

The distribution, habitat preference and population dynamics of the pale field-rat (Rattus tunneyi) at Edel Land, Shark Bay, Western Australia: the role of refuges and refugia in population persistence
Sally O’Neill A B D , Jeff Short A C and Mike Calver A
A C and Mike Calver A
A Environmental and Conservation Sciences, Murdoch University, South Street, Murdoch, WA 6150, Australia.
B TERN Ecosystem Surveillance, GPO Box 498, Adelaide, SA 5005, Australia.
C Wildlife Research and Management Pty Ltd, PO Box 1360, Kalamunda, WA 6926, Australia.
D Corresponding author. Email: sally.oneill@adelaide.edu.au
Wildlife Research 48(5) 444-457 https://doi.org/10.1071/WR20005
Submitted: 7 January 2020 Accepted: 23 January 2021 Published: 17 May 2021
Journal Compilation © CSIRO 2021 Open Access CC BY
Abstract
Context: The pale field-rat (Rattus tunneyi) is a small native rat that formerly had a wide distribution throughout Australia. It has suffered substantial range contraction since European settlement and is now largely absent from arid and semiarid Australia. In this biome, it was known to persist only at two Western Australian locations: Edel Land, on the south-western shore of Shark Bay, and islands off the Pilbara coast.
Aims: We aimed to establish the extent of the species range at Edel Land, its habitat preference, the temporal stability of its populations with respect to rainfall, and threats to its persistence.
Methods: We trapped at 54 sites to establish distribution and habitat preference, and re-trapped four of these sites at which R. tunneyi was present in each season for 2.5 years to establish trends in abundance.
Key results: Trapping resulted in the capture of 45 R. tunneyi individuals across 17 of 54 sites (4104 trap-nights; 1.1% capture success). Rattus tunneyi typically occupied localised areas of dense shrubland, often in habitats with free water or near-surface moisture from drainage from high dunes allowing denser and taller vegetation and, at some sites, year-round growth of grasses or rushes. Regular re-trapping of four sites in each season (2002 – 2004) suggested a declining population, probably owing to a sequence of dry years.
Key conclusions: Rattus tunneyi at Shark Bay occurred only in localised mesic refuges, apparently dependent on seepage from high dunes generated by major inputs of rainfall from infrequent cyclones or sequences of high-rainfall years.
Implications: This isolated population is likely to be threatened by browsing by feral goats, opening up otherwise densely vegetated habitats of refuge areas, and their trampling of R. tunneyi burrows; by the depletion of grasses from herbivory by European rabbits; and by the long-term impact of a drying climate. It is unlikely to persist without effective on-going management, particularly of the goat population.
Keywords: climate change, irruption, Mus, native rodent, outbreak, refuge, source–sink.
Introduction
The identification and understanding of refugia for biodiversity have become a key conservation goal, particularly given projected anthropogenic climate change (Keppel et al. 2012; Reside et al. 2014). Refugia provide safe havens for species over ecological or evolutionary time scales, as distinct from refuges that provide protection to individuals during their lifetime (Davis et al. 2013; Reside et al. 2014). An early attempt to identify refugia for biodiversity within Australia’s semiarid and arid zone was that of Morton et al. (1995). These authors used the presence of relictual populations of native species as one of several factors to identify refugia. In one example, relictual populations of the native rat Rattus tunneyi, isolated from the contemporary core range of the species in the north and north-east of Australia, were used to identify clusters of islands in the Pilbara and mainland peninsulas at Shark Bay that formed refugia of value to a range of other species (Morton et al. 1995).
Whereas the refugial value of offshore islands may be obvious, being isolated from many of the threatening processes operating on the adjacent mainland, the same is not always true of mainland refugia. The pale field-rat or djini (R. tunneyi) persists on the peninsulas of Shark Bay, being far removed from its other mainland range; however, the reason for this persistence is unclear.
The pale field-rat is one of seven species of native Rattus occurring on the Australian mainland (Van Dyck and Strahan 2008). The species now occurs principally in native tussock grassland environments in north-eastern New South Wales, coastal Queensland and across the tropical north of Australia, having declined from >85% of its former range at the time of European settlement (Braithwaite and Griffiths 1996). Historical and subfossil records suggest that it once occurred through much of arid and semiarid Australia. However, it is now absent from the southern Northern Territory, from South Australia and from mainland Western Australia south of Broome. This decline has been attributed to destruction of its localised riparian refuges by introduced mammalian herbivores, principally domestic stock and rabbits (Braithwaite and Griffiths 1996; Aplin et al. 2008), which may affect the species directly or render it more vulnerable to predation.
One apparent exception to the species’ widespread decline in arid and semiarid Australia is a small remnant population detected at Edel Land on the southern shore of Shark Bay in Western Australia. Specimens were collected in an area of sparsely vegetated coastal dunes in 1968 and 1970 (Kitchener and Vicker 1981). However, little is known about the status of this population and the little that can be deduced from capture locations suggests specimens derived from atypical habitat.
Subsequent widespread trapping in 1989 across three of the four peninsulas of Edel Land targeting small mammals yielded only a single R. tunneyi individual (Sanders and Harold 1990). No R. tunneyi individuals were reported from an extensive survey of the southern Carnarvon Basin from October 1994 to March 1996, that included pitfall trapping within Edel Land (McKenzie et al. 2000). In contrast, Short et al. (2018) reported the irruption and collapse of a population of R. tunneyi at Heirisson Prong, some 40 km to the north-east of the sites of previous records.
The occurrence of the Edel Land population, some 1600 km south-west of its nearest mainland neighbour, is something of a paradox. This is a landscape of sandplain and tall sand dunes, with a predominant vegetation of low heath, Triodia hummock grasses and shrubland, an annual average rainfall of ~250 mm per annum, and with nowhere that could obviously be described as riparian habitat, being very different from the habitat of the species reported elsewhere (Watts and Aslin 1981; Woinarski et al. 1992; Braithwaite and Griffiths 1996).
Braithwaite and Griffiths (1996) emphasised the importance of mesic refuges to the persistence of R. tunneyi in the wet–dry tropics and Short et al. (2018) suggested that the species on Heirisson Prong may have irrupted from one or a few isolated mesic refuges. Pavey et al. (2017) reviewed the role of drought refuges in the regional persistence and irruptive dynamics of rodents in arid Australia, highlighting the importance of refuges to persistence through dry periods and providing source populations from which to irrupt during resource pulses triggered by wet periods. There is at least some indications that the dynamics of R. tunneyi may follow a similar pattern. If these refuges are degraded through grazing and trampling by introduced herbivores or by fire, the species sheltering there may be more susceptible to exotic predators, as noted by Newsome (1975) and McGregor et al. (2015).
Rattus tunneyi is primarily nocturnal, spending the day in shallow burrows in loose sandy soils (Aplin et al. 2008). It is mostly herbivorous, consuming mainly grass stems, seeds and roots (Watts 1977), although it is known also to feed on young trees by burrowing and eating their roots (Watts and Aslin 1981). They have a gestation period of ~22 days, are capable of giving birth to litters in rapid succession, have a typical litter size of about four, and may attain sexual maturity as early as 5 weeks of age (Watts and Aslin 1981).
The aims of this study were to (i) identify the distribution of R. tunneyi within Edel Land, (ii) describe its habitat, (iii) determine the dynamics of the population over time, particularly with respect to rainfall, (iv) assess aspects of its biology relevant to its persistence at, and possible irruption from, local refuges, and (v) establish the likely threats to its persistence. An overarching aim was to understand why this species has persisted at this site, so isolated from the remainder of its mainland range. The study was conducted between May 2001 and February 2004, in a period of below-average rainfall following a series of years of above-average rainfall. Given this, we predicted that field-rats would largely be detected at mesic sites with high levels of grass cover, a more restricted distribution than would be likely in years of above-average rainfall.
Materials and methods
Study site
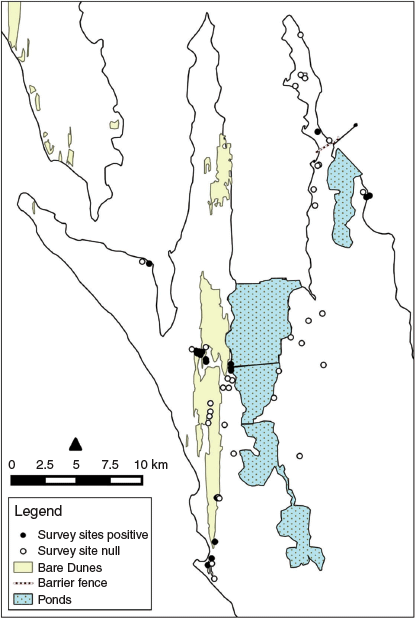
Edel Land is made up of four peninsulas, namely, Steep Point, Bellefin Prong, Heirisson Prong and Carrarang Peninsula, on the south-western margin of Shark Bay (Fig. 1). At the time of the study, the area was part of the 805 km2 Carrarang pastoral station, a commercial property running goats for live export. The pastoral history of this station was summarised by Short et al. (2018). Major concentrations of farmed goats during this study were on the southern parts of Heirisson Prong and Carrarang peninsulas. The Steep Point and Bellefin peninsulas were not utilised for stock at this time. However, derelict pastoral infrastructure (tanks, troughs and wells) was present and there were reports of cattle being grazed there in the 1980s (P. Dickenson, pers. comm., 2018).

