

A treatment for Polygala of northern Australia
Raelee A. KerriganSchool of Marine and Tropical Biology, James Cook University, Cairns, Qld 4878, Australia and Northern Territory Herbarium, Department of Natural Resources Environment, The Arts and Sport, PO Box 496, Palmerston, NT 0831, Australia. Email: raelee@ryansurveys.com.au
Australian Systematic Botany 25(2) 83-137 https://doi.org/10.1071/SB08032
Submitted: 23 June 2008 Accepted: 8 December 2011 Published: 30 April 2012
Abstract
A taxonomic treatment is presented for all Polygala L. species of northern Australia (north of 26°S). In total, 45 species are treated, of which 44 are native and 36 are endemic to Australia. Twenty-six new species and four new varieties are described. A key to all 50 Australian species is provided, including six introduced species (five of which are restricted to southern states of Australia and are not treated here in detail). Lectotypes are here designated for five names. Nomenclatural issues regarding P. linariifolia Willd., P. glaucoides L. and P. triflora L. are discussed.
Introduction
Polygala L. is a widespread genus with a global distribution, excluding Antarctica, the Arctic, New Zealand and the islands of Polynesia, although species have now been introduced to New Zealand and Polynesia. The main centres of diversity are known from South Africa and the Americas (North, Central and South) (Cronquist 1981; Paiva 1998; Bernardi 2000). Estimates of the number of species in the genus vary widely from 300–350 species (Eriksen and Persson 2007), ~500 species (Cronquist 1981) to 725 species (Paiva 1998). These discrepancies are an indication that regional work is lacking and that generic circumscription is changing. Previously, in the most recent treatment (Monro 2003), only 19 species were recognised for Australia. In the present treatment, which is focused on northern Australia, the number of species has more than doubled. In Australia, native Polygala species are typically tropical annual herbs, usually patchily distributed throughout savanna woodland, sandstone heath and seasonally inundated lowlands and occurring in a variety of substrates ranging from sandy soil to black clay. The main diversity is distributed across northern Australia; the few species recorded in the southern temperate states are mostly introduced shrubs.
In many instances, the flora of northern Australia remained relatively poorly known until well into the 20th century, and the region still contains large areas of land that are remote and difficult to access, and remain under-surveyed (Specht 1958, Clarkson and Kenneally 1988; Dunlop and Webb 1991; Wheeler 1992; Woinarski et al. 2006). As an illustration of this situation, an average of 46 ‘new’ vascular plant species per annum were added to the Northern Territory (NT) checklist between 1972 and 2004, in many cases informally recognising undescribed taxa (Cowie and Albrecht 2004). In addition, it is recognised that the Kimberley Region of Western Australia (WA), the far north of the NT and Cape York Peninsula, although sharing a suite of common species, separately have large bodies of endemic taxa, in particular in the sandstone flora (Clarkson and Kenneally 1988; Dunlop and Webb 1991; Woinarski et al. 2006). The often highly dissected and fragmented landscapes lead to high levels of endemism (Duretto and Ladiges 1997; Leach 2000; Woinarski et al. 2006). As the tropical flora has been studied by taxonomists, it has not been unusual for revisionary works to describe large numbers of new plant species. In a recent study on Spermacoce R.Br. in the NT, 39 of the 53 species described were new (Harwood and Dessein 2005). Similarly, Toelken (2010), in revising tropical Australian Hibbertia Andrews in just two subspecific groups, described 52 new taxa. Over the past 30 or so years, many of the major genera containing numerous undescribed tropical taxa have been revised. These include Boronia Sm. (Duretto 1997, 1999), Goodenia Sm. (Carolin 1990), Heliotropium L. (Craven 1996), Pityrodia R.Br. (Munir 1979), Mitrasacme Labill. (Dunlop 1996), Triumfetta L. (Halford 1997) and Triodia R.Br. (Burbidge 1953; Lazarides 1997). Polygala is among the last of these major genera to be treated.
Classification
The only monographs on Polygalaceae with a global focus were published by Chodat more than a century ago (Chodat 1893, 1896). The following 10 genera were identified: Bredemeyera Willd., Carpolobia G.Don., Monnina Ruiz & Pav., Moutabea Aubl., Mundia Kunth, Muraltia Neck., Polygala, Salomonia Lour., Securidaca L. and Xanthophyllum Roxb. These genera were classified into three tribes, namely Polygaleae, Moutabeae and Xanthophylleae, with a fourth tribe, Carpolobiae, now recognised. The Angiosperm Phylogeny Group lists 21 genera, on the basis of Eriksen and Persson (2007), in four tribes, for the family (Stevens 2001).
Polygala was first published by Linnaeus (1753) in Species Plantarum. He described 22 species, one of which, P. chinensis L., is currently applied to Australian material (Monro 2003). In his key to genera, Chodat (1893) distinguished Polygala by the combination of eight stamens united in a semi-circle, a dehiscent capsule and the absence of a very long tuft of hairs on the seed. Acknowledging the genus as the largest of the family (~500 species), Chodat (1893, 1896) divided Polygala into the following 10 sections (author citation follows Chodat 1896): Acanthocladus (Klotzsch) Chodat, Brachytropis (Willk.) Chodat, Chamaebuxus Chodat, Gymnospora Chodat, Hebecarpa Chodat, Hebeclada Chodat, Ligustrina Chodat, Phlebotaenia (Griseb.) Chodat, Orthopolygala Chodat, nom. illeg. and Semeiocardium (Hassk.) Chodat. Orthopolygala is an illegitimate name because it included the type of Polygala, namely P. vulgaris L. Orthopolygala, or Polygala as it is now known, is broken into 14 subsections (Chodat 1893, 1896). Nine subsections occur in Australia, with many of the northern Australian species initially assigned to section Polygala, subsection 11, series Chloropterae Chodat (Chodat 1893). The species belonging to subsection 11 were distinguished by ‘capsula alata; stylus rimiformis; stigma superius breve, cochleatum, subemarginatum; inferius longius, carnosum pendens’ (capsule winged; style cleft; upper stigma short, spirally twisted, subemarginate; lower stigma long, fleshy, pendulous). Species belonging to Chloropterae were distinguished by ‘petala integra; arillus conspicua carinatus’ (petals entire; arils conspicuously keeled). Chodat’s (1893) concept of Chloropterae is confusing, however, because illustrations of some species from that series were incompatible with his description, because both cleft styles with an upper and lower stigma, and hooked styles, which I would not consider cleft, are provided. This apparent contradiction was removed in Chodat (1896) in which Chloropterae (now designated as one of three series belonging to subsection 10, Deltoideae Chodat) was redefined under Deltoideae as having the following characteristics: a bent style, narrowly ribbon-like (flattened in cross section); stigma variable, anthers not sessile; upper petals broadly triangular, more colourful than the wings (alae); sepals green and acute; upper sepals not connate. Although the list of species in Chloropterae in the later publication is not comprehensive in that only 8 of the 19 species are listed, it appears to represent the same suite of species, because the missing species are not listed elsewhere.
Phylogeny
Regional treatments and recent molecular and morphological analyses of inter- and intra-family phylogeny, conducted by Eriksen (1993a, 1993b), Persson (2001), Monro (2003) and Forest et al. (2007), have questioned the classification of Polygalaceae and sectional treatment of Polygala presented by Chodat (1896). Although these authors were unable to provide a fully resolved phylogeny for all Polygalaceae, they were able to highlight issues requiring further research, such as polyphyly in several genera such as Bredemeyera and Polygala. Apparently, the polyphyletic nature of Polygala is not surprising because several infrageneric groups have previously been recognised as independent genera (Eriksen 1993b). Four new genera have been proposed for North American Polygala, on the basis of phylogenetic data indicating four morphologically diagnosable clades that are more closely related to other genera than they are to the type group of Polygala (Abbott 2011). Also, undescribed diversity is evident in the genus, and several authors have noted that infrageneric classification of Polygala cannot be attempted until the Neotropical and South-east Asian species have been revised and included in analyses (van der Meijden 1988; Pendry 2001; Monro 2003).
Current Australian knowledge
In a recent treatment of Australian Polygala, Monro (2003) assigned 19 Polygala names to Australian taxa and classified them into infrageneric groups according to Chodat (1896) (Table 1). Seven species were identified as naturalised. Nine of the 12 native species were placed in the series Chloropterae of subsection Deltoideae. The remaining three native species were placed in either series Leptaleae or series Fortificatae. All series belong to Polygala section Polygala (P. sensu stricto). All but four of the native taxa, assigned by Monro, were also described and listed by Chodat (1896), and placed in series Chloropterae, on the basis of short axillary inflorescences and flowers with greenish acute wing sepals. All Chloropterae species are found north of ~30°S in Australia and only one native species, Polygala japonica Houtt. (series Fortificatae), is found further south in any abundance. The taxa assigned to series Chloropterae formed a monophyletic group in phylogenetic analyses based on morphological and molecular data (Monro 2003). However, it is not known if such features can really be forwarded as putative synapomorphies, because taxon sampling was insufficient to test the clades robustly. Also, these analyses did not account for many of the potentially new taxa documented as phrase names in Australian checklists since then. There were four potentially new and 16 described taxa recorded in the Queensland (QLD) census (Henderson 2002) and two potentially new and 12 described taxa recorded for WA in FloraBase (Anon. 1998). The NT vascular plant checklist later recorded 11 potentially new and 17 described taxa (Kerrigan and Albrecht 2007).

