Crossref | GoogleScholarGoogle Scholar | 23253069PubMed |
Choleris, E., Thomas, A., Kavaliers, M., and Prato, F. (2001). A detailed ethological analysis of the mouse open field test: effects of diazepam, chlordiazepoxide and an extremely low frequency pulsed magnetic field. Neuroscience and Biobehavioral Reviews 25, 235–260.
| A detailed ethological analysis of the mouse open field test: effects of diazepam, chlordiazepoxide and an extremely low frequency pulsed magnetic field.Crossref | GoogleScholarGoogle Scholar | 11378179PubMed |
Cooper Jr, W. (2003). Risk factors affecting escape behavior by the desert iguana, Dipsosaurus dorsalis: speed and directness of predator approach, degree of cover, direction of turning by a predator, and temperature. Canadian Journal of Zoology 81, 979–984.
| Risk factors affecting escape behavior by the desert iguana, Dipsosaurus dorsalis: speed and directness of predator approach, degree of cover, direction of turning by a predator, and temperature.Crossref | GoogleScholarGoogle Scholar |
Cooper Jr, W. E. (2009). Optimal escape theory predicts escape behaviors beyond flight initiation distance: risk assessment and escape by striped plateau lizards Sceloporus virgatus. Current Zoology 55, 123–131.
| Optimal escape theory predicts escape behaviors beyond flight initiation distance: risk assessment and escape by striped plateau lizards Sceloporus virgatus.Crossref | GoogleScholarGoogle Scholar |
Cooper Jr, W. E., and Whiting, M. J. (2007). Universal optimization of flight initiation distance and habitat‐driven variation in escape tactics in a Namibian lizard assemblage. Ethology 113, 661–672.
| Universal optimization of flight initiation distance and habitat‐driven variation in escape tactics in a Namibian lizard assemblage.Crossref | GoogleScholarGoogle Scholar |
Cox, M. P., Dickman, C. R., and Cox, W. G. (2000). Use of habitat by the black rat (Rattus rattus) at North Head, New South Wales: an observational and experimental study. Austral Ecology 25, 375–385.
| Use of habitat by the black rat (Rattus rattus) at North Head, New South Wales: an observational and experimental study.Crossref | GoogleScholarGoogle Scholar |
Craig, M. D., Grigg, A. H., Garkaklis, M. J., Hobbs, R. J., Grant, C. D., Fleming, P. A., and Hardy, G. E. S. J. (2009). Does habitat structure influence capture probabilities? A study of reptiles in a eucalypt forest. Wildlife Research 36, 509–515.
| Does habitat structure influence capture probabilities? A study of reptiles in a eucalypt forest.Crossref | GoogleScholarGoogle Scholar |
Daly, B. G., Dickman, C. R., and Crowther, M. S. (2008). Causes of habitat divergence in two species of agamid lizards in arid central Australia. Ecology 89, 65–76.
| Causes of habitat divergence in two species of agamid lizards in arid central Australia.Crossref | GoogleScholarGoogle Scholar | 18376548PubMed |
Davis, S. A., Akison, L. K., Farroway, L. N., Singleton, G. R., and Leslie, K. E. (2003). Abundance estimators and truth: accounting for individual heterogeneity in wild house mice. The Journal of Wildlife Management 67, 634–645.
| Abundance estimators and truth: accounting for individual heterogeneity in wild house mice.Crossref | GoogleScholarGoogle Scholar |
Dickman, C. R., Letnic, M., and Mahon, P. S. (1999). Population dynamics of two species of dragon lizards in arid Australia: the effects of rainfall. Oecologia 119, 357–366.
| Population dynamics of two species of dragon lizards in arid Australia: the effects of rainfall.Crossref | GoogleScholarGoogle Scholar | 28307758PubMed |
Dingemanse, N. J., and Dochtermann, N. A. (2013). Quantifying individual variation in behaviour: mixed-effect modelling approaches. Journal of Animal Ecology 82, 39–54.
| Quantifying individual variation in behaviour: mixed-effect modelling approaches.Crossref | GoogleScholarGoogle Scholar |
Downey, F. J., and Dickman, C. R. (1993). Macro-and microhabitat relationships among lizards of sandridge desert in central Australia. In ‘Herpetology in Australia: a Diverse Discipline’. (Eds D. Lunney, and D. Ayers.) pp. 133–138.
Drew, G. (2016). Divergent habitat selection in two small marsupials in the Simpson Desert, Australia. B.Sc.(Hons) Thesis, The University of Sydney, NSW, Australia.