|
The peninsulas of Steep Point and Bellefin Prong and the northern tip of Heirisson Prong were excised from Carrarang pastoral station and returned to the State Government in January 2008 (McCluskey 2008). They form part of the Shark Bay World Heritage Area and are proposed as the future Edel Land National Park.
The peninsulas of Steep Point and Bellefin Prong fall within the Coast land system (large linear and reticulate coastal dunes with some to >60 m in height, with minor limestone rises, and coastal cliffs to 100 m high); whereas Heirisson Prong falls largely within the Edel land system (undulating sandy plains with minor dunes to ~25 m and limestone rises; Payne et al. 1987). The Coast land system is considered highly susceptible to wind erosion if vegetation cover is depleted, and at least some parts consist of large ‘blowouts’ and mobile dunes (Fig. 1).
The climate of Edel Land is dry, warm Mediterranean (Payne et al. 1987). Rainfall in Shark Bay is mostly influenced by the winter rainfall regime of the south (on average some 70% of rain falls between May and August), rather than the summer monsoonal regime of the north. However, heavy rainfall events occur as a result of the interaction between frontal systems and tropical moisture sources (Wyrwoll et al. 2000) and occasional summer or autumn cyclones. Mean annual rainfall for Steep Point (1997–2017) is 248 mm and for Denham (1894–2017; 40–50 km to north-east) is 222 mm (sourced from the Bureau of Meteorology, http://www.bom.gov.au/climate/data/).
Steep Point, Bellefin Prong and False Entrance have a vegetation association of mid-dense dwarf shrub steppe dominated by Melaleuca cardiophylla and Thryptomene dampieri shrubs less than 1 m in height, with soft-leaved Triodia hummock grasses (Beard 1976; Shepherd et al. 2002). Both peninsulas have substantial areas of unvegetated and mobile sand dunes, with the largest some 20 km long by 2 km wide, oriented north–south. Heirisson Prong is more diverse. Much of the northern part of the peninsula, like Steep Point and Bellefin Prong, is mapped as dwarf shrub steppe of heath and hummock grass. The south-west is dominated by scrub heath of Acacia and mixed shrubs with a mid-dense canopy and a central area has a vegetation of shrub steppe (umbrella bush Acacia ligulata open shrubland with Triodia plurinervata hummock grasses). The south-eastern coast is dominated by open scrub of horse mulga Acacia ramulosa, and the far south-east of Heirisson Prong and Carrarang Peninsula are dominated by a succulent steppe of Acacia and Melaleuca shrubs higher than 1 m, with a ground layer of grey saltbush (Atriplex cinerea) shrubs. There are scattered birridas (low-lying gypsum pans) located in interdune depressions with succulent samphire Tecticornia sp. and sea heath Frankenia pauciflora low shrubs. Grey mangrove Avicennia marina occurs in scattered patches along the inland coasts.
Heirisson Prong is largely separated from the western peninsulas by a series of ponds that impound water for a local salt-harvesting operation established in the early 1960s (Fig. 1). Connection across these ponds is by a series of sparsely vegetated constructed bars up to 5 km long.
Feral goats (Capra hircus) were a significant problem in the past (Beard 1976) and were still common on Steep Point and Bellefin Prong (Sanders and Harold 1990) before intensive culling in the late 1990s. European rabbits (Oryctolagus cuniculus) are widespread throughout Edel Land and feral cats (Felis catus) and red foxes (Vulpes vulpes), both being likely predators of R. tunneyi, were common and widespread. A projected changing climate (declining annual or seasonal rainfall) may also be an issue for this species (NESP Earth Systems and Climate Change Hub 2018).
Distribution within Edel Land
Pale field-rats were surveyed by trapping at 54 sites across the Bellefin, Steep Point and Heirisson Prong peninsulas (32, 2 and 20 sites respectively). The choice of sites was informed by habitat descriptions for the species elsewhere (particularly the apparent preference for dense understorey with grass, sedge or rush species), examination of satellite imagery to identify likely key habitats, past trapping records for the species, and accessibility by four-wheel drive vehicle to areas with limited track access. There was particular emphasis on locating mesic or other apparent refuge sites within the broader landscape. These included infrastructure sites (such as water points where there was a substantial local impact on vegetation through water leakage) and natural sites where high dunes and deep swales were in close juxtaposition or high dunes closely abutted a coastline. Water drainage from high dunes and a watertable close to the surface at such sites typically meant that vegetation height and density were greater than is common across the peninsulas.
Survey sites were trapped using 48 folding aluminium traps (Elliott Scientific, Melbourne, Vic., Australia; 33 × 10 × 9 cm), and six cage traps (Sheffield Wire, Perth, WA, Australia; 55 cm × 20 cm × 20 cm) baited with rolled oats mixed with peanut butter. These were arranged in a 40 × 50 m grid with four rows of six trap stations with two Elliott traps at each, and a centre row with one cage trap per station. Trap stations were located 10 m apart. Cage traps were used as part of the grid to capture burrowing bettong (Bettongia lesueur) and so minimise its disturbance impact on Elliott traps. Bettongs were common on Heirisson Prong at the time of trapping (Short and Turner 2000). Cage traps are also known to be more successful than are Elliott traps in capturing Rattus rattus (Stokes 2013), a species potentially present at Edel Land (Short et al. 2018). Survey trapping was mostly conducted for a single night, but occurred for two successive nights on some grids to assess whether this provided any additional information on the detectability of R. tunneyi.
The 54 survey sites were each trapped once over a 2-year period from May 2001. Hence, sites were trapped at different times of the year and in varying seasonal conditions. To overcome possible problems associated with this, five sites were re-trapped at a different time of the year to establish whether there was a change in the detectable presence of R. tunneyi.
Habitat preference
Each survey site was assessed for a range of habitat attributes as well as pest animal status. These are detailed in Table 1. Measurements for most attributes were taken from within the area of the grid. However, assessment of ‘slope’ and ‘runoff’ required observation of the grid within the context of the immediate landscape. Elevation was determined from a SRTM 1 s digital elevation model, with 5 m contours derived from LiDAR (GeoscienceLiDAR sourced from GeoScience Australia). Mean plant cover was assessed from the average of 30 quadrats (0.5 × 0.5 m) randomly distributed within the grid, each assessed using categories adapted from the Braun–Blanquet scale by Tiver (1994). Signs of vertebrate presence (cats, foxes, rabbits, goats and sheep) were scored at the time of trapping from opportunistic observation of tracks in the sandy soil and of animal signs within the grid.

|
Differences in attributes between sites detected as occupied by R. tunneyi versus unoccupied were explored using non-parametric multivariate analysis (one-way PERMANOVA), based on the Bray–Curtis distance measure (for a justification, see Clarke and Warwick 2001). Before analysis, we range-standardised each variable between 0 and 1 by subtracting the smallest score from each value and dividing the result by the difference between the largest and smallest scores, allowing an equal impact of variables irrespective of measurement scale. Because the PERMANOVA was significant, we used similarity percentage (SIMPER; Clarke and Warwick 2001) to determine the contribution of individual variables to the difference. A principal-component analysis was conducted on the site–habitat variable matrix to reduce the dimensionality of the dataset and generate potential explanatory variable(s) of R. tunneyi occupancy.
In addition, multifactor analysis using discriminant analysis was used to establish whether a combination of variables could effectively discriminate between sites where field-rats were trapped and those where no field-rats were trapped. In this analysis, the two groups are separated along a canonical axis to produce maximal separation. The axis is a linear combination of the original variables as in principal components analysis, and eigenvalues indicate amount of variation explained. Each site is assigned to the group that gives minimal Mahalanobis distance to the group mean. The Mahalanobis distance is calculated from the pooled within-group covariance matrix, giving a linear discriminant classifier. The given and estimated group assignments are listed for each site. In addition, group assignment is cross-validated by a leave-one-out cross-validation (jackknifing) procedure. All analyses were conducted in the program PAST 3.2 (Paleontological Statistics; Hammer et al. 2001).