|
The only species found in Australia that have been included in recent phylogenetic analyses are the introduced species P. myrtifolia L., P. paniculata, P. virgata Thunb. and P. vulgaris. Using Bayesian inference, P. myrtifolia, P. virgata and P. vulgaris fall within an Old World clade of Polygala sens. str., and P. paniculata falls within a New World clade of Polygala sens. str. (Forest et al. 2007). Support for an Old World and New World clade in Polygala s.str. is also found in Persson (2001), in which Polygala sens. str. falls in two different Old World or New World clades in a large polytomy in the strict consensus tree. Because of limited taxon sampling for Australia, very few hypotheses can be drawn about the remaining Australian species. The assumption is that, on the basis of morphology and biogeography, most of the Australian taxa are likely to be each other’s closest relatives.
The aim of the present study is to describe morphological diversity in Polygala across northern Australia, with a view to providing a key for all Australian members of the genus and to provide good population data (such as precise location data for populations and degree of variability within populations) on which to base future phylogenetic analyses. New taxa and species limits are proposed, on the basis of careful field and herbarium study of morphological characters, and new species are formally described.
Material and methods
Species concept
The present study provides species limits for several taxa, on the basis of morphological similarity. The use of morphological characters to some degree implies the use of the morphological species concept (MSC). However, in recognising species, shared morphology is assumed to reflect a shared genetic lineage, genome and phylogeny, even though no genetic or phylogenetic data were gathered. The hypothesis is that morphologically similar populations are more closely related than morphologically dissimilar populations, and that morphological differences indicate the existence of some sort of impediment to gene flow, indicating a likelihood of discrete genetic lineages.
A major criticism of the MSC is that it is unable to detect morphologically identical sibling species and may underestimate diversity. Sibling species that are difficult to distinguish morphologically may differ molecularly to the same degree as entities that are morphologically distinct (Mayr 1993; Mallet 1995a; McDade 1995). Similarly, it is possible for morphologically distinct entities to have no detectable genetic differences, especially because molecular studies typically involve only one to a few markers or gene regions. Also, because criteria for determining species boundaries are not clearly prescribed, the MSC is difficult to apply to species that are highly variable and have distinct infraspecific variants. Consequently, species limits may vary with taxonomists and taxa (Mayr 1993; Mallet 1995a, 1995b). Although this criticism is valid, the present study provides the essential first step of describing existing diversity by using morphological characters, and provides both an hypothesis and a framework on which populational studies and future phylogenetic analyses can be based.
Characters
Taxonomically informative characters typically occur consistently, invariably and in combination with other characters within and/or across populations and allow for the diagnosis of discrete taxa. Typically, taxonomically informative characters vary within the group under study; however,they are consistent within taxa, are not easily modified with environmental changes and ideally co-vary with other characters (Stuessy 1990). It could be argued under the MSC or phylogenetic species concept (PSC) that one character-state difference is sufficient to diagnose a group and therefore a species. There are alternatives to this view. Some authors have suggested that at least two independent characters are required to differentiate species, following the morphological species concept (Hedberg 1958; Davis and Heywood 1963), or five correlated character differences following the phenetic species concept (Sneath and Sokal 1973). These same authors, however, also suggested that the degree of difference required to delineate species depends to some extent on the group being studied and its own internal diversity.
Traditionally, morphological characters used to differentiate species in Polygala have included reproductive characters of floral, seed and fruit structure, and vegetative characters of indumentum, leaf shape and habit (Bentham 1863; Chodat 1893; Domin 1927; Adema 1966; van der Meijden 1988; Pendry 1999, 2001). It is possible, that small changes in floral structure are sufficient to isolate phylogenetic lineages through reproductive isolation (Brantjes 1982) and, therefore, can offer strong evidence for defining species. Similarly, in sympatric populations, the maintenance of different character states is taken as evidence of reproductive isolation; e.g. in P. triflora and P. glaucoides, differences in the size of the lateral petals relative to the keel and the presence or absence of straight hairs have provided sufficient evidence to differentiate these sympatric species (van der Meijden 1988). Following Stuessy’s (1990) guidelines for distinguishing infraspecific ranks, geography is the most important component in the recognition of infraspecific taxa. On the basis of the definition that varieties are, in the most part, morphologically and geographically distinct but have areas of gradation, two species with varietal variation are described in the present research. They have one to few conspicuous differences, are cohesive and are largely allopatric, with some overlap.
Herbarium specimens of Polygala collected from northern Australia and held by AD, BRI, CANB, DNA, MEL and PERTH were investigated for taxonomically informative characters. Additional field survey to collect material from QLD, WA and the NT was conducted. Collection of fresh material and photographs much improved access to floral characters such as appendage type and style structure. In total, 1479 collections of Polygala from north of 26°S were examined from Australian herbaria. In addition, 150 specimens from SING and L were examined to help establish endemicity of newly described Australian taxa.
Type materials in the form of specimens or photographs were borrowed from B, BM, BRI, CANB, CGE, E, G, K, LINN, L, MEL, P, PERTH and PR to facilitate the application of published names and to assess the application of previously published names to taxa in Australia.
Notes on morphology
In Australia, native Polygala species are mostly annual herbs, although two species, P. isingii Pedley and P. triflora, appear to be short-lived perennials. Habit varies from low-spreading plants to multi-stemmed subshrubs or erect single-stemmed annuals. Leaves are simple and alternate, and range from linear to orbicular. Descriptions of leaf shape in this treatment follow Radford (1986). The indumentum generally consists of fine white hairs, either straight or curving from the base (arcuate). Plant indumentum consists of either curved hairs only, or with both curved hairs and sparse straight hairs present. Often, both hair types are present, even if only on alae or capsule. The inflorescence is usually a few- to many-flowered axillary or supra-axillary raceme, occasionally leaf-opposed or lateral (Po. orbicularis Benth., P. macrobotrya Domin, P. wightiana Wight & Arn.) or terminal (P. longifolia Poir.). Inflorescences vary from tightly congested with overlapping flowers, e.g. P. eriocephala F.Muell ex Benth., to very loose with flowers widely spaced along rachis, e.g. P. rhynchocarpa R.A.Kerrigan, sp. nov.
Flowers
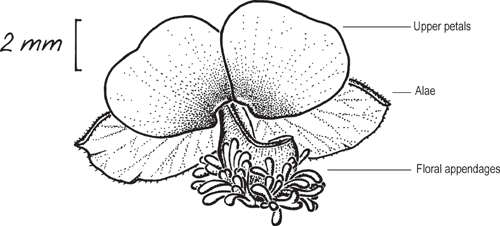
Polygala has bilaterally symmetrical flowers. The calyx consists of five sepals, including three small green unmodified outer sepals, and two enlarged, herbaceous or petaloid, inner lateral sepals (alae). In some texts, the alae are described as wings; however, in the present paper, wing is used to describe a wing sometimes present on the fruiting capsule. The corolla consists of three petals (Fig. 1). The lower petal is a clawed boat-shaped keel, with a pair of dorsal appendages attached to the keel, one on either side of the keel and often distally offset to one side. There are also two broad, clawed upper petals. In some texts, the appendages on the lower petal are referred to as crests. Flowers are bisexual. Eight stamens are fused at the base into a staminal sheath surrounding the style and variously fused to the upper petals. The style and stigmas are variable and often act as pollen presenters. The fruit consists of a laterally compressed bilocular capsule, with one seed per locule. The seeds are hairy, rarely glabrous and arillate. The aril is variously shaped, with two or three appendages running longitudinally along the seed.
|
The highly modified, keeled flowers of Polygala indicate an adaptation to specialist pollination. Bee, butterfly and bird pollination are all reported for Polygala species (Brantjes 1982; Lack and Kay 1987; van der Meijden 1988; Westerkamp and Weber 1997a). The style in Polygala is usually sheathed in the staminal column and enclosed within the keel petal. Pollen is often released and deposited on the stigma or accumulated in a modified structure on the style. Pollen transfer follows when the pollinator depresses the keel petal, usually by landing on it or the floral appendages, releasing the enclosed style from the keel and allowing the pollen to be transferred to the pollinator. In some species, tertiary pollen presentation is suspected (Westerkamp and Weber 1997b). Variations in pollen presentation have been shown to provide a mechanism for enforcing reproductive isolation in some species (Brantjes 1982; Kruger and Pretorius 1997). Some species of Polygala are reported to be autogamous (Kruger and Pretorius 1997).
According to Eriksen (1993a) and Chodat (1893), floral nectaries vary in Polygala. They can present as either annular discs or unilateral processes, or be absent. However, according to Dube and Awasthi (1985), in many cases, the glandular zone is inconspicuous, and these authors suggested that all members of Polygala should be considered as having a nectariferous zone. No floral nectaries were readily observed in the material seen in the present study. However, additional histological study is required to verify whether nectariferous zones are present.
Floral appendages
Each flower of Australian Polygala has a pair of appendages found on the distal dorsal surface of the keel petal. Floral appendage structure has been used only to a limited extent by Bentham (1863), Chodat (1893) and Monro (2003) to delimit Australian species, most likely because the appendages are quite cryptic in dried material. However, with fresh material or material preserved in spirit collections, differences can be readily seen.
Seven broad types of floral appendage are defined here and are illustrated in Fig. 2.

|
Bifurcate appendages are divided at least once to the base. They often divide further with age, but are generally no more than three- or four-lobed. When fresh, the appendages of P. petrophila and P. succulenta are very thick and succulent. The succulence of P. rhynchocarpa appendages is yet to be confirmed in fresh flowers (Fig. 2A). See P. petrophila, P. rhynchocarpa and P. succulenta.
Coralliform appendages are irregularly divided, sometimes to base, with uneven lobes and are distinguished from bifurcate appendages by their rounded apex. One lobe is usually much broader than the others, reminiscent of the branches of staghorn corals (Fig. 2B). See P. clavistyla and P. coralliformis.
Crenate appendages are irregularly notched along the margin, often with one protruding lobe like the thumb on a pair of mittens (Fig. 2C). See P. parviloba.
Entire appendages are undivided appendages (Fig. 2D). See P. integra and P. praecox.
Fimbriate appendages are split into many fine divisions, and each division is of relatively even thickness throughout. Fimbriate appendages are either straight or recurved (Fig. 2E, F). See P. bifoliata, P. canaliculata, P. crassitesta, P. dependens, P. difficilis, P. exsquarrosa Adema, P. gabrielae Domin, P. galeocephala, P. glaucifolia, P. glaucoides, P. isingii, P. kimberleyensis, P. japonica, P. longifolia, P. macrobotrya, P. obversa, P. orbicularis, P. paniculata, P. pendulina, P. persicariifolia DC., P. polifolia C.Presl, P. pterocarpa, P. rhinanthoides Sol. ex Benth., P. saccopetala, P. scorpioides, P. stenosepala, P. tepperi F.Muell., P. triflora and P. wightiana.
Horned appendages are narrowly oblong, with an acute apex and are turgid (Fig. 2G). See P. eriocephala F.Muell. ex Benth.
Spathulate appendages are similar to fimbriate appendages in that they consist of many even-sized divisions, but the divisions broaden distally (Fig. 2H). See P. barbata, P. barklyensis, P. dimorphotricha, P. geniculata, P. pycnantha, P. stenoclada Benth. and P. validiflora.
Style and stigmas
Observations of Australian Polygala indicate that they are varied and diverse in style and stigma structure; however, they generally possess either a strongly hooked style with a flat stigma on the inner surface of the hook, giving a horseshoe shape to the apex of the style, or are curved with a globular stigma. In many cases, when the style is not strongly hooked, it extends past the stigma. Chodat (1893) often referred to this extension as a sterile second stigma, and it often has a role in pollen presentation. In many cases, the style extension is grooved or sculptured and forms a pocket where pollen is captured. These observations are in keeping with those of other researchers, e.g. Westerkamp and Weber (1997b). There are some species in the genus that have two fertile stigmas (Castro et al. 2008). This is considered the ancestral form, and in some species one of the ancestral stigmas has been modified to form a pollen presenter or has been lost altogether, as is the case for many Australian species (Ladd and Donaldson 1993; Ladd 1994; Castro et al. 2008). Variations in the length, hairiness and shape of the extension and in the type of sculpturing exist amongst Australian species (Fig. 3). These differences could possibly be involved in the reproductive isolation of the species. In the present study, the degree to which the style is distinctly dorso-ventrally flattened is a useful diagnostic character for P. succulenta, P. exsquarrosa and P. petrophila, when compared with P. parviloba, P. clavistyla and P. integra. In other species, the character becomes more obscure; e.g. in P. geniculata and P. glaucifolia the distinctiveness of the flattening varies as the style thickens in places, and the orientation of the flattening is obscured by the twisting of the style. This is confounded further by the fact that the style rapidly dries out under the heat of a microscope light source during examination, exaggerating the degree of flattening. I have also noticed variation in the turgidity of the style, and therefore the interpretation of this character, with the apparent age of the flower when preserved in spirit. Caution is advised; the character is robust for the few species mentioned above but can be difficult to interpret for the remaining species and would not be useful in phylogenetic analyses. Only rehydrated flowers were available for the following species: P. bifoliata*, P. barkleyensis*, P. dependens*, P. glaucoides, P. isingii*, P. japonica, P. kimberleyensis*, P. persicariifolia, P. polifolia, P. pycnantha*, P. triflora and P. validiflora*. In some cases (marked with an asterisk), it was difficult to determine whether the flowers examined were self-pollinated and had not fully developed.

|
Staminal column and upper petals
In most of the Australian material, stamens are fused part way along their length to form a sheath. They are usually monadelphous, and fused evenly or fused in increasing lengths from the outer to the middle pair, giving the fused area a deltoid appearance. The stamens of P. wightiana are triadelphous, fused in three bundles of three, two and three. Useful staminal column and upper petal characters consist of the extent and area and/or position of fusion of the staminal sheath to the upper petals, and the presence or absence of a visible flap created by this fusion at anthesis (Fig. 4). In the present study, the width of the visible portion of the staminal sheath (here labelled staminal flap) is measured from where the upper petal claw attaches to the staminal sheath to where the keel petal obscures the staminal sheath. This character is useful in diagnosing P. dimorphotricha (0.9–1.5 mm) from P. barbata (≤0.5 mm). In some species, e.g. P. longifolia, the upper petal attaches further up the staminal sheath than does the overlapping keel petal and is difficult to measure. As with the style characters mentioned above, some species were coded from rehydrated flowers only. In the present treatment, the anthers are described as dehiscing via apical slits, following Eriksen (1993a), who declared that the anthers were commonly misinterpreted as dehiscing via apical pores. Stamens in Australian species appear to dehisce via apical pores; however, I did not systematically study stamens throughout their development and dehiscence to determine whether they are slits when young. Hence, I cannot refute or corroborate Eriksen’s (1993a) work.

|
Arils
Arils and caruncles are mechanisms for attracting and rewarding dispersal vectors (Berg 1975), or may assist in seed hydration (Forest et al. 2007). Many types of dispersal have been noted for Polygala (van der Meijden 1988), including myrmecochory (ant dispersal), ornithochory (bird dispersal), anemochory (wind dispersal), diplochory (dispersal by two separate vectors, e.g. wind-dispersed seed falls from fruit and is then animal-dispersed) and epizoochory (borne externally by animals). Although myrmecochory has been established in several Australian Polygala (Verkerke 1985), studies outside Australia have shown that aril appendages and seed indumentum act as air sacs and aid in floating, as well as providing enticements and handles for ants to carry the seeds (Oostermeijer 1989; Anon. 2002). Although advantages of myrmecochory to Australian Polygala are speculative, Berg (1975) suggested that buried seeds are protected from rapid fires and predation, and are often planted in positions suitable for germination. Although no germination trials have been conducted on Australian Polygala, fire and seed predation are prominent features of the northern Australian landscape, and work on Florida species has indicated that germination is higher if seeds are buried or treated with smoke water (Anon. 2002).
Researchers studying the ovule ontogeny in Polygala refer to the fleshy aril structure and appendages of Polygala seed as an exostome aril (Verkerke and Bouman 1980). According to Mauseth (1988), an exostome aril is an aril formed from the outer integument of the micropyle. The present treatment uses the terms, aril and aril appendages, for the outgrowth at the micropyle and hilum end of the seed, following Verkerke and Bouman (1980) and van der Meijden’s (1988) treatments. Following the convention established by previous treatments (Adema 1966; van der Meijden 1988), the aril-bearing end is treated as the apex of the seed because of the seed’s attachment to the stylar end of the capsule.
The presence, number, length and texture of aril appendages were utilised by Chodat (1893) in his subsectional classification of Polygala and keys to species. Other taxonomically useful seed characters include indumentation and aril head shape and colour. The aril head refers to the fleshy white outgrowth at the micropylar end of the seed. In Australian material, the aril heads are situated at the apex of the seed. They vary in shape, size and colour and are categorised into five types (see below, illustrated in Fig. 5. These categories represent a gradation from an intact fleshy outgrowth, i.e. the ‘domed’ category, to a much reduced aril, represented only by the attachment of the aril appendages, i.e. the ‘reduced’ category. In some species, aril head shape is variable between two categories along this continuum. In any case, the categories are useful in describing morphological diversity and represent discrete character states between ends of the continuum. Descriptions of aril-head shapes and colours do not include the appendages, which extend from the head longitudinally down the seed, and apply to mature seeds only.

