

Additions to Amanita (Amanitaceae, Agaricales) section Arenariae from south-western Australia
E. M. Davison A B F , D. Giustiniano C D , N. L. Bougher B , L. E. McGurk C E and E. L. J. Watkin C DA School of Molecular and Life Sciences, Curtin University, GPO Box U1987, Perth, WA 6845, Australia.
B Western Australian Herbarium, Department of Biodiversity, Conservation and Attractions, Locked Bag 104, Bentley Delivery Centre, WA 6983, Australia.
C School of Biomedical Sciences, Curtin University, GPO Box U1987, Perth, WA 6845, Australia.
D Curtin Medical School, Curtin University, GPO Box U1987, Perth, WA 6845, Australia.
E Present address: Human Resources, The University of Western Australia, Crawley, WA 6009, Australia.
F Corresponding author. Email: e.davison@curtin.edu.au
Australian Systematic Botany 34(6) 541-569 https://doi.org/10.1071/SB21017
Submitted: 27 4 2021 Accepted: 29 7 2021 Published: 8 September 2021
Journal Compilation © CSIRO 2021 Open Access CC BY
Abstract
A recent molecular phylogeny of Amanita recognises three subgenera and 11 sections. Members of subgenus Amanitina are characterised by amyloid spores and a mycorrhizal habit. Section Arenariae falls within this subgenus. Members of this section are known only from southern Australia; they are either sequestrate (secotioid) or agaricoid and lack clamp connections. We describe the following three additional secotioid species: Amanita arenarioides Bougher, E.M.Davison & Giustiniano, A. compacta Bougher, E.M.Davison & Giustiniano and A. pseudoarenaria E.M.Davison, Giustiniano & Bougher, which are separated on macroscopic appearance, spore shape and genetic sequences. We also describe two agaricoid species, namely, A. pupatuju E.M.Davison, Giustiniano, McGurk & E.L.J.Watkin, and A sabulosa E.M.Davison & Giustiniano, which are separated on bulb shape and genetic sequences. We provide expanded descriptions of A. arenaria (O.K.Mill. & E.Horak) Justo and A. griselloides D.A.Reid; we also synonymise A. dumosorum D.A.Reid with A. peltigera D.A.Reid. A revised diagnosis and description of section Arenariae is provided, together with a key to currently recognised member of this section.
Keywords: Basidiomycota, multi-gene phylogenetics, taxonomy, ITS variation.
Introduction
Amanita Pers. is the largest genus in Amanitaceae, with ~1000 described species worldwide; however, only ~600 are well defined (Yang et al. 2018). This genus is important ecologically because most species are mycorrhizal with woody plants. It is economically important because some species are comestible, and others are poisonous (Cui et al. 2018). The genus is defined by the field character of a membranous or friable universal veil, and by the microscopic and ontogenetic characters of bilateral lamella trama, acrophysalidic stipe trama and schizohymenial development (Bas 1969, pp. 299–333). Like many mushrooms, Amanita spp. are phenotypically variable; consequently, many species cannot be confidently identified using field characters alone; microscopic examination is also essential. Molecular analyses too have been instrumental in both identifying new species and clarifying relationships within the genus (Cui et al. 2018; Yang et al. 2018).
Within Australia, agaricoid species have been monographed by Reid (1980) and Wood (1997). Additional descriptions and clarifications have been made by Miller (1991, 1992), Grgurinovic (1997), Davison (2011), Davison and Giustiniano (2020), Davison et al. (2013, 2015, 2017a, 2017b, 2020) and McGurk et al. (2016). In addition, there are sequestrate species that were assigned to Amarrendia Bougher & T.Lebel and Torrendia Bres., genera that are now synonymised with Amanita (Miller and Horak 1992; Bougher 1999; Bougher and Lebel 2002; Justo et al. 2010). There are ~100 named Australian species, but many more await description.
The family Amanitaceae has been recently revised by Cui et al. (2018), using concatenated datasets of nuLSU (28S nuclear ribosomal large subunit rRNA gene), β-tubulin, ef1-α (elongation factor 1-α), and rpb2 (RNA polymerase II) gene regions. Within Amanita, they recognise three subgenera, namely, Amanita, Amanitina (E.-J.Gilbert) E.-J.Gilbert and Lepidella Beauseign. Most Australian species fall within subgenus Amanitina because they have amyloid spores, a bulb at the base of the stipe, and are likely to be mycorrhizal. Cui et al. (2018) recognised the following six sections within Amanitina: section Amidella (E.-J.Gilbert) Konrad & Maubl., section Arenariae Zhu L.Yang, Y.Y.Cui & Q.Cai, section Phalloideae (Fr.) Quél., section Roanokenses Singer, section Strobiliformes Singer ex Zhu L.Yang, Y.Y.Cui & Q.Cai, and section Validae (Fr.) Quél. These sections can be recognised morphologically by differences in the pileus margin, form of the universal veil, and presence or absence of clamp connections.
Members of section Arenariae are agaricoid or sequestrate, their basidia are thin- or thick-walled and lack clamp connections; agaricoid species have an appendiculate pileus margin and remnants of universal veil on the pileus (Cui et al. 2018, pp. 106, 117). Five species have been described; they are known only from southern Australia. The type is A. arenaria (O.K.Mill. & E.Horak) Justo, a sequestrate (secotioid) species from the south-west of Western Australia (WA). Amanita arenaria is unusual because it has inamyloid spores, whereas they are amyloid in agaricoid members of this section. There are a further five sequestrate species of Amanita known from Australia, and they cluster together in section Amarrendiae (Bougher & Lebel) Zhu L.Yang, Y.Y.Cui, Q.Cai & L.P.Tang, which are in subgenus Amanita, not subgenus Amanitina (Justo et al. 2010; Cui et al. 2018, pp. 19, 44).
Miller and Horak (1992) described A. arenaria (as Torrendia arenaria O.K.Mill. & E.Horak) from collections they had made while visiting WA in 1985, 1989 and 1991. Miller split two of his collections (OKM 24013, VPI 679; OKM 24018, VPI 412) and deposited duplicates in the Western Australian Herbarium (PERTH). Within T. arenaria, they described T. arenaria f. lutescens O.K.Mill. & E.Horak for material showing yellow stains on the stipe and pileus or slowly turning yellowish on bruising or cutting. Justo et al. (2010) re-examined 10 of Miller’s collections of T. arenaria, together with other Torrendia species, and used the nuLSU gene region to determine whether they were closely related to Amanita, as had been suggested by Bas (1975). They showed all Torrendia species nested within Amanita and transferred all species to Amanita. Their results showed that these secotioid species were not monophyletic because they fell into three well-supported clades in different parts of the genus. They also showed that four of the collections identified by Miller and Horak (1992) as T. arenaria were an undescribed species that was morphologically and molecularly distinct, and described it as A. pseudoinculta Justo. One of the collections examined by Justo et al. (2010) was T. arenaria f. lutescens; however, the nuLSU sequence from this collection did not differ from sequences from collections identified as T. arenaria f. arenaria. They also noted that the six A. arenaria collections consistently clustered into two subgroups that differed in spore shape; those from the type locality were ellipsoid to elongate (Q = 1.55–1.72), whereas spores of the others were broadly ellipsoid to ellipsoid (Q = 1.28–1.36). They suggested that this either represented two cryptic species, or there was high genetic divergence among collections from different localities.
While reviewing all A. arenaria collections in PERTH and in private collections, we have noted there are at least three species that are macroscopically similar but can be separated by spore shape and molecular sequences. We describe these in this paper, together with another secotioid species that differs macroscopically from A. arenaria.
There are four agaricoid species that are recognised as being members of section Arenariae; these are A. lesueurii E.M.Davison, A. peltigera D.A.Reid, A. wadjukiorum E.M.Davison and A. wadulawitu McGurk, E.M.Davison & E.L.J.Watkin (Cui et al. 2018; Davison et al. 2020). In this paper, we describe two additional species. Many more described species may be recognised as being members of this section if widespread sequencing of described Australian amanitas is conducted.
Reid (1980) described A. griselloides D.A.Reid as having a layered universal veil on the pileus; however, examination of the type by E. M. Davison found that the superficial layer was formed by hyphomycetes. We provide a revised description in this paper.
The type of A. peltigera was described from South Australia (SA) as having a saccate volva and lacking a partial veil (Reid 1980; Davison et al. 2020). However, examination of collections from SA and the south-west of WA showed that they form a single clade exhibiting variation in both the persistence of the partial veil and remains of the universal veil at the top of the bulb (Davison et al. 2020). Reid (1980) also described A. dumosorum D.A.Reid, which he considered to be similar to A. peltigera apart from having a partial veil, lacking a saccate volva, and lacking infrequent large, inflated cells in the universal veil on the pileus. However, examination of the type of A. dumosorum (K(M) 236387) by E. M. Davison found infrequent large, inflated cells in this tissue. The other differences noted by Reid (1980) occurred in collections of A. peltigera (Davison et al. 2020); because these two species cannot be separated by their field characters or micromorphology, they are synonymised in this paper.
The nuclear ribosomal internal transcribed spacer (ITS) region has been used as a barcode marker for species discrimination within the Basidiomycota (Schoch et al. 2012). Hughes et al. (2013) found less than 2% base pair divergence among haplotypes within an individual, and Cai et al. (2014) used ITS sequences to separate Amanita species within section Phalloideae. The ITS region does not appear as useful among Australian species. For example, Davison et al. (2017a) found that ITS sequences fail to separate three species from section Phalloideae, which differ in spore shape and are geographically separated, and within A. peltigera, the base pair divergence among haplotypes from the same individual is between 0.2 and 5.8% (Davison et al. 2020). In this paper, we determine the difference between ITS clones from both secotioid and agaricoid members of section Arenariae, to determine whether they show divergence similar to that observed in A. peltigera.
This paper has four aims. First, molecular sequences are used to check the placement of the undescribed species and A. griselloides D.A.Reid within section Arenariae, their relationships with one another, and the variation within the ITS region of haplotypes from the same individual and from different collections of the same species. Second, we describe three new secotioid and two new agaricoid species, together with a revised description of A. arenaria and an expanded description of A. griselloides. Third, we synonymise A. dumosorum with A. peltigera. Last, we provide revised diagnosis and description of section Arenariae and a key to currently recognised members of this section.
Materials and methods
Taxonomy
The methodology used for describing the macroscopic and microscopic characters largely follows Tulloss (2000). Colour names, including the colour of spores in deposit and other shades of white to cream (designated by the letters A–G), follow Royal Botanic Garden, Edinburgh (1969), whereas colour codes are from Kornerup and Wanscher (1983). In the descriptions of basidiospores (and basidia) the notation [x/y/z] denotes x basidiospores measured from y basidiomes from z collections. Biometric variables for spores follow Tulloss (2000), that is:
-
L =the average spore length computed for one specimen examined and the range of such averages
-
L′ = average spore length computed for all spores measured
-
W = the average spore width computed for one specimen examined and the range of such averages
-
W′ = average spore width computed for all spores measured
-
Q =the length:breadth for a single spore and the range of the ratio of length:breadth for all spores measured
-
Q =the average value of Q computed for one specimen examined and the range of such averages
-
Q′ = the average value of Q computed for all spores measured
Author citations follow Index Fungorum (see http://www.indexfungorum.org/Names/Names.asp, accessed 19 February 2021). Herbarium codes follow Index Herbariorum (see http://sweetgum.nybg.org/science/ih/, accessed 1 February 2021).
Phylogenetics
DNA extraction, amplification and cloning of the ITS, amplification of the nuLSU, ef1-α, rpb2 and β-tubulin regions follow the methodology in Davison et al. (2017b). Sequence data were assembled with Geneious (ver. 10.0.5, www.geneious.com; Kearse et al. 2012). Additional sequences were accessed from GenBank (see http://www.ncbi.nlm.nih.gov/, accessed 2 February 2021; Tables 1, 2).

