

Toward a terrestrial biogeographical regionalisation of the world: historical notes, characterisation and area nomenclature
Juan J. Morrone A * and Malte C. Ebach B C
A * and Malte C. Ebach B C
A Museo de Zoología ‘Alfonso L. Herrera’, Departamento de Biología Evolutiva, Facultad de Ciencias, Universidad Nacional Autónoma de México (UNAM), 04510 Mexico City, Mexico.
B Earth and Sustainability Science Research, School of Biological, Earth and Environmental Sciences, University of New South Wales, Sydney, NSW 2052, Australia.
C Palaeontology Department, Australian Museum Research Institute, Sydney, NSW 2010, Australia.
Australian Systematic Botany 35(3) 187-224 https://doi.org/10.1071/SB22002
Submitted: 11 January 2022 Accepted: 5 May 2022 Published: 4 July 2022
© 2022 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution-NonCommercial 4.0 International License (CC BY-NC)
Abstract
An interim hierarchical classification (i.e. biogeographical regionalisation or area taxonomy) of the world’s terrestrial regions is provided, following the work of Morrone published in Australian Systematic Botany in 2015. Area names are listed according to the International Code of Area Nomenclature so as to synonymise redundant names. The interim global terrestrial regionalisation to the subregion level recognises 3 kingdoms, 2 subkingdoms, 8 regions, 21 subregions and 5 transition zones. No new names are proposed for the regions; however, Lydekker’s Line is renamed Illiger’s Line. We note that some regions still require area classification at the subregion level, particularly the Palearctic, Ethiopian and Oriental regions. Henceforth, the following interim global regionalisation may be used as a template for further revisions and additions of new areas in the future.
Keywords: area taxonomy, bioregionalisation, Ethiopian region, Holotropical kingdom, Nearctic region, Neotropical region, Neotropical subkingdom, Oriental region, Palearctic region, Paleotropical subkingdom, transition zones.
Introduction
North of the Equator also south we trace
Of either TROPIC the imagined place;
The space between, the TORRID ZONE we all;
Whose scorching belt surrounds this earthly ball;
Beyond the tropics, nearer to each pole,
More gentle skies the TEMPERATE ZONES control;
While at each pole is placed a FROZEN ZONE,
Where frost eternal rears his icy throne [Newton 1845, p. 2, The five zones of the Earth].
Our aim is to provide a detailed global biogeographical regionalisation (i.e. hierarchical classification) for the world’s terrestrial biota to the subregion level. Our study will advance the scheme of Morrone (2015a) by using a thorough biogeographical classification or area taxonomy (see Ebach and Michaux 2017) to justify each area as well as using the International Code of Area Nomenclature (ICAN; Ebach et al. 2008) to stabilise the proliferation of names by providing a synonymy, diagnosis and discussion for each valid name. Names within the biogeographical hierarchy usually fall into five levels, namely kingdoms (also known as realms), regions, dominions, provinces and districts. In some cases subkingdoms, subregions or subprovinces are recognised. The resulting hierarchical area taxonomy may be used as the basis for future global, intercontinental or regional biogeographic studies, without the need to apply new names. This study does not propose how to discover or identify natural biogeographic areas (i.e. endemic areas), rather we use the biogeographical literature to build a database of available names with a list of synonymies within a biogeographical hierarchy called an area taxonomy. We hope our study builds towards a global standard and database for biogeographic areas and names.
To understand the vast biogeographical literature and past discussions concerning the validity of certain areas, such as the Holarctic, a historical excerpt is presented below. For a full account, we recommend von Hofsten (1916) for early history of biogeography, Egerton (2012) for the history of ecological biogeography, Wallaschek (2009, 2010a, 2010b, 2011a, 2011b, 2012a, 2012b, 2012c, 2013a, 2013b, 2014, 2015a, 2015b) for the history of zoogeography, and Ebach (2015) for the history of 18th and 19th century regionalisation. The summary focuses on past controversies, including the description of natural regions and the acceptance of the Holarctic in the 19th, 20th and 21st centuries.
Summary of the history of biogeographical regionalisation
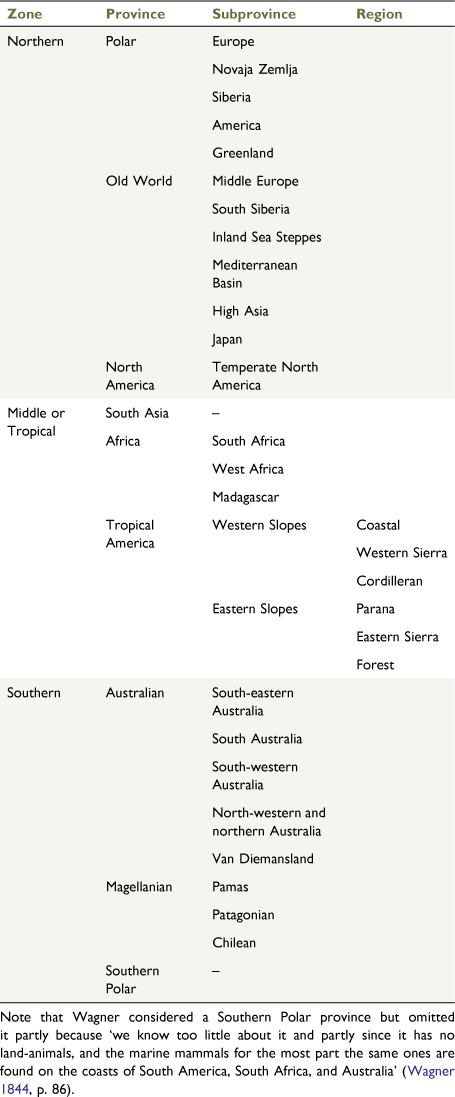
Global bioregionalisations were first proposed in the early 19th century, most notably by de Candolle (1805, 1820), Latreille (1815), Schouw (1823), Prichard (1826) and Swainson (1835). The first global hierarchical area classification was proposed by Wagner (1844, 1845, 1846a, 1846b) in a three-part monograph titled Die Geographische Verbreitung der Säugethiere Dargestellt [The geographical distribution of mammals], which was based on the mammalian distributions proposed by Illiger (1815; see also von Hofsten 1916; Kendeigh 1954; Smith 2005; Egerton 2012; Wallaschek 2015a; Ebach 2021). Wagner’s work contained the first global biogeographic map and area taxonomy (map republished in Hermogenes de Mendonça and Ebach 2020; area taxonomy republished in Ebach 2021; Fig. 1, Table 1). Unlike previous maps, such as Schouw’s Verbreitungsbezirk und Vertheilungsweise der Arten [Distributional areas and distributions of species] (Schouw 1823), Wagner’s map included all terrestrial areas rather than the distribution of economically important cereal crops (e.g. oats, wheat, maize, rice, rye). The map it most closely resembles is Zimmermann’s Tabula mundi geographico zoologica sistens quadrupedes: hucusque notos sedibus suis adscriptos [Geographical map of the world’s quadrupeds] (Zimmermann 1777, p. 1783; Fig. 2); however, Zimmermann’s map simply shows the distributions of terrestrial quadrupeds over a map of the Old and New Worlds (i.e. North and South America, Africa, Europe, Asia and Australia). Wagner’s map attempted to show the actual distributional areas of mammals as a whole. Wagner had created the first global biogeographic map that represented the first hierarchical classification (Table 1).

|
|

|
Natural areas
Prior to Wagner, there was much discussion among zoogeographers over the ‘naturalness’ of areas. Early naturalists, such as Fabricius (1778) and Latreille (1815), had proposed areas that Swainson (1835) had considered to be artificial. In fact, Kirby and Spence (1826) stated:
…any division of the globe into climates, by means of equivalent parallels and meridians, wears the appearance of an artificial and arbitrary system, rather than to one according to nature [p. 487].
Yet, this is exactly what phytogeographers such as Schouw (1823), Meyen (1846) and Grisebach (1872) and later zoogeographers (e.g. Forbes 1846; Schmarda 1853; Merriam 1892) were practising during the 19th century, a method that continues to this day (see Olson et al. 2001).
This approach differs from the identification of natural areas, that is, the earth being divided into naturally occurring biogeographical units on the basis of their endemic taxa, in the same way the living world is divided into taxa (e.g. species, genera, families). A world naturally divided into units would also apply at the biotic level; Australia has a unique flora and fauna, that is, a biota, which is very different from the biota of other continents with similar climates. Yet, when we observe this biota, we discover that plant species interact with the landscape (i.e. topography, soil, climate) and, therefore, produce plant forms and vegetation types. For example, an arid area will not produce the same types of trees you find in say, a rainforest. The plant forms in an arid area are stunted, have small leaves, where rainforest plants would be taller, form canopies and have larger leaves. The whole vegetation in fact would be different. A rainforest would have dense foliage, high trees all competing for sunlight, where an arid area would have a vegetation type that suits the dry and hot environment that it grows in.
Both approaches, the taxonomic and vegetational, can be described as ‘natural’; the taxonomic is a result of biogeographical homology and the vegetational is a result of plant forms. Yet, only the taxonomic is in anyway evolutionary in the simplest sense of the word. If we say that the family Macropodidae, namely the kangaroos and their allies, is endemic to Australasia, then we mean that kangaroos and all their nearest relatives are more closely related to each other (a monophyletic or natural taxon) than they are to anything else; that is, they are endemic to one area. With vegetation, this becomes problematic. A rainforest may be defined in a particular way, on the basis of the types of unique plant forms, rainfall, soil type and so on. This means that there is a universal rainforest, but the taxa that inhabit one rainforest may share only a distant relationship with the taxa in another. In this sense the plant forms and vegetation appear to be natural, but from an evolutionary standpoint they are artificial groups. The difference between taxonomic and vegetational regionalisations jarred with early ecologists such as Cowles (1908), who chastised taxonomy for producing artificial species, believing that taxonomists should be concerned with populations and their physiological forms, given that it is the environment that controls these forms.
Hierarchical area taxonomy
For the practical purposes of area taxonomy (i.e. biogeographical regionalisation, bioregionalisation, area classification), how one determines an area is not critical. Rather, it is how these areas relate that determines naturalness, and that relationship is based on biogeographical homology (Morrone 2001a; Ebach and Michaux 2017). Sclater (1858) hinted at this relationship being vital:
…little or no attention is given to the fact that two or more of these geographical divisions may have much closer relations to each other than to any third, and due regard being paid to the general aspect of their Zoology and Botany, only form one natural province or kingdom (as it may perhaps be termed), equivalent in value to that third [p. 130].
A decade later, Huxley (1868) made a similar statement on relationship:
I think it becomes clear that the Nearctic province is really far more closely allied with the Palæarctic than with the Neotropical region, and that the inhabitants of the Indian and the Æthiopian regions are much more nearly connected with another and with this of the Palæarctic region than they are with those of Australia [p. 314].
Yet, few 19th century plant and animal geographers tried to uncover natural biotic relationships. The biogeographic map produced by de Candolle (1805) attempted to use soils, climate and drainage to justify a relationship or connection, but failed. The link between biotic relationships and biogeographic areas came much later in the 20th century (Platnick and Nelson 1978; Rosen 1979; Nelson and Platnick 1981; Nelson and Ladiges 1996).
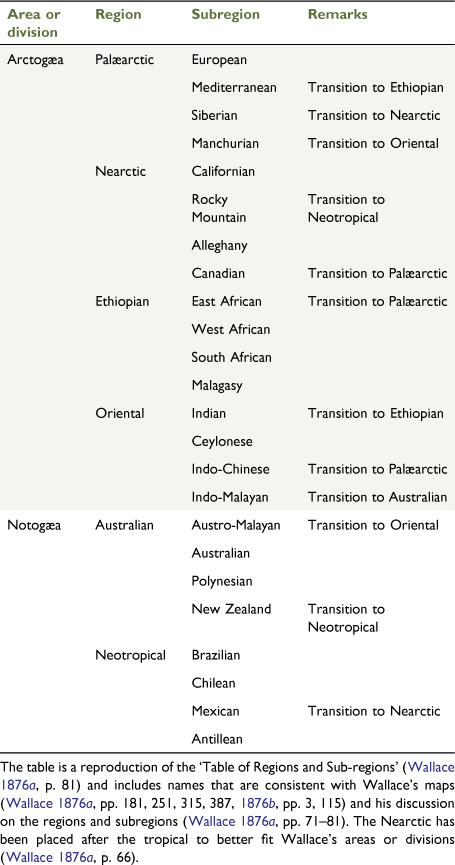
Sclater (1858) did provide a method to determine potential ‘natural’ areas by measuring endemicity and tallying the number of endemic bird taxa in each area. Essentially, Sclater’s areas were hierarchical in nature and covered all territorial areas, with the exception of the newly explored Antarctica and Subantarctic Islands. Sclater’s areas and his methodology of tallying up the numbers of taxa endemic to each area were adopted by Wallace (1876a, 1876b), who recognised Sclater’s six avian zoogeographic regions based on vertebrate distributions (Fig. 3, Table 2). Wallace preferred Sclater’s system because it initiated:
a more natural system, that of determining zoological regions, not by any arbitrary or a priori consideration but by studying the actual ranges of the more important groups of animals [Wallace 1876a, p. 53].

|
|
But not all of Sclater’s areas were adopted:
Mr. Sclater had grouped his regions primarily into Palaeogæa and Neogæa, the Old and New Worlds of geographers; a division which strikingly accords with the distribution of the passerine birds, but not so well with that of mammalia or reptiles [Wallace 1876a, p. 59].
Wallace’s biggest problem was the push back from other vertebrate zoologists, such as Heilprin (1882) and Allen (1871), who regarded the circumpolar zones as valid climatic areas based on a law of the distribution of life in circumpolar zones, which was addressed in some detail by Wallace (1876a):
But this supposed ‘law’ [of Allen 1871] only applies to the smallest details of distribution – to the range and increasing and decreasing numbers of species as well pass from north to south, or the reverse; while it has little bearing on the great features of zoological geography – the limitation of groups of genera and families to certain areas. It is analogous to the ‘law of adaptation’ in the organisation of animals, by which members of various groups are suited for an aerial, an aquatic, a desert, or an arboreal life; are herbivorous, carnivorous, or insectivorous; are fitted to live underground, or in fresh waters, or on polar ice [Wallace 1876a, p. 67, original emphasis].
Wallace here conjures up the stations and habitations of de Candolle (1820):
By the term station I mean the special nature of the locality in which each species customarily grows; and by the term habitation, a general indication of the country wherein the plant is native. The term station relates essentially to climate, to the terrain of a given place; the term habitation relates to geographical, and even geological, circumstances [de Candolle translated in Nelson 1978, p. 280, original emphasis].
Allen states:
The recognition of a ‘Nearctic’ as contradistinguished from a ‘Palæarctic region’ is almost equally arbitrary and at variance with the law of the distribution of life in circumpolar zones [Allen 1871, p. 382].
Other zoogeographers adopted the Holarctic including Blanford (1890), who used the term Aquilonian as an equivalent, and Lydekker (1896; see Arldt 1906 for further authors). The debate between Wallace and his adoptees (i.e. Sharpe 1893; Sclater and Sclater 1899; Fig. 4, 5) and North American zoologists such as Allen, Heilprin, Gill, Günther and Cope (neither of whom published a map) continued in society journals (see Ebach 2015). English ornithologist Alfred Newton, friend to both sides, tried to convince Wallace of the reality of a Holarctic by private correspondence shortly before the publication of the The Geographical Distribution of Animals (Wallace 1876a, 1876b):
Now taking the distribution of these genera from Baird and Sclater I find there are 13 genera wholly confined to the Nearctic region; 20 more of which all the species are Nearctic, but some of them extend to Mexico[,] a few more to Guatemala… [Wallace to Newton, 8 May 1875, Wallace Letters Online, see https://www.nhm.ac.uk/research-curation/scientific-resources/collections/library-collections/wallace-letters-online/4048/3992/B/details.html].
I am sorry you are so disturbed about distinctness of Nearctic and Neotrop[ic] regions. Your statistics do not in the slightest degree affect my conviction that they sh[oul]d be kept absolutely distinct [Wallace to Newton, 26 May 1875, Wallace Letters Online, see https://www.nhm.ac.uk/research-curation/scientific-resources/collections/library-collections/wallace-letters-online/4049/3993/B/details.html].
I will only remark now that you proceed on your supposition that my ‘Holarctic’ region = your Palaearctic and Nearctic – whereas the southern boundaries of this last are, in my opinion and that of several American zoologists, very uncertain… Thus a very considerable number of the genera, which you assign to your Nearctic and Palaearctic regions, belong really to more southern areas, and by their elimination your lists would present a very different aspect. Again too, you have omitted from your Nearctic list all the Palearctic genera of birds which inhabit Alaska, and if I am not mistaken these are several Mammals also, making Alaska essentially Palaearctic [Newton to Wallace, 17 June 1894, Wallace Letters Online, see https://www.nhm.ac.uk/research-curation/scientific-resources/collections/library-collections/wallace-letters-online/4300/4425/T/details.html].