|
Domed heads are symmetrical and are either longer than wide or as long as wide (Fig. 5A). See P. praecox and P. succulenta.
Round heads are symmetrical and are generally shorter than wide (Fig. 5B). See P. barbata, P. bifoliata, P. canaliculata, P. crassitesta, P. dimorphotricha, P. gabrielae, P. glaucoides, P. integra, P. obversa, P. orbicularis, P. polifolia, P. praecox, P. pterocarpa, P. rhinanthoides, P. stenosepala and P. validiflora.
Hooked or skewed heads are slightly asymmetrical and are generally shorter than wide (Fig. 5C). See P. bifoliata, P. canaliculata, P. dimorphotricha, P. eriocephala, P. gabrielae, P. japonica, P. kimberleyensis, P. macrobotrya, P. obversa, P. pendulina, P. pterocarpa, P. saccopetala, P. scorpioides, P. stenosepala and P. validiflora.
Helmet heads are more or less asymmetrical, with the head as long as it is wide or longer than wide, and often with a central, lateral groove where the aril is reduced to the seed testa (Fig. 5D–F). See P. coralliformis, P. difficilis, P. eriocephala, P. galeocephala, P. isingii, P. kimberleyensis, P. pendulina, P. pycnantha, P. rhynchocarpa, P. saccopetala, P. scorpioides, P. stenoclada, P. tepperi, P. triflora and P. wightiana.
In the case of P. longifolia and P. persicariifolia, the aril appendages can be so reduced and aril so symmetrical as to give the aril head a horseshoe shape.
Reduced heads have no aril development at the top of the seed and are reduced to appendages only (Fig. 5G, H). See P. barklyensis, P. clavistyla, P. dependens, P. exsquarrosa, P. geniculata, P. glaucifolia, P. paniculata, P. parviloba and P. petrophila.
Variations in seed indumentation and shape, which have proved useful for diagnosis, are illustrated for clarity in Fig. 6.

|
Phylogeny
Allocation of the newly described Australian taxa to Chodat’s subsectional and series classification (Chodat 1893, 1896) is difficult. Chodat defined only 9 of the 14 subsections he recognised, but his descriptions were not dichotomous, and the character states defining each group were not always exclusive. P. wightiana, for example, was listed by Chodat (1893) as belonging to Leptaleae, on the basis of style and anther characters (Table 1). However, these characters are not exclusive to Leptaleae and could also apply to series Chloropterae subsection Deltoideae (Table 2). Given that some aspects of Chodat’s classification are not supported by recent phylogenetic analyses (Eriksen 1993a, 1993b; Persson 2001; Monro 2003; Forest et al. 2007) and can be too ambiguous to apply, an infrageneric classification is not provided. However, it is possible to speculate on putative natural groups within the Australian material.

|
An intuitive assessment of Australian Polygala identifies two morphological groups (see below). Group 1 comprises taxa with green alae, monadelphous stamens, flat stigmas, secondary sterile stigma absent and distinctly hooked styles with horseshoe-shaped apex, whereas Group 2 comprises taxa with green alae, monadelphous stamens, curved to shortly hooked styles with a more or less globular stigma, the style of which extends past the fertile stigma and is considered a sterile stigma which often functions as a pollen presenter. The remaining species are more difficult to group, although they can mostly be separated on the basis of petaloid alae and monadelphous stamens versus green alae and triadelphous stamens; however, the resulting groups are heterogeneous with respect to styles and stigmas. A few species do not fall into any of these groups, as discussed below. A preliminary sorting of native Australian species into these groups is listed below. States and territories where these species are found, as well as their ranges outside Australia, are listed in parentheses, and distribution in Australia is classified as broad or narrow. The categories, broad and narrow, are subjective classifications based on the number of known populations and the extent of the area of occupancy as mapped in the present study (Figs 7–10). As a guide, narrow distributions generally consist of five or fewer known populations and are usually restricted to one state or bioregion. Broad distributions generally consist of 10 or more known populations and are usually across all northern states (QLD, NT and WA) and several bioregions.

|

|

|

|
Group 1 comprises the following species: P. barbata (NT, broad), P. barklyensis (NT, QLD, narrow), P. bifoliata (WA, NT, broad), P. coralliformis (NT, broad), P. crassitesta (WA, NT, QLD, broad), P. dependens (NT, QLD, broad), P. difficilis (NT, QLD, broad), P. dimorphotricha (WA, NT, narrow), P. gabrielae (QLD, narrow), P. galeocephala (WA, NT, broad), P. geniculata (NT, narrow), P. glaucifolia (WA, NT, SA, QLD, broad), P. glaucoides (QLD, narrow), P. isingii (WA, NT, broad), P. kimberleyensis (WA, narrow), P. macrobotrya (QLD, narrow), P. obversa (NT, QLD, broad), P. orbicularis (NT, narrow), P. pendulina (QLD, narrow), P. praecox (WA, narrow), P. pterocarpa (WA, NT, QLD, broad), P. pycnantha (QLD, narrow), P. rhinanthoides (QLD, narrow; PNG), P. saccopetala (WA, narrow), P. scorpioides (QLD, narrow), P. stenoclada (NT, QLD, broad), P. stenosepala (WA, NT, broad), P. tepperi (WA, NT, broad), P. triflora (SA, QLD, NSW, Sri Lanka to PNG) and P. validiflora (NT, narrow).
Group 2 comprises the following species: P. canaliculata (WA, narrow), P. clavistyla (NT, narrow), P. eriocephala (WA, NT, broad), P. exsquarrosa (WA, NT, QLD, broad), P. parviloba (WA, NT, QLD, broad), P. petrophila (WA, NT, broad) and P. succulenta (WA, NT, broad).
Species with petaloid alae, monadelphous stamens, styles and stigmas various include the following: P. japonica (QLD, VIC, NSW, ACT, broad, India, Asia, Philippines, Malesia and Papua New Guinea), P. longifolia (WA, NT, QLD, broad, SE Asia and Malesia), P. paniculata (NSW, QLD, broad, tropical America, Africa, SE Asia and Malesia) and P. persicariifolia (QLD, narrow, Africa, South-east Asia and Malesia).
Species with green alae, triadelphous stamens, styles and stigmas various include the following: P. polifolia (QLD, broad, Pakistan to China and from Philippines to Australia) and P. wightiana (WA, NT, QLD, broad, India, South-east Asia and Malesia).
Few biogeographical conclusions can be drawn from these groupings, and consequently any hypotheses regarding breeding strategies and character evolution would be highly speculative. Group 1 is a diverse group comprising largely Australian endemics with both very restricted (e.g. P. geniculata and P. praecox) and broad (e.g. P. pterocarpa and P. tepperi) distributions. Species from Group 1 share very similar if not identical style and/or stigma arrangements characterised by the absence of a second stigma. The extent of self-pollination in this group needs to be explored. In most of the flowers observed in the present study, pollen had accumulated in the nook of the style. It is likely that this is pollen from the same flower, as the stamens are situated directly below the stigma. If this is the case, the pollen is likely to congest the stigmatic surface, excluding cross-pollination, unless a mechanism exists that releases pollen after a pollinator has visited, as has been observed in P. myrtifolia (Westerkamp and Weber 1997b). In this case, the pollen mass released into the nook springs out from the keel when the flower is first visited by a bee. The pollen mass hits the bee dorsally, and pollen is then moved to a tertiary position between the first and second visit. Indirect evidence for both self-pollination and herkogamy (a mechanism generally involved in the prevention of self-interference) exist for Polygala. However, Castro et al. (2008) found that stigma interference did appear to influence pollen-tube growth in some herkogamous species, and so few generalisations can be made. Group 2, entirely Australian endemics, comprises species with very broad (P. exsquarrosa) and narrow (P. canaliculata) distributions; members have intricate and specific style and/or stigma arrangements characterised by the presence of the second sterile stigma, suggesting unique pollen presentation and, therefore, potentially unique pollinators. P. rhynchocarpa (WA, narrow) and P. integra (NT, broad) are not easily placed in either of the two major groups, possessing neither the hooked style of Group 1 nor the globular stigma and style extension and sterile stigma of Group 2. Which of the states represents the ancestral form is unknown, although it has been suggested that the pollen presenter form is an intermediate state between the ancestral form of two fertile stigmas and the more derived state in which the second stigma is entirely absent (Ladd and Donaldson 1993; Ladd 1994). When one considers the bilocular condition of the genus, it does seem more parsimonious to view the loss of the second stigma as a more derived feature; however, without a full phylogenetic context, this is speculative. Most of the remaining Australian species that lack the diagnostic features of Groups 1 and 2 are widely distributed outside Australia. Consequently, any attempt to understand their morphological groupings must include the appropriate global context.
Conservation
Following International Union for the Conservation of Nature (IUCN) Red List categories (IUCN 2001), most of the species covered in this revision are in the category of Least Concern. Where found, they are sufficiently abundant and/or widespread, with no identified threats, and do not qualify for any of the IUCN Red List threatened categories. The following species are considered Data Deficient and require further survey to establish abundance, distribution and potential threats: P. barklyensis, P. canaliculata, P. gabrielae, P. glaucoides, P. macrobotrya, P. persicariifolia, P. polifolia, P. praecox and P. rhynchocarpa. In these cases, there has been insufficient targeted surveying of the species to determine whether the current known distribution is an artefact of sampling effort or a reflection of real distribution and abundance.
Genus description
Extra Australian limits and conditions are provided in square brackets for a few distinctive features.
PolygalaL. Sp. Pl. 2: 701. (1753)
Type: P. vulgaris L., selected by N.L.Britton & A.Brown, Ill. Fl. of N. USA ed. 2, 2: 446 (1913).
Annual or perennial herbs, shrubs [small trees or rarely lianas]. Young stems and inflorescence rachis often angular or flattened. Leaves usually alternate, rarely opposite or whorled, rarely caducous. Flowers in a many-flowered inflorescence, rarely solitary. Inflorescence a terminal, leaf-opposed or axillary raceme or spike [or sometimes a terminal panicle]; bracts and bracteoles present. Flowers zygomorphic, white, yellow, blue, purple or mauve. Sepals 5, outer 3 unmodified, with the inner 2 lateral sepals (alae) enlarged and often petaloid. Petals 3, connate at the base and variously adnate to the stamens, lower petal a boat-shaped keel, clawed, with a pair of variously shaped appendages on the distal dorsal surface [or unadorned]; lateral upper petals clawed, often auriculate on free outer margins, variously hairy or glabrous. Stamens 8, fused at the base into a staminal sheath, monadelphous or triadelphous [or partly diadelphous outside Australia]; anthers 2-celled and dehiscent via a short apical pore-like slit. Style variously hooked, curved or more or less straight, sometimes apparently extending past fertile stigma; although considered to have two stigmatic lobes, in Australian material, the second stigma is often modified (either reduced or lost) and sterile; fertile stigma globular or flat, lateral or subapical. Ovary laterally compressed, 2-celled with 1 ovule per cell. Fruit a compressed loculidical capsule, usually obovate or orbiculate, with a broad or narrow marginal wing, dehiscing along the outer suture. Seeds hairy, with an aril, the aril with 2 or 3 appendages of various shapes and lengths running longitudinally along the seed.
The implicit norm (to be assumed in the species description) for the following characters are as follows: general plant colour green, leaves not conduplicate, leaves persistent, leaves more or less concolorous, inflorescence not scorpioid, alae green, style glabrous, lateral pocket on keel absent, keel petal glabrous abaxially, seed indumentum of one hair type only, capsule lobe apices equal (i.e. level and opposite) and no tap-root thickening. Only the exceptions are addressed in the species description. Capsule and alae positions are described relative to the rachis (Figs 11, 12). Capsule lobe apices are described as separated when neither touching nor overlapping.