Dynamics
Four of the 54 survey sites were selected for regular ongoing monitoring to establish the temporal stability of R. tunneyi populations. Three sites, namely, Perched Swamp, Clough’s Bar South, and Clough’s Bar North, were located at the base of Bellefin Prong. A fourth site, Reverse Osmosis Plant, was located on Heirisson Prong, 1 km south of the town of Useless Loop. These sites were trapped at ~3-month intervals between January 2002 and May 2004, with trapping sessions corresponding with season (spring, summer, autumn, winter).
The Perched Swamp site was positioned in a shallow swale dominated by dense rush Juncus kraussii subsp. australiensis, with a sparse overstorey of Acacia ligulata and coastal daisybush Olearia axillaris. It did not have free surface water; however, because of its low position in the landscape, groundwater was likely to be close to the surface.
The Clough’s Bar South site was located adjacent to Useless Loop Inlet, at the second in a series of two hyper-saline waterbodies created by constructing barriers of earth across the inlet. The eastern edge of the grid was bounded by the shoreline to the east and a steep 60 m dune to the west and there was a small pond, where surface water was available year-round. Water invertebrates were abundant and tracks at the water’s edge indicated that goats regularly drank from it, suggesting that this was a fresh water source of reasonable quality. The site was dominated by dense tall stands of honeymyrtle Melaleuca huegelii subsp. pristicensis thickets in areas of lowest elevation, with mixed Acacia shrubland upslope and areas of lower and more open shrubland vegetation on a steep dune slope. The M. huegelii subsp. pristicensis thickets were up to 3 m high and often sparsely covered with dodder laurel Cassytha pomiformis. A narrow linear patch of rush Juncus kraussii subsp. australiensis was present adjacent to the inlet.
The Clough’s Bar North site was similarly bounded by a tall 60 m dune to the west and the inlet to the east. Another small dune running parallel to the main dune separated the site into a deep swale and a narrow coastal plain. The swale was dominated by dense, tall (3 m) thickets of mainly Acacia didyma, occasionally Melaleuca huegelii subsp. pristicensis or Acacia ligulata, with an understorey of shrubby twinleaf Roepera fruticulosa, climbing mulla mulla (Ptilotus divaricatus) and feather speargrass (Austrostipa elegantissima). Vegetation of the intervening dune consisted of low shrubland of Melaleuca cardiophylla, Thryptomene dampieri and Triodia bromoides. The coastal plain was dominated by dense, tall Melaleuca huegelii subsp. pristicensis. There was no surface water available within the grid.
The Reverse Osmosis Plant site was located at the base of a tall dune and had a vegetation of impenetrable shrubland up to 3 m tall of Acacia ligulata and coastal hibiscus Alyogyne cuneiformis, with areas of dense, tall (to 1.5 m high) marine couch Sporobolus virginicus. Water was available year-round because of leaking water from the water treatment cooling tower and an adjacent small holding dam, with this resulting in an extensive area of green grass of S. virginicus that was present all year.
A trapping grid of 1.2 ha was established at each of the four sites. Elliott traps were positioned in a nine by nine grid with traps spaced at ~10 m intervals. A single trap was placed at each trap station. Traps were baited with rolled oats and peanut butter, and set in late afternoon and checked at sunrise for three consecutive nights. Trap success and ‘minimum number alive’ (Krebs 1998) were calculated for each grid over time. Changing trap success over time was juxtaposed against monthly rainfall and also against the known timing of the most recent irruption of numbers of pale field-rats at Heirisson Prong.
Biology
Data were collected on all captured mammals, including date of capture, trap location, species, sex, hind-foot length, weight and reproductive status. Captured R. tunneyi individuals were marked either with pit tags (Implantable ID-100, Microchips Australia, Melbourne, Vic., Australia) injected under the skin at the scruff of the neck or by a unique ear punch (for juveniles less than ~30 g). All animals were released at the capture location. Female field-rats were classed as juveniles if <40 g (or a hind foot measurement of <23 mm), subadult if 41–60 g (or if their hind-foot measurement was <25.8 mm), or as adult if larger. Males were considered juvenile if <60 g (or a hind-foot measurement of <24 mm), subadult if 61–84 g or if their hind-foot measurement was <27.5 mm (Taylor and Horner 1973), and adult if they were larger.
The reproductive status of captured field-rats was assessed. Females were classed as having button teats (small inconspicuous, therefore non-lactating), distended teats (elongated non-lactating, or recently lactating teats, with or without fur around the teat), or lactating. Abdomens of females were palpated to detect pregnancies. Males were classified as having either abdominal or scrotal testes.
Indices of condition were assessed using the method of Krebs and Singleton (1993). Predicted weight was calculated using a regression for all available data for field-rats caught on Edel Land between 1994 and 2006 and compared with measured weight, to give an index of condition. One-way ANOVA was used to compare differences in condition indices between rats of each sex assessed as being reproductive and those that were not.
Rainfall
We used rainfall at Steep Point to examine links to the biology of R. tunneyi and rainfall at Denham to look at potential long-term climatic trends. We examined the 112 years of rainfall data from Denham to establish whether there was any evidence for a declining trend in either annual or seasonal rainfall by using simple linear regression.
Results
Distribution within Edel Land
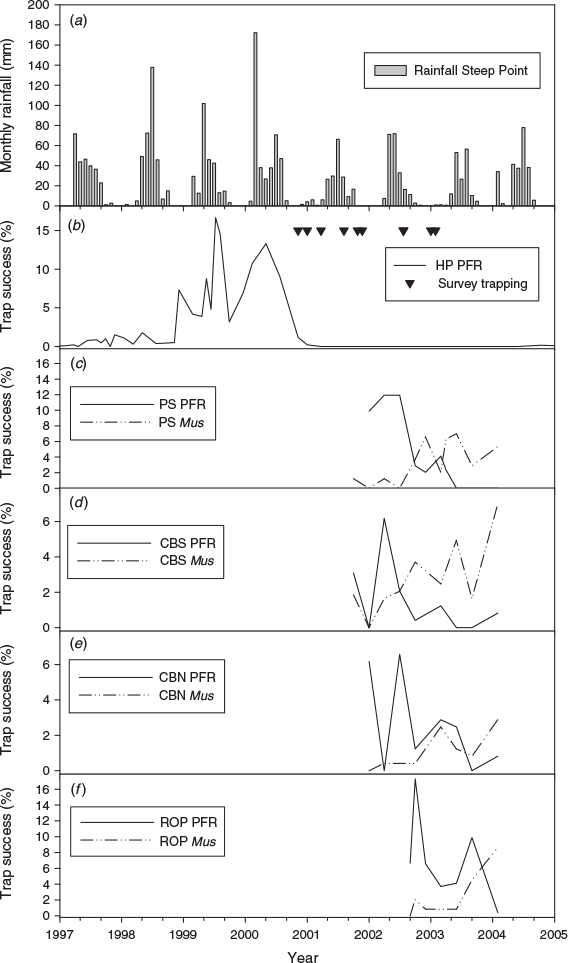
Fifty-four survey sites were trapped between May 2001 and September 2003 (Fig. 2). There were 45 captures of pale field-rats across 17 sites (31.5% of sites) in 4104 trap-nights, to give a trap success of 1.1%. Pale field-rats were trapped on all three peninsulas, predominantly on Bellefin Prong (41% of 32 sites) and Steep Point (50% of two sites), and less so on Heirisson Prong (15% of 20 sites). Figure 3a, b shows the timing of survey effort at Edel Land relative to the rainfall record and to the irruption and collapse of pale field-rats on Heirisson Prong reported by Short et al. (2018).
|
|
The adequacy of trapping for a single night on survey grids was assessed by trapping at 21 sites on multiple nights – either on two sequential nights or on two single nights separated by several months (i.e. trapped at different seasons). Sixteen survey sites were trapped on two successive nights. Twelve returned a null outcome on both nights and four returned a positive outcome on both nights. Hence, no additional information was gained by sampling over the second night. A further five survey sites were trapped on a single night on two separate occasions between 5 and 14 months apart to assess whether presence/absence was consistent over time. The same outcome (either presence or absence) was obtained at four sites (80%). No field-rats were caught at a site on Heirisson Prong where previously field-rats had been caught (re-trapped in March 2002 cf. October 2001).
Four other species of small mammals were caught (Table 2). The introduced house mouse (Mus musculus) was trapped at a rate of 1.33 individuals per 100 trap-nights at sites where R. tunneyi was trapped, and at about two-thirds that rate (0.87 per 100 trap-nights) at sites where no R. tunneyi was trapped. The sandy inland mouse (Pseudomys hermannsburgensis) showed the reverse pattern, being more common where R. tunneyi was absent (0.14 versus 0.40). The ash-grey mouse (P. albocinereus; three captures) and the little long-tailed dunnart (Sminthopsis dolichura; two captures) were rarely trapped.