|

|
Wallace was not persuaded by Newton’s argument and, by 1882, the argument has spilled over into the pages of Nature (see Ebach 2015):
[Heilprin] seeks to show that the Neoarctic [sic] and Palearctic should form one region, for which he proposes the somewhat awkward name ‘Triarctic region’, or the region of three northern continents [Wallace 1883, p. 482].
Briefly stated, it is maintained that … the Neoarctic [sic] and the Palaeoarctic [sic] faunas taken individually exhibit, in comparison with the other regional faunas (at least the Neotropical, Ethiopian, and Australian), a marked absence of positive distinguishing characters, a deficiency which in the mammal extends to families, genera, and species, and one which, in the case of the Neoarctic [sic] region, also equally (or nearly so) distinguishes the reptilian and amphibian faunas [Heilprin 1883, p. 605].
The facts of zoogeography are so involved, and often apparently contradictory, that a skilful dialectician with the requisite knowledge can make plausible argument for antithetical postulates. Prof. Heilprin, being a skilful dialectician and well informed, has submitted a pretty argument in favour of the union of the North American or ‘Nearctic’ and Eurasiatic or ‘Palaearctic’ [Gill 1885, p. 124].
We reject the term ‘Nearctic’ proposed by Mr. P. L. Sclater, and adopted by Mr. A. R. Wallace, for America north of Central America, for the reason that it seems to us an unnatural and artificial term… It is to be hoped that the term will not be adopted by American writers, as it is not by German and French writers, and we heartily endorse Mr. J. A. Allen’s protest against the use of the term by American writers on this subject [Packard 1883, p. 363; also in Allen 1892, p. 212, footnote 1; also see Ebach 2015, footnote 26].
Others retained Wallace’s usage for purely nomenclatural reasons:
I cannot understand why the word ‘Nearctic’ should be discarded. It was given by Dr. Sclater not in the sense of ‘arctic’ but ‘northern’ region of the New World, and is, in my opinion, apart from the priority which commands respect for its retention, a most simple and expressive term. My American colleagues will understand that if I have not carried their system of nomenclature unto the zoo-geographical regions of the Old World, it is not from any want of respect to their work, for I heartily agree with their conclusions as regards North America [Sharpe 1893, p. 101].
Sclater (1897) and later in their Geography of Mammals father and son Sclater and Sclater (1899) sided with Wallace for the simple reason that the Nearctic and Palearctic represented two distinct faunas:
the Palearctic and Nearctic regions have now, and have had in the past, quite sufficiently distinct faunas to warrant their division into two primary regions [Sclater 1897, p. 67].
[Allen’s] figures show that there is, as has indeed never been disputed, a great amount of similarity between the Nearctic and Palearctic faunas, but not enough to justify the junction of these two great land-masses into one ‘region’ or ‘realm’. [Sclater and Sclater 1899, p. 14].
Given this heated debate, are both sides talking past each other? If we look back to Sclater (1858), we find that his creations Neogeana and Palæogeana are based on the notion that the regions of the New World, for instance, share a closer relationship to each other than they do to the regions of the Old World and vice versa:
There are very many natural families which are quite peculiar to one or the other of these great divisions of the earth’s surface, more subfamilies, few genera really common to the two, and very few, if any, species [Sclater 1858, p. 133].
It is through classification, rather than through any distributional ‘laws’ that Sclater viewed zoogeography; after all, he firmly believed that each World was created by God (hence ‘creatio’). The notion that any distributional laws were in effect was never entertained. Wallace carried on this notion of relationship by tabling the world’s areas on the basis of vertebrates (Table 2). By placing the Australian and Neotropical regions into Notogea, he assumed relationship and not distributional laws. The idea that distributional laws shape area classification was introduced by the American zoogeographers who were more influenced by the proto-ecological ideas of the Humboldtians, who decided that the environment had a greater part to play in determining distribution. Again, here we see the division between those who see taxonomic distributions as natural versus those who feel that naturalness is something environmental. Nelson (1978) attempted to separate out these two practitioners into historical and ecological biogeographers, on the basis of the stations and habitations divide of de Candolle (1820).
One may wonder whether the Holarctic is the first major ecological zoogeographical region ever proposed. If so, it runs contrary to the Sclater–Wallacean or historical theme of natural areas based on endemicity, rather than on distributional ‘laws’.
The final word on 19th century zoogeographical regionalisation is that of Bartholomew et al. (1911). In their colourfully illustrated Physical Atlas, they redrew the published maps of Sclater (1858, 1897), Wallace (1876a, 1876b), Heilprin (1887), Lydekker (1896), Ortmann (1896) and Sclater and Sclater (1899). Bartholomew et al. (1911) admitted that:
…in a work of this kind it is impossible to attain absolute accuracy. Zoological literature has assumed such enormous proportions that a complete survey is quite impracticable… In the Text, a short historical account is given of the various systems propounded for the sub-division of the World Zoo-geographic Regions, wherein the views of the leading authorities are considered… Most of these are based upon the study of particular groups of animals –such as Mammals – but that of Dr Alfred Russel Wallace has a wider bearing, and hence has been accepted as a basis for the text of the present volume [Bartholomew et al. 1911, preface].
The short account of Bartholomew et al. (1911) starts with Sclater (1858) and ends with Sclater and Sclater (1899). Bartholomew et al. (1911) adopted the area classification of Wallace (1876a, 1876b), which is reiterated in Beddard (1895), a work that they describe as presenting ‘the subject in a different light from any of its predecessors’. Why Bartholomew et al. (1911) thought Beddard (1895) was any different to say Sclater and Sclater (1899) is unclear, as Beddard has adopted Wallace’s areas without any reference to the Holarctic. The wholesale disregard of the Holarctic is unusual, given that Wallace’s own Arctogæa incorporates the Nearctic and Palearctic.
In the spirit of Sclater and Wallace, we view the Holarctic as nothing more than a higher level area (a division of a larger area perhaps) that groups two regions on the basis of the notion of relatedness; that is, the Nearctic and the Palearctic share a greater relationship on the basis of the taxa they share, than to any other region. Yet, the adoption of the Holarctic, especially by zoologists, had taken much longer, with similar debates about its validity appearing in the early part of the 21st century (e.g. Escalante 2017), whereas others simply ignored it as a possible relationship among larger areas (e.g. Holt et al. 2013a, 2013b).
Adoption of Wallace’s areas by phytogeographers
Until the late 19th century, zoo- and phytogeography adopted separate classifications. For example, Engler (1882) divided the world’s flora into five main areas, namely the Northern and Southern Extratropical kingdoms, the Palaeotropical kingdom of the Old World, and the South American and Old Oceanic kingdoms (Fig. 6). Engler (1882) placed the Arctic region within the Northern Extratropical kingdom, an area resembling the Holotropical kingdom. Engler (1899) slightly altered the names and placed them all into the following five kingdoms: Northern Extratropical or Boreal, Palaeotropical, Central and South American, Austral (Old Oceanic) and Oceanic. The last kingdom was missing in the scheme of Engler (1882). Arldt (1907, map 1) used the same term ‘Arctic region’ to describe his circumpolar area in his Biogeographic Classification of the Continents (Fig. 7), which he classified as the Känogäisches kingdom, which includes the Holarctic region and three subregions, the Palearctic, Boreal and Nearctic. Arldt’s classification is unique, because it is based on terrestrial, freshwater, marine plant and animal distributions as well as the work of both zoo- and phytogeographers. His classification maybe seen as a review and an amalgam of 19th century plant and animal geography. Unfortunately, Arldt’s work is often overlooked in the history of biogeography (see Dowding and Ebach 2018), even though his own biogeographic classification looks incredibly modern by comparison to other early 20th century plant and animal geographers. In any case, Arldt (1907) preserved all of the Sclater–Wallacean regions as well as incorporating those of Heilprin (i.e. Holarctic) and Engler (i.e. Arctic) and served as a great revisionary work for 19th century biogeography.

|

|
Another work on global phytogeography is Good (1947, 1963; Fig. 8), who noted that:
Modern attempts to divide the world up into more or less equivalent floristic units mostly trace back to Engler’s scheme [Good 1963, p. 27].

Good (1947, 1963) simplified Engler’s classification system in his Geography of Flowering Plants, designating six floristic kingdoms, namely Boreal [sensu Holarctic], Paleotropical, Neotropical, South African, Australian and Antarctic (Table 2). Good continued:
This floristic classification may be epitomised by saying that it divides the land surfaces of the world into 37 regions [Good 1963, p. 31].
The other floristic classification scheme by Takhtajan (1978, 1986; Fig. 9) seemingly solidified phytogeography in the minds of 20th and 21st century biogeographers. Takhtajan reintroduced the Holarctic as a kingdom, converted Good’s Boreal kingdom into a subkingdom, and renamed Good’s South African and Antarctic kingdoms into the Cape and Holantarctic kingdoms respectively (Table 3). Takhtajan’s system may seem like Engler’s (1882, 1908); however, it does incorporate Heilprin’s Holarctic and several of the Sclater–Wallacean regions (Table 3), which were absent in previous floristic schemes (Engler 1882, 1899).

|

|
Later 20th and 21st century global biogeographic regionalisations
Global regionalisation after 1946 follows a similar theme of that of later 19th century area classification, including how to reconcile the Holarctic with the Nearctic and Palaearctic regions. Late 20th and 21st century biogeographical regionalisation may be divided into the following three groups: those who, knowingly or unknowingly, rejected the Holarctic in favour of the Nearctic and Palearctic (Udvardy 1975; Smith 1983; Kreft and Jetz 2010; Procheş and Ramdhani 2012; Holt et al. 2013a, 2013b; Rueda et al. 2013); those who accepted only a unified Holarctic (e.g. Schmidt 1954; de Lattin 1967; Rapoport 1968; Müller 1986; Cox 2001); and those who attempted to incorporate each under a larger division (Darlington 1957; Poynton 1959; Lopatin 1980; Morrone 2002, 2015a). Other problems raised in generating classification were the inclusion of New Zealand into the Antarctic or Holantarctic kingdom (i.e. Udvardy 1975), and whether to include or not transition zones (see Hermogenes de Mendonça and Ebach 2020 for a recent review).
Establishment of the Holarctic as a unified biogeographic area
The arguments for retaining the Holarctic are numerous. Darlington (1957) recognised six regions grouped into the following three kingdoms: Megagea (Ethiopian, Indomalayan, Palearctic and Nearctic regions), Neogea (Neotropical region) and Notogea (Australian region). Rather than choosing one classification over another, Darlington diplomatically noted:
Heilprin (1887) combined the two northern regions, the Palearctic and the Nearctic, into a Holarctic region. They are not combined here, but they may be called the Holarctic regions, and the animals which occur in both are often called Holarctic [Darlington 1957, p. 425].
If animals are Holarctic because they occur in both northern temperate zones, why not create a higher area ‘Holarctic’ in which the Nearctic and Palearctic are classified as subregions, as proposed earlier by Schmidt (1954) and Hershkovitz (1958)? Earlier 20th century zoogeographical classifications had included the Holarctic, with the Palaearctic and Nearctic as subareas, including Nichols (1943), who divided the freshwater fish fauna into ‘II. Continental: 1. Northern: A. Holarctic, a. Palaearctic and, b. Nearctic’ (Nichols 1943, p. 3); and Bobrinskiy et al. (1946), who placed the Holarctic region within Arctogea (see Beron 2018, p. 906). Herschkovitz noted that:
Darlington’s Zoogeography: the geographical distribution of animals [Darlington 1957] was received while the present paper was in press [Hershkovitz 1958, p. 594, footnote 1, original emphasis].
Poynton (1959), referring to Hershkovitz (1958), classified the Nearctic and Palearctic as subregions within a larger Holarctic region (Fig. 10), although without referring to Schmidt (1954).
|
Whereas the Holarctic was accepted by mammalogists (e.g. Hershkovitz) and herpetologists (e.g. Schmidt), entomologists were less convinced. German entomologist Gustav de Lattin in his Grundriß der Zoogeographie (de Lattin 1967) was one of the few to recognise the challenge, and formalised the Holarctic as a zoogeographic area, with the Palearctic and Nearctic as subregions (see Beron 2018), in what Illies called:
ein echter Fortschritt gegenüber der Darstellung durch DARLINGTON 1957 zu verzeichnen ist. Die wohlinformierte und ausgewogene Darstellung des holarktischen Raumes macht das Buch zu einem wichtigen Ereignis in der tiergeographischen Literatur [a real step forward when compared to the example by DARLINGTON in 1957. The well-informed and balanced representation of the Holarctic region makes the book (de Lattin 1967) an important event in the zoogeographic literature] [Illies 1968, p. 230].
Illies’ (1968) reference to Darlington is remarkable, given the number of already existing studies that accepted the Holarctic.
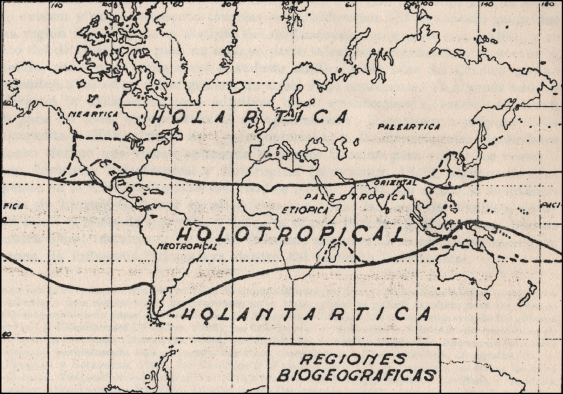
The acceptance of the Holarctic as a major area, which represents the Nearctic and Palearctic in their entirety or as subregions, was common in the second half of the 20th century. Rapoport (1968) proposed a biogeographic division of the earth into three regions or ‘biogeographic belts’, named Holarctic, Holotropical and Holantarctic (Fig. 11). Müller (1986) recognised nine divisions grouped into the following five kingdoms: Holarctic (Nearctic and Palearctic regions), Paleotropical (Ethiopian, Madagascan and Oriental regions), Australian (Australian, Oceanic, New Zealand and Hawaiian regions), Neotropical (Neotropical region) and Archinotic (Archinotic region). Bǎnǎrescu (1975) also recognised the Holarctic as a freshwater region, but not the Nearctic or Palearctic regions. Lopatin (1980) recognised the Arctogean kingdom containing the Palearctic and Nearctic regions, possibly because of the seniority of the name (Wallace 1876a, 1876b) to the later Holarctic (Heilprin 1882). The acceptance of the Holarctic continued into the late 20th and early 21st centuries. For example, Morrone (2002, 2015a) recognised the Holarctic as a kingdom and the Nearctic and Palearctic as its regions. Yet, the rejection of the Holarctic as a relational region began with the quantification of bioregionalisation.
|
Quantification of areas and the fate of the Holarctic
With the development of faster computer algorithms and the emergence of distributional databases, bioregionalisation entered a quantitative phase by the early 2000s. Early studies on global bioregionalisation included analyses using the distributional data of conifers (Sneath 1967), mammals (Smith 1983), Liliiforae (Conran 1995) and bumble-bees (Williams 1996). Both Sneath (1967) and Smith (1983) recovered the Holarctic, whereas the analyses by Conran (1995) and Williams (1996) did not.
Further bioregionalisation analyses based on bat (Procheş 2005) and gymnosperm distributions (Procheş 2006) found the Palearctic and Nearctic, but little similarity between them (i.e. they are more similar to other areas than to each other). The trend between recovering and not recovering the Holarctic continued with studies in the 2010s. In an analysis using vertebrate distributional data, Procheş and Ramdhani (2012) recovered both the Nearctic and Palearctic but failed to recover the Holarctic. They did admit, however, that using:
Wildfinder data, both of these approaches tended to yield clusters of very uneven geographic coverage, often small clusters in tropical America and large Holarctic or Palaeotropical clusters [Procheş and Ramdhani 2012, p. 261].
Rueda et al. (2013) recovered regions similar to those of Wallace (1876a, 1876b), using amphibian, bird and mammal data, and Escalante (2017) recovered Wallace’s Australian, Ethiopian, Neotropical, and Oriental regions by using mammal data. Neither study found evidence for the Holarctic.
Two studies that recovered the Holarctic were those of Kreft and Jetz (2010; Fig. 12), which used mammal distributional data, and Holt et al. (2013a, 2013b; Fig. 13), which used vertebrate data. The analysis of Kreft and Jetz (2010) found the Nearctic and Palearctic to have a greater similarity to each other, thereby justifying a Holarctic at the level of species, genus and family of volant and non-volant taxa (Kreft and Jetz 2010; Fig. 6). The analysis of Holt et al. (2013a) failed to mention the significance of their findings. Tucked away in the appendices is a dendrogram ‘of cross-taxon zoogeographic realms based on phyla-distributional data for amphibian, bird and non-marine mammal species of the world’ (Holt et al. 2013a, fig. S1), which shows the phylogenetic β diversity values between the Nearctic and Palearctic to justify a Holarctic kingdom; yet, the term was not used in the paper or in the appendices.