|

|
A genus with several species, estimated between 300–350 (Eriksen and Persson 2007) and 725 species (Paiva 1998), with a sub-cosmopolitan distribution but with its main diversity focused in tropical areas; 50 species in Australia. Herbs in the NT have a methyl salicylate (e.g. wintergreen) smell when roots are scratched on fresh specimens, as commonly reported for most members of the entire subfamily Polygaleae.
Notes on key and species descriptions
Corolla colour can be unreliable in this group because the corolla can fade to white with age in living material or can appear yellow when dried. Most floral characters require fresh flowers. Flowers in spirit or rehydrated dried material can easily be misinterpreted; e.g. in dried specimens, pollen deposition can obscure style and/or stigma details. Thus, the key provided here works most reliably with freshly collected material. As per the taxonomic norm, types that have been sighted are noted with an exclamation mark. Where available, institution barcodes are reported. In some cases, available photographs pre-dated barcodes. Sometimes attempts to provide type-specimen information were made difficult by contradictions or discrepancies between information on the labels of type specimens and information from published type citations. In such cases, the details of both are provided. There has been continuing confusion regarding citation of Domin’s ‘Beitrag zur Flora und Pflanzengeographie Australiens’ (Contribution to the Flora and Plant Geography of Australia). The correct citation for Polygala is Bibliotheca Botanica Heft 89 (Sep. 1927) 301–305 (Henderson 1984).
A key to all 50 Australian species is provided, including six introduced species. Descriptions for all 44 native Australian taxa and one introduced species in northern Australia are provided. Descriptions for the following four of the remaining five introduced southern species are available from the state floras of Victoria (VIC), New South Wales (NSW) and South Australia (SA): P. monspeliaca L. (SA, VIC), P. myrtifolia (NSW, SA, VIC), P. virgata (NSW, SA, VIC) and P. vulgaris (VIC). The sections of the key pertaining to these species, which I did not look at in detail, are largely taken from Monro (2003) and the respective state floras. P. duarteana A. St.-Hil. has not been recollected from a small area around Beaudesert in far south-eastern QLD since 1973 to confirm its persistence. A description is available in Monro (2003).
For most species, the full list of specimens examined is given. Where the list is extensive, a subset of collections has been provided. These subsets include high-quality specimens with good reproductive material, from a range of Australian herbaria, and were chosen to represent the morphological and geographic ranges of the species. For this reason, many of the older collections are not represented in the subsets. Full specimen citations are listed in Kerrigan (2008). The actual number of specimens examined, provided in parentheses for each species, represents unique collections examined (including types). The tally does not include the number of duplicates seen for each collection. Although collection numbers may be small for some species, in most cases where I have made collections, many individuals were observed from the population represented by the collection. Exceptions include those species classified above as Data Deficient. For all species except P. japonica and P. paniculata, distribution maps were generated from specimens seen. Maps of examined specimens were generated using geographic information system, ArcGIS Desktop 9.2 (ESRI, Redlands, CA). Accuracy of map points varies with accuracy of data collection. As a general guide, post-1996 collections are assumed to be collected using global positioning system (GPS) technology and be accurate to decimal degrees, whereas pre-1996 collections are assumed to be collected using map grid lines, with accuracy dependent on the scale of the maps used and the availability of good landmarks for an accurate location of position. P. japonica and P. paniculata distribution maps were extracted from the Australian Virtual Herbarium website.
Key to species
|
1. Alae herbaceous to leaf-like, more or less green, rarely tinged dark maroon and/or purple |
|
2. Slender spreading, ascending or erect herbs or subshrubs, usually <60 cm high, if taller than annual herbs |
|
3. Leaves narrow-obovate to almost linear (sometimes almost absent); inflorescences pyramidal and several-flowered, exceeding the upper leaves |
|
4. Flowers axillary or lateral; racemose or solitary |
|
5. Main colour of alae white or cream (rarely pink) |
|
6. Aril appendage 3-lobed and short, confined to apex of seed virtually absent |
|
7. Alae cream; fruit wing developed on only one margin; stems without minute glandular hairs |
|
8. Plant puberulous; style short and straight; fruit broadly winged |
|
9. Plant lax and rather diffuse; curved hairs throughout; aril head skewed [Fig. 5C] |
|
10. Staminal filaments 8 (monadelphous) |
|
11. Stem glabrous; seed 3–4 mm long, oblong [northern Australia] |
|
12. Capsules not prominently winged, if present, wing up to 0.5 mm wide 18 |
|
13. Alae ovate, elliptic, very widely ovate to orbicular, not falcate; capsule more or less symmetrical |
|
14. Floral appendages fimbriate or spathulate [Fig. 2E, F, H]; capsule wing usually more or less same width throughout |
|
15. Seed testa uniform, without thickened collar-like area |
|
16. Alae ±equal to, only slightly wider or narrower than capsule (including wing) |
|
17. Alae ±same width as capsule body (excluding wing); capsule wings usually well exserted past alae; [northern Australia] |
|
18. Style distinctly hooked at apex to form horseshoe shape [Fig. 3C]; pollen accumulation in bend of hook |
|
19. Floral appendages not horned; style not cleft |
|
20. Style not truncated into broad flattened apex, straight or curved but not hooked |
|
21. Apex of capsule lobes acute |
|
22. Style more or less terete, not distinctly dorso-ventrally flattened when fresh, if distinctly dorso-ventrally flattened then with groove above stigma; seed usually ovoid |
|
23. Each floral appendage entire [Fig. 2D], coralliform [Fig. 2B] or crenate [Fig. 2C] |
|
24. Floral appendages variously divided, not fimbriate (<4 prongs per appendage), leaves 6–55 mm long |
|
25. Floral appendages not coralliform |
|
26. Alae broadly ovate to ovate 2–5 mm wide; capsules deflexed [Fig. 11B]; floral appendages entire (rarely notched); stigma flat situated laterally at tip of style; style glabrous |
|
27. Seed with one hair type, fine whitish or ferruginous hairs throughout or glabrous (hairs can be of different lengths) [Fig. 6B–G] |
|
28. Floral appendage fimbriate; leaves elliptic, ovate, lanceolate or linear |
|
29. Leaves linear, lanceolate to narrowly elliptic; plant erect, branches spreading and ascending in the form of a candelabra |
|
30. Aril head not reduced and either helmet-shaped, hooked or rounded [Fig. 5A–F]; if somewhat reduced [Fig. 5F], then plant erect, foliage green and leaf apex obtuse |
|
31. Indumentum of curved hairs only; leaf apex of most leaves acuminate, acute, obtuse or rounded, leaves not glaucous; plant erect, often with lateral branches |
|
32. Capsule perpendicular to deflexed; alae 3.5–8.0 mm long; inflorescence up to 40 mm long |
|
33. Leaves mostly linear to oblanceolate, rarely some leaves obovate2. P. barklyensis |
|
34. Stem and leaf indumentum usually of curved and straight hairs (P. pendulina and P. dimorphotricha very rarely with curved hairs only) |
|
35. Leaves mostly linear to obovate or elliptic |
|
36. Lateral pocket on keel absent or poorly developed |
|
37. Most leaves other than obovate and if obovate, seed ovoid–oblong or oblong |
|
38. Plant erect to ascending, sometimes with spreading lateral branches; flowers purple; in a wide variety of habitats, but not known from sandy coastal ridges |
|
39. Capsule perpendicular to reflexed apex rounded; leaves various, linear to obovate; plants up to 60 cm tall, never 65 cm tall |
|
40. Leaves linear to obovate, 1–14 mm wide; seed <4 mm long, if >4 mm, leaves not linear (check that no hairs are extending past base to ~1 mm) |
|
41. Infloresence often exserted past leaves but not beyond plant [Fig. 13B], up to 125 mm long; alae in mature fruit up to 8 mm long |
|
42. Aril head hooked to rounded, not strongly helmet-shaped |
|
43. Alae 4.9–8.0 mm long |
|
44. Alae more or less equal to or narrower than capsule |
|
45. Most leaves linear, lanceolate, elliptic, oblong or ovate, if oblanceolate or obovate then ≤10 mm wide |
|
46. Aril head rounded or hooked, not helmet-shaped; floral appendages spathulate [Fig. 2H] |
|
47. Leaves mostly obovate to very widely obovate to orbicular; seed ovoid, staminal flap ~0.5 mm wide or less, upper petal claw ~2 mm wide at staminal sheath [Fig. 4B] |
|
48. Inflorescence sparsely hairy (straight hairs) or glabrous, if approaching densely hairy then inflorescence well exserted from leaf axils extending up to 90 mm long |
|
49. Keel petal glabrous abaxially |
|
50. Most leaves linear to oblanceolate, occasionally some leaves narrowly elliptic or obovate, up to 46 mm long; seed with short hairs restricted to just below aril, rest of seed with longer hairs |
|
51. Seed 3.0–4.2 mm long; alae 3.3–6.4 mm long (mature fruit); leaves 2–9 mm wide |
|
52. Inflorescence up to 90 mm long; tap root usually thickened; no pocket on keel petal |
Taxonomy
1. Polygala barbata R.A.Kerrigan, sp. nov.
Polygalae obversae affinis, a qua pilis rectis indumentorum, indumento petalorum lateralium et lobis latioribus cristae florala differt. Ab P. coralliformis lobis angustatioribus cristae floralis differt; ab P. kimberleyensis arillo galeiformi differt; ab P. orbicularis Benth. foliis obverse-ovatis et pilis rectis indumentorum differt; ab P. dimorphotricha tubo stamineo marginibus extimis petalorum lateralium non conjugenti et foliis latioribus differt.
Type: Litchfield National Park, Wangi Falls, rainforest track, 16.i.2004, R.A. Kerrigan 723 & D.J. Dixon; holo: DNA 161431!; iso: BRI, CANB, K, PERTH.
Annual herb, erect often with spreading or decumbent branches or prostrate, up to 40 cm high; indumentum of curved and straight hairs. Leaves elliptic to orbicular, oblanceolate to depressed obovate, 2–42 mm long, 2.5–30.0 mm wide; apex rounded or retuse, mucro 0.2–0.4 mm long; petiole 0–3 mm long. Inflorescence axillary or supra-axillary, a solitary flower or usually a short raceme, up to 15 mm long, not exceeding the leaves. Pedicel up to 2 mm long. Alae position antrorse, perpendicular and deflexed in fruit; herbaceous, ovate, usually somewhat dimidiate, 3.6–6.0 mm long, 2.0–3.6 mm wide; with curved and straight hairs. Corolla purple; floral appendages spathulate; upper petal claw 2 mm wide. Stamens terminate at stigma, monadelphous; staminal flap ≤0.5 mm wide. Style hooked (horseshoe-shaped at apex), more or less terete, with flat stigma along inner surface of hook. Capsule antrorse, perpendicular and deflexed, the capsule more or less aligned with alae. Capsule lacking wing, symmetrical, very widely ovate to widely oblong or squarish, 3–6 mm long, 3.5–5.5 mm wide; capsule lobe apices rounded and separated. Seed ovoid, 3.0–4.2 mm long, 1.5–2.1 mm wide; indumentum of fine white or ferruginous hairs, sometimes glabrous or with glabrous patches. Aril head round, 0.5–0.7 mm long, white, with fine short hairs. Aril appendages 3, linear, 0.9–2 mm long (Figs 2H, 4B).
Distribution
Australia (NT). This species is endemic to the NT and is distributed widely across the far north of the NT, from Borroloola to Port Keats (Fig. 7A).
Habitat
Found in a variety of habitats and substrates, including river margins, bulwaddy (Macropteranthes kekwickii F.Muell. ex Benth.), lancewood (Acacia shirleyi Maiden) and dry vine thickets, but mostly in woodlands on sandy loam.
Notes