|

|
Any polymerase chain reaction (PCR) chimeras in ITS sequences were detected using the ‘Remove chimeric reads’ function in Geneious, using the UCHIME program (ver. 11.0.667, see https://www.drive5.com/usearch/manual/uchime_algo.html; Edgar et al. 2011). Default settings were used (minimum reporting score 0.3, weight no vote 8, minimum divergence ratio 0.5). The recommended database for chimera detection in fungal ITS sequences was selected as the reference database (Abarenkov et al. 2020).
Maximum-likelihood phylogenetic trees were built using MEGA (ver. 5, see https://www.megasoftware.net; Tamura et al. 2011). The best model for each dataset was determined using the Model Function in MEGA. The general time-reversible model (Tavaré 1986) with gamma distribution rates was used to determine the placement of the undescribed species in subgenus Amanitina by using the nuLSU gene region. The Kimura 2-parameter model (Kimura 1980) with invariant sites was used for the ef1-α and β-tubulin gene regions. The Kimura 2-parameter model (Kimura 1980) with gamma distribution rates was used for the rpb2 gene regions. The Tamura–Nei model (Tamura and Nei 1993) with invariant sites and gamma distribution rates was used for concatenated nuLSU, rpb2, ef1-α and β-tubulin gene regions. The Tamura 3-parameter model with gamma distribution was used for the ITS gene regions for the secotioid species (Tamura 1992). Bootstrap consensus trees were inferred from 500 replicates.
Results
Phylogenetics
Placement of undescribed species in subgenus Amanitina
The nuLSU is the only gene region available from GenBank for all type species of sections within subgenus Amanitina. These, together with additional species (Table 1), show that A. arenarioides sp. nov., A. compacta sp. nov., A. griselloides, A. pseudoarenaria sp. nov., A. pupatju sp. nov. and A. sabulosa sp. nov. form clades within section Arenariae with good support (Fig. 1). There are two clades within this section. One well-supported clade contains the secotioid species A. arenaria, A. arenarioides, A. compacta and A. pseudoarenaria and the agaricoid species A. griselloides, A. pupatju and A. sabulosa. Taking into account the nuLSU phylogeny and the phylogenies of other regions (see below), these are considered as species-level clades and are described below (see Taxonomy section). The names of these novel taxa are indicated on the phylogenetic trees, and previous misidentifications are updated.

|
Additional gene regions
Additional gene regions (ef1-α, rpb2 and β-tubulin) are available for some collections within section Arenariae (Table 2). Phylogenetic analysis of the ef1-α (Fig. 2A), rpb2 (Fig. 2B), β-tubulin (Fig. 2C) and concatenated nuLSU, rpb2, ef1-α and β-tubulin gene regions (Fig. 2D) all show that the secotioid species (A. arenaria, A. arenarioides, A. compacta and A. pseudoarenaria) together with A. griselloides, A. pupatju and A. sabulosa form a well-supported clade, separate from A. peltigera, A. wadjukiorum and A. wadulawitu.

|
ITS region
Cloned haplotypes of the ITS region are available for both secotioid and agaricoid species from section Arenariae. No chimeric reads were detected in these sequences.
In the secotioid species, the ITS region is 569–599 base pairs long. Molecular phylogenetic analysis by the maximum-likelihood method shows that GQ925393 and GQ925388 (sequences originally identified as A. arenaria by Justo et al. 2010) are widely separated. All the new species form distinct clades, some with good bootstrap support (Fig. 3). The bootstrap support for A. arenaria is 92% (13 clones from four collections), for A. arenarioides it is 67% (8 clones from two collections), for A. pseudoarenaria it is 77% (20 clones from five collections) and for A. compacta it is 53% (4 clones from a single collection).

|
As can be seen from Fig. 3, there is considerable variation between the clones; they differ by 0.0–8.1% within an individual, by 0.0–9.6% among collections of the same species, and by 10.3–18.8% among different species (Table 3).

|
Examination of the ITS region shows that there is an insertion of up to 20 base pairs (AAAGGAAGCTTCACTTTTGA in MW775323) in ITS1 in all haplotypes of A. arenaria; this is not present in any haplotypes of A. pseudoarenaria. There is a shorter, 15 base pair insertion (AGGGGAATCTTTTGA) in ITS1 in all haplotypes of A. arenarioides. There is a 10 base pair deletion in ITS1 in all haplotypes of A. arenaria and A. arenarioides, which is not present in A. pseudoarenaria and A. compacta.
In the agaricoid species A. griselloides, A. pupatju and A. sabulosa, the ITS region is 582–588 base pairs long. There is considerable variation among the clones: they differ by 0.2–9.9% among haplotypes from the same individual, by 0.7–10.1% among collections of the same species and by 12.2–18.0% among different species (Table 4).

|
Examination of the ITS region shows that there is a five base pair deletion in ITS1 in all haplotypes of A. griselloides and A. pupatju and a four base pair insertion in ITS1 in all haplotypes of A. pupatju.
Taxonomy
Section Arenariae
Cui et al. (2018, pp. 106, 117) stated that members of section Arenariae are agaricoid or sequestrate and lack clamp connections. Since their publication, more species have been recognised as being members of section Arenariae (including those detailed below), enabling us to provide a revised diagnosis and description of this section.
Diagnosis
Basiomata agaricoid or sequestrate; basiodiospores thin-walled or very slightly thickened, smooth, amyloid in agaricoid species, inamyloid in sequestrate species; clamp connections absent in all parts of the basidiome. When basidiomata is agaricoid, pileus with remnants of universal veil, lamellulae attenuate, stipe base variable. When basidiomata is sequestrate, gleba loculate, stipitate, basidia thin-walled, collapsing early. Known from southern Australia on sandy soil, associated with Agonis, Allocasuarina, Corymbia, Eucalyptus, Hibbertia, Jacksonia, Melaleuca, Mirbelia, Podocarpus and Taxandria.
Description
Basiomata agaricoid or sequestrate
When basidiomata agaricoid, pileus plane, margin non-striate, appendiculate when young, context white or cream, unchanging, or becoming vinaceous-buff when bruised. Universal veil on pileus present. Lamellae adnexed to adnate, white to cream; lamellulae attenuate. Stipe subcylindric, stipe base turbinate and elongating with age or ovoid or conic. Partial veil present. Universal veil at base of stipe indistinct or as soft warts or as small, flimsy free limb. Basidiospores amyloid, thin-walled, smooth. Sub-basidial cell becoming inflated. Clamp connections absent.
When sequestrate, basidiomata hypogeus to emergent, sequestrate and stipitate; pileus subglobose or pulvinate or cylindric, white or cream, yellowing with age; margin inrolled, in contact with stipe. Universal veil on pileus usually absent. Gleba loculate, white to cream. Stipe subcylindric, apex hardly extending into gleba or extending almost to top of gleba. Partial veil absent. Universal veil at stipe base a small cup or absent. Basidiospores statismosporic, inamyloid or very slightly amyloid, wall slightly thickened, smooth. Basidia thin-walled, collapsing early, thick-walled crassobasidia sometimes present. Clamp connections absent.
Known from southern Australia on sandy soil, associated with Agonis, Allocasuarina, Corymbia, Eucalyptus, Hibbertia, Jacksonia, Melaleuca, Mirbelia, Podocarpus and Taxandria.
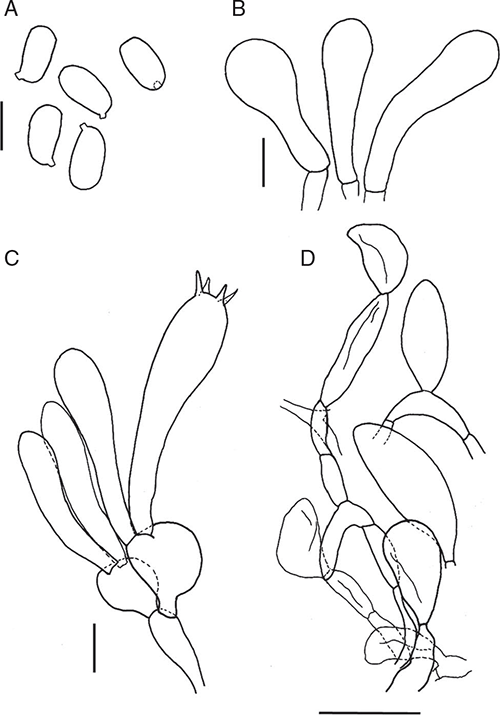
Amanita arenaria (O.K.Mill. & E.Horak) Justo, Mycologia 84(1): 65–69 (1992)
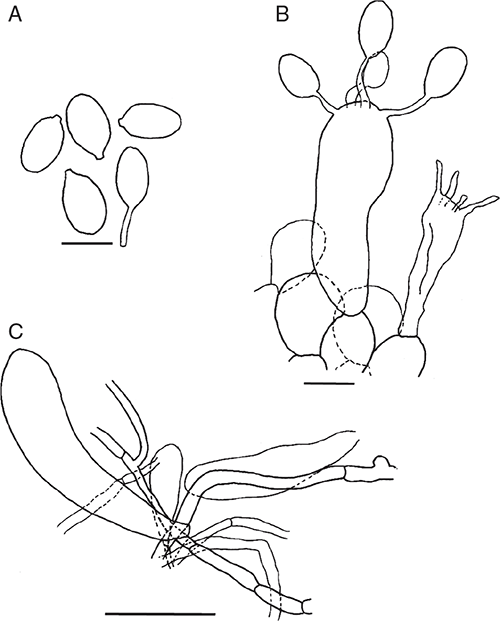
Basidiomata sequestrate, secotioid, capitate, very small. Pileus white to ivory, globose to pulvinate, margin incurved. Gleba chambered, white to cream. Stipe cylindric, hardly reaching into gleba, solid, scabrous, white. Universal veil on pileus as thin white flakes, usually missing; at stipe base a small, white to cream shallow cup. Spores ellipsoid to elongate, inamyloid. Clamp connections absent.
Type: AUSTRALIA. Western Australia: city of Albany, Two Peoples Bay Nature Reserve, 34°57′S, 118°10′E, 22 June 1989, O.K. Miller OKM 24013, H.H. Miller & N.L. Bougher E697 (holo: VPI 679, iso: PERTH 07586329!).
Mycobank number: MB515037.
Published sequence
Type: nuLSU (GQ925382).
Illustration
Miller and Horak (1992, fig. 5–7); Bougher and Syme (1998, pp. 174–175).
Description
Basidiomata sequestrate, secotioid, capitate, very small. Pileus 5–16 mm wide, up to 4 mm thick, white to ivory (B) without surface staining or bruising, globose to pulvinate, surface dry, silky, smooth to lumpy, margin incurved. Universal veil on pileus adnate, thin, membranous flakes, usually missing, white. Gleba white to ivory to cream (B–D), chambers round to elongate. Stipe 3–33 mm × 3–6 mm, cylindric or narrowing downward, apex domed, white; surface scabrous or smooth; context solid, white to ivory (B), unchanging. Universal veil at base of stipe a small broad membranous cup up to 0.5 mm thick with smooth, dry inner surface, white to cream. Odour not distinctive or of mushrooms or coconut.
Basidiospores [160/8/8] (10–)10.5–14(–14.5) × (6.5–)7–8.5(–9) μm, (L = 11.4–13.4 μm; L′ = 12.2 μm; W = 7.1–8.2 μm; W′ = 7.5 μm; Q = (1.25–)1.44–1.86(–2.00); Q = 1.54–1.70; Q′ = 1.62), colourless, thick-walled, smooth, inamyloid or very slightly amyloid, ellipsoid to elongate, contents monoguttulate; apiculus terminal occasionally sublateral, tapered or cylindric, 0.5–1 × 1–1.5 μm, rounded.
Peridium up to 20 μm thick, colourless to pale yellow, not highly gelatinised; filamentous hyphae 3–15 μm wide, walls slightly thickened, colourless, periclinal orientation, gelatinising; inflated cells and vascular hyphae not seen. Pileus context consisting of dominant or equal filamentous hyphae, inflated cells and very infrequent vascular hyphae; filamentous hyphae 3–15 μm wide, with widest hyphae constricted at septa, slightly thickened walls, colourless, gelatinising; inflated cells up to 120 × 25 μm, walls thin or slightly thickened, gelatinising, clavate or ovoid or ventricose or ellipsoidal or spherical or cylindrical, colourless; vascular hyphae 2–7 μm wide, occasionally branched, pale yellow. Dissepiments of dominant filamentous hyphae and inflated cell (vascular hyphae not seen); filamentous hyphae 4–30 μm wide, with widest hyphae constricted at septa, walls thin or slightly thickened, colourless, gelatinising; inflated cells up to 110 × 35 μm, walls thin or slightly thickened, gelatinising, clavate or ovoid or ventricose or ellipsoidal or spherical, colourless. Subhymenium cellular, up to 40 μm wide, of up to 4 cell layers, with basidia arising terminally from ovoid or clavate or globose terminal segments up to 13 μm wide. Basidia: [30/2/2] (23–)24–46(–50) × 9–14 μm, thin-walled, colourless, collapsing early, ~67% 4-spored, ~13% 3-spored, ~20% 2-spored, sterigmata up to 15 × 2 μm, cylindrical; crassobasidia present. Universal veil on pileus not layered, elements with periclinal orientation, consisting of filamentous hyphae and frequent to very infrequent inflated cells (vascular hyphae not observed); filamentous hyphae 4–12 μm wide, with thin or slightly thickened walls, colourless, gelatinising; inflated cells up to 80 × 20 μm when ventricose or up to 150 × 35 μm when clavate or up to 20 × 10 μm when ovoid, with thin or slightly thickened walls, terminal, colourless, gelatinising. Universal veil at stipe base layered, elements with axial orientation. Outer layer narrow, compact, consisting of filamentous hyphae (inflated cells and vascular hyphae not observed), 3–10 μm wide, thick-walled, colourless or pale yellow, gelatinising, often interlocking. Inner layer wide, loose, consisting of dominant or equal filamentous hyphae, inflated cells and very infrequent vascular hyphae; filamentous hyphae 4–17 μm wide, thick-walled, colourless or pale yellow, gelatinising; inflated cells up to 85 × 25 μm when clavate or up to 85 × 50 μm when ovoid or up to 50 × 50 μm when spherical, thin-walled, colourless, gelatinising; vascular hyphae 4 μm wide, pale yellow. Stipe context longitudinally acrophysalidic, consisting of filamentous hyphae, dominant acrophysalides and very infrequent vascular hyphae; filamentous hyphae 2–10 μm wide, colourless or pale yellow, gelatinising; acrophysalides up to 400 × 40 μm clavate, terminal, colourless or pale yellow, gelatinising; vascular hyphae 2–15 μm wide, occasionally branched, pale yellow. Clamp connections not seen.
Habit, habitat and distribution
Gregarious or scattered, buried in sandy soil, with only the pileus showing. In native vegetation, nearby plants include Agonis flexuosa, A. parviceps, Allocasuarina fraseriana, Corymbia calophylla, Eucalyptus marginata, E. megacarpa, Hakea sp., Jacksonia horrida, Podocarpus drouynarius and Taxandria parviceps. Occurs in the Warren WAR01 IBRA region (Department of the Environment 2013). Fruiting period is June to August.
Notes
On the basis of their nuLSU sequences and spore shape, Justo et al. (2010) suggested that the A. arenaria collections of Miller and Horak (1992) represent two cryptic species. Our analysis supports this conclusion. The nuLSU sequence GQ925382 from the holotype of A. arenaria forms a well-supported clade with other collections from the City of Albany and shire of Denmark, on the southern coast of WA (Fig. 1); their β-tubulin and ITS sequences also form well supported clades when compared with other secotioid species (Fig. 2C, 3). All collections of A. arenaria have ellipsoid to elongate spores (Q = 1.54–1.70).
Specimens examined
WESTERN AUSTRALIA. City of Albany, O.K. 24012, H.H. Miller & N.L. Bougher E696 (PERTH 07586310); loc. id., O.K. Miller 24018 & H.H. Miller (PERTH 02241757); loc. id., O.K. 24012, H.H. Miller & N.L. Bougher E696 (PERTH 07586310); loc.id., K. Syme 87/91 & M. Hart (PERTH 04987322); loc. id., K. Syme 428/92 & M. Hart (PERTH 05372178); loc.id., K. Syme 753/94, M. Hart & N.L. Bougher H 7333 (PERTH 05505968); loc.id., KS 3059 (PERTH 09316213); shire of Denmark, K. Syme 1361/04 (PERTH 07367813, MEL 2292314); loc. id., I.C. Tommerup, K. Syme, M.E. Bougher, N.L. Bougher H 7362 (PERTH 07571925).