|
Habitat preference
Positive records of R. tunneyi were spread across two land systems, namely, Coast (largely Bellefin Prong: 14 of 33 sites) and Edel (Heirisson Prong: 3 of 21). Positive records were at sites mapped by Beard (1976) as ‘bare areas, drift sands’ (8 of 17), ‘hummock grassland; dwarf shrub steppe; mixed ericoid shrubs and spinifex’ (7 of 17), or ‘hummock grasslands, shrub steppe; Acacia ligulata over Triodia plurinervata’ (2 of 17).
The majority of sites at which R. tunneyi was trapped were in shrubland environment (tall shrubland (4 sites); shrubland (8); low shrubland (2)), with the remainder being rushland (2), or in samphire on mangrove fringe (1). Key overstorey species typically included Acacia ligulata (8 sites), A. didyma (5 sites), Pileanthus limacis (4 sites), M. huegelii var. pristicensis (3 sites), Melaleuca cardiophylla (2 sites), and Juncus kraussii var. australiensis (2 sites). Hence, there was little or no correspondence between mapped vegetation association and assessed habitat type.
Grasses or rushes were present at most sites (15 of 17), but were considered a dominant part of the understorey only at a few sites (3 of 17). The exceptions were at sites with rushland or those with free water or coastal margins where Sporobolus virginicus was typically present. In total, 11 of 17 sites were located in a depression, swale or valley. Mean vegetation cover scores and maximum height of vegetation (Table 3) were typically higher at sites where R. tunneyi was present than at sites where it was absent.

|
Positive sites tended to be low in the landscape, often with high adjacent or surrounding dunes (landform means 2.18 vs 2.95). The mean altitude above sea level of all sites as determined from a digital elevation model was typically low, with occupied sites having a mean of ~12.1 m above sea level, compared with that of 17.3 m for null sites.
Signs of goats and/or sheep were detected at only 2 of 17 sites where R. tunneyi was caught (12%), compared with 8 of 37 null sites (22%). Signs of rabbits were detected at 4 of 17 sites where R. tunneyi was caught (24%), compared with 16 of 37 null sites (43%). Signs of foxes and/or feral cats were detected at only 9 of 54 sites (17%).
One-way PERMANOVA found significant differences between occupied and unoccupied sites (pseudo F1,917 = 3.12; P = 0.014). SIMPER indicated that five variables, namely, ‘myrtaceous understorey’ (10.6%), ‘rabbits present’ (9.9%), ‘monocots present’ (8.5%), ‘cover mean’ (7.8%) and ‘maximum species height’ (7.0%), caused 50% of the difference between the sites with and without field-rats. A principal component analysis showed that ‘rabbits present’ and ‘myrtaceous understorey’ (loadings of –0.501 and –0.470 respectively on PC1) were key variables indicating an absence of R. tunneyi, whereas ‘runoff’, ‘monocots dominant’ and ‘cover mean’ (0.359, 0.331 and 0.294) were key variables indicating the presence of R. tunneyi.
A discriminant analysis using a subset of six variables (selected to minimise incorrect assignment of occupied sites) correctly classified 79.6% of sites (63% jackknifed). One occupied site was misclassified (a mangrove fringe site) as were 10 null sites, giving an error of commission of 1.8% and an error of omission of 18.5%. The variables ‘rabbits present’ (–0.145), ‘landform’ (–0.142), and ‘sheep/goats present’ (–0.073) were all negative for the presence of R. tunneyi; ‘cover mean’ (0.153), ‘monocots present’ (0.152) and ‘runoff’ (0.059) were positive.
Dynamics
Four sites were trapped each season from July 2002 to March 2004. There were 586 captures of five mammal species from 8109 trap-nights conducted at the four sites combined (7.2% trap success). Rattus tunneyi and Mus musculus were the most abundant species trapped (310 and 204 captures respectively). Only low numbers of ash-grey mouse, sandy inland mouse, and little long-tailed dunnart were caught (56, 8, and 8 respectively). Overall trap success for R. tunneyi was 3.8% and that for M. musculus was 2.5%.
Overall, R. tunneyi showed a downward trend in trap success across the years of monitoring and M. musculus showed an increasing trend (Fig. 3c–f). Rattus tunneyi ‘minimum number alive’ declined from maxima in 2002 (Reverse Osmosis Plant: 23, Perched Swamp: 18; Clough’s Bar North: 14, and Clough’s Bar South: 11) to zero or near zero in 2003 (0–2) and 2004 (0–1).
Biology
All R. tunneyi individuals caught during both survey and monitoring had a mean weight of 81.4 g for males (n = 74; range 7.5–146 g) and 65.7 g for females (n = 78; range 20–102 g). Condition of R. tunneyi was calculated from the regression of weight on hind foot for all available captures, combining data from Heirisson Prong (Short et al. 2018) and the present study. This gave the equation
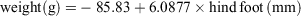
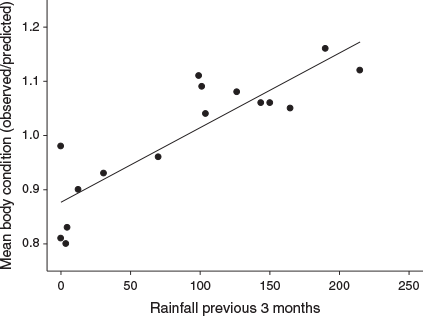
(r2 = 27.8%, F1,490 = 188.96, P < 0.01). Mean condition (observed weight/predicted weight) of field-rats was positively related to rainfall over the previous 3 months:

(r2 = 76.0%, F1,14 = 44.39, P < 0.001; Fig. 4). The mean condition index exceeded 1.0 when rainfall for the previous 3 months exceeded ~100 mm. Rainfall exceeded this 100 mm threshold for a 3-month period in late winter–early spring (August–October) in 2001, for a 3-month period in winter and early spring (July–September) in 2002, and a single month (September) in 2003. There were no cyclonic inputs of rainfall during the period of our study.
|
Male field-rats (≥60 g) assessed as reproductive had higher condition scores than did unreproductive males (scrotal versus abdominal testes; 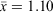 versus 0.883; F1,51 = 31.52; P < 0.001). Female field-rats (≥40 g) assessed as reproductive (distended teats or pregnant versus button teats) also had higher, albeit not significantly, condition scores than did non-reproductive females (
versus 0.883; F1,51 = 31.52; P < 0.001). Female field-rats (≥40 g) assessed as reproductive (distended teats or pregnant versus button teats) also had higher, albeit not significantly, condition scores than did non-reproductive females (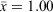 versus 0.90; F1,58 = 3.93; P = 0.052).
versus 0.90; F1,58 = 3.93; P = 0.052).
Rattus tunneyi appeared capable of breeding for most of the year with a hiatus in late autumn–early winter (Table 4). Males were recorded with scrotal testes in all months except May and June. Females were recorded as either pregnant or with distended teats in all months except May and September (although, no data were available for June or August). Juveniles were recorded in the population in September, October and January to April. However, this seasonal pattern appeared strongly influenced by rainfall. For example, there were no records of pregnancy or lactation for females captured in trapping from April 2003 to March 2004, the period of the study with lowest rainfall (only 148 mm of rain fell over the 18-month period to February 2004).

|
Rainfall
A simple regression of annual rainfall at Denham against year gave a non-significant result (F1,114 = 1.23; P = 0.269). However, a regression of winter rainfall (May to September) gave a significant negative slope of –0.44 (F1,113 = 7.74; P = 0.006), suggesting a 25% decline in winter rainfall since records began.
Discussion
Braithwaite and Griffiths (1996) identified the paradox of R. tunneyi, namely, why a species that can be a pest in some areas can show signs of vulnerability to local extinction in others. Our study at Edel Land looks at another paradox of that species, namely, how a small population can persist isolated by some 1600 km from its nearest mainland neighbour in a semiarid landscape, with no apparent riparian habitat.
Distribution within Edel Land
Rattus tunneyi is present in multiple sites in Edel Land, but within a restricted range of habitats. Its distribution appeared closely tied to the major dune system extending south–north along the length of Bellefin Prong and areas of denser, taller vegetation associated with favourable drainage of subsurface moisture from these high dunes.