Efford, M. (2004). Density estimation in live‐trapping studies. Oikos 106, 598–610.
| Density estimation in live‐trapping studies.Crossref | GoogleScholarGoogle Scholar |
Enge, K. M. (2001). The pitfalls of pitfall traps. Journal of Herpetology 35, 467–478.
| The pitfalls of pitfall traps.Crossref | GoogleScholarGoogle Scholar |
Friend, G., Smith, G. T., Mitchell, D., and Dickman, C. (1989). Influence of pitfall and drift fence design on capture rates of small vertebrates in semi-arid habitats of Western-Australia. Australian Wildlife Research 16, 1–10.
| Influence of pitfall and drift fence design on capture rates of small vertebrates in semi-arid habitats of Western-Australia.Crossref | GoogleScholarGoogle Scholar |
Garamszegi, L. Z., Eens, M., and Török, J. (2009). Behavioural syndromes and trappability in free-living collared flycatchers, Ficedula albicollis. Animal Behaviour 77, 803–812.
| Behavioural syndromes and trappability in free-living collared flycatchers, Ficedula albicollis.Crossref | GoogleScholarGoogle Scholar |
García-Muñoz, E., and Sillero, N. (2010). Two new types of noose for capturing herps. Acta Herpetologica 5, 259–264.
Garden, J. G., McAlpine, C. A., Possingham, H. P., and Jones, D. N. (2007). Using multiple survey methods to detect terrestrial reptiles and mammals: what are the most successful and cost-efficient combinations? Wildlife Research 34, 218–227.
| Using multiple survey methods to detect terrestrial reptiles and mammals: what are the most successful and cost-efficient combinations?Crossref | GoogleScholarGoogle Scholar |
Garvey, P. M., Banks, P. B., Suraci, J. P., Bodey, T. W., Glen, A. S., Jones, C. J., McArthur, C., Norbury, G. L., Price, C. J., Russell, J. C., and Sih, A. (2021). Leveraging motivations, personality, and sensory cues for vertebrate pest management. Trends in Ecology & Evolution 35, 990–1000.
Glen, A. S., Sutherland, D. R., and Cruz, J. (2010). An improved method of microhabitat assessment relevant to predation risk. Ecological Research 25, 311–314.
| An improved method of microhabitat assessment relevant to predation risk.Crossref | GoogleScholarGoogle Scholar |
Greenville, A. C., and Dickman, C. R. (2005). The ecology of Lerista labialis (Scincidae) in the Simpson Desert: reproduction and diet. Journal of Arid Environments 60, 611–625.
| The ecology of Lerista labialis (Scincidae) in the Simpson Desert: reproduction and diet.Crossref | GoogleScholarGoogle Scholar |
Greenville, A. C., Wardle, G. M., Nguyen, V., and Dickman, C. R. (2016). Spatial and temporal synchrony in reptile population dynamics in variable environments. Oecologia 182, 475–485.
| Spatial and temporal synchrony in reptile population dynamics in variable environments.Crossref | GoogleScholarGoogle Scholar | 27337964PubMed |
Harrison, R. L., Barr, D. J., and Dragoo, J. W. (2002). A Comparison of Population Survey Techniques for Swift Foxes (Vulpes velox) in New Mexico. American Midland Naturalist 148, 320–337.
| A Comparison of Population Survey Techniques for Swift Foxes (Vulpes velox) in New Mexico.Crossref | GoogleScholarGoogle Scholar |
Herremans, M. (1989). Habitat and sampling related bias in sex-ratio of trapped blackcaps Sylvia atricapilla. Ringing & Migration 10, 31–34.
| Habitat and sampling related bias in sex-ratio of trapped blackcaps Sylvia atricapilla.Crossref | GoogleScholarGoogle Scholar |
Kay, B., Gifford, E., Perry, R., and van de Ven, R. (2000). Trapping efficiency for foxes (Vulpes vulpes) in central New SouthWales: age and sex biases and the effects of reduced fox abundance. Wildlife Research 27, 547–552.
| Trapping efficiency for foxes (Vulpes vulpes) in central New SouthWales: age and sex biases and the effects of reduced fox abundance.Crossref | GoogleScholarGoogle Scholar |
Krebs, C. J. (1966). Demographic changes in fluctuating populations of Microtus californicus. Ecological Monographs 36, 239–273.
| Demographic changes in fluctuating populations of Microtus californicus.Crossref | GoogleScholarGoogle Scholar |
Laves, K. S., and Loeb, S. C. (2006). Differential estimates of southern flying squirrel (Glaucomys volans) population structure based on capture method. American Midland Naturalist 155, 237–243.
| Differential estimates of southern flying squirrel (Glaucomys volans) population structure based on capture method.Crossref | GoogleScholarGoogle Scholar |
Liang, J., Tang, S., Nieto, J. J., and Cheke, R. A. (2013). Analytical methods for detecting pesticide switches with evolution of pesticide resistance. Mathematical Biosciences 245, 249–257.