|
|
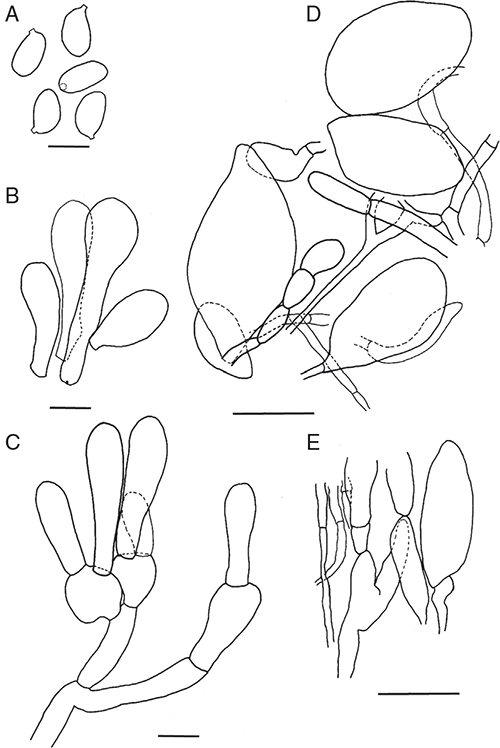
Amanita pseudoarenaria E.M.Davison, Giustiniano & Bougher, sp. nov.
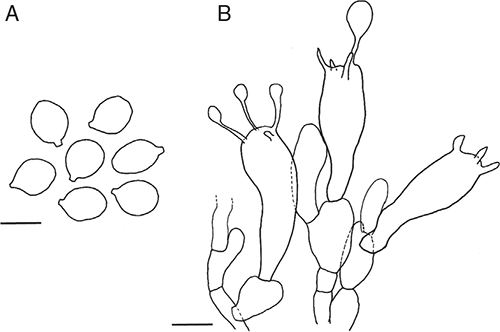
Basidiomata sequestrate, secotioid, capitate, very small. Pileus white to cream, sometimes yellow stained, globose to pulvinate, umbilicate. Gleba chambered, white to cream. Stipe cylindric, hardly reaching into gleba, solid, scabrous, white to cream, sometimes staining yellow. Universal veil a shallow cup at stipe base, white to cream. Spores subglobose to broadly ellipsoid, inamyloid. Clamp connections absent.
Type: AUSTRALIA. Western Australia: shire of Manjimup, 34°10′30″S, 116°35′41″E, 2 June 2013, N.L. Bougher 1072 (holo: PERTH 08606765!).
Mycobank number: MB839131.
Published sequences
Type: ITS (MW775312), nuLSU (MW775284), ef1-α (MW820656), rpb2 (MW820678), β-tubulin (MW820669).
Description
Basidiomata sequestrate, secotioid, capitate, very small. Pileus 5–15 mm wide, up to 5 mm thick, white to ivory to cream (B–D; 4A1–4A2), sometimes yellow stained, globose to pulvinate, umbilicate, surface smooth to lumpy. Universal veil on pileus absent. Gleba white to ivory to cream (B–D; 4A1–4A2), chambers elongate. Stipe 5–33 × 3–8 mm, cylindric or narrowing upward or narrowing downward, apex domed, white to ivory (B), sometimes yellow stained, surface scabrous or smooth; context solid, white. Universal veil at base of stipe a cup up to 5 mm high and up to 1 mm thick, white to ivory (B) often missing. Odour none.
Basidiospores [160/8/7] (8.5–)9–12(–14) × (7–)7.5–10(–11) μm, (L = 9.9–11.0 μm; L′ = 10.6 μm; W = 8.1–9.2 μm; W′ = 8.7 μm; Q = (1.05–)1.06–1.37(–1.60); Q = 1.11–1.27; Q′ = 1.22), colourless, thick-walled, smooth, inamyloid or very slightly amyloid, subglobose to broadly ellipsoid, contents monoguttulate; apiculus terminal, occasionally sublateral, tapered or cylindric, 1–2 × 1–2 μm, rounded.
Peridium up to 50 μm thick, colourless; filamentous hyphae 2–15 μm wide, walls slightly thickened, hyaline, contents colourless or very pale yellow, periclinal orientation, gelatinising; inflated cells not seen; vascular hyphae very infrequent, 6 μm wide, pale yellow. Pileus context consisting of dominant, equal or less frequent filamentous hyphae, inflated cells and very infrequent vascular hyphae; filamentous hyphae 2–15 μm wide, with widest hyphae constricted at septa, slightly thickened walled, colourless, gelatinising; inflated cells up to 135 × 30 μm thin-walled, clavate or ovoid or ventricose or ellipsoidal or spherical, colourless, gelatinising; vascular hyphae 5–8 μm wide, pale yellow. Dissepiments of filamentous hyphae 3–15 μm wide, walls thin or slightly thickened, colourless; inflated cells and vascular hyphae not seen. Subhymenium ramose, with basidia arising terminally from barely inflated to barrel-shaped terminal segments up to 12 μm wide. Basidia [55/4/4] (18–)30–47(–54) × (6–)9–13(–15) μm, thin-walled, colourless, collapsing early, ∼42% 4-spored, ∼29% 3-spored, ∼22% 2-spored, ∼7% 1-spored, sterigmata up to 10 × 2 μm, cylindrical; crassobasidia present. Universal veil at stipe base layered, elements with axial orientation. Outer layer narrow, elements sometimes interlocking, consisting of dominant filamentous hyphae and very infrequent inflated cells (vascular hyphae not seen); filamentous hyphae 2–10 μm wide, thick-walled, colourless or pale yellow, gelatinising; inflated cells up to 90 × 25 μm when clavate or up to 35 × 18 μm when ovoid or up to 30 × 30 μm when spherical, colourless. Inner layer wide, loose, consisting of dominant or frequent, inflated cells and very infrequent vascular hyphae; filamentous hyphae 3–15 μm wide, with widest hyphae constricted at septa, colourless or pale yellow, gelatinising; inflated cells up to 300 × 35 μm when clavate or 75 × 55 μm when ovoid or 40 × 40 μm when spherical or 35 × 18 μm when ellipsoidal or 140 × 50 μm when ventricose or 40 × 20 μm when pyriform, colourless or pale yellow, gelatinising; vascular hyphae 4–12 μm wide, pale yellow. Stipe context longitudinally acrophysalidic, consisting of filamentous hyphae, dominant acrophysalides and very infrequent vascular hyphae; filamentous hyphae 3–12 μm wide, colourless, gelatinising; acrophysalides up to 440 × 35 μm when clavate or 375 × 35 μm when cylindric or 150 × 35 μm when ventricose, colourless or pale yellow, gelatinising; vascular hyphae 2–7 μm wide, occasionally branched, pale yellow. Clamp connections not seen.
Habit, habitat and distribution
Gregarious, initially buried in sand or clayey sand or clayey lateritic gravel or loam, emerging as the stipe elongates. In native vegetation, nearby plants include Agonis linearifolia, A. parviceps, Corymbia calophylla, Eucalyptus marginata, E. patens, Hibbertia sp. and Mirbelia dilatata. Occurs in the Swan Coastal Plain Perth SWA02, Northern Jarrah Forest JAF01, Southern Jarrah Forest JAF02, and Warren WAR01 IBRA subregions (Department of the Environment 2013). Fruiting period is May to August.
Etymology
From the Greek pseudo- meaning ‘false’, referring to its similarity to A. arenaria.
Notes
This is the cryptic secotioid species, which was discussed by Justo et al. (2010). The nuLSU sequences form a well supported clade, as do ef1-α, rpb2, β-tubulin and ITS sequences that differ from those of A. arenaria (Fig. 1, 2A–C, 3, Table 3). This species is widespread in the south-west of WA. It is, in macroscopic appearance, similar to A. arenaria, but can be distinguished from it microscopically by its subglobose to broadly ellipsoid (Q = 1.11–1.27), not ellipsoid to elongate (Q = 1.54–1.70) spores. It differs from secotioid species in section Amarrendiae, and from A. torrendii Justo because of the absence of clamp connections. On this basis A. pseudoarenaria is described as a new species.
Some collections produce a bright yellow liquid that dries as a yellow stain where they have been damaged by insects (Fig. 6). This is not specific to A. pseudoarenaria because it has been observed on some collections of at least one additional, yet to be described, secotioid species from this section (E. M. Davison, pers. obs.).
Specimens examined
WESTERN AUSTRALIA. City of Albany, K. Syme 428/92 (PERTH 05372178); shire of Busselton, E.M. Davison 61-2015 & P.J.N. Davison (PERTH 09316167); shire of Manjimup, E.M. Davison 20-2013 & P.J.N. Davison (PERTH 09316175); shire of Mundaring, E.M. Davison 46-2015 & P.J.N. Davison (PERTH 09316191); shire of Serpentine–Jarrahdale, E.M. Davison 19-2014 & P.J.N. Davison (PERTH 09316183); loc. id. E.M. Davison 25-2020 & P.J.N. Davison (PERTH 09316205).