The survey effort at Edel Land was during a period of well below-average annual rainfall (2001: 193 mm; 2002: 214 mm, and 2003: 165 mm) following 3 years of above-average rainfall (1998: 383 mm, 1999, 263 mm and 2000: 403 mm; Fig. 3a, b) and at a time when the population of R. tunneyi on Heirisson Prong had largely collapsed, contracting back to a single known refuge location (Short et al. 2018). Five of six survey grids north of the predator-barrier fence on Heirisson Prong (see Fig. 1), trapped after mid-2001 for the present study, yielded no field-rats, despite captures having occurred at the same sites 6–12 months earlier (Short et al. 2018), reflecting the collapse in the population.
Previous surveys targeting R. tunneyi or other small mammals in Edel Land have met with variable success, possibly being linked to the habitats surveyed and timing relative to cycles of wet and dry years. For example, the previous survey that caught most R. tunneyi in Edel Land (18 specimens, August 1970: Kitchener and Vicker 1981) had a rainfall in the prior 12-months of 313 mm compared with that of the survey by Sanders and Harold (1990; one specimen trapped in dense beach spinifex Spinifex longifolius on a sand ridge; July to November 1989) that had a rainfall of 195 mm. Although we were successful in trapping R. tunneyi at Edel Land by targeting specific habitats, further surveys might do well to employ camera traps in addition to traps (De Bondi et al. 2010; Rendall et al. 2014). These have been used to successfully establish site occupancy of a wide variety of rodents, including Rattus.
Edel Land has a much lower annual average rainfall (~250 mm) than do many sites where R. tunneyi has been reported, presumably owing to its dependence on seepage of groundwater from high dunes. For example, Woinarski et al. (1992) found that R. tunneyi occupied sites to the wetter end of the sampled gradient of mean annual rainfall (600–1700 mm) of fauna surveys in north-western Australia. In the site of least rainfall (Purnululu National Park, in north-east Western Australia), R. tunneyi occupied wet gorges, springs and dense riparian vegetation (Woinarski 1992).
Habitat preference
Beard (1976; see also Shepherd et al. 2002) mapped and described the vegetation communities of Shark Bay at a scale of 1:250 000, which was too coarse to identify refuge habitat for R. tunneyi. Hence, there was a disparity between mapped habitat and assessed habitat. Rattus tunneyi used localised mesic refuges with a vegetation different from that of the surrounding matrix.
The analyses of habitat attributes suggested important factors linked to mesic sites low in the landscape with dense vegetation (higher values of ‘maximum species height’, ‘cover mean’, ‘runoff’ and ‘landform’), with the presence or abundance of grasses (low values of ‘myrtaceous understorey’ and ‘rabbits present’ and high values of ‘monocots present’, ‘monocots dominant’ and ‘cover mean’). Sites with high values of ‘myrtaceous understorey’ typically had high vegetation cover, but may not have provided much grass for food. High densities of rabbits in Edel Land are associated with a virtual absence of grasses from the understorey, particularly after periods of below-average rainfall. For example, grass cover at Heirisson Prong decreased from 9% of projected foliage cover in a winter of above-average rainfall to <1% in the following winter and two summers when grasses were a major part of the winter diet of rabbits (Robley et al. 2001). The presence of domestic and/or feral goats at some sites probably decreased grass cover and overall vegetation cover.
Mesic sites were variously created by water drainage from nearby high sand dunes into deep swales or to narrow plains on the bay edge. In some cases, these sites had been utilised before as water points for stock or for trapping and mustering feral goats. It is likely that at such sites the watertable would be close to the surface, providing a mesic habitat for plant growth. Heights above sea level for such sites were typically 8–15 m, even when at some distance from the coast, suggesting close proximity to the watertable. The dense vegetation of such sites (typically shrubland) is likely to have provided shelter from foxes and feral cats and a continuity of shelter and food supply. The predominance of ash-grey mouse (a south-western species; Morris 2008) versus sandy inland mouse (an inland species; Breed 2008) in captures on the four monitoring grids suggested selection by R. tunneyi for more mesic sites. This is consistent also with the predominance of sandy inland mouse relative to ash-grey mouse on survey grids, the bulk of which did not have R. tunneyi present.
However, in contrast to prediction, the understorey of most sites was not dominated by grasslands. Green, growing grasses or rushes were present at some sites and appeared likely to be persistent despite local variations in rainfall. Marine couch Sporobolus virginicus was moderately common at some sites and Austrostipa elegantissima was moderately common growing on and through shrubs at some sites. The rush Juncus kraussii subsp. australiensis dominated the understory in swales at several sites, both in those with free water and those without. These were at low points in the landscape, often surrounded by a large catchment area of dunes, low areas of outcropping limestone, or sandplains draining to the swale. However, at other sites where R. tunneyi was captured, grasses were not a major component of the understory.
The two populations detected on Heirisson Prong were somewhat different and have been discussed at length in Short et al. (2018). The northern-most site was of a field-rat caught in October 2001, in an area of dense low shrubland of weeping pittosporum Pittosporum phillyreoides, some 4 km south of a 17-ha fox- and cat-proof enclosure where rabbits were controlled and tussock grasses (Austrostipa sp.) were notably more abundant than outside. Rattus tunneyi had been in high numbers at this latter site following a succession of years of above-average rainfall, but crashed in mid-2001 (Short et al. 2018). At the second southerly site, leaked water from the reverse osmosis plant allowed the growth of dense impenetrable shrubland with areas of dense tall grass. It is probable that this vegetation provided both cover and regular food to the field-rat population.
It is likely that many sites at which R. tunneyi was detected during the current study were refuge sites (Pavey et al. 2017) into which the species contracts during extended dry periods. Factors supporting this supposition include the location of sites at the base of large dunes or at permanent sources of water or areas of subsurface seepage, populations detected during years of low annual rainfall, and populations detected well after the collapse of the population on Heirisson Prong, attributed to a return to years of average or below-average rainfall.
However, the enduring survival of these enclaves of dense vegetation is likely to depend on the absence of farmed goats, sheep and cattle, low densities of feral goats, low densities of rabbits, and infrequent fires. A wide range of declared rare and priority fauna in this area is considered as being at risk also to over-browsing by goats (Desmond and Chant 2003). Domestic goats and sheep were not grazed on the two most westerly peninsulas during recent pastoral history and are not currently grazed there. In the late 1990s and early 2000s, feral goat control was undertaken by the pastoralist, who had the expertise, infrastructure (goat traps around water points), and economic incentive (by on-selling captured goats) to manage the feral population. Unfortunately, with the change in tenure in 2008, much of this management has lapsed. Feral goat activity was widely detected at many R. tunneyi sites in 2018 (J. Short, pers. obs.) that were recorded free of such activity during the present study.
The habitat of Edel Land shows parallels with that described for the species in northern Australia. In the central Kimberley, in northern Western Australia, R. tunneyi was detected in four major environments, one of which was sandseep (Legge et al. 2011). The greatest number of individuals of R. tunneyi was caught in sandseep relative to the other habitats (2.90/survey, cf. riparian zone 1.59, blacksoil, 0.29; and coolibah woodlands 0.26). Sandseeps were described as moist, thickly vegetated areas at the base of sandstone ranges. Sampling over a 4-year period coincided with destocking of habitats by cattle and other stock. All habitats were considered as preferred by cattle for foraging and/or resting.
Rattus tunneyi at Kakadu National Park in the Northern Territory was strongly associated with riparian environments, particularly waterholes maintained by seeping groundwater during the dry season (Braithwaite and Muller 1997). The extent and continuity of seeping groundwater was dependent on runs of years with high or low rainfall. A lack of seepage in runs of dry years led to waterholes drying earlier in the dry season, greatly affecting a range of species present, including R. tunneyi. Similarly, the description of habitat on Cobourg Peninsula appears close to that on Edel Land: ‘inland from tidal mangroves (and) in sand dunes’ (Watts and Aslin 1981, p. 23).
The habitat of R. tunneyi elsewhere in Australia consists largely of mesic grassland sites. Watts and Aslin (1981, p. 23) described R. tunneyi as ‘predominantly a grassland animal’. In the Northern Territory, the species was frequently found close to watercourses where grasses were thicker and, in south-eastern Queensland, it occurred in open forest with a dense grass understorey. This description of their habitat is consistent with what is known of their diet. Rattus tunneyi feeds on grass stems, seeds and roots (Watts 1977; Braithwaite and Griffiths 1996). Braithwaite and Griffiths (1996) suggested that R. tunneyi had a specialised diet consisting of nutritious food sources highly dependent on the availability of moisture typical in riparian environments.
The picture of habitat requirements is confused by the apparent ability of this species to greatly expand its range into less favourable habitat at times of high rainfall to make use of temporary high levels of cover and food (Short et al. 2018). However, there is likely to be a spatial as well as a temporal component to the species occupying habitat beyond mesic refuges. Braithwaite and Muller (1997) trapped R. tunneyi on pairs of grids, one in riparian habitat and one 500 m upslope. Over several years of sampling, captures averaged approximately four times higher on the riparian grids than on the corresponding upslope sites. Hence, field-rats were able to occupy both habitats, although clearly favouring one over the other.