This species is variable. Individuals may be prostrate or erect, have small orbicular or large obovate leaves and have hairy or glabrous seeds. Similar to P. coralliformis and P. obversa, it can be distinguished from the former by the evenly divided floral appendages, and from the latter by the presence of long straight hairs on the stems and leaves, a pilose tuft of hairs on the upper petal auricle, floral appendages that broaden at the apex and a staminal tube adnate across the whole upper petal.
Etymology
Latin: barbatus (bearded), referring to the presence of a tuft of hairs on the upper petal auricle.
Selected specimens (60 examined)
2. Polygala barklyensis R.A.Kerrigan, sp. nov.
Polygalae exsquarrosae affinis, a qua stigmate plano et stylo uncato differt.
Type: Mallapunyah Station, 80 km S of Cape Crawford, 1 km E of Carpentaria Hwy, 29.iv.1991, I.D. Cowie 1605 & B.A.Wilson; holo: DNA 59745!; iso: BRI, MEL.
Annual herb, erect with branches spreading to ascending, up to 16 cm high; indumentum of curved hairs. Leaves linear, narrowly elliptic, oblanceolate or rarely some leaves obovate, 5–38 mm long, 1–5 mm wide; apex acute or acuminate, occasionally obtuse, mucro ~0.2 mm long; petiole up to 0.5 mm long. Inflorescence supra-axillary or axillary, a raceme, up to 40 mm long, sometimes exceeding the leaves, although usually shorter than leaves. Pedicel up to 2 mm long. Alae perpendicular to deflexed in fruit; herbaceous, ovate, strongly dimidiate, 3.6–5.2 mm long, 1.6–2.2 mm wide; with scattered curved hairs only, especially along the margin. Corolla purple; floral appendages spathulate; upper petal claw ~0.7 mm wide. Stamens terminate at stigma, monadelphous; staminal flap ~0.7 mm wide. Style hooked (horseshoe-shaped at apex), in dried state style dorso-ventrally flattened and twisted in apical half, with flat stigma along inner surface of hook. Capsule deflexed, apex of alae diverging at ~45° from capsule. Capsule lacking wing, more or less symmetrical, ovoid, 2.25–2.75 mm long, 3.0–3.2 mm wide; capsule lobe apices rounded and separated. Seed ovoid, 1.5–2.7 mm long, 1.25–1.60 wide; indumentum of fine white or ferruginous hairs. Aril head reduced, 0.25–0.50 mm long, brown where reduced to seed testa and white where appendages originate, with short hairs. Aril appendages 3, spathulate, 1.3–2.1 mm long.
Distribution
Australia (NT, QLD). This species is known in the NT from Mallapunyah Station north of the Barkly Tableland and in QLD from around Musselbrook Mining Camp and Lawn Hill National Park (Fig. 7B).
Habitat
Found in Corymbia dichromophloia (F.Muell.) K.D.Hill & L.A.S.Johnson or Eucalyptus leucophloia Brooker woodlands, usually on stony hill slopes, rarely in seepage areas with Melaleuca viridiflora Sol. ex Gaertn.
Notes
This species is very similar to P. exsquarrosa but can be distinguished from it by the hooked style and flat stigma.
Etymology
Latinised place name, barklyensis, referring to the general region where this species is found.
Selected specimens (6 examined)
3. Polygala bifoliata R.A.Kerrigan, sp. nov.
Polygalae stenosepalae affinis, a qua caulibus prostratis et foliis ellipticioribus differt. Ab congeneribus in Australia semine pilis cavis, crista florali fimbriata, et semine parvo distinguenda.
Type: Black Rock Pool, adjacent car park, 17.iv.2006, R.A. Kerrigan 1137; holo: DNA 177040!; iso: BRI, CANB, PERTH.
[Polygala arvensis [NT material] auct. non Willd. (1802), Benth. Fl. Austral. 1: 140 (1863) p.p. as to Upper Victoria R. F. Muell. K356709 (photograph DNA!); A.J. Ewart & O.B. Davies, Fl. N. Terr., 160 (1917); I.D. Cowie & D.A. Albrecht (eds), Checklist N. Terr. Vasc. Pl. Sp. (2004).]
[Polygala pycnophylla [NT material] auct. non Domin (1927), I.D. Cowie & D.A. Albrecht (eds), Checklist N. Terr. Vasc. Pl. Sp. (2004).]
Annual herb, prostrate or erect with decumbent branches, up to 15 cm high; indumentum of curved and straight hairs or occasionally with curved hairs only. Stems and leaves occasionally tinged maroon. Leaves of two general leaf shapes, linear, lanceolate or narrowly elliptic leaves, often on erect branches distally located, and elliptic or ovate leaves, usually proximal on decumbent branches, 2–46 mm long, 1.5–8.0 mm wide; apex acute or obtuse, mucro 0.2–0.5 mm long; petiole 0–1 mm long. Inflorescence supra-axillary or axillary, rarely a solitary flower or usually a raceme, up to 140 mm long, exceeding the leaves. Pedicel up to 9 mm long. Alae perpendicular, becoming reflexed in fruit; herbaceous, lanceolate to oblanceolate or elliptic, dimidiate to falcate, 2.8–6.3 mm long, 1.0–2.9 mm wide; with scattered curved hairs or glabrous. Corolla purple; floral appendages fimbriate; upper petal claw 1.5 mm wide. Stamens terminate at stigma, monadelphous; staminal flap ~0.5 mm wide. Style hooked (horseshoe-shaped at apex), in dried state style laterally compressed, with flat stigma along inner surface of hook. Capsule reflexed, apex of alae diverging at ~45° from capsule. Capsule lacking wing, usually strongly asymmetrical, rhomboid or squarish, 2.5–4.5 mm long, 2.6–3.9 mm wide; capsule lobes rounded, and more or less unequal. Seed ovoid–oblong, 2.0–3.6 mm long, 1.0–1.8 mm wide; indumentum of fine white hairs over whole seed and thicker recurved clearly hollow hairs adjacent to aril. Aril head round to hooked, 0.3–0.5 mm long, white, with fine short hairs or rarely glabrous. Aril appendages 3, linear, 0.75–2.10 mm long (Fig. 6A).
Distribution
Australia (NT, WA). This species is distributed across the far north of the NT N of 17°S and throughout the Kimberley region of northern WA (Fig. 7C).
Habitat
Usually found in eucalypt woodlands on a variety of soils, sometimes associated with rocky situations; rarely found in seasonally inundated habitats.
Notes
This species is very similar to P. stenosepala, and although extremes of both species are easily distinguished by leaf shape and habit, several specimens are difficult to assign clearly and may represent intergrades between the two species. These include C.R. Dunlop 6322, K.G. Brennan 4868 and 4986, and J.L. Egan 3117. P. bifoliata is generally prostrate or erect with decumbent branches, whereas P. stenosepala is usually erect with ascending branches. P. bifoliata is found in open eucalypt woodlands, in contrast to P. stenosepala which is commonly associated with seasonally inundated Melaleuca L. shrublands. Only a few flowering specimens are available for this species, and further differences may become apparent with additional material. These two taxa may represent phenotypic extremes affected by the different conditions of the habitat they occupy, with anomalous specimens linking the two extremes. This theory, however, is rejected as the few individuals that do occupy the other taxon’s habitat do appear to maintain morphological differences.
The total geographic overlap in the distribution of P. bifoliata and P. stenosepala makes it difficult to apply the infraspecific concepts proposed by Stuessy (1990) of 75% geographic and morphological separation. This concept proposes that the morphological distinction between most of the taxa is a consequence of geographic separation influencing the exchange of genes and that proximity would allow full and unrestricted gene exchange. It is likely that the intergrades represent hybrids between the two taxa, and it could be argued that the presence of only a few intergrades indicates some mechanism of reproductive isolation restricting interbreeding between the two populations.
Etymology
Latin bi- (two) and foliatus (leaf-bearing), referring to the two leaf shapes found on most specimens.
Selected specimens (59 examined)
4. Polygala canaliculata R.A.Kerrigan, sp. nov.
Polygalae rhynchocarpae affinis, a qua stylo canaliculato differt et lobis capsulae apicibus obtusis differt. Ab congeneribus in Australia apice styli canaliculato truncato, et lobis capsulae apicibus obtusis distinguenda.
Type: Kimberley, 142.6 km E from Kalumburu turnoff on Gibb River Rd, 16.v.2006, R.A. Kerrigan 1131; holo: DNA 176690!; iso: BRI, CANB, K, L, MEL, NT, PERTH.
Annual herb, erect with ascending branches, up to 45 cm high; indumentum of curved hairs. Leaves sparse and caducous, linear, 12–47 mm long, 0.5–1.5 mm wide; apex acute, mucro ~0.3 mm long; sessile. Inflorescence supra-axillary, usually a raceme, up to 170 mm long. Pedicel up to 3.6 mm long, exceeding the leaves. Alae position deflexed, becoming reflexed in fruit; herbaceous, lanceolate, narrowly elliptic or oblanceolate, dimidiate to falcate, 5–7 mm long, 1.6–2.4 mm wide; with scattered curved hairs. Corolla purple; floral appendages fimbriate; upper petal claw 1.2–1.5 mm wide. Stamens terminate at stigma, monadelphous; staminal flap <0.5 mm wide. Style curved, more or less terete, flattened at apex above globular stigma, flattened edge shallowly notched or grooved. Capsule deflexed, becoming reflexed, more or less aligned with alae. Capsule lacking wing, more or less asymmetrical, obovate, 5.0–5.5 mm long, 2.6–3.0 mm wide; capsule lobe apices rounded, obtuse, separated and more or less unequal. Seed oblong, 3.5–3.9 mm long, 1.2–1.3 mm wide; indumentum of fine white hairs. Aril head round sometimes hooked to one side, ~0.6 mm long, white, with short hairs. Aril appendages 3, linear, 0.6–1.0 mm long (Fig. 3F).
Distribution
Australia (WA). This species is endemic to WA and is only known from the Cockburn and Pentecost Ranges in the Kimberley Region (Fig. 7D).
Habitat
Usually growing amongst rocky rubble of sandstone in eucalypt woodlands with Triodia spp.
Notes
This species is very similar to P. macrobotrya, P. pendulina, P. rhynchocarpa and P. stenosepala. It can be distinguished from P. macrobotrya, P. pendulina and P. stenosepala by the curved, rather than hooked style and the narrow linear leaves. From P. rhynchocarpa, it can be distinguished by the groove on the end of the broad truncated style and the obtuse apex of the capsule lobes. P. rhynchocarpa has a more twisted and narrowly truncated style, with an extension that gives it a hammer-head appearance. This species is considered Data Deficient following IUCN Red List categories (IUCN 2001). Although only known from a few collections, this is likely a consequence of inadequate surveying rather than a reflection of the species’ real distribution and abundance.
Etymology
Latin canaliculatus (grooved), referring to the shallow groove at the top of the style.
Selected specimens (3 examined)
5. Polygala clavistyla R.A.Kerrigan, sp. nov.
Polygalae geniculatae affinis, a qua stigmate globoso et stylo clavato differt. Ab congeneribus in Australia stylo clavato distinguenda.
Type: Edith Falls, 13.iv.1995, J.L. Egan 4733; holo: DNA 124519!; iso: BRI, CANB, K, PERTH.
Annual herb, erect with spreading or decumbent branches, up to 10 cm high; indumentum of curved hairs. Stems and leaves often tinged red or maroon. Leaves oblanceolate to obovate, 4–26 mm long, 1.5–8.0 mm wide; apex rounded, truncate or retuse, mucro 0.2–0.3 mm long; petiole 0–1.5 mm long. Inflorescence terminal, a raceme, up to 65 mm long, exceeding the leaves. Pedicels upto 3.5 mm long. Alae position antrorse to perpendicular in fruit; herbaceous, ovate, dimidiate, 4.0–5.5 mm long, 1.3–3.0 mm wide; with curved hairs or glabrescent. Corolla purple; floral appendages irregularly divided, coralliform; upper petal claw 1.2–1.5 mm wide. Stamens terminate at stigma, monadelphous; staminal flap ~0.5 mm wide. Style curved, extending beyond globular stigma, more or less terete, becoming clavate at apex. Capsule antrorse or perpendicular, apex of alae diverging at ~45° from capsule. Capsule lacking wing, more or less symmetrical, ovoid, apex acute, often with curved overlapping horns or mucro on each lobe, 3.0–3.5 mm long, 2.65–3.00 mm wide; capsule lobe apices acute, frequently overlapping. Seed ovoid, 2.0–2.9 mm long, 1.2–1.5 mm wide; indumentum of fine white to ferruginous hairs. Aril head reduced, 0.3–0.5 mm long, brown where reduced to seed testa and white where appendages originate, hairy. Aril appendages 3, cuneate, 0.7–2.0 mm long (Figs 2B, 3J).