|
|
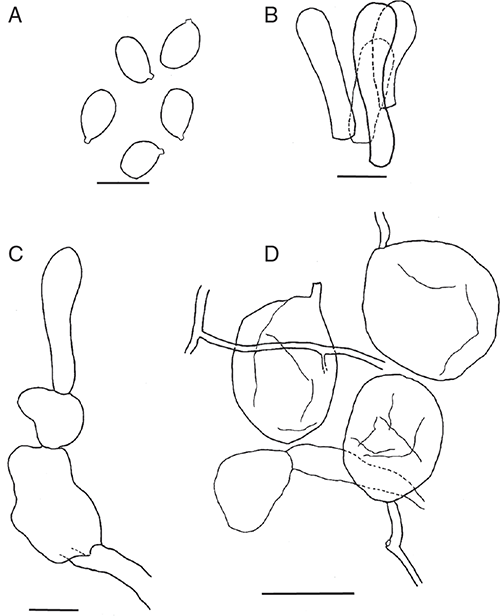
Amanita arenarioides Bougher, E.M.Davison & Giustiniano, sp. nov.
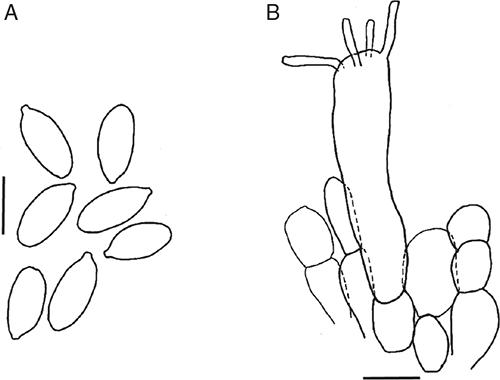
Basidiomata sequestrate, secotioid, capitate, very small. Pileus white or ivory white, bruising dull yellow, globose to pulvinate, margin incurved. Gleba chambered, white or ivory. Stipe cylindric, hardly reaching into gleba, solid, white, staining yellow. Universal veil on pileus not seen; at stipe base a small, shallow cup. Spores elongate to cylindric, inamyloid. Clamp connections absent.
Type: AUSTRALIA. City of Wanneroo, Yanchep National Park, 31°30′S, 115°42′E, 13 June 1997, N.L. Bougher & R.T. Wills H 7501 (holo: PERTH 07627025!).
Mycobank number: MB839132.
Published sequences
Type: ITS: (MW775309), nuLSU (MW775283), ef1-α (MW820655), rpb2 (MW820678), β-tubulin (MW820664).
Description
Basidiomata sequestrate, secotioid, capitate, very small. Pileus 5–16 mm wide, up to 8 mm thick, white, or ivory white, bruising dull yellow, globose to pulvinate, margin incurved, clasping, surface dry. Gleba chambered, chambers rounded, white or ivory. Stipe 7–15 × 2–6 mm, cylindric, white bruising dull yellow, surface smooth or with recurved scales; context solid, white. Universal veil a small basal cup up to 3 mm high. Odour none.
Basidiospores [160/8/2] (9–)10.5–14(–15) × (5–)5.5–7 μm, (L = 10.7–12.8 μm, L′ = 12.2 μm; W = 5.8–6.5 μm, W′ = 6.2 μm; Q = (1.50–)1.69–2.33(–2.60); Q = 1.85–2.13, Q′ = 1.99) colourless, wall slightly thickened, smooth, not adaxially flattened, inamyloid or very slightly amyloid, elongate to cylindric, contents monoguttulate; apiculus terminal or slightly sublateral, tapered, ∼1 × 1.5 μm, rounded or truncate.
Peridium up to 10 μm thick, not continuous, very pale yellow; filamentous hyphae 3–5 μm wide, walls slightly thickened, periclinal orientation, gelatinising; inflated cells not observed; vascular hyphae 6 μm wide, pale yellow. Pileus context consisting of dominant or frequent filamentous hyphae, inflated cells and very infrequent vascular hyphae; filamentous hyphae 4–18 μm wide, with widest hyphae constricted at septa, thin or slightly thickened walls, colourless, gelatinising; inflated cells up to 70 × 30 μm, thin or slightly thickened walls, ellipsoid or clavate or ovoid or cylindric, terminal or in chains of 2; vascular hyphae 3–5 μm wide, pale yellow. Dissepiments of dominant filamentous hyphae encircling chambers, frequent to infrequent inflated cells and very infrequent vascular hyphae; filamentous hyphae 3–12 μm wide, thin-walled, colourless, gelatinising; inflated cells up to 50 × 20 μm thin or slightly thickened walls, ovoid or ventricose or clavate or cylindrical or spherical, colourless; vascular hyphae 4–6 μm wide, pale yellow. Subhymenium inflated, with basidia arising terminally from segments up to 15 μm wide, basidioles frequent. Basidia [24/2/2] (30–)32–45 × (8–)9–12 μm, thin-walled, collapsing, colourless ∼67% 4-spored, ∼17% 3-spored, ∼8% 2-spored, ∼8% 1-spored, sterigmata up to 10 × 2 μm. Universal veil on pileus absent. Universal veil at stipe base layered with axial orientation; outer layer very narrow, of filamentous hyphae (inflated cells and vascular hyphae not observed), 3–15 μm wide, thick, pale yellow walls, gelatinising; inner layer wide, of dominant filamentous hyphae and frequent inflated cells (vascular hyphae not observed), filamentous hyphae 2–15 μm wide, thick, pale yellow gelatinising walls; inflated cells up to 85 × 25 μm when clavate or 70 × 60 μm when ellipsoidal or 40 × 35 μm when ovoid or to 75 × 20 μm when ventricose or 25 × 25 μm when spherical, thick, pale yellow gelatinising walls. Stipe context longitudinally acrophysalidic, consisting of frequent filamentous hyphae, dominant acrophysalides and very infrequent vascular hyphae; filamentous hyphae 3–12 μm wide, slightly thickened, pale yellow gelatinising walls; acrophysalides up to 240 × 40 μm, slightly thickened, pale yellow gelatinising walls, clavate; vascular hyphae 3–8 μm wide, occasionally branched, pale yellow. Clamp connections not seen.
Habit, habitat and distribution
Gregarious, barely emerging from sand. In native vegetation, nearby plants include Banksia grandis, Eucalyptus marginata. Occurs in the Swan Coastal Plain SWA02 IBRA subregion (Department of the Environment 2013). Fruiting period is June to July.
Etymology
From the Latin -oideus meaning ‘like’, referring to its similarity to A. arenaria.
Notes
The nuLSU sequences form a well-supported clade, as do ef1-α and ITS sequences that differ from those of the secotioid species A. arenaria and A. pseudoarenaria (Fig. 1, 2A–C, 3, Table 3). This species has been found only north of Perth. It is, in macroscopic appearance, similar to A. arenaria and A. pseudoarenaria, apart from the basidiomes slowly developing a dull yellowish colour after handling. It can be distinguished from A. arenaria and A. pseudoarenaria microscopically by its elongate to cylindric (Q = 1.85–2.13), not ellipsoid to elongate (Q = 1.54–1.70, A. arenaria) or subglobose to broadly ellipsoid (Q = 1.11–1.27, A. pseudoarenaria), spores. It differs from secotioid species in section Amarrendiae, and from A. torrendii because of the absence of clamp connections. On this basis, A. arenarioides is described as a new species.
Specimens examined
WESTERN AUSTRALIA. Shire of Gingin, I.C.P. Tommerup H 7508 (PERTH 07626959).

|
|
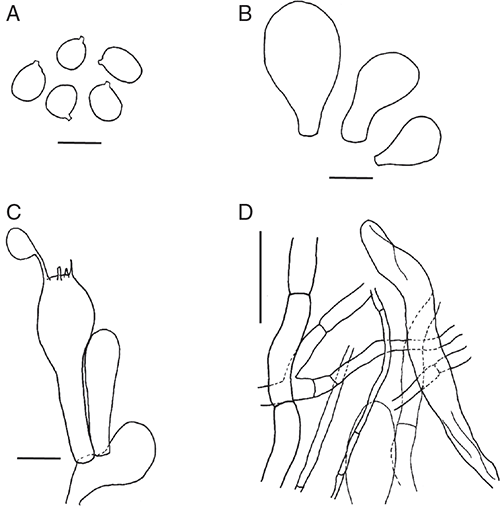
Amanita compacta Bougher, E.M.Davison & Giustiniano, sp. nov.
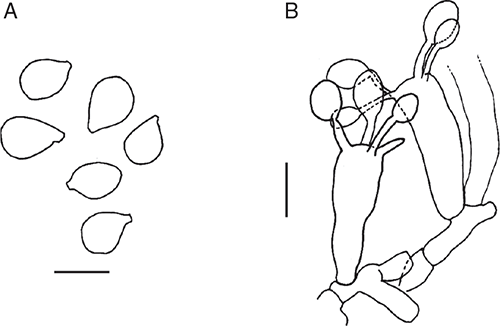
Basidiomata sequestrate, secotioid, cylindric, very small. Pileus white, unchanging, thimble-shaped, thin, clasping. Gleba chambered, white or ivory. Stipe cylindric, almost reaching top of pileus, solid, white. Universal veil absent. Spores inamyloid, broadly ellipsoid to ellipsoid. Clamp connections absent.
Type: AUSTRALIA. Western Australia: Shire of Wongan–Ballidu, Elphin Nature Reserve, 30°51′43″S, 116°41′34″E, 10 Aug. 2015, N.L. Bougher 1180 (holo: PERTH 08700524!).
Mycobank number: MB839133.
Published sequences
Type: ITS (MW775277), nuLSU (MW775285), ef1-α (MW820657), rpb2 (MW820679), β-tubulin (MW820665).
Description
Basidiomata sequestrate, secotioid, cylindric, very small. Pileus 7–11 mm wide, up to 3 mm thick, white, without surface staining or bruising, thimble-shaped, covering stipe apex, clasping, surface dry. Gleba with elongate chambers, white or ivory. Stipe 15–18 × 5–11 mm, cylindric tapered within pileus, white, surface smooth; context solid, white. Partial veil absent. Universal veil absent on pileus and at stipe base. Odour none.
Basidiospores [80/4/1] 9–12(–13) × (6–7.5(–8.5) μm, (L = 1.40–1.63 μm; L′ = 10.6 μm; W = 6.8–7.2 μm; W′ = 7.0 μm; Q = (1.18–)1.33–1.83(–1.92); Q = 1.40–1.63; Q′ = 1.53) colourless, slightly thickened wall, smooth, not adaxially flattened, some distorted, inamyloid or very slightly amyloid, broadly ellipsoid to ellipsoid to elongate, contents monoguttulate; apiculus terminal or slightly sublateral, cylindric or tapered, ∼1 × 1.5 μm, truncate.
Peridium present, not continuous, colourless; filamentous hyphae 4–8 μm wide, with some terminal segments up to 10 μm, walls slightly thickened, periclinal orientation, gelatinising; inflated cells not observed. Pileus context consisting of dominant filamentous and infrequent or very infrequent inflated cells; filamentous hyphae 3–12 μm wide, walls slightly thickened, interlocking in places, colourless, gelatinising; inflated cells up to 70 × 15 μm, slightly thickened walls, clavate or ovoid or ventricose, colourless, gelatinising. Dissepiments of dominant filamentous hyphae encircling chambers and very infrequent inflated cells; filamentous hyphae 3–7 μm wide, thin-walled, colourless; inflated cells up to 55 × 12 μm clavate, colourless. Subhymenium ramose, with basidia arising terminally from segments 4 μm wide, becoming inflated, up to 7 μm. Basidia [20/1/1] (20–)25–36(–37) × (7–)8–11(–15) μm, thin-walled, collapsing, colourless ∼65% 4-spored, ∼20% 3-spored, ∼15% 2-spored, sterigmata up to 7 × 2 μm. Stipe context longitudinally acrophysalidic, consisting of frequent filamentous hyphae and dominant acrophysalides; filamentous hyphae 2–6 μm wide, slightly thickened walls, colourless, gelatinising; acrophysalides up to 300 × 35 μm, slightly thickened walls, clavate or cylindric, colourless, gelatinising. Vascular hyphae not seen. Clamp connections not seen.
Habit, habitat and distribution
Gregarious, barely emerging from sand. In native vegetation, nearby plants Allocasuarina campestris. Occurs in the Lesueur Sandplain GES02 IBRA subregion (Department of the Environment 2013). Fruiting period is August.
Etymology
From the Latin compactus meaning ‘close-packed’, referring to the compact, rather stumpy appearance of the basidiomata.
Notes
Only one collection of this distinctive species is available. It differs macroscopically from the secotioid species A. arenaria, A. pseudoarenaria and A. arenarioides because the basidiomes are cylindric, not capitate, and the gleba is thimble-shaped, not globose to pulvinate. Also, the universal veil is absent. Microscopically it differs from A. pseudoarenaria and A. arenarioides because the spores are ellipsoid to elongate (Q = 1.40–1.63), not subglobose to broadly ellipsoid (Q = 1.11–1.27, A. pseudoarenaria) or elongate to cylindric (Q = 1.85–2.13). It differs from secotioid species in section Amarrendiae, and from A. torrendii because of the absence of clamp connections. The nuLSU, ef1-α and ITS sequences differ from those of A. arenaria, A. pseudoarenaria and A. arenarioides (Fig. 1, 2A 3, Table 3). On this basis, A. compacta is described as a new species.