| Analytical methods for detecting pesticide switches with evolution of pesticide resistance.Crossref | GoogleScholarGoogle Scholar | 23891583PubMed |
Losos, J. B. (1987). Postures of the military dragon (Ctenophorus isolepis) in relation to substrate temperature. Amphibia-Reptilia 8, 419–423.
| Postures of the military dragon (Ctenophorus isolepis) in relation to substrate temperature.Crossref | GoogleScholarGoogle Scholar |
Masters, P. (1996). The effects of fire-driven succession on reptiles in spinifex grasslnads at Uluru National Park, Northern Territory. Wildlife Research 23, 39–47.
| The effects of fire-driven succession on reptiles in spinifex grasslnads at Uluru National Park, Northern Territory.Crossref | GoogleScholarGoogle Scholar |
Melville, J., and Schulte, J. A. (2001). Correlates of active body temperatures and microhabitat occupation in nine species of central Australian agamid lizards. Austral Ecology 26, 660–669.
| Correlates of active body temperatures and microhabitat occupation in nine species of central Australian agamid lizards.Crossref | GoogleScholarGoogle Scholar |
Merrick, M. J., and Koprowski, J. L. (2017). Should we consider individual behavior differences in applied wildlife conservation studies? Biological Conservation 209, 34–44.
| Should we consider individual behavior differences in applied wildlife conservation studies?Crossref | GoogleScholarGoogle Scholar |
Michael, D. R., Cunningham, R. B., Donnelly, C. F., and Lindenmayer, D. B. (2012). Comparative use of active searches and artificial refuges to survey reptiles in temperate eucalypt woodlands. Wildlife Research 39, 149–162.
| Comparative use of active searches and artificial refuges to survey reptiles in temperate eucalypt woodlands.Crossref | GoogleScholarGoogle Scholar |
Michelangeli, M., Wong, B. B. M., and Chapple, D. G. (2016). It’s a trap: sampling bias due to animal personality is not always inevitable. Behavioral Ecology 27, 62–67.
| It’s a trap: sampling bias due to animal personality is not always inevitable.Crossref | GoogleScholarGoogle Scholar |
Montiglio, P. O., Ferrari, C., and Reale, D. (2013). Social niche specialization under constraints: personality, social interactions and environmental heterogeneity. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 368, 20120343.
| Social niche specialization under constraints: personality, social interactions and environmental heterogeneity.Crossref | GoogleScholarGoogle Scholar | 23569291PubMed |
Moseby, K., and Read, J. (2001). Factors affecting pitfall capture rates of small ground vertebrates in arid South Australia. II. Optimum pitfall trapping effort. Wildlife Research 28, 61–71.
| Factors affecting pitfall capture rates of small ground vertebrates in arid South Australia. II. Optimum pitfall trapping effort.Crossref | GoogleScholarGoogle Scholar |
Nersesian, C. L., Banks, P. B., and McArthur, C. (2011). Behavioural responses to indirect and direct predator cues by a mammalian herbivore, the common brushtail possum. Behavioral Ecology and Sociobiology 66, 47–55.
| Behavioural responses to indirect and direct predator cues by a mammalian herbivore, the common brushtail possum.Crossref | GoogleScholarGoogle Scholar |
Norbury, G., Spencer, N., Webster, R., Bailey, J., Walker, R., Wilson, R., Hunter, M., and Reed, C. (2002). Best-practice trapping of ferret populations. Landcare Research Contract Report LC 1, 140.
Noss, A. J. (1999). Censusing rainforest game species with communal net hunts African Journal of Ecology 37, 1–11.
| Censusing rainforest game species with communal net huntsCrossref | GoogleScholarGoogle Scholar |
NSW Department of Primary Industries (2020). ‘Use of pitfall traps.’ (NSW Department of Primary Industries.)
O’Farrell, M. J., Clark, W. A., Emmerson, F., Juarez, S. M., Kay, F. R., O’Farrell, T. M., and Goodlett, T. Y. (1994). Use of a mesh live trap for small mammals: are results from Sherman live traps deceptive? Journal of Mammalogy 75, 692–699.
| Use of a mesh live trap for small mammals: are results from Sherman live traps deceptive?Crossref | GoogleScholarGoogle Scholar |
Pianka, E. R. (1971). Ecology of the agamid lizard Amphibolurus isolepis in Western Australia. Copeia , 527–536.
| Ecology of the agamid lizard Amphibolurus isolepis in Western Australia.Crossref | GoogleScholarGoogle Scholar |
Pierce, C. L., Rasmussen, J. B., and Leggett, W. C. (1990). Sampling littoral fish with a seine: corrections for variable capture efficiency. Canadian Journal of Fisheries and Aquatic Sciences 47, 1004–1010.
| Sampling littoral fish with a seine: corrections for variable capture efficiency.Crossref | GoogleScholarGoogle Scholar |
Prout, D., and King, C. (2006). The effect of handling under anaesthetic on the recapture rate of wild ship rats (Rattus rattus). Animal Welfare 15, 63.