Distribution
Australia (NT). This species is endemic to the NT and is only known from a restricted area around Nitmiluk National Park, extending north into southern Kakadu National Park and east to Big River Station near the Roper River (Fig. 7E).
Habitat
Found in woodlands in skeletal soil on plateaux and rocky hills, amongst laterite or sandstone.
Notes
This species is very similar to P. geniculata but is distinguished by the globular stigma and club-shaped style.
Etymology
Latin clavatus- (club-shaped) and -stylus (style), referring to the club-shaped apex of the style.
Selected specimens (21 examined)
6. Polygala coralliformis R.A.Kerrigan, sp. nov.
Polygalae barbatae et P. obversae affinis, a quibis crista florali coralliformi differt. Ab congeneribus in Australia crista florali coralliformi, et semine pilis cavis distinguenda.
Type: N of Wildman River Ranger Station, 13.iii.2001, R.K. Harwood 1032; holo: DNA 145411!; iso: BRI, CANB, PERTH.
Annual herb, erect often with decumbent and/or spreading branches, up to 20 cm high; indumentum of curved and straight hairs, leaves glabrescent. Occasionally, leaf margins and alae tinged red. Leaves usually elliptic or obovate to depressed obovate, rarely very widely ovate or circular, 5–29 mm long, (2–)4–22(–32) mm wide; apex rounded, retuse or emarginate, rarely acute, mucro 0.3–0.4 mm long; petiole 0–2 mm long. Inflorescence supra-axillary or axillary, a solitary flower or sometimes 2–5 flowers arising separately along internode or very rarely on a short raceme, to <1 mm long, not exceeding the leaves. Pedicel up to 2 mm long. Alae position antrorse to perpendicular in fruit; herbaceous, obovate, oblanceolate or ovate, dimidiate, 5–11 mm long, 2–6 mm wide; with curved and straight hairs, or straight hairs only. Corolla dark purple, with white throat; floral appendages irregularly divided, coralliformis; upper petal claw 1.6 mm wide. Stamens terminate at stigma, monadelphous; staminal flap not visible up to 0.7 mm wide. Style hooked (horseshoe-shaped at apex), more or less terete, with flat stigma along inner surface of hook. Capsule antrorse to perpendicular, more or less aligned with alae. Capsule lacking wing, more or less symmetrical, widely ovate to widely oblong, 4.5–7.5 mm long, 3.5–5.5 mm wide, capsule lobe apices rounded and separated. Seed ovoid-oblong, 3.7–5.8 mm long, 1.4–4.5 mm wide; indumentum of fine white or ferruginous hairs over whole seed and thick clearly hollow hairs adjacent to aril head. Aril head distinctly helmet-shaped, 0.6–1.0 mm long, white to pale brown, rarely with fine short hairs. Aril appendages 3, linear, 0.35–1.25 mm long.
Distribution
Australia (NT). This species is endemic to the NT and occurs east of Darwin across to Bickerton Is. in north-eastern Arnhem Land and south to Borroloola (Fig. 7F).
Habitat
Found in a variety of habitats including sandstone heath, along the margins of swamps, monsoon vine forest and Allosyncarpia S.T.Blake forest, but mostly in woodlands on sandy loam or sand.
Notes
The species is similar to P. obversa and P. barbata but is readily distinguished by the coral-like floral appendages and the thick, hollow hairs below the aril of the seed. Specimens found around the Mount Brockman outlier in Kakadu National Park tend to be generally smaller in habit and leaf size.
Etymology
Latin coralliformis (coral-like in form), referring to the irregularly-lobed keel appendages, with some lobes broader than others, reminiscent of staghorn corals.
Selected specimens (87 examined)
7. Polygala crassitesta R.A.Kerrigan, sp. nov.
Polygalae pterocarpae affinis, a qua testa incrassata ad apicem seminis differt. Ab congeneribus in Australia testa incrassata ad apicem seminis distinguenda.
Type: Victoria Hwy, Bullo River Station turnoff, 09.iii.2006, R.A. Kerrigan 1032; holo: DNA 176646!; iso: BRI, CANB, K, NSW, PERTH.
[Polygala gabrielae [NT material] auct. non Domin (1927), I.D. Cowie & D.A. Albrecht (eds), Checklist N. Terr. Vasc. Pl. Sp. (2004).]
Annual herb, erect with ascending branches or more or less decumbent with spreading branches, up to 56 cm high; indumentum of curved and straight hairs, occasionally with only curved hairs on stems. Leaves linear, narrowly elliptic to elliptic, oblanceolate to obovate, 3–85 mm long, 1–16 mm wide; apex rounded, truncate or retuse, rarely acute, mucro 0.4–0.5 mm long; petiole 0–2.5 mm long. Inflorescence axillary or supra-axillary, rarely a solitary flower usually a raceme, up to 80 mm long, not exceeding the leaves. Pedicel up to 2.5 mm long. Alae position perpendicular, becoming deflexed in fruit; herbaceous, ovate and elliptic to very widely elliptic, 3.9–7.0 mm long, 2.9–5.7 mm wide; with straight and curved hairs. Corolla purple; floral appendages fimbriate; upper petal claw 1.2 mm wide; keel petal with slight lateral pocket development or pocket absent. Stamens terminate at stigma, monadelphous; staminal flap 1.0–1.2 mm wide. Style hooked (horseshoe-shaped at apex), often with beak on outer edge of bend visible in dried material, more or less terete at base but becoming dorso-ventrally flattened below hook and twisting in apical half, with flat stigma along inner surface of hook. Capsule deflexed, more or less aligned with alae. Capsule with wing, wing 0.55–2.00 mm wide, more or less equal throughout, symmetrical, widely elliptic, ovate, oblong or occasionally orbicular, 5.3–6.5 mm long, 3.7–5.7 mm wide; capsule lobe apices rounded and separated. Seed shape irregularly oblong (hour-glass shaped), 4.2–5.7 mm long, 1.7–2.4 mm wide; testa thickened adjacent to aril; indumentum of fine white hairs. Aril head round, 0.4–0.8 mm long, white and brown or black, short hairs present. Aril appendages 3, linear, 2.4–4.6 mm long (Fig. 6F).
Distribution
Australia (NT, QLD, WA). The main distribution of this species lies within NT and QLD and just extends into WA east of Kununurra. In the NT, it occurs in latitudes south of Katherine from the Victoria River District (VRD) to the Gulf of Carpentaria and south to the Barkly Tableland. In QLD, it occurs as far east as Mount Coolon, south-west of Mackay and south to Blackall (Fig. 7G).
Habitat
Almost always found on clay or cracking clay soils in grasslands with Dichanthium Willemet, Astrebla F.Muell. and Iseilema Andersson, often with Eucalyptus microtheca F.Muell., E. pruinosa Schauer, Corymbia terminalis (F.Muell.) K.D.Hill & L.A.S.Johnson or Bauhinia cunninghamiana (Benth.) Benth.
Notes
Specimens from QLD tend to be shorter and have smaller alae than those of other districts, and specimens from around the Gulf of Carpentaria and Barkly Tablelands tend to be taller with more linear leaves.
Etymology
Latin crassus- (thick) and -testa (outer coat of seed), referring to the thickened seed coat forming a collar at the base of the aril.
Selected specimens (50 examined)
8. Polygala dependens R.A.Kerrigan, sp. nov.
Polygalae exsquarrosae similis, sed ab ea aliis parvioribus, stylo uncato, inflorescentiis brevioribus distinguitur. Ab congeneribus in Australia capsulis pendulinis, et arillo seminis redacto distinguenda.
Type: Buchanan Hwy, 2 km from Stuart Hwy turnoff, S side of road, 12.ii.2004, R.A.Kerrigan 738 & P.S. Short; holo: DNA 161700!; iso: BRI, PERTH.
Annual herb, rounded to compact, erect often with spreading branches, up to 12 cm high; indumentum of curved hairs. Whole plant often tinged maroon. Leaves narrowly oblong to oblanceolate or obovate, 6–30 mm long, 2–10 mm wide; apex mostly acute, obtuse or rounded, rarely some leaves emarginate, mucro 0.2–0.5 mm long; petiole 0–2 mm long. Inflorescence axillary or supra-axillary, a raceme, rarely with a solitary flower on internode preceding raceme, up to 5(–20) mm long, not exceeding the leaves. Pedicel up to 0.5(–1) mm long. Alae position perpendicular in fruit; herbaceous, lanceolate, oblanceolate, falcate to narrowly dimidiate, 2.0–3.9 mm long, 0.8–1.8 mm wide; with curved hairs or glabrous. Corolla recorded as purple (fresh material not seen); floral appendages shortly fimbriate; upper petal claw ~0.5 mm wide. Stamens terminate at stigma, stamen monadelphous; staminal flap ~0.5 mm wide. Style hooked (horseshoe-shaped at apex), flattened dorso-ventrally at base, becoming thickened and terete in middle and dorso-ventrally flattened below hook in dried state, with flat stigma along inner surface of hook. Capsule deflexed or reflexed, apex of alae diverging at ~45° from capsule. Capsule lacking wing, symmetrical, oblong, 2.5–4.0 mm long, 2.6–3.7 mm wide, capsule lobe apices rounded and separated. Seed ovoid, 2.5–3.3 mm long, 1.2–1.8 mm wide; indumentum of fine white or ferruginous hairs. Aril head reduced, 0.4–0.6 mm long, brown where reduced to seed testa and white where appendages originate, with short hairs. Aril appendages 3, linear rarely spathulate or cuneate, 1.0–2.3 mm long (Fig. 5G).
Distribution
Australia (NT, QLD). In the NT, this species occurs south of Katherine to the Dulcie Ranges, west to Wave Hill in the VRD and east to Lake Nash on the Barkly Tablelands. In QLD, it extends across the border into Lawn Hill, north of Mt Isa (Fig. 7H).
Habitat
Found in a variety of habitats, including open woodlands and Acacia Mill. shrublands, often with Triodia R.Br. spp., on sand, clay loam, laterite or silt, or in skeletal soils on stony hill slopes.
Notes
Fresh flowers of this species have not been observed, and floral appendage type, corolla colour and arrangement of the staminal column are difficult to determine from dried material. I was able to find only a few specimens with old flowers; appendage type, in particular, was difficult to determine because appendages were very short and appeared poorly developed on these specimens.
Etymology
Latin dependens (hanging down), referring to the way the capsules are suspended in this species.
Selected specimens (25 examined)
9. Polygala difficilis R.A.Kerrigan, sp. nov.
Polygalae pycnanthae affinis, a qua pilis carinae differt. Ab congeneribus in Australia pilis carinae distinguenda.
Type: 33.6 km S of Tennant Creek, 01.v.2007, R.A. Kerrigan 1253 & D.J. Dixon; holo: DNA180895!; iso: AD, BRI, CANB, K, NSW, NT, PERTH.
Annual herb, erect with ascending branches, up to 30 cm high; indumentum of short curved and long straight or bent hairs. Leaves linear to narrowly oblong, rarely oblanceolate or obovate, occasionally conduplicate, 3.5–29 mm long, 1–6 mm wide; apex rounded, truncate, retuse or emarginate, mucro 0.2–0.3 mm long; petiole 0–2 mm long. Inflorescence supra-axillary or axillary, a raceme, up to 17 mm long, not exceeding the leaves. Pedicel up to 2 mm long. Alae position antrorse to perpendicular in fruit; herbaceous, narrowly ovate, usually falcate to dimidiate, 4.6–7.0 mm long, 1.4–2.4 mm wide; densely hairy with long straight hairs present, occasionally with curved hairs also. Corolla purple; keel petal with few strigose hairs along abaxial surface of keel; floral appendages fimbriate; upper petal claw 0.5–0.7 mm wide. Stamens terminate at stigma, monadelphous; staminal flap ~0.5 mm wide. Style hooked (horseshoe-shaped at apex), more or less terete, with flat stigma along inner surface of hook. Capsule antrorse or perpendicular, mostly more or less aligned with alae, or apex of alae diverging ~45° from capsule. Capsule lacking wing, oblong to ovate, 3.5–4.5 mm long, 2.3–3.3 mm wide; capsule lobe apices rounded and separated. Seed ovoid–oblong, 3.0–3.9 mm long, 1.25–1.50 mm wide; indumentum of fine white or ferruginous hairs. Aril head narrowly helmet-shaped, 0.6–1.0 mm long, white or white and brown, with fine short hairs over whole of aril head. Aril appendages 3, oblong, 0.50–0.75 mm long (Figs 5F, 12).
Distribution