|

|
The usage of the Holarctic is justified both historically and quantitatively. Most quantitative studies have recovered the Holarctic, and, in doing so, the Holotropical. Given this long tradition that extends back to Heilprin (1882), we feel confident that future studies will recover the Holarctic, and by definition the Holotropical. A recent quantitative study using plant data did, indeed, recover three clusters that correspond strongly to the Holarctic, Austral and Holotropical.
Towards a terrestrial biogeographical regionalisation of the world
The biogeographical regionalisation presented below divides the world into three kingdoms, following Newbigin (1950), Kuschel (1963), Rapoport (1968), Morrone (2002, 2015a), Moreira-Muñoz (2007) and Carta et al. (2022). Within these kingdoms, we recognise 2 subkingdoms, 8 regions, 21 subregions and 5 transition zones (Fig. 14, Table 4). This interim global biogeographic classification to the subregion level takes into account all previous classifications and names within a formal nomenclatural synonymy that follows the ICAN (Ebach et al. 2008; and see below). Each list of synonymies is followed by a diagnosis and a list of regions. The name of each region is followed by a list of synonymies, a diagnosis and remarks. Within each region, the proposed subregions are provided, although we note that some of them, particularly in the Palearctic, Ethiopian and Oriental regions, still need analysis. In the areas of overlap between regions belonging to different kingdoms, five transition zones are delimited, as formerly recognised by several authors (e.g. Müller 1986; Halffter 1987; Morrone 2006, 2014, 2015a, 2015b; Kreft and Jetz 2013). No new names are proposed in this study.

|
|
The naming system within this and other biogeographic classifications follows the ICAN (Ebach et al. 2008). The ICAN was proposed so as to stem the flow of area names and to formalise a practise, so that one diagnosed area is assigned a single name. A biogeographical nomenclature as proposed by Wallace (1894) did not question:
who is right and who is wrong in the naming and grouping of these regions, or of determining what are the true primary regions. All proposed regions are, from some points of view, natural, but the whole question of their grouping and nomenclature is one of convenience and of utility in relation to the object aimed at [p. 613].
In other words, nomenclature, that is how biogeographers name areas, is not part of the discovery process of biogeography, but rather a convenient way to organise names without confusion. We may draw an analogy to the Botanical and Zoological Codes of Nomenclature that ensure one taxon is assigned a single name for the purposes of unambiguous communication of names. How biogeographers discover natural areas is another topic entirely and should not be confused with area nomenclature.
The biogeographic names proposed by de Candolle (1820), Prichard (1826), Schouw (1823), Wagner (1844, 1845, 1846a, 1846b) and Schmarda (1853) are not included in the synonyms, because they were published before Sclater’s (1858) regionalisation. We apply a criterion analogous to the nomen conservandum convention of taxonomical nomenclature to provide a greater stability (Morrone 2014). Changing well known area names from the Sclaterian–Wallacean system to the names used by these authors will only confuse the area nomenclature further. Given that areas such as the Nearctic and Palearctic are already widely used in the biogeographic literature, keeping them seems to be prudent and in keeping with a stable area nomenclature.
Area taxonomy
Holarctic kingdom: Heilprin (1882)
Diagnosis
North America, Greenland, Europe, Africa north of the Atlas mountains and Asia north of the Himalayan mountains.
Remarks
In contrast to some authors that have considered the Holarctic to represent a region, we treat it as a kingdom and subordinate the Nearctic and Palearctic to it (Morrone 2002, 2015a; Carta et al. 2022). From a palaeogeographic viewpoint, it corresponds to the palaeocontinent of Laurasia (Amorim and Tozoni 1994).
Regions
The Holarctic kingdom comprises two regions, Nearctic and Palearctic.
Nearctic region: Sclater (1858)
Diagnosis
Canada, mainland USA, central and northern Mexico and Greenland.
Remarks
So as to accommodate both the Nearctic and Holarctic in a single classification, without revising or synonymising these areas, we place the Nearctic and Palearctic as regions within the Holarctic kingdom.
Subregions
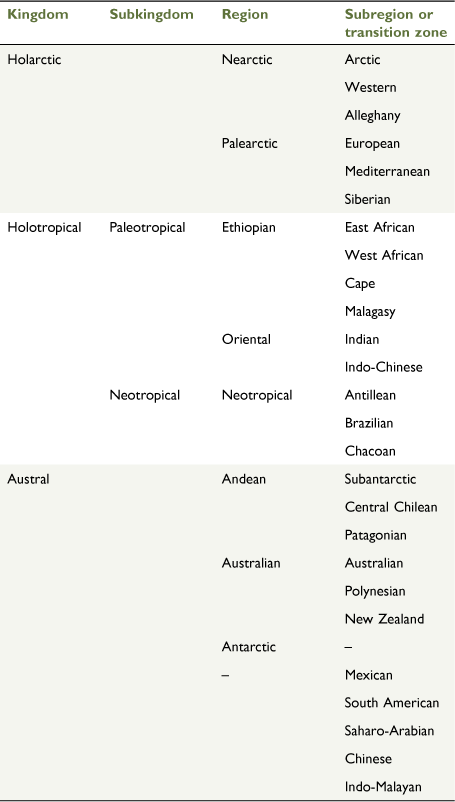
The Nearctic region comprises the Arctic, Western and Alleghany subregions (Escalante et al. 2021; Fig. 15).
Arctic subregion: LeConte (1859)
Diagnosis
Alaska (including the St Lawrence, Diomede and Aleutian Islands), northern Canada (Yukon, Northwest Territories and Nunavut, northern parts of British Columbia, Alberta, Saskatchewan, Manitoba, Ontario and Quebec, and Newfoundland and Labrador) and Greenland.
Western subregion: Allen (1871)
Diagnosis
The area west of, and including, the Rocky Mountains and its eastern foothills, Graham Island (British Columbia) in the north-west, following the lowlands south of Peace River (British Columbia) to Winnipeg (Manitoba). The eastern boundary follows the whole Eastern System of the Western Cordillera of North America, starting from Regina (Saskatchewan). The southern boundary extends to the Sierras Madre and the Baja California Peninsula to the south-west in Mexico.
Alleghany subregion: Cooper (1859)
Diagnosis
The area east of the Eastern System of the Western Cordillera of North America, south of the Arctic subregion (including Nova Scotia) and east of the Mid Atlantic Ridge (not including Iceland).
Palearctic region: Sclater (1858)
Diagnosis
Eurasia, northern China, Japan and Africa north of the Sahara.
Remarks
The Palearctic is included as a region within the Holarctic kingdom so as to accommodate to conflicting classifications (see discussion above).
Subregions
The Palearctic region has been classically divided into three subregions, namely European, Mediterranean and Siberian (Fig. 16). Further studies are still needed to corroborate them.