Purdie, R. (1984). ‘Land systems of the Simpson Desert region.’ (CSIRO Publishing: Melbourne, Vic., Australia.)
Read, J., and Moseby, K. (2001). Factors affecting pitfall capture rates of small ground vertebrates in arid South Australia. I. The influence of weather and moon phase on capture rates of reptiles. Wildlife Research 28, 53–60.
| Factors affecting pitfall capture rates of small ground vertebrates in arid South Australia. I. The influence of weather and moon phase on capture rates of reptiles.Crossref | GoogleScholarGoogle Scholar |
Réale, D., Reader, S. M., Sol, D., McDougall, P. T., and Dingemanse, N. J. (2007). Integrating animal temperament within ecology and evolution. Biological Reviews of the Cambridge Philosophical Society 82, 291–318.
| Integrating animal temperament within ecology and evolution.Crossref | GoogleScholarGoogle Scholar | 17437562PubMed |
Rodda, G. H., Savidge, J. A., Tyrrell, C. L., Christy, M. T., and Ellingson, A. R. (2007). Size bias in visual searches and trapping of brown treesnakes on Guam. The Journal of Wildlife Management 71, 656–661.
| Size bias in visual searches and trapping of brown treesnakes on Guam.Crossref | GoogleScholarGoogle Scholar |
Rolfe, J. K., and McKenzie, N. L. (2000). Comparison of methods used to capture herpetofauna: an example from the Carnarvon Basin. Records of the Western Australian Museum 61, 361–371.
| Comparison of methods used to capture herpetofauna: an example from the Carnarvon Basin.Crossref | GoogleScholarGoogle Scholar |
Schirmer, A., Herde, A., Eccard, J. A., and Dammhahn, M. (2019). Individuals in space: personality-dependent space use, movement and microhabitat use facilitate individual spatial niche specialization. Oecologia 189, 647–660.
| Individuals in space: personality-dependent space use, movement and microhabitat use facilitate individual spatial niche specialization.Crossref | GoogleScholarGoogle Scholar | 30826867PubMed |
Skalski, J. R. (1994). Estimating wildlife populations based on incomplete area surveys. Wildlife Society Bulletin (1973–2006) 22, 192–203.
Stokes, V. L. (2013). Trappability of introduced and native rodents in different trap types in coastal forests of south-eastern Australia. Australian Mammalogy 35, 49–53.
| Trappability of introduced and native rodents in different trap types in coastal forests of south-eastern Australia.Crossref | GoogleScholarGoogle Scholar |
Thompson, G., and Withers, P. (2005). The relationship between size-free body shape and choice of retreat for Western Australian Ctenophorus (Agamidae) dragon lizards. Amphibia-Reptilia 26, 65–72.
| The relationship between size-free body shape and choice of retreat for Western Australian Ctenophorus (Agamidae) dragon lizards.Crossref | GoogleScholarGoogle Scholar |
Walsh, R. N., and Cummins, R. A. (1976). The open-field test: a critical review. Psychological Bulletin 83, 482.
| The open-field test: a critical review.Crossref | GoogleScholarGoogle Scholar | 17582919PubMed |
Walsh, M. G., Fenner, D. B., and Winkelman, D. L. (2002). Comparison of an electric seine and prepositioned area electrofishers for sampling stream fish communities. North American Journal of Fisheries Management 22, 77–85.
| Comparison of an electric seine and prepositioned area electrofishers for sampling stream fish communities.Crossref | GoogleScholarGoogle Scholar |
Ward‐Fear, G., Rangers, B., Pearson, D., Bruton, M., and Shine, R. (2019). Sharper eyes see shyer lizards: collaboration with indigenous peoples can alter the outcomes of conservation research. Conservation Letters 12, e12643.
| Sharper eyes see shyer lizards: collaboration with indigenous peoples can alter the outcomes of conservation research.Crossref | GoogleScholarGoogle Scholar |
Williams, D. F., and Braun, S. E. (1983). Comparison of pitfall and conventional traps for sampling small mammal populations. The Journal of Wildlife Management 47, 841–845.
| Comparison of pitfall and conventional traps for sampling small mammal populations.Crossref | GoogleScholarGoogle Scholar |
Wilson, A. D. M., Binder, T. R., McGrath, K. P., Cooke, S. J., Godin, J.-G. J., and Kraft, C. (2011). Capture technique and fish personality: angling targets timid bluegill sunfish, Lepomis macrochirus. Canadian Journal of Fisheries and Aquatic Sciences 68, 749–757.
| Capture technique and fish personality: angling targets timid bluegill sunfish, Lepomis macrochirus.Crossref | GoogleScholarGoogle Scholar |