Australia (NT, QLD). In the NT, this species is known from latitudes between Renner Springs and Ti-Tree and extends east along the Barkly Hwy to Camooweal in QLD. In QLD, it is recorded from Lawn Hill National Park in the north and is mostly collected along the Kennedy Hwy from Camooweal to White Mountains National Park, excluding the Mitchell Grass Plains (Fig. 7I).
Habitat
Usually found in a variety of eucalypt woodlands including Eucalyptus crebra F.Muell., E. pruinosa, E. leichhardtii F.M.Bailey and Acacia shrublands in sand and sandy loam.
Notes
This species is very similar to P. pycnantha and P. isingii but differs from the former by the presence of hairs on the keel and from the latter by the clustered inflorescence. Specimens of this species from QLD tend to have longer, conduplicate leaves with a more acute apex.
Etymology
Latin difficilis (difficult), referring to the difficulty in penning an available and suitable Latin name for this species.
Selected specimens (18 examined)
10. Polygala dimorphotricha R.A.Kerrigan, sp. nov.
Polygalae barbatae similis, a qua differt tubo stamineo marginibus extimis petalorum lateralium conjugenti et foliis magis obverse-ovatis. A P. kimberleyensis foliis angustatioribus et longioribus, et sepalis lateralibus longioribus et latioribus differt. A P. obversa crista florali lobis spathulatis, et indumento pilis rectis differt.
Type: Bullo River Station, 09.iii.2006, R.A. Kerrigan 1028; holo: DNA 176650!; iso: BRI, CANB, K, MEL, NSW, NT, PERTH.
Annual herb, erect with ascending or decumbent or spreading branches, up to 50 cm high; indumentum of curved and straight hairs, very rarely straight hairs only sparsely present. Leaves usually obovate or oblanceolate, 5–46 mm long, 5–18 mm wide; apex rounded, truncate or emarginate, mucro 0.3–0.5 mm long; petiole 0–2 mm long. Inflorescence axillary or supra-axillary, a raceme, up to 20 mm long, not exceeding the leaves. Pedicel to ~3 mm long. Alae position antrorse, occasionally perpendicular in fruit; herbaceous, ovate to lanceolate, usually somewhat dimidiate, 5–9 mm long, 2.0–3.5 mm wide; with curved and straight hairs or curved hairs only or glabrescent. Corolla purple; floral appendages spathulate; upper petal claw 1 mm wide. Stamens terminate at stigma, monadelphous; staminal flap 0.9–1.5 mm wide. Style hooked (horseshoe-shaped at apex), more or less terete, with flat stigma along inner surface of hook. Capsule antrorse to occasionally perpendicular, more or less aligned with alae. Capsule lacking wing, more or less symmetrical, very widely ovate to widely elliptic, 4.7–5.8 mm long, 3.7–4.6 mm wide; capsule lobe apices rounded and separated. Seed ovoid–oblong, 3.0–4.5 mm long, 1.3–2.0 mm wide; indumentum of fine white or ferruginous hairs present. Aril head round to hooked, 0.5–0.8 mm long, white or white and brown, with fine short hairs. Aril appendages 3, linear to triangular, 0.45–1.80 mm long (Fig. 4A).
Distribution
Australia (NT, WA). This species occurs west of the Victoria River in the NT to Beverley Springs in the Kimberley, WA (Fig. 7J).
Habitat
Mostly found in eucalypt woodlands amongst broken sandstone rubble on slopes or plains.
Notes
This species is very similar to P. obversa, P. barbata and P. kimberleyensis. This species is distinguished from P. kimberleyensis by having mostly obovate leaves, with the leaves generally being narrower and longer, and by having longer and wider alae. It is also distinguished from P. obversa by the broader appendages and the presence of straight hairs. P. dimorphotricha differs from P. barbata by the size of the flap created by the fusion of the staminal tube to the upper petal (Fig. 4A) and by having mostly obovate and generally narrower leaves. Like for P. barbata, the upper petal claw is adnate to staminal sheath thoughout. Two atypical specimens from Jasper Gorge Gregory National Park, K.G. Brennan 4381 (DNA) and R. Booth & N. Woodward 1556 (DNA), either have very few straight hairs or straight hairs are absent.
Etymology
Latin dimorphus- (having two forms) and -trichoma (hair), referring to the two hair types on this species, curved and straight.
Selected specimens (16 examined)
11. Polygala eriocephala F.Muell. ex Benth., Fl. Austral. 1 : 139 (1863)
Type: N Australia. Upper Victoria River, F. Muell.; holo: K 279777 (photograph DNA!).
Annual herb, erect with ascending branches, up to 40 cm high; indumentum of curved and straight hairs. Leaves linear or oblanceolate, 7–74 mm long, 1–8 mm wide; apex acute, obtuse or rounded, mucro ~0.4 mm long; petiole 0–1 mm. Inflorescence supra-axillary, a raceme, up to 35 mm long, not exceeding the leaves. Pedicel more or less lacking. Alae position antrorse to perpendicular in fruit; herbaceous, ovate, dimidiate, 6.4–9.8 mm long, 3.3–4.5 mm wide; densely villous with straight hairs. Corolla purple; floral appendages horned, offset to one side of keel; upper petal claw 1.5–2.0 mm wide. Stamens terminate at stigma, monadelphous; staminal flap not visible up to ~0.05 mm. Style sinuous, terete, cleft into a short stalk perpendicular to main axis with a globular stigma, and longer stalk persistent past stigma and sterile. Capsule antrorse to perpendicular, more or less aligned with alae. Capsule lacking wing, more or less symmetrical, widely ovate, oblong to widely obovate, 4.5–6.1 mm long, 3.5–4.5 mm wide; capsule lobe apices rounded and separated. Seed ovoid–oblong, 4.0–4.6 mm long, 1.5–2.0 mm wide; indumentum of fine white and ferruginous hairs. Aril head hooked to helmet-shaped, 0.6–1.0 mm long, white or brown and white, with a tuft of hairs. Aril appendages 3, oblong to linear, 0.20–0.65 mm long (Figs 2G, 3D).
Distribution
Australia (NT, WA). Recorded in WA from the Cockburn Range to the Ord River and in the NT from Dunmarra to the WA border (Fig. 7K).
Habitat
Found in woodlands on sandy soils often adjacent to creek lines. Predominantly occurs in red sand on flats in open woodlands, although also recorded in sandy clay loam adjacent to gullies, in riparian situations and on rocky slopes.
Notes
This species is very easily distinguished by the horned floral appendages and cleft style.
Nomenclatural notes
The specimen K 279777 is considered the holotype because it was the only specimen at K or MEL that matched the type citation.
Selected specimens (27 examined)
12. Polygala exsquarrosa Adema, Blumea 14: 268 (1967)
Lectotype (here designated): Endeavour R., R. Brown; lecto: BM 566301!; isolectotype: BRI 391425!, CANB 900371219!; remaining syntype: Upper Victoria R., F. Muell.; syn- K 356710!; excluded syntype: Upper Victoria R., F. Muell.; syn- K 356709!.
Polygala arvensis var. squarrosa Benth., Fl. Austral. 1: (1863); Polygala chinensis var. squarrosa (Benth.) Domin, Biblioth. Bot. 89: 303 (1927).
Annual herb, erect with spreading or ascending branches, up to 40 cm high; indumentum of mostly curved and straight hairs; stems. Leaves and alae often tinged maroon. Leaves linear to narrowly oblong or oblanceolate, 4–33 mm long, 1–5 mm wide; apex mostly acute or acuminate, rarely obtuse, mucro 0.3–0.4 mm long; petiole 0–1 mm long. Inflorescence supra-axillary or axillary, occasionally leaf-opposed, rarely terminal, a raceme, up to 200 mm long, although often much shorter ~25 mm long, exceeding the leaves or shorter than leaves. Pedicel up to 1 mm long. Alae position antrorse to perpendicular in fruit; herbaceous, narrowly elliptic to oblanceolate, dimidiate or falcate, 4.0–7.8 mm long, 1.0–2.2 mm wide; with straight hairs present and occasionally curved hairs also. Corolla purple; floral appendages fimbriate; upper petal claw ~0.7 mm wide. Stamens terminate at stigma, monadelphous; staminal flap <0.5 mm wide. Style curved and dorso-ventrally flattened, prominently grooved above globular stigma. Capsule reflexed or perpendicular, more or less aligned with alae or apex of alae diverging from capsule. Capsule lacking wing, usually asymmetrical, ovoid, 1.8–2.8 mm long, 1.4–2.5 mm wide, capsule lobe apices acute and separated. Seed ovoid, 1.7–2.8 mm long, 0.7–1.4 mm wide; indumentum of fine white and ferruginous hairs. Aril head reduced, 0.2–0.5 mm long, brown where reduced to seed testa and white where appendages originate, with hairs. Aril appendages 3, linear, 0.35–1.50 mm long (Figs 2E, 3B).
Distribution
Australia (NT, QLD, WA), Malesia and New Guinea. In Australia, this species is widely distributed north of 15°S in WA, NT and QLD (Fig. 7L).
Habitat
Usually found in eucalypt woodlands on rocky hillslopes or in Melaleuca shrublands, sedgelands or herbfields, often associated with sandy soils.
Nomenclatural notes
Adema (1966) did not see the Mueller specimen ‘Upper Victoria R.’ from K when he created a new name at species rank based on P. arvensis var. squarrosa. He did, however, cite a specimen with the same collection details from BM, which I have not been able to locate via the Australian Botanical Liaison Officer (ABLO) based at Kew. Adema (1966) did not lectotypify in this instance, perhaps because he did not see all the syntypes. Specimen K 356709 is a different species, P. bifoliata, and is thus excluded.
There must be an error in Bentham’s citation of collections for P. arvensis var. squarrosa as ‘Endeavour R., Brown’ in Flora Australiensis (Bentham 1863), because the Flinders expedition did not land there. They were most likely Banks’ specimens from the ‘Endeavour’ voyage that Brown had on the ‘Investigator’ as a working set (Alex George, ABLO, pers. comm., 2005).
Notes
This species is highly variable. Floral appendage divisions vary from once to many times divided, and gross morphology varies from single-stemmed specimens with short linear leaves to multi-stemmed specimens with longer and broader leaves. The inflorescence varies from short (~25 mm long) and congested, to long (up to 200 mm long) and loose. They also occur in two main habitats, stony hillslopes or wet swamps. Despite efforts to sort this species further, on the basis of habit, habitat and geography, no consistent combination of characters has allowed further differentiation. Specimens distributed in a very small area within Kakadu National Park, including Kapalga, Mardugal campground and Nourlangie, have a much more compact habit, longer leaves and a more reduced aril head. P. exsquarrosa is most similar to P. petrophila but can be distinguished from it by the smaller capsule and seed and the narrower alae.
Selected specimens (81 examined)
13. Polygala gabrielae Domin, Biblioth. Bot. 89: 304 (1927)
Type citation: Nord-Queensland: auf halbfreien Stellen in den Savannenwäldern bei Chillagoe in Gesellschaft zahlreicher Annuellen (Domin II. 1910); Type details: locis subnudis in xerodrymis prope opp. Chillagoe, K. Domin 5697; holo: PR 528257!
Annual herb, erect with ascending branches, up to 40 cm high; indumentum of curved hairs, rarely with a few straight hairs. Leaves linear, 12–66 mm long, 1.5–5.0 mm wide; apex acute, obtuse, mucro ~0.5 mm long; petiole 0–1 mm. Inflorescence supra-axillary or leaf-opposed, a raceme, up to 90 mm long, exceeding the leaves, but generally not extending past leaves at the top of the plant. Pedicel 3 mm long. Alae position perpendicular and deflexed in fruit; herbaceous, elliptic to ovate, slightly dimidiate to falcate, 3.5–6.0 mm long, 1.0–3.9 mm wide; with curved hairs. Corolla purple; floral appendages fimbriate; upper petal claw 1 mm wide. Stamens terminate at stigma, monadelphous; staminal flap 1 mm wide. Style distinctly hooked (horseshoe-shaped at apex), more or less terete, with flat stigma on inner surface of hook. Capsule deflexed, occasionally some reflexed, more or less aligned with alae or apex of alae slightly diverging from capsule. Capsule lacking wing, more or less symmetrical, oblong to squarish, 4.25–5.00 mm long, 3–4 mm wide; capsule lobe apices rounded and separated. Seed ovoid-oblong, 3.75–4.00 mm long, 1.50–1.65 mm wide; indumentum of fine white becoming ferruginous hairs. Aril head hooked to round, 0.50–0.65 mm long, white or white and brown, with a tuft of hairs. Aril appendages 3, oblong to linear, 1.0–1.5 mm long (Figs 11A–C, 13B).