European subregion: Wallace (1876a)
Diagnosis
European continent south of the geographical North Pole (including Svalbard, Novaya Zemlya and Franz Joseph Land); north of the Pyrenees, Alps, Carpathian, Caucasus and Balkan mountains and Black Sea (including the Sea of Azov); west of the Mid Atlantic Ridge (including Iceland and Jan Mayen island); and west of the Ural Mountains, Volga and Don rivers.
Mediterranean subregion: Wallace (1876a, 1876b)
Diagnosis
Area south of the Pyrenees, Alps, Carpathian, Caucasus and Balkan mountains, the Sea of Azov, the Volga and Don Rivers, the Caspian Sea, and the Tibetan Plateau; west of the Indus River valley; north of the Persian Gulf (from Bahrain) and the Gulf of Oman. The southern boundary runs from Bahrain to the north of the Red Sea (Suez, Egypt) to the southern border of Western Sahara (including Macaronesia).
Siberian subregion: Wallace (1876a, 1876b)
Diagnosis
Area east of the Ural Mountains, including Siberia, Caspian Sea ending at Kamchatka and the Bering Strait. Includes the Tibetan Plateau and Mongolia and Gobi deserts.
Holotropical kingdom: Rapoport (1968)
Diagnosis
Tropical areas of the world, between 30° south latitude and 30° north latitude.
Remarks
It corresponds to Tropical Gondwanaland (Amorim and Tozoni 1994).
Subkingdoms
We follow the recent analysis of Carta et al. (2022), who postulated the existence of two subkingdoms, namely Paleotropical and Neotropical.
Paleotropical subkingdom: Engler (1882)
Diagnosis
Tropical areas within the continents of Africa and Asia (Carta et al. 2022).
Regions
The Paleotropical subkingdom comprises two regions, namely Ethiopian and Oriental.
Ethiopian region: Sclater (1858)
Diagnosis
Central and southern Africa, the northern part of the Arabian peninsula, Madagascar and the West Indian Ocean islands.
Remarks
The Cape region of previous regionalisations is demoted to a subregion of the Ethiopian region following Carta et al. (2022) and Santos and Ribeiro (2022). Further studies using phylogenetic data within a comparative biogeographic analysis are needed to confirm this classification.
Subregions
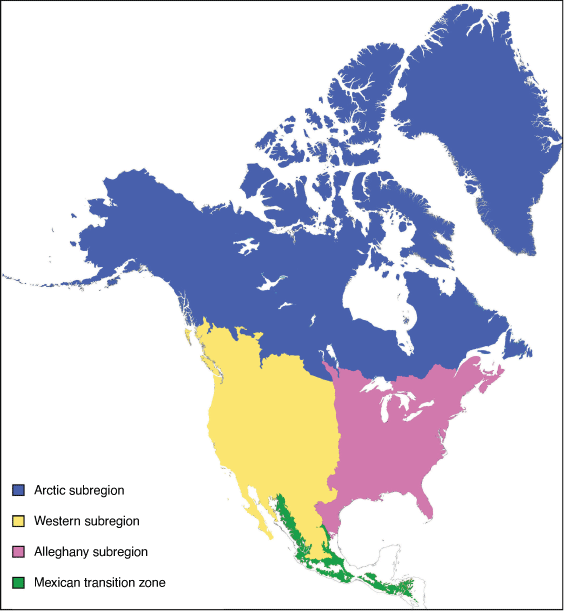
The Ethiopian region comprises the following four subregions: East African, West African, Cape and Malagasy (Fig. 17).
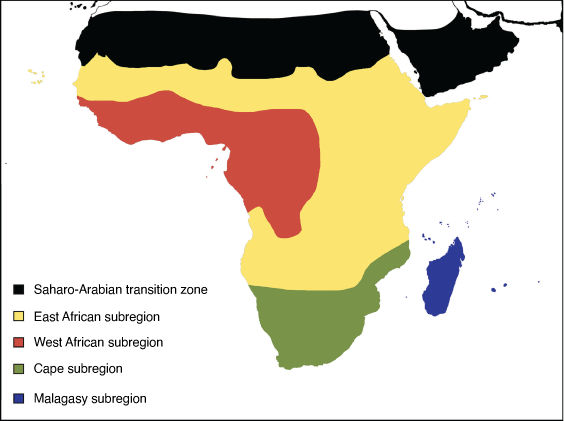
East African subregion: Wallace (1876a)
Diagnosis
Highland areas along the eastern and south-western part of the African continent (including Angola and northern part of Namibia); including a portion of the south-western and southern Sahara (southern parts of Mauritania, Mali, Niger, Chad, Sudan), the Rift Valley (including the Horn of Africa), and the southern part of the Arabian Peninsula (from Bahrain to Suez, Egypt), going as far south, but not including, the Cape subregion.
West African subregion: Wallace (1876a)
Diagnosis
Lowland sub-Saharan areas to the west and north of the East African subregion.
Cape subregion: Grisebach (1872)
Diagnosis
The area south-east of Lake Malawi in the Rift Valley (Mozambique), south of the Zambezi River (Zimbabwe) and along the northern borders of Botswana and Namibia.
Malagasy subregion: Wallace (1876a)
Diagnosis
Madagascar and surrounding islands (Seychelles, Comoros, Mayotte and Mascarene islands).
Oriental region: Wallace (1876a)
Diagnosis
All areas south of the Tibetan Plateau and the Yellow River and west of Weber’s Line.
Remarks
A comparative biogeographic analysis by King and Ebach (2017) has shown that Palaeogene genera from Australia, southern New Guinea and eastern Sulawesi share a greater relationship to another than they do to Neogene genera (ages based on molecular dating). The results support Wallace’s Line as the border between the Oriental and Australian region, rather than Weber’s or Lydekker’s lines.
Subregions
The Oriental region comprises two subregions, Indian and Indo-Chinese (Fig. 18).
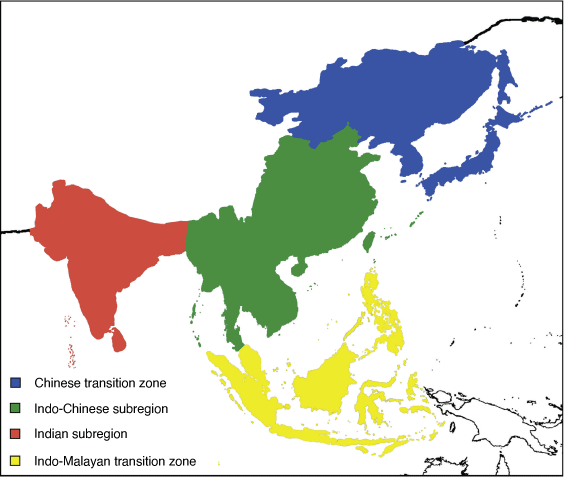
Indian subregion: Sclater (1858)
Diagnosis
Indian subcontinent, including the island of Sri Lanka, south of the Himalayas bordered by the Indus to the west and the Chittagongs to the east.
Indo-Chinese subregion: Wallace (1876a)
Diagnosis
Areas east of the Chittagongs, including the Andaman and Nicobar Islands, north of the Ta Pi River including the Phuket Range, bordered by the South China Sea in the west, by the islands of Taiwan and Hainan as well as the Ryukyu islands south of and including the Amami Islands (Japan) marked in the north-east by the Watase Line. Area bordered in the south by the Balintang Channel north of the Philippines, the Parcel Islands south of China, and Phú Quý and Côn Đảo islands to the south-east of Vietnam.
Neotropical subkingdom: Sclater (1858)
Diagnosis
Tropical South America, Central America, south-central Mexico and the Caribbean.
Regions
The Neotropical subkingdom comprises the Neotropical region.
Neotropical region: Sclater (1858)
Diagnosis
The same as the subkingdom.
Subregions
The Neotropical region comprises three subregions, namely Antillean, Brazilian and Chacoan (Morrone 2014, 2017; Fig. 19).
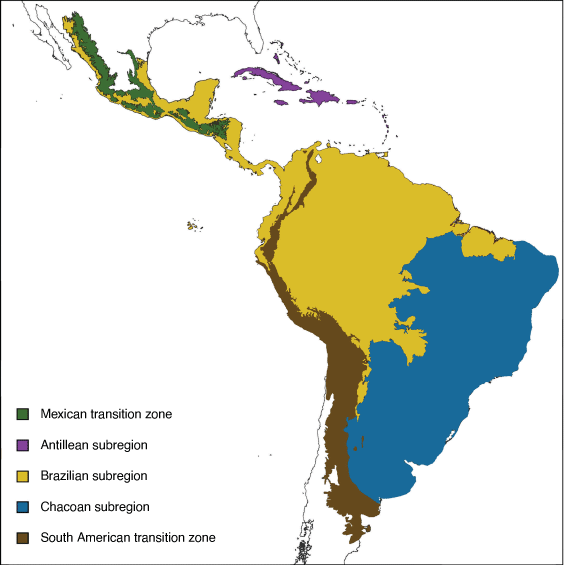
Antillean subregion: Wallace (1876a)
Diagnosis
The Antillean subregion comprises the Antilles or West Indies (Greater and Lesser Antilles) and the Bahamas. The Greater Antilles include Cuba, Jamaica, Hispaniola and Puerto Rico, and the Lesser Antilles include Grenada, The Grenadines, St Vincent, Barbados, St Lucia, Martinique, Dominica, Marie Galante, Guadeloupe, La Desirade, Montserrat, Antigua, Nevis, St Kitts, Barbuda, St Eustatius, Saba, St Barthélemy, St Martin and Anguilla (Morrone 2017).
Brazilian subregion: Blyth (1871)
Diagnosis
The Brazilian subregion comprises central and southern Mexico, Central America, and north-western South America (Morrone 2017).
Chacoan subregion: Cabrera (1951)
Diagnosis
The Chacoan subregion comprises south-eastern South America (Morrone 2017).
Austral kingdom: Engler (1899)
Diagnosis
Southern temperate areas in South America, South Africa, Australasia and Antarctica.
Remarks
It corresponds to Temperate Gondwanaland (Amorim and Tozoni 1994; Ebach et al. 2015; Morrone 2015a, 2018). The Cape region, previously assigned to this kingdom, is here transferred to the Ethiopian region.
Regions
The Austral kingdom comprises three regions, namely Andean, Australian and Antarctic.
Andean region: Engler (1882)
Diagnosis
Southern South America below 30° south latitude.
Remarks
Roig-Juñent et al. (2018) considered that the Patagonian biogeographic province should be considered as belonging to the South American transition zone, but we keep it in the Andean region.
Subregions
The Andean regions comprises three subregions, namely Subantarctic, Central Chilean and Patagonian (Morrone 2015b, 2018; Fig. 20).
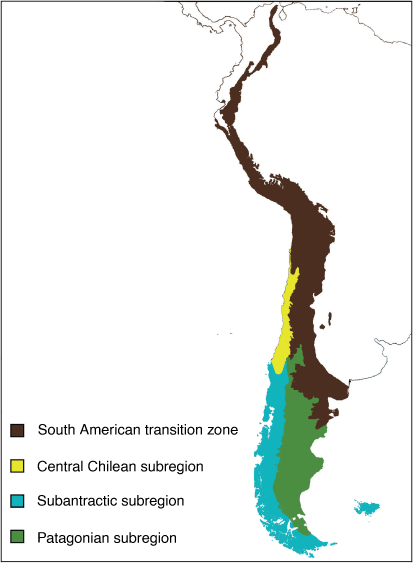
Subantarctic subregion: Skottsberg (1905)
Diagnosis
This subregion represents the core of the Andean region, and corresponds to Austral Chile from 37° south latitude to Cabo de Hornos, the archipelago of southern Chile and Argentina, and the Falklands (Malvinas), South Georgia and Juan Fernández islands (Morrone 2018).
Central Chilean subregion: Hauman (1931)
Diagnosis
Area that corresponds to Central Chile between 26 and 37° south latitude (Morrone 2018).
Patagonian subregion: Lorentz (1876)
Diagnosis
Southern Argentina, from central Mendoza, widening through Neuquén, Río Negro, Chubut and Santa Cruz, to northern Tierra del Fuego; and reaching Chile in Malleco, Aisén and Magallanes provinces (Morrone 2018).
Australian region: Sclater (1858)
Diagnosis
Australian continent, New Guinea and western islands, eastern Sulawesi, southern Philippines, New Caledonia, and New Zealand.
Remarks
Wallace’s original classification is used herein, which includes the southern Phillipines.
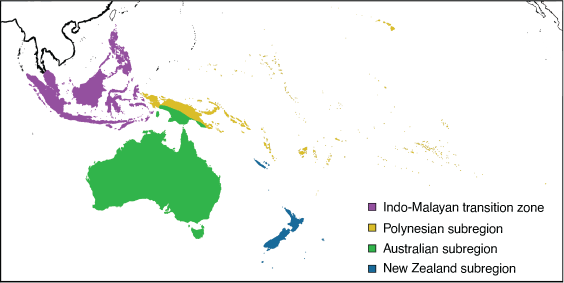
Subregions
The Australian region comprises three subregions, namely Australian, Polynesian and New Zealand (Fig. 21).
|
Australian subregion: Wallace (1876a)
Diagnosis
The Australian continent, including southern New Guinea (south of the Central Range) and Aru Islands Regency.
Polynesian subregion: Wallace (1876a)
Diagnosis
The islands of the tropical Pacific Ocean, from the Pitcairn Islands in the east to the Hawaiian Islands in the north-east, to Palau and Mariana Islands in the north-west, to Illiger’s Line in the west, including Northern Papua, the Central Ranges and the Solomons, to Vanuatu and Austral islands to the south.
Remarks
New Caledonia (Grande Terre and the Loyalty Islands) is not included in the Polynesian subregion because it shares Australian taxa and taxa that are sister to New Zealand. Although there are connections to both the Oriental and the Neotropical regions, the relationship with the Australian biota is significant (Heads 2014). Given that the boundary known as Lydekker’s Line (Lydekker 1896) was proposed by Illiger (1815) 81 years earlier in his Überblick der Säugthiere nach ihrer Vertheilung über die Welttheile [A Review of Mammals based on their Global Distribution], the boundary is renamed herein Illiger’s Line, after the German zoologist Johann Karl Wilhelm Illiger (1775–1813).
New Zealand subregion: Wallace (1876a)
Diagnosis
The area encompassing New Zealand and surrounding islands (Stewart, Lord Howe, Norfolk, Chathams, Campbell) and New Caledonia (see remarks above).
Antarctic region: Blyth (1871)
Diagnosis
Antarctica and surrounding islands.
Remarks
Udvardy (1987) has recognised three provinces within this region, namely Insulantarctica, Marielandia and Maudlandia.
Transition zones
Areas that are defined through the overlap of two different kingdoms are called transition zones (sensu lato Morrone 2015a; sensu stricto Hermogenes de Mendonça and Ebach (2020; Table 5). Transition zones, by their composite nature, have no place in a hierarchal classification; rather they form a separate non-hierarchal classification detailed below.