Dynamics
Captures of R. tunneyi on monitoring grids on Edel Land were highly variable, with maxima of 6–12%. This compares with 25% at a predator-free refuge on Heirisson Prong (Short et al. 2018), 6–10% at Kakadu (Braithwaite and Muller 1997), 10% in hoop pine plantations and 0.8% in native habitat (Taylor and Horner 1973).
Short et al. (2018) found that trap success for R. tunneyi on Heirisson Prong was significantly correlated with rainfall over the previous 12- and 18-month periods. Rainfall for the years of this study (2001–03) was well below average, presumably accounting for the decline in trap success over time on the monitoring grids. This is reinforced by the strong relationship between body condition of field-rats and recent past rainfall. The year before the study (2000) had a high annual total (403 mm) and a significant aseasonal input of rainfall (172 mm in March; Cyclone Steve). It is unclear what time lag might exist between the current rainfall and the replenishment of soil and surface water at the base of major dunes by seepage to the benefit of R. tunneyi.
Biology
Much of R. tunneyi’s extant range is dominated by summer rainfall, but this does not apply to Edel Land. Edel Land has regular winter rainfall, supplemented on occasion by unpredictable cyclonic rainfall through summer and early autumn. Hence, a flexible breeding strategy would seem warranted. Substantial rainfall (>100 mm in a 3-month period) occurred only in late winter–early spring during our study. Despite low-rainfall years, we recorded evidence of breeding and recruitment of R. tunneyi across most months of the year.
Elsewhere, seasonality of breeding in R. tunneyi appears highly variable depending on location and, presumably, the seasonality of rainfall. Taylor and Horner (1973) reported that males with scrotal testes were trapped in mid-June in the Dampier Archipelago and there were juveniles in the population in November and January at sites on the north-western coast of Western Australia and in March at Alice Springs in the southern Northern Territory. Braithwaite and Griffiths (1996) reported a peak in breeding activity of R. tunneyi in March–May in the Northern Territory. Taylor and Calaby (2004) found that the species bred in the latter part of the wet season and the middle of the dry period in the wet–dry tropics, at which time seasonal suppression of breeding occurred in both adults and maturing individuals.
Conservation issues and prognosis for persistence at Shark Bay
The R. tunneyi population at Shark Bay appears to be small, localised and threatened on a range of fronts, from disruption of its mesic refuges by feral or domestic stock, from heightened predation from feral cats and other predators, from a drying climate, and from the interplay between these factors. Two factors in play elsewhere within its range are of less significance at Shark Bay, namely, uncontrolled fires and the presence of introduced black rats (R. rattus).
Mesic refuges are favoured by stock (Legge et al. 2019) and are highly vulnerable to grazing by stock and feral herbivores, particularly during drought (Braithwaite and Griffiths 1996). Stock, feral goats and other feral ungulates typically have a profound impact on native vegetation close to the water point and are also likely to affect R. tunneyi via soil compaction and trampling of their burrow systems (Ward-Fear et al. 2017). Trampling of R. tunneyi burrows by banteng (Bos javanicus) on the Cobourg Peninsula, Northern Territory was reported by Calaby and Keith (1974).
The concentration of R. tunneyi activity in localised mesic refuges may potentially make this species more vulnerable to predation from feral cats and other predators, particularly if the vegetation cover of refuges is reduced by grazing or fire. Feral cats may target small isolated populations of rats (Frank et al. 2014). Leahy et al. (2016) found that R. tunneyi declined after fire in savanna in northern Australia, largely because the decline in ground cover exposed the species to greater predation from feral cats, dingoes, and other predators. McGregor et al. (2015) found that the hunting success of feral cats was closely tied to habitat density, being some 70% in open habitat in contrast to 17% in habitats with dense grass or complex rock.
Similarly, Legge et al. (2019) found that predation by feral cats was enhanced and prolonged in areas where livestock have access to burnt areas, presumably due to the ongoing reduction in ground cover, resulting in a decrease in native mammal richness, including the presence and abundance of R. tunneyi. Burbidge and Manly (2002) showed that the impact of cats on native wildlife on islands was dependent on rainfall, being greatest in arid environments, due to higher densities of cats and, presumably, their more effective hunting in open habitats.
The continued viability of refuge populations on Edel Land appears to depend on large rainfall events and sequences of years of above-average rainfall. If rainfall is declining as a result of climate change, as suggested by the apparent decline in winter rainfall over the period of records, then the population could well be at risk.
One factor possibly contributing to the successful persistence of R. tunneyi on Edel Land may be the absence of R. rattus. Rattus rattus has not been detected from settlements on Edel Land, Peron Peninsula or Dirk Hartog Island (Palmer and Morris 2014). Burbidge and Manly (2002) suggested that the absence of R. tunneyi from near-coastal areas of north-western Australia where R. rattus is now abundant might be evidence of a negative effect of one species on the other. Rattus tunneyi occurs on at least 30 islands on this coast (with R. rattus being absent) and subfossils indicate that the species formerly occurred on the adjacent mainland (Burbidge and Manly 2002; Baynes and McDowell 2010; Burbidge and Abbott 2017).
In south-eastern Australia, an experimental reduction in R. rattus numbers in littoral rainforest resulted in significant and sustained increases in populations of the bush rat R. fuscipes (Stokes et al. 2009). Juvenile bush rats appeared particularly vulnerable, being largely absent from control untreated sites despite breeding in females, but responding rapidly to the removal of R. rattus via juvenile recruitment, immigration and increases in residency of females. Similarly, R. colletti appears to have a negative impact on R. tunneyi (Braithwaite and Griffiths 1996); when flooded out of their normal wetland habitat, they move up creek lines, displacing R. tunneyi.
On Melville Island, in the northern Australian savanna, fire frequency is a significant predictor of abundance of R. tunneyi and other small mammals (Davies et al. 2018). Rattus tunneyi was found to be most abundant in plots burnt triennially in the early dry season, presumably owing to this fire regime promoting and maintaining grass cover relative to annual burning. Similarly, a significant increase in numbers of R. tunneyi were reported over 4 years in the central Kimberley as a result of intensive fire management, combined with control of introduced stock (Legge et al. 2011).
Rattus tunneyi formerly occurred on Peron Peninsula and the adjacent mainland to the east of Edel Land, but is now locally extinct there (Baynes 1990). Morris (2000) proposed that R. tunneyi be translocated to Peron Peninsula to enhance its conservation status in this region. However, the likely success of such an action seems far from assured, given the lower rainfall at this site than at Edel Land. Success would likely depend on the local availability of permanent mesic sites protected from grazing.
Morris (2000) proposed that the western subspecies Rattus tunneyi tunneyi be listed as ‘Lower risk (near threatened)’ under IUCN guidelines. Woinarski et al. (2014) listed the western subspecies as ‘Near Threatened’, with island populations (19 in the Pilbara, seven in the Kimberley and 11 in the Northern Territory) considered important to its long-term conservation. However, they reported that it had been lost from some islands (in the Northern Territory) and was believed to be continuing to decline in range on the mainland (>50% over the past 10–20 years). This is in contrast to a significant increase in numbers reported over 4 years in the central Kimberley as a result of intensive fire management combined with control of introduced stock (Legge et al. 2011).
Morton et al. (1995) identified Edel Land as part of a refugia for biological conservation, in part on the basis of the presence of a remnant population of R. tunneyi. It seems that seepage from high dunes provides localised habitat that has buffered the species against past short- and long-term climate cycles. However, populations of R. tunneyi appear small and localised and subject to trampling and overgrazing by feral goats and excessive predation by foxes and feral cats, which may exacerbate the impact of drought and climate change. Hence, the long-term viability of the population of R. tunneyi at Shark Bay seems far from certain.
Conflicts of interest
The authors declare no conflicts of interest.
Acknowledgements
We thank Shark Bay Resources, Useless Loop Community Biosphere Group, CSIRO Sustainable Ecosystems and Earthwatch Australia for their support; Bruce Turner, Blair Parsons and numerous volunteers for their assistance with field work; and Paul Dickenson (Steep Point) and Tony Millar (Carrarang) for sharing their local knowledge. This research did not receive any specific funding. Animal ethics approval for trapping was granted by CSIRO Sustainable Ecosystems (#00/01-36) and a scientific licence by the Western Australian Department of Conservation and Land Management (SF003607). We thank Andrew Burbidge, Russell Palmer and four anonymous referees for reviewing an earlier draft of this manuscript.