|
|
Amanita pupatju E.M.Davison, Giustiniano, McGurk & E.L.J.Watkin, sp. nov.
Basidiomata agaricoid, very small to large. Pileus white. Universal veil on pileus white, felted to floccose patches over disc, not evident at base of stipe. Lamellae white. Stipe white with mealy or floccose surface and conic bulb. Partial veil arachnoid, superior, white. Spores amyloid, elongate to cylindric. Universal veil not layered, with no dominant orientation, composed of terminal inflated cells that are occasionally in short chains. Clamp connections absent.
Type: AUSTRALIA. Western Australia: city of Melville, Murdoch University grounds, 32°47′27″S, 115°50′04″E, 31 May 2015, E.M. Davison 9–2015 & P.J.N. Davison, (holo: PERTH 09269525!).
Mycobank number: MB839134.
Published sequences
Type: ITS (MW775301), nuLSU (MW775287), ef1-α (MW820659), β-tubulin (MW820668).
Description
Pileus 22–115 mm wide, up to 6 mm thick, white to very pale clay pink (6A2) with age, without surface staining or bruising, initially convex becoming plane with slightly depressed centre, surface tacky when moist; margin non-striate, appendiculate. Universal veil on pileus adnate, thin, felted to floccose, breaking into small patches over disc, white. Lamellae adnexed to adnate, subcrowded, white, 5–10 mm broad, margin concolorous, fimbriate; lamellulae plentiful to infrequent in several lengths, rounded truncate or truncate or rounded attenuate or attenuate. Stipe 35–75 mm long, 8–40 mm wide, cylindrical or tapering upward or tapering downward, white, surface mealy or floccose or fibrillose. Partial veil superior, arachnoid, white. Bulb 14–35 × 16–39 mm, conic to tapered, occasionally ovoid when young, white to grey. Remains of universal veil at top of bulb not evident, or as a slight ridge. Pileus and stipe context white, unchanging, stipe solid becoming hollow. Smell none when young, ammoniac when old. Spore deposit white or B (ivory).
Basidiospores [200/10/8] (8.5–)9.5–13.5(–17) × (4.5–)5–7(–7.5) μm, (L = 10.6–13.6 μm; L′ = 11.5 μm; W = 5.2–6.5 μm; W′ = 5.8 μm; Q = (1.55–)1.67–2.33(–3.00); Q = 1.80–2.12; Q′ = 1.98), colourless, thin-walled, smooth, amyloid, ellipsoid to elongate to cylindrical, contents monoguttulate or granular; apiculus sublateral, cylindric or tapered, ~1 × 1 μm, rounded or truncate.
Pileipellis up to 300 μm thick in old specimens, with a colourless gelatinised suprapellis up to 200 μm thick and colourless subpellis, consisting of filamentous hyphae and infrequent vascular hyphae (inflated cells not observed), filamentous hyphae 3–15 μm wide, thick-walled, walls hyaline, gelatinising, contents colourless, radially orientated with some interweaving; vascular hyphae 3–6 μm wide, pale yellow or yellowish-brown. Pileus context consisting of filamentous hyphae (dominant), inflated cells and very infrequent vascular hyphae, filamentous hyphae 3–40 μm wide, with widest hyphae constricted at septa, thin-walled, colourless; inflated cells to 250 × 40 μm, thin-walled, clavate or ventricose or ovoid, terminal, colourless; vascular hyphae 3–11 μm wide, pale yellow. Lamella trama bilateral, divergent. Central stratum up to 40 μm wide, consisting of filamentous hyphae (inflated cells and vascular hyphae not observed), filamentous hyphae 3–10 μm wide, thin-walled, colourless. Subhymenial base with angle of divergence ~30° from central stratum, with filamentous hyphae following smooth broad curve to subhymenium, consisting of dominant filamentous hyphae, inflated cells (infrequent to frequent) and very infrequent vascular hyphae; filamentous hyphae 3–20 μm wide, widest close to subhymenium and constricted at septa, frequently branched, thin-walled, colourless; inflated cells up to 200 × 20 μm when clavate or up to 160 × 12 μm when ventricose or up to 50 × 18 μm when elliptic or up to 60 × 20 μm when cylindric, terminal, colourless; vascular hyphae 3–5 μm wide, occasionally branched, pale greenish-yellow. Subhymenium with basidia arising terminally from pyriform to spherical terminal segments up to 24 μm wide. Lamella edge tissue sterile, with inflated cells infrequent to frequent, up to 40 × 11 μm when clavate or up to 35 × 11 μm when capitate or up to 30 × 14 μm when broadly clavate or up to 30 × 14 μm when pyriform or up to 27 × 7 μm when cylindric, colourless. Basidia [140/7/7] (35–)38–57(–65) × (9–)10–13(–14) μm, thin-walled, colourless, ∼93% 4-spored, ∼3% 3-spored, ∼4% 2-spored, sterigmata to 8 × 2 μm. Universal veil on pileus not layered, elements with no dominant orientation, consisting of frequent to dominant filamentous hyphae, dominant to frequent inflated cells and very infrequent vascular hyphae, filamentous hyphae 5–15 μm wide, with widest hyphae sometimes constricted at septa, colourless, some gelatinising; inflated cells up to 90 × 75 μm when ovoid or up to 220 × 55 μm when ventricose or up to 100 × 100 μm when spherical or up to 80 × 60 μm when elliptic or up to 95 × 25 μm when cylindrical, terminal or in chains of up to 3, colourless, some gelatinising; vascular hyphae 6–12 μm wide, occasionally branched, pale yellow. Universal veil at stipe base absent. Stipe context longitudinally acrophysalidic, consisting of frequent filamentous hyphae, dominant acrophysalides and very infrequent to frequent vascular hyphae, filamentous hyphae 2–8 μm wide, colourless; acrophysalides to 420 × 35 μm clavate or cylindric, terminal, colourless or pale yellow, gelatinising; vascular hyphae 3–10 μm wide, occasionally branched, pale yellow or pale brownish-yellow. Partial veil not examined. Clamp connections not observed.
Habit, habitat and distribution
Singly or gregarious in sand. In degraded native vegetation, nearby plants include Corymbia calophylla and Eucalyptus marginata. Occurs in the Swan Coastal Plain Perth SWA02 IBRA subregion (Department of the Environment 2013). Fruiting period is April to June.
Etymology
Pupatju means ‘to cover the head’ in the Nhanda dialect of the Western Australian Aboriginal Noongar language, reflecting the patches of universal veil on the pileus. The epithet is formed as a noun in apposition.
Notes
Figure 1 shows A. pupatju is a member of section Arenariae. There are several species with a white or pale pileus, in which clamp connections are either absent or very infrequent. Amanita pupatju has similar shaped, but longer, spores than does A. preissii (A. pupatju [200/10/8] L = 10.6–13.6 μm, W = 5.2–6.5 μm, Q = 1.80–2.12; A. preissii [340/11/11] L = 8.8–11.5 μm, W = 5.2–6.0 μm, Q = 1.65–2.13), but the pileus, universal veil and partial veil do not age pale cinnamon to saffron (Davison et al. 2017b). It has spores of shape and size similar to those of A. lesueurii ([200/10/5] L = 11.1–12.4 μm, W = 5.5–6.2 μm, Q = 1.90–2.26), but lacks the felted or floccose universal veil on the pileus which becomes vinaceous-buff to grey with age (Davison et al. 2013). Amanita pupatju has spores that are wider than those of A. wadulawitu ([859/21/21] L = 10.2–12.5 μm, W = 4.4–5.2 μm, Q = 2.21–2.55) and a much smaller and less well defined basal bulb (McGurk et al. 2016). It differs from A. clelandii (E.-J. Gilbert) E.-J. Gilbert, and A. variabilis (E.-J. Gilbert) E.-J. Gilbert & Cleland, two poorly known species from South Australia, by its larger size and different-shaped spores (Reid 1980). The nuLSU sequence, ef1-α, β-tubulin show that it clusters with A. griselloides (Fig. 1, 2A, 2C), but the ITS sequences differ by 12.2–13.8% (Table 4). Amanita pupatju differs from A. griselloides because it has a white, not pale grey or brown, pileus and the inflated cells in the universal veil on the pileus have colourless, not brown, contents (Reid 1980; see below). On this basis, Amanita pupatju is described as a new species.
Specimens examined
WESTERN AUSTRALIA. City of Melville, E.M. Davison 22-1990 & P.J.N. Davison (PERTH 03097129); loc. id., E.M. Davison 37-1990 & P.J.N. Davison (PERTH 03096955); loc. id., E.M. Davison 52-1990 & P.J.N. Davison (PERTH 03097080); loc. id., E.M. Davison 8-1991 & P.J.N. Davison (PERTH 03096726); loc. id., E.M. Davison 11-1991 & P.J.N. Davison (PERTH 03096947); loc. id., E.M. Davison 15-1992 & P.J.N. Davison (PERTH 03096998); loc. id., E.M. Davison 3-1995 & P.J.N. Davison (PERTH 09269479); loc. id., L.E. McGurk 41–2005 (PERTH 09269533).