|
Mexican transition zone: Grisebach (1872)
Diagnosis
Mountainous areas of Mexico, Guatemala, Honduras, El Salvador and Nicaragua north of lake Nicaragua (Morrone 2014, 2015a; Halffter and Morrone 2017). It corresponds with the boundary between the Nearctic and Neotropical regions, and comprises the Sierra Madre Occidental, Sierra Madre Oriental, Sierra Madre del Sur, Transmexican Volcanic Belt and Chiapas highlands (Morrone 2014, 2015a).
Remarks
Halffter (1987) considered that the Mexican transition zone extends to southern United States as well as the Mexican lowlands.
Chinese transition zone: Sclater and Sclater (1899)
Diagnosis
It corresponds to the boundary between the Palearctic and Oriental regions (Palestrini et al. 1985; Müller 1986; Kreft and Jetz 2013).
Remarks
Müller (1979) suggested that this zone extends from the Yang Tsê–Kiang River to the 21st parallel, including also Taiwan. Palestrini et al. (1985) analysed the geographical distribution of some groups of Scarabaeoidea (Coleoptera) of this area and detected the overlap of Palearctic, Oriental and Sino-Japanese cenocrons.
Saharo-Arabian transition zone: Holt et al. (2013a)
Diagnosis
It comprises the Sahara Desert and the Arabian Peninsula (Müller 1986; Kreft and Jetz 2013). It corresponds to the boundary between the Palearctic and Ethiopian regions.
South American transition zone: Morrone (2004)
Diagnosis
It comprises the Andean highlands between western Venezuela and northern Chile and central western Argentina (Morrone 2006, 2014; Martínez et al. 2016). It corresponds to the boundary between the Neotropical and Andean regions, which was analysed by Rapoport (1968), who discussed the alternative placements given by different authors to the ‘subtropical line’ that separates these regions.
Remarks
Urtubey et al. (2010) analysed the distribution of some Asteraceae within this transition zone. Escalante (2017) recovered this transition zone in an endemicity analysis of the terrestrial mammals of the world and noted its close relationship with the Andean region. More recently, Roig-Juñent et al. (2018) considered that the Patagonian biogeographic province should be considered as belonging to the South American Transition Zone instead of the Andean region in the strict sense.
Indo-Malayan transition zone: Wallace (1876a)
Diagnosis
It corresponds to the boundary between the Oriental and Australian regions (Illiger’s Line in the east).
Remarks
Müller (1986) discussed its boundaries and gave examples of Oriental and Australian taxa overlapping in this transition zone. Michaux (2010) analysed the geological development of this area, identified areas of endemism and concluded that the latter are linked to geological processes resulting from the interaction between the Eurasian and Australian continents, and the Philippine Sea Plate. King and Ebach (2017) showed that it is a temporally composite area. Michaux (2019) undertook a parsimony analysis of endemicity, finding that the areas assigned to this transition zone may constitute a natural area if the Philippines are included, as proposed previously by Dickerson et al. (1928).
Data availability
Data sharing is not applicable as no new data were generated or analysed during this study.
Conflicts of interest
Malte Ebach is an Associate Editor of Australian Systematic Botany but did not at any stage have editor-level access to this manuscript while in peer review, as is the standard practice when handling manuscripts submitted by an editor to this journal. Australian Systematic Botany encourages its editors to publish in the journal and they are kept totally separate from the decision-making processes for their manuscripts. The authors have no further conflicts of interest to declare.
Declaration of funding
This research did not receive any specific funding.
Acknowledgements
We acknowledge the valuable comments by three anonymous reviewers.
References
Acosta LE, Maury EA (1998a) Scorpiones. In ‘Biodiversidad de artrópodos argentinos: un enfoque biotaxonómico’. (Eds JJ Morrone, S Coscarón) pp. 545–559. (Ediciones Sur: La Plata, Argentina)Acosta LE, Maury EA (1998b) Opiliones. In ‘ Biodiversidad de artrópodos argentinos: un enfoque biotaxonómico’. (Eds JJ Morrone, S Coscarón) pp. 569–580. (Ediciones Sur: La Plata, Argentina)
Allen JA (1871) On the mammals and winter birds of east Florida, with an examination of certain assumed specific characters in birds, and a sketch of the bird-faunae of eastern North America. Bulletin of the Museum of Comparative Zoology at Harvard College 2, 161–450.
Allen JA (1892) The geographical distribution of North American mammals. Bulletin of the American Museum of Natural History 4, 199–243.
Allen JA (1893) The geographical origin and distribution of North American birds, considered in relation to faunal areas of North America. Auk 10, 97–150.
| The geographical origin and distribution of North American birds, considered in relation to faunal areas of North America.Crossref | GoogleScholarGoogle Scholar |
Almirón A, Azpelicueta M, Casciotta J, López Cazorla A (1997) Ichthyogeographic boundary between the Brazilian and Austral subregions in South America. Argentina. Biogeographica 73, 23–30.
Amorim DS, Pires MRS (1996) Neotropical biogeography and a method for maximum biodiversity estimation. In ‘Biodiversity in Brazil: a first approach’. (Eds CEM Bicudo, NA Menezes) pp. 183–219. (CNPq: São Paulo, Brazil)
Amorim DS, Tozoni SHS (1994) Phylogenetic and biogeographic analysis of the Anisopodoidea (Diptera, Bibionomorpha), with an area cladogram for intercontinental relationships. Revista Brasileira de Entomologia 38, 517–543.
Apodaca MJ, Crisci JV, Katinas L (2015a) Las provincias fitogeográficas de la República Argentina: definición y sus principales áreas protegidas. In ‘El deterioro del suelo y del ambiente en la Argentina’. (Eds RR Casas, GF Albarracín) (Fundación Ciencia, Educación y Cultura: Buenos Aires, Argentina)
Apodaca MJ, Crisci JV, Katinas L (2015b) Andean origin and diversification of the genus Perezia, an ancient lineage of Asteraceae. Smithsonian Contributions to Botany 102, 1–28.
Arana MD, Morrone JJ, Oggero AJ (2011) Licofitas (Tracheophyta: Lycopodiophyta) de las Sierras Centrales de Argentina: un enfoque panbiogeográfico. Gayana. Botánica 68, 16–21.
| Licofitas (Tracheophyta: Lycopodiophyta) de las Sierras Centrales de Argentina: un enfoque panbiogeográfico.Crossref | GoogleScholarGoogle Scholar |
Arana MD, Martínez GA, Oggero AJ, Natale ES, Morrone JJ (2017) Map and shapefile of the biogeographic provinces of Argentina. Zootaxa 4341, 420–422.
| Map and shapefile of the biogeographic provinces of Argentina.Crossref | GoogleScholarGoogle Scholar | 29245664PubMed |
Arldt T (1906) Die tiergeographischen Reiche und Regionen. Geographische Zeitschrift 12, 212–222.
Arldt T (1907) ‘Die Entwicklung der Kontinente und ihrer Lebewelt: Ein Beitrag zur vergleichenden Erdgeschichte.’ (W. Engelmann: Leipzig, German Empire)
Artigas JN (1975) Introducción al estudio por computación de las áreas zoogeográficas de Chile continental basado en la distribución de 903 especies de animales terrestres. Gayana. Miscelánea 4, 1–25.
| Introducción al estudio por computación de las áreas zoogeográficas de Chile continental basado en la distribución de 903 especies de animales terrestres.Crossref | GoogleScholarGoogle Scholar |
Bănărescu P (1975) ‘Principles and problems of zoogeography.’ (Nolit: Belgrade, SFR Yugoslavia)
Bănărescu P, Boşcaiu N (1978) ‘Biogeographie: Fauna und Flora der Erde und ihre geschichtliche Entwicklung.’ (Veb Gustav Fischer Verlag: Jena, German Democratic Republic)
Baranzelli MC, Johnson LA, Cosacov A, Sérsic AN (2014) Historial and ecological divergence among populations of Monttea chilensis (Plantaginaceae), an endemic endangered shrub bordering the Atacama Desert, Chile. Evolutionary Ecology 28, 751–774.
| Historial and ecological divergence among populations of Monttea chilensis (Plantaginaceae), an endemic endangered shrub bordering the Atacama Desert, Chile.Crossref | GoogleScholarGoogle Scholar |
Bartholomew JG, Clark WE, Grimshaw PH (1911) ‘Atlas of zoogeography: a series of maps illustrating the distribution of over seven hundred families, genera, and species of existing animals.’ (Edinburgh Geographical Institute: Edinburgh, UK)
Beddard FE (1895) ‘A text‐book of zoogeography.’ (Clay and Son: London, UK)
Berg LS (1916) ‘Les Poissons des Eaux douces de la Russie’. (Department of Agriculture: Moscow, USSR) [In Russian]
Beron P (2018) ‘Zoogeography of Arachnida.’ (Springer: Dordrecht, Netherlands)
Blanford WT (1890) Anniversary address. Proceedings of the Annual Meeting of the Geological Society of London 1890, 13–80.
Blyth E (1871) A suggested new division of the Earth into zoological regions. Nature 3, 427–429.
| A suggested new division of the Earth into zoological regions.Crossref | GoogleScholarGoogle Scholar |
Bobrinskiy NA, Zenkevich LA, Birstein YA (1946) ‘Geography of animals.’ (Sov. Nauka: Moscow, USSR)
Born J, Linder HP, Desmer P (2007) The Greater Cape floristic region. Journal of Biogeography 34, 147–162.
| The Greater Cape floristic region.Crossref | GoogleScholarGoogle Scholar |
Brown DE, Reichenbacher F, Franson SE (1998) ‘A classification of North American biotic communities.’ (The University of Utah Press: Salt Lake City, UT, USA)
Burkart R, Bárbaro NO, Sánchez RO, Gómez DA (1999) ‘Eco-regiones de la Argentina.’ (Administración de Parques Nacionales: Buenos Aires, Argentina)
Cabrera AL (1951) Territorios fitogeográficos de la República Argentina. Boletín de la Sociedad Argentina de Botánica 4, 21–65.
Cabrera AL (1971) Fitogeografía de la República Argentina. Boletín de la Sociedad Argentina de Botánica 14, 1–42.
Cabrera AL, Willink A (1973) ‘Biogeografía de América Latina. Monografía 13, Serie de Biología.’ (OEA: Washington, DC, USA)
Cabrera A, Yepes J (1940) ‘Mamíferos sud-americanos (vida, costumbres y descripción).’ (Historia Natural Ediar: Buenos Aires, Argentina)
Cano E, Ramírez AV, Cano-Ortiz A, Esteban Ruiz FJE (2009) Distribution of Central American Melastomataceae: biogeographical analysis of the Caribbean islands. Acta Botanica Gallica 156, 527–557.
| Distribution of Central American Melastomataceae: biogeographical analysis of the Caribbean islands.Crossref | GoogleScholarGoogle Scholar |
Carta A, Peruzzi L, Ramırez-Barahona S (2022) A global phylogenetic regionalization of vascular plants reveals a deep split between Gondwanan and Laurasian biotas. New Phytologist 233, 1494–1504.
| A global phylogenetic regionalization of vascular plants reveals a deep split between Gondwanan and Laurasian biotas.Crossref | GoogleScholarGoogle Scholar | 34758121PubMed |
Cekalovic T (1974) Divisiones biogeográficas de la XII Región Chilena (Magallanes). Boletín de la Sociedad de Biología de Concepción 48, 297–314.
Clarke CB (1892) On biologic regions and tabulation areas. Philosophical Transactions of the Royal Society of London 183, 371–387.
| On biologic regions and tabulation areas.Crossref | GoogleScholarGoogle Scholar |
Conran JG (1995) Distributions in the Liliiflorae and their biogeographical implications. Journal of Biogeography 22, 1023–1034.
Cooper JG (1859) On the distribution of the forests and trees of North America, with notes on its physical geography. Annual Report of the Board of Regents of the Smithsonian Institution, Showing the Operations, Expenditures, and Condition of the Institution for the Year 1858, 246–280.
Cope ED (1873) Zoological description. In ‘Gray’s Atlas of the United States, with General Maps of the World, Accompanied by Descriptions Geographical, Historical, Scientific and Statical’. (Ed. OW Gray) pp. 32–36. (Stedman Brown and Lyon: Philadelphia, PA, USA)
Cope ED (1875) Check-list of North American Batrachia and Reptilia. Part III. On geographical distribution of the Vertebrata of the Regnum Nearcticum. Bulletin of the United States National Museum 1, 55–100.
Corona A, Morrone JJ (2005) Track analysis of the species of Lampetis (Spinthoptera) Casey, 1909 (Coleoptera: Buprestidae) in North America, Central America, and the West Indies. Caribbean Journal of Science 41, 37–41.
Coscarón S, Coscarón-Arias CL (1995) Distribution of Neotropical Simuliidae (Insecta, Diptera) and its areas of endemism. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales 19, 717–732.
Cowles HC (1908) An ecological aspect of the conception of species. The American Naturalist 42, 265–271.
Cox CBC (2001) The biogeographic regions reconsidered. Journal of Biogeography 28, 511–523.
| The biogeographic regions reconsidered.Crossref | GoogleScholarGoogle Scholar |
Cracraft J (1985) Historical biogeography and patterns of differentiation within the South American avifauna: areas of endemism. Ornithological Monographs 36, 49–84.
| Historical biogeography and patterns of differentiation within the South American avifauna: areas of endemism.Crossref | GoogleScholarGoogle Scholar |
Daniels LD, Veblen TV (2000) ENSO effects on temperature and precipitation of the Patagonian–Andean region: implications for biogeography. Physical Geography 21, 223–243.
| ENSO effects on temperature and precipitation of the Patagonian–Andean region: implications for biogeography.Crossref | GoogleScholarGoogle Scholar |
Darlington PJ Jr (1957) ‘Zoogeography: the geographical distribution of animals.’ (Wiley: New York, NY, USA)
de Candolle AP (1805) Explication de la carte Botanique de la France. In ‘Flore française, ou descriptions succinctes de toutes les plantes qui croissent naturellement en France, disposées selon une nouvelle méthode d’analyse, et précédées par un exposé des principes élémentaires de la botanique’. (Eds JBPAM Lamarck, AP de Candolle) (Desray: Paris, France)
de Candolle AP (1820) Géographie botanique. In ‘Dictionnaire des Sciences Naturelles’. (Ed. FG Levrault) pp. 359–422. (Imprimeur du Roi)
de Lattin G (1967) ‘Grundriss der Zoogeographie.’ (Gustav Fischer Verlag: Jena, German Democratic Republic)
de Mello-Leitão C 1937 ‘Zoo-geografia do Brasil.’ (Biblioteca Pedagógica Brasileira, Brasiliana: São Paulo, Brazil)
de Mello-Leitão C (1943) Los alacranes y la zoogeografía de Sudamérica. Revista Argentina de Zoogeografía 2, 125–131.
de Paggi SJ (1990) Ecological and biogeographical remarks on the rotifer fauna of Argentina. Revue d’Hydrobiologie Tropicale 23, 297–311.
Del Risco E, Vandama A (1989) ‘Regionalización florística.’ (Nuevo Atlas Nacional de Cuba, Instituto Cubano de Geodesia y Cartografía and Instituto Geográfico Nacional de España: Madrid, Spain)
Delachaux EAS (1920) Las regiones físicas de la República Argentina. Revista del Museo de La Plata 15, 102–131.
Dickerson RE, Merrill ED, McGregor RC, Schultze W, Taylor EH, Herre AW (1928) Distribution of life in the Philippines. Manila Bureau of Science Monographs 21, 1–322.
Diels L (1908) ‘Pflanzengeographie.’ (Göschensche Verlagshandlung: Leipzig)