References
Aplin, K. P., Braithwaite, R. W., and Baverstock, P. R. (2008). Pale Field-rat Rattus tunneyi (Thomas, 1904). In ‘The Mammals of Australia’. (Eds S. Van Dyck and R. Strahan.) pp. 698–699. (Reed New Holland: Australia.)Baynes, A. (1990). The mammals of Shark Bay, Western Australia. In ‘Research in Shark Bay. Report of the France-Australe Bicentenary Expedition Committee’. (Eds P. F. Berry, S. D. Bradshaw, and B. R. Wilson.) pp. 313–325. Western Australian Museum, Perth, Western Australia.
Baynes, A., and McDowell, M. C. (2010). The original mammal fauna of the Pilbara biogeographic region of north-western Australia. Records of the Western Australian Museum 78, 285–298.
| The original mammal fauna of the Pilbara biogeographic region of north-western Australia.Crossref | GoogleScholarGoogle Scholar |
Beard, J. S. (1976). ‘The vegetation of the Shark Bay and Edel areas. Western Australia. Map and explanatory memoir. 1:250 000 series.’ (Vegmap Publications: Perth, WA, Australia.)
Braithwaite, R. W., and Griffiths, A. D. (1996). The paradox of Rattus tunneyi: endangerment of a native pest. Wildlife Research 23, 1–21.
| The paradox of Rattus tunneyi: endangerment of a native pest.Crossref | GoogleScholarGoogle Scholar |
Braithwaite, R. W., and Muller, W. (1997). Rainfall, groundwater and refuges: predicting extinctions of Australian tropical mammal species. Australian Journal of Ecology 22, 57–67.
| Rainfall, groundwater and refuges: predicting extinctions of Australian tropical mammal species.Crossref | GoogleScholarGoogle Scholar |
Breed, W. G. (2008). Sandy Inland Mouse Pseudomys hermannsburgensis. In ‘The Mammals of Australia’. (Eds S. Van Dyck and R. Strahan.) pp. 636–637. (Reed New Holland: Australia.)
Burbidge, A. A., and Abbott, I. J. (2017). Mammals on Western Australian islands: occurrence and preliminary analysis. Australian Journal of Zoology 65, 183–195.
| Mammals on Western Australian islands: occurrence and preliminary analysis.Crossref | GoogleScholarGoogle Scholar |
Burbidge, A. A., and Manly, B. J. F. (2002). Mammal extinctions on Australian islands: causes and conservation implications. Journal of Biogeography 29, 465–473.
| Mammal extinctions on Australian islands: causes and conservation implications.Crossref | GoogleScholarGoogle Scholar |
Calaby, J. H., and Keith, K. (1974). Mammals. In ‘Fauna survey of the Port Essington District, Cobourg Peninsula, NT, Australia. Technical Paper No. 28’. (Ed. H.J. Frith and J.H. Calaby.) pp. 179–208. (CSIRO Division of Wildlife Research: Canberra, ACT, Australia.)
Clarke, K. R., and Warwick, R. M. (2001). ‘Change in Marine Communities: an Approach to Statistical Analysis and Interpretation.’ (Primer-E: Plymouth, UK.)
Davies, H. F., McCarthy, M. A., Rioli, W., Puruntatameri, J., Roberts, W., Kerinaiua, C., Kerinaula, V., Womatakimi, K. B., Andersen, A. N., and Murphy, B. P. (2018). An experimental test of whether pyrodiversity promotes mammal diversity in a northern Australian savanna. Journal of Applied Ecology 55, 2124–2134.
| An experimental test of whether pyrodiversity promotes mammal diversity in a northern Australian savanna.Crossref | GoogleScholarGoogle Scholar |
Davis, J., Pavlova, A., Thompson, R., and Sunnucks, P. (2013). Evolutionary refugia and ecological refuges: key concepts for conserving Australian arid zone freshwater biodiversity under climate change. Global Change Biology 19, 1970–1984.
| Evolutionary refugia and ecological refuges: key concepts for conserving Australian arid zone freshwater biodiversity under climate change.Crossref | GoogleScholarGoogle Scholar | 23526791PubMed |
De Bondi, N., White, J. G., Stevens, M., and Cooke, R. (2010). A comparison of the effectiveness of camera trapping and live trapping for sampling terrestrial small-mammal communities. Wildlife Research 37, 456–465.
| A comparison of the effectiveness of camera trapping and live trapping for sampling terrestrial small-mammal communities.Crossref | GoogleScholarGoogle Scholar |
Desmond, A., and Chant, A. (2003). Geraldton Sandplains 1 (GS1 - Edel subregion). In ‘A Biodiversity Audit of Western Australia’s 53 Biogeographical Subregions’. (Eds J. E. May, N. L. McKenzie, and S. McKenna.) pp. 252–264. (Department of Conservation and Land Management: Perth, WA, Australia.)
NESP Earth Systems and Climate Change Hub (2018). Climate change and the Shark Bay World Heritage Area: foundations for a climate change adaptation strategy and action plan. Earth Systems and Climate Change Hub Report No. 7, NESP Earth Systems and Climate Change Hub, Australia.
Frank, A. S. K., Johnson, C. N., Fisher, A., Lawes, M. J., Woinarski, J. C. Z., Tuft, K., Radford, I. J., Gordon, I. J., Collis, M., and Legge, S. (2014). Experimental evidence that feral cats cause local extirpation of small mammals in Australia’s tropical savannas. Journal of Applied Ecology 51, 1486–1493.
| Experimental evidence that feral cats cause local extirpation of small mammals in Australia’s tropical savannas.Crossref | GoogleScholarGoogle Scholar |
Hammer, Ø., Harper, D. A. T., and Ryan, P. D. (2001). PAST: paleontological statistics software package for education and data analysis. Palaeontologia Electronica 4, 4.
Keppel, G., Van Niel, K. P., Wardell-Johnson, G. W., Yates, C. J., Byrne, M., Mucina, L., Schut, A. G. T., Hopper, S. D., and Franklin, S. E. (2012). Refugia: identifying and understanding safe havens for biodiversity under climate change. Global Ecology and Biogeography 21, 393–404.
| Refugia: identifying and understanding safe havens for biodiversity under climate change.Crossref | GoogleScholarGoogle Scholar |
Kitchener, D. J., and Vicker, E. (1981). ‘Catalogue of Modern Mammals in the Western Australian Museum 1895–1981.’ (Western Australian Museum: Perth, WA, Australia.)
Krebs, C. J. (1998). ‘Ecological Methodology.’ (Benjamin/Cummings: Menlo Park, CA, USA.)
Krebs, C. J., and Singleton, G. R. (1993). Indices of condition for small mammals. Australian Journal of Zoology 41, 317–323.
| Indices of condition for small mammals.Crossref | GoogleScholarGoogle Scholar |
Leahy, L., Legge, S., Tuft, K., McGregor, H., Barmuta, L. A., Jones, M. E., and Johnson, C. N. (2016). Amplified predation after fire suppresses rodent populations in Australia’s tropical savannas. Wildlife Research 42, 705–716.
| Amplified predation after fire suppresses rodent populations in Australia’s tropical savannas.Crossref | GoogleScholarGoogle Scholar |
Legge, S., Kennedy, M. S., Lloyd, R., Murphy, S. A., and Fisher, A. (2011). Rapid recovery of mammal fauna in the central Kimberley, northern Australia, following the removal of introduced herbivores. Austral Ecology 36, 791–799.
| Rapid recovery of mammal fauna in the central Kimberley, northern Australia, following the removal of introduced herbivores.Crossref | GoogleScholarGoogle Scholar |
Legge, S., Smith, J. G., James, A., Tuft, K. D., Webb, T., and Woinarski, J. C. Z. (2019). Interactions among threats affect conservation management outcomes: livestock grazing removes the benefits of fire management for small mammals in Australian tropical savannas. Conservation Science and Practice 1, e52.
| Interactions among threats affect conservation management outcomes: livestock grazing removes the benefits of fire management for small mammals in Australian tropical savannas.Crossref | GoogleScholarGoogle Scholar |
McCluskey, P. (2008). ‘Shark Bay World Heritage Property Strategic Plan 2008 – 2020.’ (Department of Environment and Conservation: Perth, WA, Australia.)