|
|
Amanita sabulosa E.M.Davison & Giustiniano, sp. nov.
Basidiomata agaricoid, small to medium. Pileus white to ivory, aging pale smoke grey to pale vinaceous-buff to milky coffee to clay buff to drab, paler at margin, margin appendiculate. Universal veil on pileus white to ivory to pale vinaceous-buff, thin, felted, crustose or as flat patches. Lamellae white to ivory. Stipe white. Partial veil superior, white, fugacious. Universal veil at stipe base a cup forming a globose to subglobose marginate bulb. Spores amyloid, elongate to cylindrical. Universal veil of equal filamentous hyphae and inflated cells. Clamp connections absent.
Type: Australia, Western Australia: city of Melville, Murdoch University grounds, 32°4′27″S, 115°14′20″E, 28 May 2017, E.M. Davison 34-2017 &P.J.N. Davison (holo: PERTH 09178759!).
Mycobank number: MB839135.
Published sequences
Type: ITS (MW775291), nuLSU (MW775279), ef1-α (MW820650), rpb2 (MW820675), β-tubulin (MW820660).
Description
Pileus 27–90 mm wide, up to 9 mm thick, white to ivory, aging pale cream to pale smoke grey to pale vinaceous-buff to milky coffee to clay buff to drab (5A2–4B2–5B2–6C3–6E3) with pale margin, without surface staining or bruising, initially convex becoming plane with slightly depressed centre, surface tacky when moist; margin non-striate, appendiculate. Universal veil on pileus adnate, thin, felted, crustose or breaking into flat patches over disc, white to ivory to very pale vinaceous-buff (5B2). Lamellae adnate to adnexed to free, close to subcrowded, white or ivory, 2–16 mm broad, margin concolorous, fimbriate; lamellulae plentiful, in several lengths, rounded truncate or truncate or rounded attenuate or attenuate. Stipe 26–60 mm long, 10–20 mm wide, cylindrical, or tapering upward, white, surface smooth or floccose or fibrillose below partial veil. Partial veil superior, descendent, striate above, soft to arachnoid, fugacious, white. Universal veil at stipe base a cup, forming a marginate globose to rounded bulb, 10–18 × 12–22 mm, sometimes with free limb up to 5 mm high, white. Pileus and stipe context white, unchanging, stipe solid. Smell none. Spore deposit white or B (ivory).
Basidiospores [220/11/11] (8.5–)10–12.5(–13) × (4.5–)5–7(–7.5) μm, (L = 9.8–11.8 μm; L′ = 11.0 μm; W = 5.2–6.4 μm; W′ = 6.0 μm; Q = (1.39–)1.55–2.10(–2.44); Q = 1.60–2.06; Q′ = 1.84), colourless, thin-walled, smooth, amyloid, elongate to cylindrical occasionally ellipsoid, contents monoguttulate or granular; apiculus sublateral, cylindric or tapered, ~1 × 1 μm, rounded or truncate.
Pileipellis up to 150 μm thick in old specimens, with a colourless gelatinised suprapellis up to 80 μm thick and colourless or pale yellow subpellis, consisting of filamentous hyphae and very infrequent vascular hyphae (inflated cells not observed), filamentous hyphae 3–12 μm wide, thick-walled, walls hyaline, gelatinising, contents colourless, radially orientated with some interweaving; vascular hyphae 2–5 μm wide, pale yellow. Pileus context consisting of filamentous hyphae (dominant), inflated cells and very infrequent vascular hyphae, filamentous hyphae 3–40 μm wide, with widest hyphae constricted at septa, thin-walled, colourless; inflated cells to 350 × 25 μm thin-walled ventricose or clavate or cylindrical or ovoid, terminal, colourless; vascular hyphae 3–10 μm wide, occasionally branched, pale yellow. Lamella trama bilateral, divergent. Central stratum up to 20 μm wide, consisting of filamentous hyphae and very infrequent vascular hyphae (inflated cells not observed), filamentous hyphae 2–15 μm wide, with widest hyphae constricted at septa, thin-walled, colourless; vascular hyphae 3–4 μm wide, some sinuous, pale yellow or colourless. Subhymenial base with angle of divergence 20–30° from central stratum with filamentous hyphae following broad curve to subhymenium, consisting of dominant filamentous hyphae, inflated cells (infrequent to frequent) and very infrequent vascular hyphae; filamentous hyphae 2–35 μm wide, widest close to subhymenium and constricted at septa, frequently branched, thin-walled, colourless; inflated cells up to 120 × 30 μm when clavate or up to 120 × 15 μm when cylindric or up to 110 × 25 μm when ventricose or up to 35 × 20 μm when ovoid, terminal, colourless; vascular hyphae 2–10 μm wide, occasionally branched, some sinuous, pale yellow or greenish-yellow. Subhymenium with basidia arising terminally from pyriform to spherical terminal segments, becoming inflated, up to 22 μm wide. Lamella edge tissue sterile, with inflated cells infrequent to frequent, up to 45 × 12 μm when clavate or up to 50 × 20 μm when pyriform or up to 25 × 13 μm when ovoid or up to 80 × 8 μm when cylindric, disarticulating, colourless. Basidia [140/7/7] (38–)40–60(–66) × (9–)10–15(–17) μm, thin-walled, colourless, ∼90% 4-spored, ∼3% 3-spored, ∼7% 2-spored, sterigmata to 7 × 2 μm. Universal veil on pileus layered. Basal layer of elements with somewhat periclinal orientation, consisting of filamentous hyphae (equal or less frequent) and inflated cells (vascular hyphae not seen), filamentous hyphae 3–25 μm wide, with widest hyphae constricted at septa, colourless, some gelatinising; inflated cells up to 110 × 90 μm when ovoid or up to 120 × 50 μm when ellipsoid or up to 70 × 70 μm when spherical or up to 95 × 45 μm when clavate, terminal or occasionally in chains of up to 3, disarticulating, colourless. Superficial layer very narrow, often absent, with periclinal orientation, consisting of filamentous hyphae (inflated cells and vascular hyphae not seen), 3–12 μm wide, colourless, gelatinising. Universal veil at stipe base layered. Inner layer narrow, often absent, of elements with axial orientation, consisting of filamentous hyphae (dominant to equal) and inflated cells (vascular hyphae not seen), filamentous hyphae 3–17 μm wide, some thick-walled, colourless, gelatinising; inflated cells up to 80 × 50 μm when ovoid or up to 150 × 35 μm when clavate, terminal, colourless. Middle layer wide, consisting of filamentous hyphae (dominant to infrequent), inflated cells and very infrequent vascular hyphae, filamentous hyphae 2–12 μm wide, some thick-walled, colourless, gelatinising; inflated cells up to 140 × 140 μm when spherical or up to 105 × 60 μm when ovoid or up to 150 × 50 μm when ellipsoidal or up to 200 × 20 μm when clavate or up to 75 × 35 μm when pyriform, terminal, colourless, disarticulating; vascular hyphae 3–6 μm wide, pale yellow or pale green. Outer layer narrow, often absent, of elements with axial orientation, consisting of dominant filamentous hyphae and frequent to infrequent inflated cells (vascular hyphae not seen), filamentous hyphae 3–20 μm wide, colourless, gelatinising; inflated cells up to 30 × 25 μm when ovoid or up to 50 × 20 μm when clavate, terminal, colourless. Stipe context longitudinally acrophysalidic, consisting of frequent filamentous hyphae, dominant acrophysalides and very infrequent to frequent vascular hyphae, filamentous hyphae 2–10 μm wide, colourless; acrophysalides up to 300 × 40 μm when clavate or up to 500 × 25 μm when cylindric or up to 250 × 35 μm when fusiform, terminal, colourless, gelatinising; vascular hyphae 2–20 μm wide, occasionally branched, pale yellow or pale green. Ornamentation on stipe surface of filamentous hyphae (frequent or equal) and inflated cells (vascular hyphae not seen), filamentous hyphae 2–8 μm wide, colourless, disarticulating; inflated cells up to 150 × 25 μm when clavate or up to 45 × 25 μm when pyriform, terminal, colourless, disarticulating. Partial veil of filamentous hyphae (equal, or frequent), inflated cells and frequent vascular hyphae, filamentous hyphae 3–8 μm, wide, colourless, disarticulating; inflated cells up to 150 × 25 μm clavate, colourless, disarticulating; vascular hyphae 2–8 μm wide occasionally branched, colourless or pale yellow or pale greenish yellow. Clamp connections not observed.
Habit, habitat and distribution
Singly or gregarious in sand, in native vegetation; nearby plants include Allocasuarina fraseriana, Brachyloma preissii, Corymbia calophylla, Eucalyptus marginata, Jacksonia furcellata and Melaleuca lanceolata. Occurs in the Swan Coastal Plain Perth SWA02 and IBRA subregion and the Murray–Darling Depression Murray Mallee MDD02 IBRA subregion (Department of the Environment 2013). Fruiting period is May to July.
Etymology
From the Latin sabulosus meaning ‘sandy’, referring to the sandy soil where it occurs.
Notes
Figure 1 shows A. sabulosa is a member of section Arenariae. The pileus is, in appearance, similar to that of A. pupatju and the spores are of similar shape (A. sabulosa [220/11/11] L = 9.8–11.8 μm, W = 5.2–6.4 μm, Q = 1.60–2.06; A. pupatju [200/10/8] L = 10.6–13.6 μm, W = 5.2–6.5 μm, Q = 1.80–2.12); however, these two species differ in the shape of the stipe base, which is conic to tapered in A. pupatju, whereas A. sabulosa has a marginate bulb (Fig. 12, 14). The nuLSU, ef1-α, β-tubulin sequences show that these two species differ (Fig. 1, 2A, C), and ITS sequences differ by 14.6–18.0% (Table 4).
Amanita sabulosa has spores of similar shape to those of A. lesueurii ([200/10/5] L = 11.1–12.4 μm, W = 5.5–6.2 μm, Q = 1.90–2.26), but lacks the felted or floccose universal veil on the pileus, which becomes vinaceous-buff to grey with age (Davison et al. 2013). The spores of A. sabulosa are wider than those of A. wadulawitu ([859/21/21] L = 10.2–12.5 μm, W = 4.4–5.2 μm, Q = 2.21–2.55) and it has a universal veil, which forms a marginate bulb, not a bulb that is initially turbinate and which elongates with age (McGurk et al. 2016).
There are other species described from Australia that have amyloid spores, a felted universal veil and no clamp connections, that may fall with section Arenariae. Amanita sublutea (Cleland) E.-J.Gilbert is a poorly known species from South Australia. From the descriptions given by Cleland (1931), Reid (1980), Bas (1969) and Grgurinovic (1997), A. sabulosa has a paler pileus, wider stipe, shorter and narrower spores (A. sublutea: [20/2] (11–)11.5–13(–13.5) × 6.5–7.5(–8) μm (Bas 1969), [50/2] 11–13.8 × 5.8–7.7 μm (Grgurinovic 1997); A. sabulosa [220/11/11] (8.5–)10–12.5(–13) × (4.5–)5–7(–7.5) μm), the subhymenium is inflated rather than inflated ramose and its cells are not coralloid. Some of these differences may be the result of environmental conditions; more collections of A. sublutea from the type locality, or sequences from the holotype, would clarify whether these are two different species.
Amanita sabulosa differs from A. gracilenta A.E.Wood by its larger size, more robust form, marginate, not slightly swollen, bulb, presence of a partial veil, shorter and narrower spores (A. gracilenta 8.1–9.9(–11.0) × 4.5–6.1(–6.6) μm; A. sabulosa [200/10/10] (9–)10–12.5(–13) × (4.5–)5–7(–7.5) μm), and larger inflated cells in the universal veil on the pileus (Wood 1997). It differs from A. clelandii and A. variabilis, two poorly known species from South Australia, by its larger size and different-shaped spores (Reid 1980). On this basis, A. sabulosa is described as a new species.
Specimens examined
WESTERN AUSTRALIA. City of Melville, E.M. Davison 10-1995 & P.J.N. Davison (PERTH 09269452); loc. id., E.M. Davison 11-1995 & P.J.N. Davison, (PERTH 09269487); loc. id., E.M. Davison 11-2002 & P.J.N. Davison (PERTH 09178694); loc. id., E.M. Davison 23-2003 & P.J.N. Davison (PERTH 09178708); loc. id., E.M. Davison 15-2014 & P.J.N. Davison (PERTH 09178716); loc. id., E.M. Davison 17-2014 & P.J.N. Davison (PERTH 09178724); loc. id., E.M. Davison 15-2015 & P.J.N. Davison (PERTH 09178732); loc. id., E.M. Davison 41-2016 & P.J.N. Davison (PERTH 09178740); loc. id., E.M. Davison 35-2017 & P.J.N. Davison (PERTH 09178767); loc. id., E.M. Davison 57-2017 & P.J.N. Davison (PERTH 09178775).
SOUTH AUSTRALIA. Monarto Conservation Park, P.S. Catcheside 2509 & D.E.A. Catchside (AD-C54481).