Dinerstein E, Olson DM, Graham DJ, Webster AL, Primm SA, Bookbinder MP, Ledec G (1995) ‘A conservation assessment of the terrestrial ecoregions of Latin America and the Caribbean.’ (The World Bank: Washington, DC, USA)
Doing H (1970) Botanical geography and chorology in Australia. Belmontia (Miscellaneous Papers) 13, 81–88.
Donato M (2006) Historical biogeography of the family Tristiridae (Orthoptera: Acridomorpha) applying dispersal–vicariance analysis. Journal of Arid Environments 66, 421–434.
| Historical biogeography of the family Tristiridae (Orthoptera: Acridomorpha) applying dispersal–vicariance analysis.Crossref | GoogleScholarGoogle Scholar |
Dowding EM, Ebach MC (2018) An interim global bioregionalisation of Devonian areas. Palaeobiodiversity and Palaeoenvironments 98, 527–547.
| An interim global bioregionalisation of Devonian areas.Crossref | GoogleScholarGoogle Scholar |
Drude O (1890) ‘Handbuch der Pflanzengeographie.’ (Bibliothek Geographischer Handbücher, Verlag J. Engelhorn: Stuttgart, German Empire)
Ebach MC (2015) ‘Origins of biogeography.’ (Springer: Dordrecht, Netherlands)
Ebach MC (2021) Origins of biogeography: a personal perspective. In ‘An integrative approach of the evolution of living’. (Ed. E Guilbert) pp. 1–26. (ISTE Science Publishing: London, UK)
Ebach MC, Michaux B (2017) Establishing a framework for a natural area taxonomy. Acta Biotheoretica 2017, 1–11.
Ebach MC, Morrone JJ, Parenti LR, Viloria AL (2008) International Code of Area Nomenclature. Journal of Biogeography 35, 1153–1157.
| International Code of Area Nomenclature.Crossref | GoogleScholarGoogle Scholar |
Ebach MC, Murphy DJ, González-Orozco CE, Miller JT (2015) A revised area taxonomy of phytogeographical regions within the Australian Bioregionalization Atlas. Phytotaxa 208, 261–277.
| A revised area taxonomy of phytogeographical regions within the Australian Bioregionalization Atlas.Crossref | GoogleScholarGoogle Scholar |
Echeverry A, Morrone JJ (2013) Generalised tracks, area cladograms and tectonics in the Caribbean. Journal of Biogeography 40, 1619–1637.
| Generalised tracks, area cladograms and tectonics in the Caribbean.Crossref | GoogleScholarGoogle Scholar |
Egerton FN (2012) ‘Roots of ecology: antiquity to Haeckel.’ (University of California Press: Berkeley, CA, USA)
Engler A (1882) ‘Versuch einer Entwicklungsgeschichte der Pflanzenwelt, insbesondere der Florengebiete seit der Tertiärperiode. Vol. 2. Die extratropischen Gebiete der Südlichen Hemisphäre und die tropischen Gebiete.’ (Verlag von W. Engelmann: Leipzig, German Empire)
Engler A (1899) ‘Die Entwicklung der Pflanzengeographie in den letzten hundert Jahren und weitere Aufgaben derselben: Wissenschaftliche Beiträge zum Gedächtnis der hundertjährigen Wiederkehr des Antritts von Alexander von Humboldt’s Reise nach Amerika.’ (Gesellschaft für Erdkunde zu Berlin: Berlin, German Empire)
Escalante T (2017) A natural regionalization of the world based on primary biogeographic homology of terrestrial mammals. Biological Journal of the Linnean Society. Linnean Society of London 120, 349–362.
Escalante T, Morrone JJ, Rodríguez-Tapia G (2013) Biogeographic regions of North American mammals based on endemism. Biological Journal of the Linnean Society. Linnean Society of London 110, 485–499.
| Biogeographic regions of North American mammals based on endemism.Crossref | GoogleScholarGoogle Scholar |
Escalante T, Rodríguez-Tapia G, Morrone JJ (2021) Toward a biogeographic regionalization of the Nearctic region: area nomenclature and digital map. Zootaxa 5027, 351–375.
| Toward a biogeographic regionalization of the Nearctic region: area nomenclature and digital map.Crossref | GoogleScholarGoogle Scholar | 34811166PubMed |
Espinosa Organista D, Ocegueda Cruz S, Aguilar Zúñiga C, Flores Villela O, Llorente-Bousquets J (2008) El conocimiento biogeográfico de las especies y su regionalización natural. In ‘Capital natural de México, Conocimiento actual de la biodiversidad, vol I’. (Ed. J Sarukhán) pp. 33–65. (Conabio: Mexico City, Mexico)
Fabricius JC (1778) ‘Philosophia Entomologica.’ (Impensis Carol. Ernest. Bohnii: Hansestadt Hamburg)
Fiaschi P, Pirani JR (2009) Review of plant biogeographic studies in Brazil. Journal of Systematics and Evolution 47, 477–496.
| Review of plant biogeographic studies in Brazil.Crossref | GoogleScholarGoogle Scholar |
Fittkau EJ (1969) The fauna of South America. In ‘Biogeography and ecology in South America, 2’. (Eds E Fittkau, JJ Illies, H Klinge, GH Schwabe, H Sioli) pp. 624–650. (Junk: The Hague, Netherlands)
Fleming CA (1987) Comments on Udvardy’s biogeographical realm Antarctica. Journal of the Royal Society of New Zealand 17, 195–200.
| Comments on Udvardy’s biogeographical realm Antarctica.Crossref | GoogleScholarGoogle Scholar |
Flint OS (1989) Studies of Neotropical caddisflies, XXXIX: the genus Smicridea in the Chilean subregion (Trichoptera: Hydropsychidae). Smithsonian Contributions to Zoology 472, 1–45.
| Studies of Neotropical caddisflies, XXXIX: the genus Smicridea in the Chilean subregion (Trichoptera: Hydropsychidae).Crossref | GoogleScholarGoogle Scholar |
Forbes E (1846) On the connexion between the distribution of the existing fauna and flora of the British Isles and the geological changes which have affected their area, especially during the epoch of the Northern Drift. Memoirs of the Geological Survey of Great Britain 1, 336–432.
Forsyth Major CJ (1884) Zoogeographische Übergangsregionen. Kosmos (Stockholm) 14, 102–113.
Galley C, Linder HP (2006) Geographical affinities of the Cape flora, South Africa. Journal of Biogeography 33, 236–250.
| Geographical affinities of the Cape flora, South Africa.Crossref | GoogleScholarGoogle Scholar |
Galley C, Bytebier B, Bellstedt DU, Linder HP (2007) The Cape element in the Afrotemperate flora: from Cape to Cairo? Proceedings of the Royal Society of London – B. Biological Sciences 274, 535–543.
| The Cape element in the Afrotemperate flora: from Cape to Cairo?Crossref | GoogleScholarGoogle Scholar |
Gill T T (1885) The principles of zoogeography. Proceedings of the Biological Society of London 2, 1–39.
Glasby CJ (2005) Polychaete distribution patterns revisited: An historical explanation. Marine Ecology 26, 235–245.
| Polychaete distribution patterns revisited: An historical explanation.Crossref | GoogleScholarGoogle Scholar |
Goetsch W (1931) Estudios sobre zoogeografía chilena. Boletín de la Sociedad de Biología de Concepción 5, 1–19.
Good R (1947) ‘The geography of the flowering plants.’ (Longman: London, UK)
Good R (1963) ‘The geography of the flowering plants’, 3rd edn. (Longman: London, UK)
Grisebach A (1872) ‘Die Vegetation der Erde nach ihrer klimatischen Anordnung: Ein Abriss der vergleichenden Geographie der Pflanzen.’ (Verlag von Wilhelm Engelmann: Leipzig, German Empire)
Hagmeier EM (1966) A numerical analysis of distributional patterns of North America mammals, II. Re-evaluation of the provinces. Systematic Zoology 15, 279–299.
| A numerical analysis of distributional patterns of North America mammals, II. Re-evaluation of the provinces.Crossref | GoogleScholarGoogle Scholar |
Hagmeier EM, Stults CD (1964) A numerical analysis of distributional patterns of North America mammals. Systematic Zoology 13, 125–155.
| A numerical analysis of distributional patterns of North America mammals.Crossref | GoogleScholarGoogle Scholar |
Halffter G (1987) Biogeography of the montane entomofauna of Mexico and Central America. Annual Review of Entomology 32, 95–114.
| Biogeography of the montane entomofauna of Mexico and Central America.Crossref | GoogleScholarGoogle Scholar |
Halffter G, Morrone JJ (2017) An analytical review of Halffter’s Mexican transition zone, and its relevance for evolutionary biogeography, ecology and biogeographic regionalization. Zootaxa 4226, 1–46.
| An analytical review of Halffter’s Mexican transition zone, and its relevance for evolutionary biogeography, ecology and biogeographic regionalization.Crossref | GoogleScholarGoogle Scholar |
Hauman L (1920) Ganadería y geobotánica en la Argentina. Revista del Centro de Estudiantes de Agronomía y Veterinaria de la Universidad de Buenos Aires 102, 45–65.
Hauman L (1931) Esquisse phytogéographique de l’Argentine subtropicale et de ses relations avec la géobotanique sud-américaine. Bulletin de la Société Royale de Botanique de Belgique 64, 20–79.
Heads M (2014) ‘Biogeography of Australasia: a molecular analysis.’ (Cambridge University Press)
Heilprin A (1882) On the value of the “Nearctic” as one of the primary zoological regions. Proceedings of the Academy of Natural Sciences of Philadelphia 34, 316–334.
Heilprin A (1883) On the value of the ‘Neoarctic’ [sic] as one of the primary zoological regions. Nature 27, 606
| On the value of the ‘Neoarctic’ [sic] as one of the primary zoological regions.Crossref | GoogleScholarGoogle Scholar |
Heilprin A (1887) ‘The geographical and geological distribution of animals.’ (D. Appleton and Company: New York, NY, USA)
Hemsley WB (1887). Botany, Vol. IV. In ‘Biologia Centrali-Americana or, contributions to the knowledge of the fauna and flora of Mexico and Central America’. (Eds ED Godman, O Salvin) pp. 1–448. (London, UK)
Hermogenes de Mendonça L, Ebach MC (2020) A review of transition zones in biogeographical classification. Biological Journal of the Linnean Society 131, 717–736.
| A review of transition zones in biogeographical classification.Crossref | GoogleScholarGoogle Scholar |
Hershkovitz P (1958) A geographic classification of neotropical mammals. Fieldiana. Zoology 36, 1–646.
Hershkovitz P (1969) The evolution of mammals on southern continents. VI: The Recent mammals of the Neotropical region: a zoogeographic and ecological review. The Quarterly Review of Biology 44, 1–70.
| The evolution of mammals on southern continents. VI: The Recent mammals of the Neotropical region: a zoogeographic and ecological review.Crossref | GoogleScholarGoogle Scholar |
Holdgate MW (1961) Vegetation and soils in the south Chilean islands. Journal of Ecology 49, 559–580.
| Vegetation and soils in the south Chilean islands.Crossref | GoogleScholarGoogle Scholar |
Holt BG, Lessard JP, Borregaard MK, Fritz SA, Araújo MB, Dimitrov D, Fabre PH, Graham CH, Graves GR, Jønsson KA, Nogués-Bravo D, Wang Z, Whittaker RJ, Fjeldså RJ, Rahbek C (2013a) An update of Wallace’s zoogeographic regions of the world. Science 339, 74–78.
| An update of Wallace’s zoogeographic regions of the world.Crossref | GoogleScholarGoogle Scholar | 23258408PubMed |
Holt BG, Lessard JP, Borregaard MK, Fritz SA, Araújo MB, Dimitrov D, Fabre PH, Graham CH, Graves GR, Jønsson KA, Nogués-Bravo D, Wang Z, Whittaker RJ, Fjeldså RJ, Rahbek C (2013b) Response to comment on ‘An update of Wallace’s zoogeographic regions of the world’. Science 341, 343-d
| Response to comment on ‘An update of Wallace’s zoogeographic regions of the world’.Crossref | GoogleScholarGoogle Scholar |
Huang C, Ebach MC, Ahyong ST (2020) Bioregionalisation of the freshwater zoogeographical areas of mainland China. Zootaxa 4742, 271–298.
| Bioregionalisation of the freshwater zoogeographical areas of mainland China.Crossref | GoogleScholarGoogle Scholar |
Huber O, Riina R (1997) ‘Glosario ecológico de las Américas. Vol. I. México, América Central e islas del Caribe: Países hispanoparlantes.’ (UNESCO: Paris, France)
Huber O, Riina R (2003) ‘Glosario ecológico de las Américas. Vol. 2. América del Sur: Países hispanoparlantes.’ (UNESCO: Caracas, Venezuela)
Hueck K (1957) Las regiones forestales de Sudamérica. Boletín del Instituto Forestal Latinoamericano de Investigación y Capacitación (Mérida) 2, 1–40.
Hueck K (1966) ‘Die Wälder Südamerikas.’ (Fischer: Stuttgart, Federal Republic of Germany)
Huxley TH (1868) On the classification and distribution of Alectoromorphae and Heteromorphae. Proceedings of the Zoological Society of London 1868, 294–319.
Illies J (1968) Buch Besprechung. Beitrage zur Neotropischen Fauna 5, 229–234.
Illiger JKW (1815) Überblick der Säugthiere nach ihrer Vertheilung über die Welttheile. Abhandlungen der physikalische Klasse der Koeniglich-Preussischen Akademie der Wissenschaften 1804–1811, 39–159.
Kang B, Deng J, Wu Y, Chen L, Zhang J, Qiu H, Lu Y, He D (2014) Mapping China’s freshwater fishes: diversity and biogeography. Fish and Fisheries 15, 209–230.
| Mapping China’s freshwater fishes: diversity and biogeography.Crossref | GoogleScholarGoogle Scholar |
Kendeigh SC (1954) History and evaluation of various concepts of plant and animal communities in North America. Ecology 35, 152–171.
| History and evaluation of various concepts of plant and animal communities in North America.Crossref | GoogleScholarGoogle Scholar |
King AR, Ebach MC (2017) A novel approach to time-slicing areas within biogeographic-area classifications: Wallacea as an example. Australian Systematic Botany 30, 495–512.
| A novel approach to time-slicing areas within biogeographic-area classifications: Wallacea as an example.Crossref | GoogleScholarGoogle Scholar |
Kirby W, Spence W (1826) ‘Introduction to entomology.’ (Longman, Rees, Orme, Brown and Green: London, UK)
Kirby WF (1872) On the geographical distribution of the diurnal Lepidoptera as compared with that of the birds. Journal of the Linnean Society of London 11, 431–439.
| On the geographical distribution of the diurnal Lepidoptera as compared with that of the birds.Crossref | GoogleScholarGoogle Scholar |
Klassa B, Santos CMD (2015) Areas of endemism in the Neotropical region based on the geographical distribution of Tabanomorpha (Diptera: Brachycera). Zootaxa 4058, 519–534.
| Areas of endemism in the Neotropical region based on the geographical distribution of Tabanomorpha (Diptera: Brachycera).Crossref | GoogleScholarGoogle Scholar | 26701543PubMed |
Kreft H, Jetz W (2010) A framework for delineating biogeographical regions based on species distributions. Journal of Biogeography 37, 2029–2053.
| A framework for delineating biogeographical regions based on species distributions.Crossref | GoogleScholarGoogle Scholar |
Kreft H, Jetz W (2013) Comment on ‘An update of Wallace’s zoogeographic regions of the world’. Science 341, 343-c
| Comment on ‘An update of Wallace’s zoogeographic regions of the world’.Crossref | GoogleScholarGoogle Scholar |
Kuschel G (1960) Terrestrial zoology in southern Chile. Proceedings of the Royal Society of London – B. Biological Sciences 152, 540–550.
| Terrestrial zoology in southern Chile.Crossref | GoogleScholarGoogle Scholar |
Kuschel G (1963) Problems concerning an Austral region. In ‘Pacific basin biogeography: a symposium: Tenth Pacific Science Congress’, 21 August–6 September 1961, Honolulu, HI, USA. (Eds JL Gressitt, CH Lindroth, FR Fosberg, CA Fleming, EG Turbott) pp. 443–449. (Bishop Museum Press: Honolulu, HI, USA)
Kuschel G (1969) Biogeography and ecology of South American Coleoptera. In ‘Biogeography and ecology in South America, 2’. (Eds E Fittkau, JJ Illies, H Klinge, GH Schwabe, H Sioli) pp. 709–722. (Junk: The Hague, Netherlands)