McGregor, H., Legge, S., Jones, M. E., and Johnson, C. N. (2015). Feral cats are better killers in open habitats, revealed by animal-borne video. PLoS One 10, e0133915.
| Feral cats are better killers in open habitats, revealed by animal-borne video.Crossref | GoogleScholarGoogle Scholar | 26288224PubMed |
McKenzie, N. L., Hall, N. J., and Muir, W. P. (2000). Non-volant mammals of the southern Carnarvon Basin, Western Australia. Records of the Western Australian Museum 61, 479–510.
| Non-volant mammals of the southern Carnarvon Basin, Western Australia.Crossref | GoogleScholarGoogle Scholar |
Morris, K. (2000). The status and conservation of native rodents in Western Australia. Wildlife Research 27, 405–419.
| The status and conservation of native rodents in Western Australia.Crossref | GoogleScholarGoogle Scholar |
Morris, K. (2008) Ash-grey mouse Pseudomys albocinereus. In ‘The Mammals of Australia’. (Eds S. Van Dyck and R. Strahan.) pp. 611–613. (Reed New Holland: Australia.)
Morton, S. R., Short, J., and Barker, R. D. (1995). Refugia for Biological Diversity in Arid and Semi-arid Australia. A report to the Biodiversity Unit of the Department of Environment, Sport, and Territories. Biodiversity Series, Paper No. 4. Department of Environment, Sport, and Territories, Canberra.
Newsome, A. E. (1975). An ecological comparison of the two arid-zone kangaroos of Australia, and their anomalous prosperity since the introduction of ruminant stock to their environment. The Quarterly Review of Biology 50, 389–424.
| An ecological comparison of the two arid-zone kangaroos of Australia, and their anomalous prosperity since the introduction of ruminant stock to their environment.Crossref | GoogleScholarGoogle Scholar | 1221459PubMed |
Palmer, R., and Morris, K. D. (2014). A survey for black rats (Rattus rattus) in the Shark Bay communities of Denham, Monkey Mia and Useless Loop. Unpublished report to Department of Parks and Wildlife, Perth, WA, Australia.
Pavey, C. R., Addison, J., Brandle, R., Dickman, C. R., McDonald, P. J., Moseby, K. E., and Young, L. I. (2017). The role of refuges in the persistence of Australian dryland mammals. Biological Reviews of the Cambridge Philosophical Society 92, 647–664.
| The role of refuges in the persistence of Australian dryland mammals.Crossref | GoogleScholarGoogle Scholar | 26685752PubMed |
Payne, A. L., Curry, P. J., and Spencer, G. F. (1987). An inventory and condition survey of rangelands in the Carnarvon Basin, Western Australia. Western Australian Department of Agriculture Technical Bulletin No. 73, 478 pp.
Rendall, A. R., Sutherland, D. R., Cooke, R., and White, J. (2014). Camera trapping: a contemporary approach to monitoring invasive rodents in high conservation priority ecosystems. PLoS One 9, e86592.
| Camera trapping: a contemporary approach to monitoring invasive rodents in high conservation priority ecosystems.Crossref | GoogleScholarGoogle Scholar | 24599307PubMed |
Reside, A. E., Webergen, J. A., Phillips, B. L., Wardell-Johnson, G. W., Keppel, G., Ferrier, S., Williams, S. E., and Vanderwal, J. (2014). Characteristics of climate change refugia for Australian biodiversity. Austral Ecology 39, 887–897.
| Characteristics of climate change refugia for Australian biodiversity.Crossref | GoogleScholarGoogle Scholar |
Robley, A. J., Short, J. C., and Bradley, J. S. (2001). Dietary overlap between the burrowing bettong (Bettongia lesueur) and the European rabbit (Oryctolagus cuniculus) in semi-arid coastal Western Australia. Wildlife Research 28, 341–349.
| Dietary overlap between the burrowing bettong (Bettongia lesueur) and the European rabbit (Oryctolagus cuniculus) in semi-arid coastal Western Australia.Crossref | GoogleScholarGoogle Scholar |
Sanders, A., and Harold, G. (1990). ‘Search for the Shark Bay Mouse Pseudomys praeconis at Shark Bay on the Western Australian Mainland.’ (World Wildlife Fund and Department of Conservation and Land Management: Perth, WA, Australia.)
Shepherd, D. P., Beeston, G. R., and Hopkins, A. J. M. (2002). Native Vegetation in Western Australia, Extent, Type and Status. Resource Management Technical Report 249. Department of Agriculture, Western Australia.
Short, J., and Turner, B. (2000). Reintroduction of the burrowing bettong Bettongia lesueur (Marsupialia: Potoroidae) to mainland Australia. Biological Conservation 96, 185–196.
| Reintroduction of the burrowing bettong Bettongia lesueur (Marsupialia: Potoroidae) to mainland Australia.Crossref | GoogleScholarGoogle Scholar |
Short, J. C., O’Neill, S., and Richards, J. D. (2018). Irruption and collapse of a population of pale field-rat Rattus tunneyi at Heirisson Prong, Shark Bay, Western Australia. Australian Mammalogy 40, 36–40.
| Irruption and collapse of a population of pale field-rat Rattus tunneyi at Heirisson Prong, Shark Bay, Western Australia.Crossref | GoogleScholarGoogle Scholar |
Stokes, V. L. (2013). Trappability of introduced and native rodents in different trap types in coastal forest of south-eastern Australia. Australian Mammalogy 35, 49–53.
| Trappability of introduced and native rodents in different trap types in coastal forest of south-eastern Australia.Crossref | GoogleScholarGoogle Scholar |
Stokes, V. L., Banks, P. B., Pech, R. P., and Spratt, D. M. (2009). Competition in an invaded rodent community reveals black rats as a threat to native bush rats in littoral rainforest of south-eastern Australia. Journal of Animal Ecology 46, 1239–1247.
| Competition in an invaded rodent community reveals black rats as a threat to native bush rats in littoral rainforest of south-eastern Australia.Crossref | GoogleScholarGoogle Scholar |
Taylor, J. M., and Calaby, J. H. (2004). Seasonal reproductive dynamics in a tropical Australian rodent, Rattus tunneyi (Rodentia: Muridae). Australian Journal of Zoology 52, 159–168.
| Seasonal reproductive dynamics in a tropical Australian rodent, Rattus tunneyi (Rodentia: Muridae).Crossref | GoogleScholarGoogle Scholar |
Taylor, J. M., and Horner, B. E. (1973). Reproductive characteristics of wild native Australian Rattus (Rodentia: Muridae). Australian Journal of Zoology 21, 437–475.
| Reproductive characteristics of wild native Australian Rattus (Rodentia: Muridae).Crossref | GoogleScholarGoogle Scholar | 4803465PubMed |
Tiver, F. (1994). Vegetation patterns of eastern South Australia: edaphic control and effects of herbivory. Doctoral dissertation, Department of Botany, University of Adelaide, Adelaide, SA, Australia.
Van Dyck, S., and Strahan, R. (2008). ‘The Mammals of Australia.’ (Reed New Holland: Sydney, NSW, Australia.)
Ward-Fear, G., Brown, G. P., Pearson, D. J., and Shine, R. (2017). An invasive tree facilitates the persistence of native rodents on an over-grazed floodplain in tropical Australia. Austral Ecology 42, 385–393.
| An invasive tree facilitates the persistence of native rodents on an over-grazed floodplain in tropical Australia.Crossref | GoogleScholarGoogle Scholar |
Watts, C. H. S. (1977). The foods eaten by some Australian rodents (Muridae). Australian Wildlife Research 4, 151–157.
| The foods eaten by some Australian rodents (Muridae).Crossref | GoogleScholarGoogle Scholar |
Watts, C. H. S., and Aslin, H. J. (1981). ‘The Rodents of Australia.’ (Angus & Robertson Publishers: Australia.)
Woinarski, J. C. Z. (1992). A survey of the wildlife and vegetation of Purnululu (Bungle Bungle) National Park and adjacent area. Department of Conservation and Land Management, Perth, WA, Australia. Research Bulletin No. 6.
Woinarski, J. C. Z., Braithwaite, R. W., Menkhorst, K. A., Griffin, S., Fisher, A., and Preece, N. (1992). Gradient analysis of the distribution of mammals in Stage III of Kakadu National Park, with a review of the distribution patterns of mammals across north-western Australia. Wildlife Research 19, 233–262.
| Gradient analysis of the distribution of mammals in Stage III of Kakadu National Park, with a review of the distribution patterns of mammals across north-western Australia.Crossref | GoogleScholarGoogle Scholar |
Woinarski, J. C. Z., Burbidge, A. A., and Harrison, P. L. (2014). ‘The action plan for Australian mammals 2012.’ (CSIRO Publishing: Melbourne, Vic., Australia.)
Wyrwoll, K. H., Courtney, J., and Sandercock, P. (2000). The climatic environment of the Carnarvon Basin, Western Australia. Records of the Western Australian Museum 61, 13–27.
| The climatic environment of the Carnarvon Basin, Western Australia.Crossref | GoogleScholarGoogle Scholar |