|
|
Amanita griselloides D.A.Reid, Victorian Naturalist 95(2): 47 (1978)
Basidiomata agaricoid, small to medium. Pileus pale grey to grey to pale brown to brown, paler at margin, margin appendiculate. Universal veil on pileus white, becoming greyish-brown to brown, felted to floccose, as one or several patches. Lamellae white. Stipe white with ovoid to conic bulb that elongates with age. Partial veil superior to median, white, fugacious. Universal veil at top of bulb very inconspicuous or absent. Spores amyloid, ellipsoid to elongate. Universal veil on pileus of dominant inflated cells with brown or greyish-brown contents. Clamp connections absent.
Type: AUSTRALIA. Western Australia: 2 km north of Bow River on Highway 1, near Walpole, 34°37′S, 116°58′E, 13 May 1976, D.A & D.G Reid, R. Hilton, N. Brittan (holo: K(M) 236386!; iso: DAR 32037!).
Mycobank number: MB308554.
Description
Pileus 33–65 mm wide, up to 10 mm thick, silvery-grey to smoke grey to silvery-beige to cinnamon to pale brown to brown, margin paler, without surface staining or bruising, initially convex, becoming plane with decurved margin and slightly depressed centre, surface slightly tacky when moist shiny when dry; margin non-striate, slightly appendiculate when young. Universal veil on pileus adnate, felted to floccose, as one or several patches in centre of disc, white to light brown to greyish-brown to brown. Lamellae adnexed to adnate, close to subcrowded, white to pale buff (5B2), 6–10 mm broad, margin concolorous, fimbriate; lamellulae frequent to infrequent in several lengths, shortest truncate or rounded truncate, longest attenuate or subattenuate. Stipe 20–48 mm long, 8–27 mm wide, cylindrical or tapering upwards, white or cream, surface at top of stipe powdery, below smooth or minutely floccose or fibrillose. Partial veil superior to median, descendent, striate above, fugacious or not observed, white or pale buff. Bulb 25–30 × 8–32 mm, ovoid or conic becoming tapered, white or pale buff. Remains of universal veil at top of bulb not apparent or a soft, floccose rim at top of bulb, white. Pileus and stipe context white, unchanging, stipe solid, becoming hollow. Smell not distinctive or unpleasant. Spore deposit white.
Basidiospores [180/9/8] (8–)8.5–12(–12.5) × (5–)5.5–7 μm, (L = 9.2–11.3 μm; L′ = 10.0 μm; W = 5.9–6.6 μm; W′ = 6.2 μm; Q = (1.29–)1.38–1.83(–2.00); Q = 1.51–1.76; Q′ = 1.61), colourless, thin-walled, smooth, amyloid, ellipsoid to elongate, contents monoguttulate or granular; apiculus sublateral, cylindric or tapered, ~1 × 1 μm, truncate or rounded.
Pileipellis poorly developed in young specimens, up to 200 μm thick in old specimens, with a colourless gelatinised suprapellis up to 150 μm thick, and yellow or brown subpellis, consisting of filamentous hyphae and frequent to infrequent vascular hyphae (inflated cells not observed), filamentous hyphae 2–12 μm wide, thick-walled, walls hyaline, gelatinising, contents colourless to yellow to brown, radially orientated; vascular hyphae 2–10 μm wide, occasionally branched, pale yellow or yellowish-brown. Pileus context consisting of filamentous hyphae (dominant or equal), inflated cells and frequent to infrequent vascular hyphae, filamentous hyphae 3–25 μm wide, with widest hyphae constricted at septa, thin-walled, colourless; inflated cells up to 300 × 35 μm, thin-walled, some gelatinising, clavate or ventricose or ovoid or cylindrical, colourless; vascular hyphae 3–15 μm wide, occasionally branched, pale yellow. Lamella trama bilateral, divergent. Central stratum up to 30 μm wide, consisting of filamentous hyphae (inflated cells and vascular hyphae not observed), filamentous hyphae 4–6 μm wide, colourless. Subhymenial base with angle of divergence ~30° from central stratum, with filamentous hyphae following smooth broad curve to subhymenium, consisting of dominant filamentous hyphae, inflated cells (infrequent or not seen; vascular hyphae not seen), filamentous hyphae 3–15 μm wide, widest close to subhymenium and constricted at septa, frequently branched, thin-walled, colourless; inflated cells up to 70 × 20 μm ventricose or cylindric, terminal, colourless. Subhymenium with basidia arising terminally from barely inflated to pyriform terminal segments up to 18 μm wide. Lamella edge tissue sterile, with inflated cells infrequent to frequent, to 20–35 × 10–18 μm clavate or pyriform or ovoid, colourless, disarticulating. Basidia [140/7/7] (31–)35–59(–66) × (8–)9–13(–15) μm, thin-walled, colourless, ∼86% 4-spored, ∼8% 3-spored, ∼6% 2-spored, sterigmata up to 8 × 2 μm. Universal veil on pileus not layered, elements with no dominant orientation, consisting of frequent to dominant filamentous hyphae, dominant to frequent inflated cells and very infrequent to frequent vascular hyphae, filamentous hyphae 3–15 μm wide, colourless or pale brown or pale greyish-brown, gelatinising; inflated cell up to 85 × 70 μm when ovoid or up to 70 × 70 μm when spherical or up to 90 × 50 μm when ellipsoidal or up to 110 × 25 μm when ventricose or up to 110 × 45 μm when clavate, terminal, occasionally in chains of up to 3 cells, colourless or grey or greyish-brown or brown, gelatinising; vascular hyphae 3–12 μm wide, occasionally branched, brownish-yellow. Universal veil on stipe base often absent, not layered, with no dominant orientation, consisting of dominant filamentous hyphae, frequent inflated cells and very infrequent vascular hyphae, filamentous hyphae 4–10 μm wide, thick-walled, colourless or pale grey, gelatinising; inflated cells up to 110 × 20 μm when clavate or up to 110 × 40 μm when pyriform, very pale grey; vascular hyphae 5 μm wide, pale yellow. Stipe context longitudinally acrophysalidic, consisting of frequent to equal filamentous hyphae, acrophysalides and infrequent to frequent vascular hyphae, filamentous hyphae 2–10 μm wide, colourless; acrophysalides up to 190 × 35 μm, clavate or cylindric, terminal, colourless; vascular hyphae 3–35 μm wide, occasionally branched, yellowish-brown. Partial veil not examined. Clamp connections not observed.
Habit, habitat and distribution
Gregarious in sand. In native vegetation; nearby plants include Agonis hypericifolia, Allocasuarina fraseriana, Eucalyptus diversicolor and E. marginata. Occurs in the Southern Jarrah Forest JAF02, Warren WAR01, and Recherche ESP02 IBRA subregions (Department of the Environment 2013). Fruiting period is April to July.
Notes
Reid (1980) described the universal veil on the pileus of A. griselloides as layered, with a grey felty floccose layer of inflated cells with brown contents intermixed with thin-walled, hyaline hyphae, covered by a white arachnoid superficial layer formed entirely of thin-walled, prostrate hyphae. He commented that the structures of the universal veil on the pileus of an additional collection (DAR 32036) were exactly as in the type. A layered universal veil in which there is a superficial layer of mainly filamentous hyphae occurs in some members of subgenus Amanitina (Bas 1969, pp. 307–308). However, in the type K(M) 236386, this superficial layer has phialides, indicating it is a saprophyte growing on the basidiome. This superficial layer was not seen in DAR 32036. It has not been seen in the other collections of A. griselloides.
Specimens examined
WESTERN AUSTRALIA. City of Albany, G. Byrne 5790 (PERTH 09175938); loc. id., G. Byrne 626628 (PERTH 09004831); loc. id., D.A. & D.G. Reid, R Hilton & N. Brittan s.n. (DAR 32036); shire of Esperance, E.M. Davison 2–1990 & P.J.N. Davison (PERTH 03096866); loc. id., E.M. Davison 5-1990 & P.J.N. Davison (PERTH 03096858); shire of Denmark, K. Syme 5/91 (PERTH 04988442); city of Albany, K. Syme 400/92 (PERTH 05254299).

|
|
Amanita peltigera D.A.Reid, Victorian Naturalist 95(2): 49 (1978)
Type: AUSTRALIA. South Australia: Stirling West, 35°00′S, 138°43′E, Mar. 1976, J. Randles s.n. (holo: K(M) 236385!).
Mycobank number: MB308577.
Mycobank number: MB308550.
Notes
As mentioned in the Introduction, Reid (1980) considered A. dumosorum and A. peltigera to be similar, apart from the presence of a partial veil and the absence of a saccate volva (Fig. 18). However, a study of collections of A. peltigera from SA and WA showed variation in the presence of the partial veil and the development and thickness of the universal veil at the top of the bulb (Davison et al. 2020). Reid (1980) also stated that the structure of the universal veil on the pileus was entirely hyphal; however, we have noted infrequent, large, inflated cells in this tissue (Fig. 19). Basidiospores are amyloid and of similar size: A. peltigera [499/25/25] (6.5–)8–10.5(–11.5) × (5.5–)6–8(–9) μm, (L = 8.0–10.0 μm; L′ = 9.1 μm; W = 6.3–7.7 μm; W′ = 7.1 μm; Q = (1.06–)1.14–1.43(–1.62); Q = 1.20–1.38; Q′ = 1.29), A. dumosorum: [40/1/1] 7–10(–10.5) × 5.5–7.5 μm, (L′ = 9.0 μm; W′ = 6.5 μm; Q = (1.17–)1.20–1.54; Q′ = 1.38), and their microscopic features are similar (Fig. 19). On this basis, A. dumosorum is synonymised with A. peltigera.