Lamas CJE, Nihei SS, Cunha AM, Couri MS (2014) Phylogeny and biogeography of Heterostylum (Diptera: Bombyliidae): Evidence for an ancient Caribbean diversification model. The Florida Entomologist 97, 952–966.
| Phylogeny and biogeography of Heterostylum (Diptera: Bombyliidae): Evidence for an ancient Caribbean diversification model.Crossref | GoogleScholarGoogle Scholar |
Latreille PA (1815) Introduction à la géographie générale des arachnides et des insectes; ou des climats propres à ces animaux. Mémoire lu à l’Académie des Sciences 3, 37–67.
LeConte JL (1859) ‘The Coleoptera of Kansas and eastern New Mexico. Smithsonian Contributions to Knowledge.’ (The Smithsonian Institution: New York, NY, USA)
Li SZ (1981) ‘Distribution division of Chinese freshwater fishes.’ (Science Press: Beijing, PR China) [In Chinese]
Linder HP (1994) Afrotemperate phytogeography: Implications of cladistic biogeographical analysis. In ‘Proceedings of the XIIIth Plenary Meeting AETFAT’, 2–11 April 1991, Zomba, Malawi. (Eds JH Seyani, AC Chikuni) pp. 913–930. (National Herbarium and Botanic Gardens of Malawi)
Locklin C (2017) Southeastern South America: Central Chile. In ‘Terrestrial Ecoregions of Latin America and the Caribbean’. WWF NT1201. (World Wildlife Fund)
Lopatin IK (1980) ‘Osnovy zoogeografii [Fundamentals of zoogeography].’ (Visheyshaya Shkola: Minsk, USSR) [In Russian]
Lorentz PG (1876) Cuadro de la vegetación de la República Argentina. In ‘La República Argentina’. (Ed. R Napp) pp. 77–136. (Sociedad Anónima de Tipografía, Litografía y Fundición de Tipos: Buenos Aires, Argentina)
Lücking R (2003) Takhtajan’s floristic regions and foliicolous lichen biogeography: a compatibility analysis. Lichenologist 35, 33–53.
| Takhtajan’s floristic regions and foliicolous lichen biogeography: a compatibility analysis.Crossref | GoogleScholarGoogle Scholar |
Lydekker BA (1896) ‘A geographical history of mammals.’ (Cambridge University Press)
Martínez GA, Arana MD, Oggero AJ, Natale ES (2016) Biogeographic relationships and new regionalisation of high-altitude grasslands and woodlands of the central Pampean Ranges (Argentina), based on vascular plants and vertebrates. Australian Systematic Botany 29, 473–488.
| Biogeographic relationships and new regionalisation of high-altitude grasslands and woodlands of the central Pampean Ranges (Argentina), based on vascular plants and vertebrates.Crossref | GoogleScholarGoogle Scholar |
Merriam CH (1892) The geographic distribution of life in North America with special reference to the Mammalia. Proceedings of the Biological Society of Washington 7, 1–64.
Meyen FJF (1846) ‘Outlines of the geography of plants: with particular enquiries concerning the native country, the culture, and the uses of the principal cultivated plants on which the prosperity of nations is based.’ (Ray Society: London, UK)
Michaux B (2010) Biogeography of Wallacea: Geotectonic models, areas of endemism, and natural biogeographic units. Biological Journal of the Linnean Society. Linnean Society of London 101, 193–212.
| Biogeography of Wallacea: Geotectonic models, areas of endemism, and natural biogeographic units.Crossref | GoogleScholarGoogle Scholar |
Michaux B (2019) ‘Biogeology: evolution in a changing landscape.’ (CRC Press: Boca Raton, FL, USA)
Monrós F (1958) Consideraciones sobre la fauna del sur de Chile y revisión de la tribus Stenomelini (Coleoptera, Chrysomelidae). Acta Zoológica Lilloana 15, 143–153.
Morain SA (1984) ‘Systematic and regional biogeography.’ (Van Nostrand Reinhold Company: New York, NY, USA)
Moreira GRP, Ferrari A, Mondin CA, Cervi AC (2011) Panbiogeographical analysis of passion vines at their southern limit of distribution in the Neotropics. Revista Brasileira de Biociências 9, 28–40.
Moreira-Muñoz A (2007) The Austral floristic realm revisited. Journal of Biogeography 34, 1649–1660.
| The Austral floristic realm revisited.Crossref | GoogleScholarGoogle Scholar |
Moreira-Muñoz A (2014) Central Chile ecoregion. In ‘Endemism in vascular plants’. (Ed. C Hobohm) pp. 221–233. (Springer Science: Dordrecht, Netherlands)
Morrone JJ (1996) The biogeographical Andean subregion: a proposal exemplified by Arthropod taxa (Arachnida, Crustacea, and Hexapoda). Neotrópica 42, 103–114.
Morrone JJ (1999) Presentación preliminar de un nuevo esquema biogeográfico de América del Sur. Biogeographica 75, 1–16.
Morrone JJ (2001a) Homology, biogeography and areas of endemism. Diversity & Distributions 7, 297–300.
| Homology, biogeography and areas of endemism.Crossref | GoogleScholarGoogle Scholar |
Morrone JJ (2001b) ‘Biogeografía de América Latina y el Caribe. Vol. 3.’ (M&T-Manuales and Tesis SEA, Sociedad Entomológica Aragonesa: Zaragoza, Spain)
Morrone JJ (2002) Biogeographic regions under track and cladistic scrutiny. Journal of Biogeography 29, 149–152.
| Biogeographic regions under track and cladistic scrutiny.Crossref | GoogleScholarGoogle Scholar |
Morrone JJ (2004a) Panbiogeografía, componentes bióticos y zonas de transición. Revista Brasileira de Entomologia 48, 149–162.
| Panbiogeografía, componentes bióticos y zonas de transición.Crossref | GoogleScholarGoogle Scholar |
Morrone JJ (2004b) La zona de transición Sudamericana: caracterización y relevancia evolutiva. Acta Entomológica Chilena 28, 41–50.
Morrone JJ (2005) Hacia una síntesis biogeográfica de México. Revista Mexicana de Biodiversidad 76, 207–252.
| Hacia una síntesis biogeográfica de México.Crossref | GoogleScholarGoogle Scholar |
Morrone JJ (2006) Biogeographic areas and transition zones of Latin America and the Caribbean Islands based on panbiogeographic and cladistic analyses of the entomofauna. Annual Review of Entomology 51, 467–494.
| Biogeographic areas and transition zones of Latin America and the Caribbean Islands based on panbiogeographic and cladistic analyses of the entomofauna.Crossref | GoogleScholarGoogle Scholar | 16332220PubMed |
Morrone JJ (2011) Island evolutionary biogeography: analysis of the weevils (Coleoptera: Curculionidae) of the Falkland Islands (Islas Malvinas). Journal of Biogeography 38, 2078–2090.
| Island evolutionary biogeography: analysis of the weevils (Coleoptera: Curculionidae) of the Falkland Islands (Islas Malvinas).Crossref | GoogleScholarGoogle Scholar |
Morrone JJ (2014) Biogeographic regionalization of the Neotropical region. Zootaxa 3782, 1–110.
| Biogeographic regionalization of the Neotropical region.Crossref | GoogleScholarGoogle Scholar | 24871951PubMed |
Morrone JJ (2015a) Biogeographic regionalization of the world: a reappraisal. Australian Systematic Botany 28, 81–90.
| Biogeographic regionalization of the world: a reappraisal.Crossref | GoogleScholarGoogle Scholar |
Morrone JJ (2015b) Biogeographical regionalisation of the Andean region. Zootaxa 3936, 207–236.
| Biogeographical regionalisation of the Andean region.Crossref | GoogleScholarGoogle Scholar | 25947431PubMed |
Morrone JJ (2017) ‘Neotropical biogeography: regionalization and evolution.’ (CRC Press: Boca Raton, FL, USA)
Morrone JJ (2018) ‘Evolutionary biogeography of the Andean region.’ (CRC Press: Boca Raton, FL, USA)
Morrone JJ (2020) ‘The Mexican Transition Zone: a natural biogeographic laboratory to study biotic assembly.’ (Springer: Cham, Switzerland)
Morrone JJ, Espinosa-Organista D, Aguilar C, Llorente J (1999) Preliminary classification of the Mexican biogeographic provinces: a parsimony analysis of endemicity based on plant, insect and bird taxa. The Southwestern Naturalist 44, 507–514.
| Preliminary classification of the Mexican biogeographic provinces: a parsimony analysis of endemicity based on plant, insect and bird taxa.Crossref | GoogleScholarGoogle Scholar |
Müller P (1973) ‘The dispersal centres of terrestrial vertebrates in the Neotropical realm: a study in the evolution of the Neotropical biota and its native landscapes.’ (Junk: The Hague, Netherlands)
Müller P (1979) ‘Introducción a la biogeografía.’ (Blume: Madrid, Spain)
Müller P (1986) ‘Biogeography.’ (Harper and Row: New York, NY, USA)
Murray A (1866) ‘The geographical distribution of mammals.’ (Day and Son Limited: London, UK)
Nelson G (1978) From Candolle to Croizat: comments on the history of biogeography. Journal of the History of Biology 11, 269–305.
Nelson G, Ladiges PY (1996) Paralogy in cladistic biogeography and analysis of paralogy-free subtrees. American Museum Novitates 3167, 1–58.
Nelson G, Platnick NI (1981) ‘Systematics and biogeography: cladistics and vicariance.’ (Columbia University Press: New York, NY, USA)
Newbigin MI (1913) ‘Animal geography: the faunas of the natural regions of the world.’ (Clarendon Press: Oxford, UK)
Newbigin MI (1950) ‘Plant and animal geography.’ (Methuen: London, UK)
Newton A (1893) ‘A dictionary of birds.’ (Adam and Charles Black: London, UK)
Newton JW (1845) ‘Animal topography arm the geographical distribution of animals illustrated in familiar verse.’ (William Spooner: London, UK)
Nichols JT (1943) ‘The freshwater fishes of China. Natural history of Central Asia: Volume IX.’ (The American Museum of Natural History: New York, NY, USA)
Nihei SS, de Carvalho CJB (2004) Taxonomy, cladistics and biogeography of Coenosopsia Malloch (Diptera, Anthomyiidae) and its significance to the evolution of anthomyiids in the Neotropics. Systematic Entomology 29, 260–275.
| Taxonomy, cladistics and biogeography of Coenosopsia Malloch (Diptera, Anthomyiidae) and its significance to the evolution of anthomyiids in the Neotropics.Crossref | GoogleScholarGoogle Scholar |
Ojeda RA, Borghi CE, Roig VG (2002) Mamíferos de Argentina. In ‘Diversidad y conservación de mamíferos neotropicales’. (Eds G Ceballos, JA Simonetti) pp. 23–63. (Conabio and UNAM: Mexico City, Mexico)
Olson DM, Dinerstein E, Wikramanayake ED, Burgess ND, Powell GVN, Underwood EC, D’Amico JA, Itoua IE, Strand HE, Morrison JC, Loucks CJ, Allnutt TF, Ricketts TH, Kura Y, Lamoreux JF, Wettengel WW, Hedao P, Kassem KR (2001) Terrestrial ecoregions of the world: a new map of life on Earth. Bioscience 51, 933–938.
| Terrestrial ecoregions of the world: a new map of life on Earth.Crossref | GoogleScholarGoogle Scholar |
Orfila RN (1941) Apuntaciones ornitológicas para la zoogeografía neotropical. I. Distrito Sabánico. Revista Argentina de Zoogeografía 1, 85–92.
Ortmann AE (1896) ‘Grundzüge der marinen Tiergeographie.’ (Gustav Fischer: Jena, German Empire)
Packard AS (1883) Monograph of Phyllopod crustacea. Twelfth annual report of the US Geological and Geographical Survey of the Territories 1, 1–592.
Palestrini C, Simonis A, Zunino M (1985) Modelli di distribuzione dell’entomofauna della Zona di Transizione Cinese, analisi di esempi e ipotesi sulle sue origini. Biogeographia 11, 195–209.
Parenti LR, Ebach MC (2010) Wallacea deconstructed. In ‘Beyond cladistics: the branching of a paradigm’. (Eds DM Williams, S Knapp) pp. 303–318. (University of California Press: Berkeley, CA, USA)
Parenti LR, Ebach MC (2013) Evidence and hypothesis in biogeography. Journal of Biogeography 40, 813–820.
| Evidence and hypothesis in biogeography.Crossref | GoogleScholarGoogle Scholar |
Parodi LR (1934) Las plantas indígenas no alimenticias cultivadas en la Argentina. Revista Argentina de Agronomía 1, 165–212.
Paulson DR (1979) Odonata. In ‘Biota Acuática de Sudamérica austral’. (Ed. SH Hurlbert) pp. 170–171. (San Diego State University: San Diego, CA, USA)
Pellegrino KCM, Rodrigues MT, Waite AN, Morando M, Yassuda YY, Sites JW (2005) Phylogeography and species limits in the Gymnodactylus darwinii complex (Gekkonidae, Squamata): genetic structure coincides with river systems in the Brazilian Atlantic forest. Biological Journal of the Linnean Society 85, 13–26.
| Phylogeography and species limits in the Gymnodactylus darwinii complex (Gekkonidae, Squamata): genetic structure coincides with river systems in the Brazilian Atlantic forest.Crossref | GoogleScholarGoogle Scholar |
Peña LE (1966) A preliminary attempt to divide Chile into entomofaunal regions, based on the Tenebrionidae (Coleoptera). Postilla 97, 1–17.
Pires AC, Marinoni L (2011) Distributional patterns of the Neotropical genus Thecomyia Perty (Diptera, Sciomyzidae) and phylogenetic support. Revista Brasileira de Entomologia 55, 6–14.
| Distributional patterns of the Neotropical genus Thecomyia Perty (Diptera, Sciomyzidae) and phylogenetic support.Crossref | GoogleScholarGoogle Scholar |
Platnick NI, Nelson G (1978) A method of analysis for historical biogeography. Systematic Zoology 27, 1–16.
| A method of analysis for historical biogeography.Crossref | GoogleScholarGoogle Scholar |
Porzecanski AL, Cracraft J (2005) Cladistic analysis of distributions and endemism (CADE): using raw distributions of birds to unravel the biogeography of the South American aridlands. Journal of Biogeography 32, 261–275.
| Cladistic analysis of distributions and endemism (CADE): using raw distributions of birds to unravel the biogeography of the South American aridlands.Crossref | GoogleScholarGoogle Scholar |
Posadas P, Estévez JM, Morrone JJ (1997) Distributional patterns and endemism areas of vascular plants in the Andean subregion. Fontqueria 48, 1–10.
Prichard JC (1826) ‘Researches into the physical history of mankind’, 2nd edn. (Houlfton and Stoneman: London, UK)
Procheş S (2005) The world’s biogeographical regions: cluster analyses based on bat distributions. Journal of Biogeography 32, 607–614.
Procheş S (2006) Latitudinal and longitudinal barriers in global biogeography. Biology Letters 2, 69–72.
Procheş Ş, Ramdhani S (2012) The world’s zoogeographical regions confirmed by cross-taxon analyses. Bioscience 62, 260–270.
| The world’s zoogeographical regions confirmed by cross-taxon analyses.Crossref | GoogleScholarGoogle Scholar |
Poynton JC (1959) Zoogeography. Nucleon 1, 24–30.
Poynton JC (2000) Evidence of an Afrotemperate amphibian fauna. African Journal of Herpetology 49, 33–41.
| Evidence of an Afrotemperate amphibian fauna.Crossref | GoogleScholarGoogle Scholar |
Ramos KS, Melo GAR (2010) Taxonomic revision and phylogenetic relationships of the bee genus Parapsaenythia Friese (Hymenoptera, Apidae, Protandrenini), with biogeographic inferences for the South American Chacoan subregion. Systematic Entomology 35, 449–474.
| Taxonomic revision and phylogenetic relationships of the bee genus Parapsaenythia Friese (Hymenoptera, Apidae, Protandrenini), with biogeographic inferences for the South American Chacoan subregion.Crossref | GoogleScholarGoogle Scholar |
Rapoport EH (1968) Algunos problemas biogeográficos del Nuevo Mundo con especial referencia a la región Neotropical. In ‘Biologie de l’Amerique Australe’. (Eds C Delamare Debouteville, EH Rapoport) Vol. 4, pp. 55–110. (CNRS: Paris, France)
Raven HC (1935) Wallace’s Line and the distribution of Indo-Australian mammals. Bulletin of the American Museum of Natural History 68, 177–293.
Rensch B (1936) ‘Die Geschichte des Sundabogens; eine tiergeographische Untersuchung.’ (Von Bornträger: Berlin, Germany)
Ribeiro GC, Santos CMD, Olivieri LT, Santos D, Berbert JM, Eterovic A (2014) The world’s biogeographical regions revisited: global patterns of endemism in Tipulidae (Diptera). Zootaxa 3847, 241–258.
| The world’s biogeographical regions revisited: global patterns of endemism in Tipulidae (Diptera).Crossref | GoogleScholarGoogle Scholar | 25112336PubMed |
Ringuelet RA (1955) Vinculaciones faunísticas de la zona boscosa del Nahuel Huapi y el dominio zoogeográfico Australcordillerano. Notas del Museo de La Plata. Zoologia 18, 21–121.
Ringuelet RA (1961) Rasgos fundamentales de la zoogeografía de la Argentina. Physis 22, 151–170.
Ringuelet RA (1975) Zoogeografía y ecología de los peces de aguas continentales de la Argentina y consideraciones sobre las áreas ictiológicas de América del Sur. Ecosur 2, 1–122.
Ringuelet RA (1978) Dinamismo histórico de la fauna brasílica en la Argentina. Ameghiniana 15, 255–262.
Rivas-Martínez S, Navarro G (1994) ‘Mapa biogeográfico de Suramérica.’ (Published by the authors: Madrid, Spain)
Rivas-Martínez S, Tovar O (1983) Síntesis biogeográfica de los Andes. Collectanea Botanica (Barcelona) 14, 515–521.
Rivas-Martínez S, Navarro G, Penas Á, Costa M (2011) Biogeographic map of South America. A preliminary survey. International Journal of Geobotanical Research 1, 21–40.
| Biogeographic map of South America. A preliminary survey.Crossref | GoogleScholarGoogle Scholar |
Roig-Juñent S, Domínguez MC (2001) Diversidad de la familia Carabidae (Coleoptera) en Chile. Revista Chilena de Historia Natural 74, 549–571.
| Diversidad de la familia Carabidae (Coleoptera) en Chile.Crossref | GoogleScholarGoogle Scholar |
Roig-Juñent S, Domínguez MC, Flores GE, Mattoni C (2006) Biogeographic history of South American arid lands: a view from its arthropods using TASS analysis. Journal of Arid Environments 66, 404–420.
| Biogeographic history of South American arid lands: a view from its arthropods using TASS analysis.Crossref | GoogleScholarGoogle Scholar |
Roig-Juñent SA, Griotti M, Domínguez MC, Agrain FA, Campos-Soldini P, Carrara R, Cheli G, Fernández-Campón F, Flores GR, Katinas L, Muzón JR, Neita-Moreno JC, Pessacq P, San Blas G, Scheibler EE, Crisci JV (2018) The Patagonian Steppe biogeographic province: Andean region or South American transition zone? Zoologica Scripta 47, 623–629.
| The Patagonian Steppe biogeographic province: Andean region or South American transition zone?Crossref | GoogleScholarGoogle Scholar |
Rosen DE (1979) Fishes of the uplands and intermontane basins: revisionary studies and comparative geography. Bulletin of the American Museum of Natural History 162, 1–176.
Rueda M, Rodríguez MA, Hawkins B (2013) Identifying global zoogeographical regions: lessons from Wallace. Journal of Biogeography 40, 2215–2225.
| Identifying global zoogeographical regions: lessons from Wallace.Crossref | GoogleScholarGoogle Scholar |
Rzedowski J (1978) ‘La vegetación de México.’ (Editorial Limusa: Mexico City, Mexico)
Samek V, del Risco E, Vandana R (1988) Fitorregionalización del Caribe. Revista del Jardín Botánico Nacional (La Habana) 9, 25–38.
Sánchez Osés C, Pérez-Hernández R (2005) Historia y tabla de equivalencias de las propuestas de subdivisiones biogeográficas de la región Neotropical. In ‘Regionalización biogeográfica en Iberoamérica y tópicos afines– Primeras Jornadas Biogeográficas de la Red Iberoamericana de Biogeografía y Entomología Sistemática (RIBES XII. I–CYTED). Las Prensas de Ciencias’. (Eds J Llorente Bousquets, JJ Morrone) pp. 495–508. (UNAM: Mexico City, Mexico)
Santos D, Ribeiro GC (2022) Areas of endemism in the Afrotropical region based on the geographical distribution of Tipulomorpha (Insecta: Diptera). Austral Ecology 47, 92–113.
| Areas of endemism in the Afrotropical region based on the geographical distribution of Tipulomorpha (Insecta: Diptera).Crossref | GoogleScholarGoogle Scholar |
Schilder FA (1956) ‘Lehrbuch der Allgemeinen Zoogeographie.’ (Gustav Fischer Verlag: Jena, German Democratic Republic)
Schmarda KL (1853) ‘Die geographische Verbreitung der thiere.’ (Carl Gerold and Son: Vienna, Austria)
Schmidt KP (1954) Faunal realms, regions, and provinces. The Quarterly Review of Biology 29, 322–331.
| Faunal realms, regions, and provinces.Crossref | GoogleScholarGoogle Scholar | 13237405PubMed |
Schouw JF (1823) ‘Grundzüge einer allgemeinen Pflanzengeographie.’ (Reimer: Berlin, Prussia)
Sclater PL (1858) On the general geographic distribution of the members of the class Aves. Proceedings of the Linnean Society of London, Zoology 2, 130–145.
Sclater WL (1894) The geography of mammals: No. I. Introductory. The Geographical Journal 3, 95–105.
| The geography of mammals: No. I. Introductory.Crossref | GoogleScholarGoogle Scholar |
Sclater WL (1897) The geography of mammals. No. VI. The Nearctic Region (continued). The Geographical Journal 9, 67–76.
Sclater WL, Sclater PL (1899) ‘The geography of mammals.’ (Kegan Paul, Trench, Trübner and Co.: London, UK)
Sérsic AN, Cosacov A, Cocucci AA, Johnson LA, Pozner R, Ávila LJ, Sites JW, Morando M (2011) Emerging phylogeographical patterns of plants and terrestrial vertebrates from Patagonia. Biological Journal of the Linnean Society. Linnean Society of London 103, 475–494.
| Emerging phylogeographical patterns of plants and terrestrial vertebrates from Patagonia.Crossref | GoogleScholarGoogle Scholar |
Shannon RC (1927) Contribución a los estudios de las zonas biológicas de la República Argentina. Revista de la Sociedad Entomológica Argentina 4, 1–14.
Sharpe RB (1884) On the zoo-geographical areas of the world, illustrating the distribution of birds. Nature and Science 3, 100–108.
Sharpe RB (1893) On the zoo‐geographical areas of the world, illustrating the distribution of birds. Natural Science 3, 100–108.
Sick WD (1969) Geographical substance. Monographiae Biologicae 19, 449–474.
Sigrist MS, de Carvalho CJB (2009) Historical relationships among areas of endemism in the tropical South America using Brooks Parsimony Analysis (BPA). Biota Neotropica 9, 79–90.
| Historical relationships among areas of endemism in the tropical South America using Brooks Parsimony Analysis (BPA).Crossref | GoogleScholarGoogle Scholar |
Skottsberg C (1905) Some remarks upon the geographical distribution of vegetation in the colder Southern Hemisphere. Ymer 4, 402–427.
Smith CH (1983) A system of world mammal faunal regions. I. Logical and statistical derivation of the regions. Journal of Biogeography 10, 455–466.
| A system of world mammal faunal regions. I. Logical and statistical derivation of the regions.Crossref | GoogleScholarGoogle Scholar |
Smith CH (2005) Some biogeographers, evolutionists, and ecologists: chrono-biographical sketches. Alphabetical listing of the naturalists. Available at http://people.wku.edu/charles.smith/chronob/homelist.htm
Smith HM (1941) An analysis of the biotic provinces of Mexico, as indicated by the distribution of the lizards of the genus Sceloporus. Anales. Escuela Nacional de Ciencias Biologicas (Mexico) 2, 95–103.
Sneath PHA (1967) Conifer distributions and continental drift. Nature 215, 467–470.
| Conifer distributions and continental drift.Crossref | GoogleScholarGoogle Scholar |
Spinelli GR, Marino PI, Posadas P (2006) The Patagonian species of the genus Atrichopogon Kieffer, with a biogeographic analysis based on Forcipomyiinae (Diptera: Ceratopogonidae). Insect Systematics & Evolution 37, 301–324.
| The Patagonian species of the genus Atrichopogon Kieffer, with a biogeographic analysis based on Forcipomyiinae (Diptera: Ceratopogonidae).Crossref | GoogleScholarGoogle Scholar |
Stoddart DR (1992) Biogeography of the tropical Pacific. Pacific Science 46, 276–293.
Swainson W (1835) ‘A treatise on the geography and classification of animals.’ (Longman, Brown, Green, and Longman: London, UK)
Takhtajan A (1978) ‘The floristic regions of the world.’ (“Nauka” Publishing: Leningrad, USSR) [In Russian]
Takhtajan A (1986) ‘Floristic regions of the world.’ (University of California Press: Berkeley, CA, USA)
Tatewaki M (1963) Phytogeography of the islands of the North Pacific Ocean. In ‘Pacific Ocean biogeography: a symposium: Tenth Pacific Science Congress’, 21 August–6 September 1961, Honolulu, HI, USA. (Ed. JL Gressitt, CH Lindroth, FR Fosberg, CA Fleming, EG Turbott) pp. 23–28. (Bishop Museum Press: Honolulu, HI, USA)
Thorne RF (1963) Biotic distributions in the tropical Pacific. In ‘Pacific basin biogeography: a symposium: Tenth Pacific Science Congress’, 21 August–6 September 1961, Honolulu, HI, USA. (Eds JL Gressitt, CH Lindroth, FR Fosberg, CA Fleming, EG Turbott) pp. 311–350. (Bishop Museum Press: Honolulu, HI, USA)
Udvardy MDF (1975) A classification of the biogeographical provinces of the world. Occasional Paper 18. (International Union for Conservation of Nature and Natural Resources: Morges, Switzerland) Available at https://portals.iucn.org/library/sites/library/files/documents/OP-018.pdf
Udvardy MDF (1987) The biogeographical realm Antarctica: a proposal. Journal of the Royal Society of New Zealand 17, 187–194.
| The biogeographical realm Antarctica: a proposal.Crossref | GoogleScholarGoogle Scholar |
Urtubey E, Stuessy TF, Tremetsberger K, Morrone JJ (2010) The South American biogeographic transition zone: An analysis from Asteraceae. Taxon 59, 505–509.
| The South American biogeographic transition zone: An analysis from Asteraceae.Crossref | GoogleScholarGoogle Scholar |
Vallejo B (2011) The Philippines in Wallacea. In ‘Biodiversity, biogeography and nature conservation in Wallacea and New Guinea’ (Ed. D Telnov) Vol. I, pp. 27–42. (The Entomological Society of Latvia: Riga, Latvia)
von Hofsten NGE (1916) Zur älteren Geschichte des Diskontinuitätsproblems in der Biogeographie. Zoologische Annalen Zeitschrift für Geschichte der Zoologie 7, 197–353.
van Rooy J, van Wyk AE (2012) Phytogeographical and ecological affinities of the bryofloristic regions of southern Africa. Polish Botanical Journal 57, 109–118.
Viloria AL 2005. Las mariposas (Lepidoptera: Papilionoidea) y la regionalización biogeográfica de Venezuela. In ‘Regionalización biogeográfica en Iberoamérica y tópicos afines – Primeras Jornadas Biogeográficas de la Red Iberoamericana de Biogeografía y Entomología Sistemática (RIBES XII. I–CYTED). Las Prensas de Ciencias’. (Eds J Llorente Bousquets, JJ Morrone) pp. 441–459. (UNAM: Mexico City, Mexico)
Wagner JA (1844) Die Geographische Verbreitung der Säugethiere Dargestellt. Abhandlungen der Königlich Bayerischen Akademie der Wissenschaften. Mathematisch‐Physikalische Klasse 1, 1–146.
Wagner JA (1845) Die Geographische Verbreitung der Säugethiere Dargestellt. Abhandlungen, Bayerische Akademie der Wissenschaften 2, 37–108.
Wagner JA (1846a) Die Geographische Verbreitung der Säugethiere Dargestellt. Abhandlungen, Bayerische Akademie der Wissenschaften 3, 1–114.
Wagner JA (1846b) Die Geographische Verbreitung der Säugethiere Dargestellt. Abhandlungen, Bayerische Akademie der Wissenschaften 4, 241–244.
Wallace AR (1876a) ‘The geographical distribution of animals. With a study of the relations of living and extinct faunas as elucidating the past changes of the earth’s surface. Vol. I.’ (Harper and Brothers: New York, NY, USA)
Wallace AR (1876b) ‘The geographical distribution of animals. With a study of the relations of living and extinct faunas as elucidating the past changes of the earth’s surface. Vol. II.’ (Harper and Brothers: New York, NY, USA)
Wallace AR (1883) On the value of the ‘Neoarctic’ [sic] as one of the primary zoological regions. Nature 27, 482–483.
Wallace AR (1894) What are zoological regions? Nature 49, 610–613.
Wallaschek M (2009) ‘Fragmente zur Geschichte und Theorie der Zoogeographie: Die Begriffe Zoogeographie, Arealsystem und Areal.’ (Published by the author: Halle, Germany)
Wallaschek M (2010a) ‘Fragmente zur Geschichte und Theorie der Zoogeographie: II. Die Begriffe Fauna und Faunistik.’ (Published by the author: Halle, Germany)
Wallaschek M (2010b) ‘Fragmente zur Geschichte und Theorie der Zoogeographie: III. Die Begriffe Verbreitung und Ausbreitung.’ (Published by the author: Halle, Germany)
Wallaschek M (2011a) ‘Fragmente zur Geschichte und Theorie der Zoogeographie: IV. Die chorologische Zoogeographie und ihre Anfänge.’ (Published by the author: Halle, Germany)
Wallaschek M (2011b) ‘Fragmente zur Geschichte und Theorie der Zoogeographie: V. Die chorologische Zoogeographie und ihr Fortgang.’ (Published by the author: Halle, Germany)
Wallaschek M (2012a) ‘Fragmente zur Geschichte und Theorie der Zoogeographie: VI. Vergleichende Zoogeographie.’ (Published by the author: Halle, Germany)
Wallaschek M (2012b) ‘Fragmente zur Geschichte und Theorie der Zoogeographie: VI. Vergleichende Zoogeographie.’ (Published by the author: Halle, Germany)
Wallaschek M (2012c) ‘Fragmente zur Geschichte und Theorie der Zoogeographie: VII. Die ökologische Zoogeographie.’ (Published by the author: Halle, Germany)
Wallaschek M (2013a) ‘Fragmente zur Geschichte und Theorie der Zoogeographie: VIII. Die historische Zoogeographie.’ (Published by the author: Halle, Germany)
Wallaschek M (2013b) ‘Fragmente zur Geschichte und Theorie der Zoogeographie: IX. Fazit, Literatur, Glossar, Zoogeographenverzeichnis.’ (Published by the author: Halle, Germany)
Wallaschek M (2014) ‘Ludwig Karl Schmarda (1819–1908): Leben und Werk.’ (Published by the author: Halle, Germany)
Wallaschek M (2015a) Johann Andreas Wagner (1797–1861) und “Die geographische Verbreitung der Säugthiere”. Beiträge zur Geschichte der Zoogeographie 1, 3–24.
Wallaschek M (2015b) Johann Friedrich Blumenbach (1752–1840) und die Zoogeographie im “Handbuch der Naturgeschichte”. Philippia 16, 237–262.
West RC (1964) The natural regions of Middle America. In ‘Handbook of Middle American Indians, vol 1’. (Ed. RC West) pp. 363–383. (University of Texas Press: Austin, TX, USA)
Williams PH (1996) Mapping variations in the strength and breadth of biogeographic transition zones using species turnover. Proceedings of the Royal Society of London – B. Biological Sciences 263, 579–588.
| Mapping variations in the strength and breadth of biogeographic transition zones using species turnover.Crossref | GoogleScholarGoogle Scholar |
Zhang CL (1954) The distribution of the freshwater fishes of China. Journal of Geographical Sciences 3, 279–284. [In Chinese]
Zimmermann EAW (1777) ‘Specimen zoologiae geographicae, Quadrupedum domicilia et migrationes sistens.’ (Theodorum Haak: Leiden, Netherlands)
Zimmermann EAW (1783) ‘Geographische Geschichte des Menschen, und der allgemein verbreiteten vierfüssigen Thiere.’ (Weygandschen Buchhandlung: Leiden, Netherlands)
Zuloaga FO, Morrone O, Rodríguez D (1999) Análisis de la biodiversidad en plantas vasculares de la Argentina. Kurtziana 27, 17–167.