|
|
|
1 Basidiome secotioid |
|
2 Pileus aging grey, brown or greyish-brown |
|
3 Bulb conic or marginate |
|
4 Universal veil on pileus becoming pale brown to grey, spores elongate to cylindrical |
|
5 Bulb marginate |
|
6 Universal veil on pileus grey to brown, spores ellipsoid to elongate |
|
7 Pileus aging grey, universal veil on pileus felted, breaking into aerolate patches |
|
8 Clamp connections absent |
|
9 Basidiome capitate |
|
10 Spores ellipsoid to cylindric |
|
11 Spores elongate to cylindrical (Q = 1.85–2.13) |
Discussion
The recent revision of Amanitaceae by Cui et al. (2018), by using concatenated datasets of nuLSU, β-tubulin, ef1-α and rpb2 gene regions, provides a phylogeny for Amanita species with amyloid spores. Our work, using only the nuLSU gene region, shows that section Arenariae is sister to sections Phalloideae, Strobiliformes and Validae, with good bootstrap support (Fig. 1). Within section Arenariae, there is a well-supported clade containing the secotioid species A. arenaria, A. arenarioides, A. compacta and A. pseudoarenaria, and the agaricoid species A. griselloides, A. pupatju and A. sabulosa (Fig. 2B–D). There are insufficient sequences to resolve their relationships with A. lesueurii, A. peltigera, A. wadjukiorum and A. wadulawitu. the other known members of this section.
The multicopy ITS region is the DNA barcode marker for fungi (Schoch et al. 2012). We have found that cloning is necessary to consistently obtain high-quality sequences (McGurk 2013; McGurk et al. 2016; Davison and Giustiniano 2020; Davison et al. 2017a, 2017b, 2020). However, cloning has shown inconsistent utility of this region for delineating species from different sections of Amanita from southern Australia (Davison et al. 2017a, 2020). In section Phalloideae, ITS sequences varied by up to 1.42% among collections of the same species, and by 1.42–1.60% between different species which are morphologically distinct and geographically separated (Davison et al. 2017a); however, in section Arenariae, amplicons from the same individual vary from 0.0 to 9.9% (Tables 3, 4; Davison et al. 2020). There were no changes to the methodology and processing of samples from sections Phalloideae and Arenariae. Examination of the ITS region showed consistent insertions and deletions among different species, and some consistent transitions and transversions among collections of the same species. We have assumed that these represent real differences among clones, not being PCR-induced errors; no chimeras were detected by UCHIME. We were not able to determine whether these differences represent past hybridisation events (Hughes et al. 2015, 2018).
Section Arenariae
It is likely that there are many more described agaricoid species in Australia that are unrecognised members of this section. These would be expected to have a slightly appendiculate pileus margin, and inflated sub-basidial cell; possible examples are A. gracilenta, A. albidannulata A.E.Wood, and A. annulalbida A.E.Wood.
Data availability statement
The data that support this study are available in the article.
Conflicts of interest
The authors declare that they have no conflicts of interest.
Declaration of funding
This work was supported by the Australian Biological Resources Study Grants CN211-40, and CN216-04, by Lotterywest Grant Application 421011392 and the Western Australian Naturalists’ Club Serventy Memorial Fund. L. E. McGurk was supported by a Curtin University Postgraduate Scholarship.
Acknowledgements
We thank the Fungarium Collections Manager, Royal Botanic Gardens, Kew, for the loan of K(M) 236386 and K(M) 236387, and DAR for the loan of DAR 32034, DAR 32037 and DAR 32036. We thank T. W. May for reviewing the manuscript, K. Syme for permission to use Fig. 4, and G. Byrne for permission to use Fig. 16. Associate Professor Elder in Residence Simon Forrest is thanked for cultural and linguistic advice in naming A. pupatju.
References
Abarenkov K, Zirk A, Piirmann T, Pöhönen R, Ivanov F, Nilsson RH, Kõljalg U (2020) UNITE USEARCH/UTAX release for Fungi. Version 04.02.2020. (UNITE Community) Available at https://plutof.ut.ee/#/doi/10.15156/BIO/786375 [Verified 15 April 2021]Bas C (1969) Morphology and subdivision of Amanita and a monograph of its section Lepidella. Persoonia 5, 285–579.
Bas C (1975) A comparison of Torrendia (Gasteromycetes) with Amanita (Agaricales). Beihefte zur Nova Hedwigia 51, 53–61.
Bougher NL (1999) New species of Torrendia (fungi, Agaricales) from remnant woodlands in the wheatbelt region of Western Australia. Australian Systematic Botany 12, 145–156.
| New species of Torrendia (fungi, Agaricales) from remnant woodlands in the wheatbelt region of Western Australia.Crossref | GoogleScholarGoogle Scholar |
Bougher NL, Lebel T (2002) Australasian sequestrate (truffle-like) fungi. XII. Amarrendia gen. nov.: an astipitate, sequestrate relative of Torrendia and Amanita (Amanitaceae) from Australia. Australian Systematic Botany 15, 513–525.
| Australasian sequestrate (truffle-like) fungi. XII. Amarrendia gen. nov.: an astipitate, sequestrate relative of Torrendia and Amanita (Amanitaceae) from Australia.Crossref | GoogleScholarGoogle Scholar |
Bougher NL, Syme K (1998) ‘Fungi of Southern Australia.’ (University of Western Australia Press: Perth, WA, Australia)
Cai Q, Tulloss RE, Tang LP, Tolgor B, Zhang P, Chen ZH, Yang ZL (2014) Multi-locus phylogeny of lethal amanitas: implications for species diversity and historical biogeography. BMC Evolutionary Biology 14, 143
| Multi-locus phylogeny of lethal amanitas: implications for species diversity and historical biogeography.Crossref | GoogleScholarGoogle Scholar | 24950598PubMed |
Cleland JB (1931) Australian fungi: notes and descriptions. No. 8. Transactions and Proceedings of the Royal Society of South Australia 55, 152–160.
Cui Y-Y, Cai Q, Tang LP, Liu J-W, Yang ZL (2018) The family Amanitaceae: molecular phylogeny, higher-rank taxonomy and the species in China. Fungal Diversity 91, 5–230.
| The family Amanitaceae: molecular phylogeny, higher-rank taxonomy and the species in China.Crossref | GoogleScholarGoogle Scholar |
Davison EM (2011) Amanita ochroterrea and Amanita brunneiphylla (Basidiomycota), one species or two? Nuytsia 21, 177–184.
Davison EM, Giustiniano D (2020) Amanita hiltonii (Amanitaceae), a common but frequently misidentified mushroom in southwestern Australia, and a reconsideration of A. albifimbriata and A. brunneibulbosa. Muelleria 39, 59–73.
Davison EM, McGurk LE, Bougher NL, Syme K, Watkin ELJ (2013) Amanita lesueurii and A. wadjukiorum (Basidiomycota), two new species from Western Australia, and an expanded description of A. fibrillopes. Nuytsia 23, 589–606.
Davison EM, Giustiniano D, McGurk LE, Syme K, Robinson RM (2015) Amanita drummondii and A. quenda (Basidiomycota), two new species from Western Australia, and an expanded description of A. walpolei. Nuytsia 25, 1–13.
Davison EM, Giustiniano D, Busetti F, Gates GM, Syme K (2017a) Death cap mushrooms from southern Australia: additions to Amanita (Amanitaceae, Agaricales) sect. Phalloideae Clade IX. Australian Systematic Botany 30, 371–389.
| Death cap mushrooms from southern Australia: additions to Amanita (Amanitaceae, Agaricales) sect. Phalloideae Clade IX.Crossref | GoogleScholarGoogle Scholar |
Davison EM, Giustiniano D, McGurk LE, Watkin ELJ, Bougher NL (2017b) Neotypification and redescription of Amanita preissii (Basidiomycota), and a reconsideration of the status of A. griseibrunnea. Nuytsia 28, 193–204.
Davison EM, Giustiniano D, Haska JF (2020) Clarification of the type locality of Amanita peltigera (Agaricales, Amanitaceae), phylogenetic placement within subgenus Amanitina, and an expanded description. Swainsona 33, 51–61.
Department of the Environment (2013) Australia’s bioregions (IBRA), IBRA7. (Commonwealth of Australia) Available at https://www.environment.gov.au/system/files/pages/5b3d2d31-2355-4b60-820c-e370572b2520/files/subregions-new.pdf [Verified 3 February 2021]
Edgar RC, Haas BJ, Clemente JC, Quince C, Knight R (2011) UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 27, 2194–2200.
| UCHIME improves sensitivity and speed of chimera detection.Crossref | GoogleScholarGoogle Scholar | 21700674PubMed |
Grgurinovic CA (1997) ‘Larger Fungi of South Australia.’ (The Botanic Gardens of Adelaide and State Herbarium: Adelaide, SA, Australia)
Hughes KW, Petersen RH, Lodge DJ, Bergemann SE, Baumgartner K, Tulloss RE, Lickey E, Cifuentes J (2013) Evolutionary consequences of putative intra- and interspecific hybridization in agaric fungi. Mycologia 105, 1577–1594.
| Evolutionary consequences of putative intra- and interspecific hybridization in agaric fungi.Crossref | GoogleScholarGoogle Scholar | 23928423PubMed |
Hughes KW, Morris SD, Reboredo-Segovia A (2015) Cloning of ribosomal ITS PCR products creates frequent, non-random chimeric sequences – a test involving heterozygotes between Gymnopus dichrous taxa I and II. MycoKeys 10, 45–56.
| Cloning of ribosomal ITS PCR products creates frequent, non-random chimeric sequences – a test involving heterozygotes between Gymnopus dichrous taxa I and II.Crossref | GoogleScholarGoogle Scholar |
Hughes KW, Tulloss RH, Petersen RH (2018) Intragenomic nuclear RNA variation in a cryptic Amanita taxon. Mycologia 110, 93–103.
| Intragenomic nuclear RNA variation in a cryptic Amanita taxon.Crossref | GoogleScholarGoogle Scholar | 29864000PubMed |
Justo A, Morgenstern I, Hallen-Adams HE, Hibbert DS (2010) Convergent evolution of sequestrate forms in Amanita under Mediterranean climate conditions. Mycologia 102, 675–688.
| Convergent evolution of sequestrate forms in Amanita under Mediterranean climate conditions.Crossref | GoogleScholarGoogle Scholar | 20524599PubMed |
Kearse M, Moir R, Wilson A, Stones-Havas S, Cheung M, Sturrock S, Buxton S, Cooper A, Markowitz S, Duran C, Thierer T, Ashton B, Meintjes P, Drummond A (2012) Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 28, 1647–1649.
| Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data.Crossref | GoogleScholarGoogle Scholar | 22543367PubMed |
Kimura M (1980) A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. Journal of Molecular Evolution 16, 111–120.
| A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences.Crossref | GoogleScholarGoogle Scholar | 7463489PubMed |
Kornerup A, Wanscher JH (1983) ‘Methuen Handbook of Colour’, 3rd edn. (Methuen: London, UK)
McGurk LE (2013) Delineation of selected Western Australian Amanita mycoflora by macroscopic, microscopic and DNA sequence analysis. PhD thesis, Curtin University, Perth, WA, Australia.
McGurk LE, Giustiniano D, Davison EM, Watkin EJ (2016) Amanita wadulawitu (Basidiomycota), a new species from Western Australia, and an expanded description of A. kalamundae. Nuytsia 27, 21–30.
Miller OK (1991) New species of Amanita from Western Australia. Canadian Journal of Botany 69, 2692–2703.
| New species of Amanita from Western Australia.Crossref | GoogleScholarGoogle Scholar |
Miller OK (1992) Three new species of Amanita from Western Australia. Mycologia 84, 679–686.
| Three new species of Amanita from Western Australia.Crossref | GoogleScholarGoogle Scholar |
Miller OK, Horak E (1992) Observations on the genus Torrendia and a new species from Australia. Mycologia 84, 64–71.
| Observations on the genus Torrendia and a new species from Australia.Crossref | GoogleScholarGoogle Scholar |
Reid DA (1980) A monograph of the Australian species of Amanita Pers. ex Hook. (fungi). Australian Journal of Botany, Supplementary Series 10, 1–96.
Royal Botanic Garden, Edinburgh (1969) ‘Flora of British fungi: colours identification chart.’ (Her Majesty’s Stationery Office: Edinburgh, UK)
Schoch CL, Seifert KA, Huhndorf S, Robert V, Spouge JL, Levesque CA, Chen W, Fungal Barcoding Consortium (2012) Nuclear ribosomal internal transcribed spacer (ITS) region, a universal DNA barcode marker for fungi. Proceedings of the National Academy of Sciences of the United States of America 109, 6241–6246.
| Nuclear ribosomal internal transcribed spacer (ITS) region, a universal DNA barcode marker for fungi.Crossref | GoogleScholarGoogle Scholar | 22454494PubMed |
Tamura K (1992) Estimation of the number of nuceotide substitutions when there are strong transition-transversion and G + C content biases. Molecular Biology and Evolution 9, 678–687.
Tamura K, Nei M (1993) Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Molecular Biology and Evolution 10, 512–526.
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S (2011) Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Molecular Biology and Evolution 28, 2731–2739.
| Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods.Crossref | GoogleScholarGoogle Scholar | 21546353PubMed |
Tavaré L (1986) Some probabilistic and statistical problems on the analysis of DNA sequences. Lectures on Mathematics in the Life Sciences 17, 57–86.
Tulloss RE (2000) Notes on methodology for study of Amanita (Agaricales) Available at http://www.amanitaceae.org/content/uploaded/pdf/methodsb.pdf [Verified 11 August 2016]
Wood AE (1997) Studies in the genus Amanita (Agaricales) in Australia. Australian Systematic Botany 10, 723–854.
| Studies in the genus Amanita (Agaricales) in Australia.Crossref | GoogleScholarGoogle Scholar |
Yang ZL, Cai Q, Cui Y-Y (2018) Phylogeny, diversity and morphological evolution of Amanitaceae. Biosystematics and Ecology-Series 34, 359–380.