|
Distribution
Australia (QLD). This species is known only from a restricted area around Chillagoe and Undara Lava Tubes, in far-northern QLD (Fig. 8A).
Habitat
Only known from a few collections where it is recorded growing in eucalypt woodlands.
Notes
In Australia this species is very similar to P. scorpioides and P. macrobotrya. It differs from P. macrobotrya by the larger capsules and shorter inflorescence. From P. scorpioides it differs by the erect, single-stemmed rather than multi-stemmed sprawling to ascending habit and the indumentum’s mostly lacking straight hairs. The above differences notwithstanding, there are no discrete character states separating P. gabrielae, P. macrobotrya and P. scorpioides. P. gabrielae and P. macrobotrya are both geographically restricted and known from few collections. They occur on the southern edge of the range of P.scorpioides, which is distributed throughout Cape York Peninsula. Although on the available evidence, P. gabrielae and P. macrobotrya appear to be distinct species, it is recognised that further collecting may reveal that they represent extremes of one variable species complex related to P. scorpioides. Domin published the names P. gabrielae and P. macrobotrya in 1927, and until additional evidence is accumulated to support their merging, I am compelled to maintain them. P. macrobotrya is not a synonym of P. wightiana, as queried by Adema (1969).
Nomenclatural notes
In line with recommendation 9A.4 and Art. 37.3 of the International Code of Botanical Nomenclature (McNeill et al. 2006), the Specimen PR 528257 is considered the type, because it was the only specimen at PR, Domin’s home herbarium, which best matched the type citation in terms of the locality description.
Selected specimens (4 examined)
14. Polygala galeocephala R.A.Kerrigan, sp. nov.
Ab congeneribus in Australia haec species indumento pilis curvis non nisi, arillo galeiformi, crista florali lobis spathulatis, stylo uncato, foliis ellipticis, lanceolatis et obverse-ovatis, et sepalis lateralibus longioribus (5–8 mm longis) distinguenda.
Type: Bullo River Station, on road to homestead, 07.iii.2006, R.A. Kerrigan 949; holo: DNA 177025!; iso: BRI, CANB, K, MEL, NSW, PERTH.
Polygala rhinanthoides var. minor Benth., Fl. Austral. 1 : 140 (1863). Type citation: Upper Victoria R., F. Muell.; Type detail: rocks of the Upper Victoria R., F. Muell.; holo: -K279799!
Annual herb, erect with ascending branches, up to 20 cm high; indumentum of curved hairs. Occasionally leaves and alae tinged red. Leaves linear to elliptic, lanceolate to ovate, rarely obovate, 6–46 mm long, 2.5–14.0 mm wide; apex acute, obtuse or occasionally emarginate, mucro ~0.4 mm long; petiole 0–1 mm long. Inflorescence supra-axillary or axillary, sometimes a solitary flower arising along internodes or usually a raceme, up to 70 mm long, usually not exceeding the leaves, sometimes longer than leaves. Pedicel up to 3 mm long. Alae position perpendicular in fruit; Alae herbaceous, ovate, elliptic or obovate, dimidiate, 4.95–8.00 mm long, 1.9–4.2 mm wide; glabrous or with curved hairs. Corolla purple; floral appendages fimbriate; upper petal claw ~0.7 mm wide. Stamens terminate at stigma, monadelphous; staminal flap ~0.6 mm wide. Style hooked (horseshoe-shaped at apex), more or less terete, becoming dorso-ventrally flattened below hook and twisting in apical half, with flat stigma along inner surface of hook. Capsule perpendicular, becoming reflexed, more or less aligned with alae, sometimes apex of alae diverging from capsule. Capsule lacking wing, oblong to obovate, 4–5 mm long, 2.7–3.5 mm wide; capsule lobe apices rounded and separated. Seed ovoid–oblong, 3.4–4.2 mm long, 1.1–1.5 mm wide; indumentum with fine white or ferruginous hairs. Aril head helmet-shaped, 0.6–1.0 mm long, white or white and brown, rarely all brown, with short erect hairs. Aril appendages 3, linear, 0.4–2.0 mm long.
Distribution
Australia (NT, WA). This species occurs throughout the Kimberley in WA and the western half of the NT, as far south as Supplejack Station (northern Tanami) and north to Bradshaw Station (Victoria River District). Although the identification of B.G. Thomson 589 (NSW) from Mallapunyah Station, Kilgour Gorge, has been confirmed as P. galeocephala, it has not been re-collected east of Gregory National Park (Fig. 8B).
Habitat
Found in a variety of habitats, mostly in woodlands on rocky sandstone hill slopes.
Notes
When in bud, keel and floral appendages look larger than lateral flag petals. However, this is not the case when flowers are fully mature. One specimen from WA, Warralong Station, Burbidge NT 737 (PERTH), has affinities with this species (sharing a similar habit, indumentum, leaf shape, seed shape and aril shape) but has a more open inflorescence similar to P. gabrielae. Further collecting in the area is required to determine the extent of this variation.
Etymology
Latin galea- (helmet) and Greek -cephalus (head), referring to the helmet shape of the aril.
Selected specimens (50 examined)
15. Polygala geniculata R.A.Kerrigan, sp. nov.
Polygalae clavistylae affinis, a qua stigmate plano et stylo uncato differt. Ab congeneribus in Australia indumento pilis curvis non nisi, arillo seminis redacto, crista florali lobis spathulatis vel coraliformibus, stylo uncato, foliis obverse-ovatis, et capsulis non pendulinis distinguenda.
Type: c. 5 km SE of Lake Bennett, 02.iv.2003, I.D. Cowie 9688; holo: DNA 157569!; iso: BRI, CANB, K, MEL, NSW, PERTH.
[Polygala glaucoides [NT material] auct. non. L. (1753), I.D. Cowie & D.A. Albrecht (eds), Checklist N. Terr. Vasc. Pl. Sp. (2004).]
Annual herb, erect with ascending branches, up to 20 cm high; indumentum of curved hairs. Stems and leaves often tinged maroon. Leaves obovate, 5–30 mm long, 3–12 mm wide; apex acute, obtuse or rounded, rarely some leaves truncate or emarginate, mucro 0.3–0.5 mm long; petiole 0–1 mm long. Inflorescence supra-axillary, axillary or terminal, a raceme, up to 35 mm long, exceeding the leaves, equal to leaves or shorter than leaves. Pedicel up to 2.5 mm long. Alae position antrorse to perpendicular in fruit; herbaceous, ovate, strongly dimidiate, 4–8 mm long, 2–4 mm wide; glabrous or with curved hairs. Corolla purple; floral appendages spathulate, rarely approaching coralliformis; upper petal 1.3–1.7 mm wide. Stamens terminate at stigma, monadelphous; staminal flap 1 mm wide. Style hooked (horseshoe-shaped at apex), more or less terete at base, becoming dorso-ventrally flattened below hook and twisting in apical half, with flat stigma along inner surface of hook. Capsule perpendicular, not reflexed, more or less aligned with alae, sometimes apex of alae diverging ~30° from capsule. Capsule lacking wing, more or less symmetrical, ovoid, 3.5–4.3 mm long, 3.5–4.3 mm wide; capsule lobe apices obtuse or subacute and separated. Seed ovoid, 2.7–3.4 mm long, 1.5–1.9 wide; indumentum of fine white and ferruginous hairs. Aril head reduced, 0.4–0.5 mm long, brown where reduced to seed testa and white where appendages originate, with short hairs. Aril appendages 3, spathulate, 1.5–2.5 mm long.
Distribution
Australia (NT). This species is endemic to the NT and is known only from a restricted area from Hayes Creek, Litchfield National Park, outer Darwin and the Mary River, with an outlier specimen recorded from Bradshaw Field Training Area, I.D. Cowie 11469 (BRI, DNA, PERTH) (Fig. 8C).
Habitat
Found in woodlands, usually on stony hill slopes amongst laterite, quartzite or shale rubble.
Notes
This species is very similar to P. clavistyla but can be distinguished from that species by the hooked style and flat stigma. The outlier specimen, I.D. Cowie 11469, differs from typical P. geniculata by the narrowly elliptic leaves and more coralliform floral appendages.
Etymology
Latin geniculatus (hooked or bent), referring to the hooked style rather than club-shaped style.
Selected specimens (13 examined)
16. Polygala glaucifolia R.A.Kerrigan, sp. nov.
Polygalae isingii similis, a qua habitu rotundato, foliis glaucis, arillo redacto et pilis mollioribus differt.
Type: 70 km N of Newman on main road to Port Headland, Hamersley Ranges, 06.v.2006, R.A. Kerrigan 1093; holo: DNA 177046!; iso: AD, BRI, CANB, PERTH.
Annual herb, rounded with spreading and decumbent branches, up to 11 cm high; indumentum of curved and soft straight hairs. Leaves obovate, oblanceolate, 2.5–21.0 mm long, 1–10(–14) mm wide; apex rounded, truncate, retuse or emarginate, mucro ~0.2 mm long; petiole up to 5 mm long. Inflorescence axillary in leaf and branch axils, a raceme, up to 11 mm long, not exceeding the leaves. Pedicel up to 2 mm long. Alae position antrorse to perpendicular in fruit; herbaceous, ovate to lanceolate, dimidiate, 2.0–5.2 mm long, 1.2–2.5 mm wide; with curved and straight hairs or straight hairs only. Corolla light blue to purple or pink; keel rarely sparsely pilose; floral appendages fimbriate; upper petal claw 1 mm wide. Stamens terminate at stigma, monadelphous; staminal flap not visible up to 0.5 mm wide. Style hooked (horseshoe-shaped at apex), beaked at apex of bend, more or less terete at base, becoming dorso-ventrally flattened below hook and twisting in apical half, with flat stigma along inner surface of hook. Capsule antrorse to perpendicular, apex of alae diverging at ~30° from capsule. Capsule lacking wing, symmetrical, elongate ovoid, 3.0–5.5 mm long, 2.5–4.0 mm wide; capsule lobe apices obtuse and touching laterally. Seed elongate ovoid, 3.50–4.25 mm, 1.25–1.65 mm wide; indumentum of fine white or ferruginous hairs. Aril head reduced, 0.5–1.0 mm long, brown where reduced to seed testa and white where appendages originate, with short erect hairs. Aril appendages 3, linear, 0.7–1.0(–2.8) mm long (Fig. 5H).
Distribution
Australia (NT, QLD, SA, WA). Occurs in the arid zones of NT, QLD, WA and SA, extending from coastal regions in WA to around the shared borders of NT, QLD and SA, possibly also present in the arid zone of north-western NSW (Fig. 8D).
Habitat
Found in a variety of habitats including Acacia shrublands and Triodia grasslands, generally associated with gravelly or rocky slopes and foot hills but also found in drainage lines, flats and subsaline conditions, often on loam, sandy loam or clay.
Notes
This species is similar to P. isingii and P. difficilis but differs from both by having a prostrate to spreading compact habit, greyish foliage when fresh and a reduced aril head. Four specimens, J. Devitt 171 (DNA), M.E. Trudgen 15945 & 6995 & 1155 (PERTH), from Mount Skinner (NT), Hammersley National Park (WA) and Yampire Gorge are considered atypical specimens of this species and are not included in the above description. Although these specimens share many of the typical characters of P. glaucifolia including, importantly, the seed characters, they have strikingly narrow and longer leaves and in gross morphology appear similar to P. dependens.
Etymology
Latin glaucus- (blue grey) and -folium (leaf), referring to the blue-grey colour of the leaves.
Selected specimens (54 examined)
17. Polygala glaucoides L., Sp. Pl. 2: 705 (1753)
Type: ‘Habitat in Zeylona; Fl. Zey. 270’; lecto: BM 621889 (photograph DNA!); fide D.B.Sumithraarachchi, Rev. Hand. Fl. Ceylon 6 : 310 (1988).
Annual herb, prostrate or erect with spreading or decumbent branches, up to 20 cm high; indumentum of curved hairs. Leaves oblanceolate, narrowly elliptic or narrowly oblong, 4–30 mm long, 2–8 mm wide; apex rounded or truncate, mucro ~0.2 mm; petiole 0–1 mm long. Inflorescence supra-axillary, a raceme, up to 2 mm long, not exceeding the leaves. Pedicel up to 0.5 mm long. Alae antrorse or perpendicular in fruit; herbaceous, ovate to lanceolate, falcate or dimidiate, 4.3–5.8 mm long, 2.0–2.4 mm wide; with curved hairs or glabrous. Corolla yellow (from dried material only); floral appendages fimbriate; upper petal ~0.5 mm wide. Stamens terminate at stigma, monadelphous; staminal flap <0.5 mm wide. Style hooked (horseshoe-shaped at apex), in dried state style laterally compressed, with flat stigma along inner surface of hook. Capsule perpendicular to deflexed, apex of alae diverging ~30° from capsule. Capsule lacking wing, more or less symmetrical, widely elliptic to widely ovoid, 4.0–4.5 mm long, 2.8–3.2 mm wide; capsule lobe apices rounded and separated. Seed ovoid–oblong, 3.0–3.5 mm long, ~1.5 mm wide; indumentum of fine white hairs. Aril head round, 0.55–0.80 mm long, white, with short erect hairs. Aril appendages 3, linear, up to 0.8 mm long.
Distribution
A very widespread species distributed from Sri Lanka to continental South-east Asia to Australia (QLD). In Australia, it is known from only a few collections from Rockingham Bay near Cardwell, South Molle Island and the Herbert River. Attempts to relocate the populations near Cardwell in March 2007 failed (Fig. 8E).
Habitat
With so few collections, it is difficult to determine the habitat of this species in Australia. Van der Meijden (1988) reported that this species occupies grassy places at low altitudes. The lack of collections may reflect the fact that much of the accessible coastal lowland savanna is under cultivation.
Notes
This species very similar to P. polifolia and P. triflora, but is easily distinguished from the former by the fusion of stamens in a bundle of eight rather than three bundles (3, 2, 3) and by a hooked style rather than a curved style with an apical point. It is distinguished from P. triflora by the absence of straight hairs on the capsule margin. With so few collections of this species in Australia and with such a poor type for P. glaucoides, it is difficult to assign the specimens of the taxon described here unequivocally to P. glaucoides. Using the description of van der Meijden (1988), there is little evidence to suggest it is not P. glaucoides. Issues regarding synonymy of this species and other taxa mentioned by van der Meijden (1988) cannot be fully resolved until a monograph of the group is completed and specimens throughout the range are examined. Reports on flower colour based on dried material should be treated with caution.
Selected specimens (5 examined)
18. Polygala integra R.A.Kerrigan, sp. nov.
Polygalae crassitestae et P. pterocarpae affinis, a quibis differt crista florali integra, stigmate plano ad apicem styli, et stylo non uncato. Ab congeneribus in Australia crista florali integra, stigmate plano ad apicem styli, et stylo non uncato distinguenda.
Type: 250 m NE along Green Ant Ck towards East Springs, off Adelaide River Scenic Hwy, 02.ii.2004, R.A. Kerrigan 725 & R.K. Harwood; holo: DNA 161692!; iso: BRI, CANB, NT, PERTH.
[Polygala eriocephala [NT material] auct. non F.Muell. ex Benth. (1863), I.D. Cowie & D.A. Albrecht (eds), Checklist N. Terr. Vasc. Pl. Sp. (2004).]
Annual herb, erect with ascending branches, rarely decumbent, up to 80 cm high; indumentum of curved and straight hairs. Base of stem often tinged red or maroon. Leaves linear to oblong, narrowly elliptic or lanceolate, 8–70 mm long, 1–7 mm wide; apex acute, obtuse or rounded, mucro 0–0.5 mm long; petiole 0–1 mm long. Inflorescence supra-axillary or leaf-opposed, a raceme, up to 60(–80) mm long, usually not exceeding the leaves, sometimes longer than leaves. Pedicel up to 1.5 mm long. Alae position perpendicular, rarely antrorse or deflexed in fruit; herbaceous, broadly ovate, dimidiate, 3.5–7.0 mm long, 2–5 mm wide; with curved and long straight hairs. Corolla purple or blue; floral appendages entire and bent at right angles to apex of keel to form a horizontal platform; upper petal claw 1.5 mm wide. Stamens terminate well below stigma, monadelphous; staminal flap ~0.5 mm wide. Style curved, slightly capitate at apex, more or less terete at base, becoming dorso-ventrally flattened below hook and twisting in apical half, with flat stigma on inner surface immediately below apex. Capsule deflexed, apex of alae diverging at ~45° from capsule. Capsule with wing, wing 0.2–1.4 mm wide, usually widest at apex of locules or capsule lobes, more or less symmetrical, very widely ovate to very widely obovate or orbicular to squarish, 3.2–4.4 mm long, 3.5–4.0 mm wide, capsule lobe apices rounded and separated. Seed ovoid, 3.0–3.5 mm long, 1.5–1.9 mm wide; indumentum of fine white becoming ferruginous hairs. Aril head round, ~0.5 mm long, brown cap on white, with a tuft of hairs. Aril appendages 3, spathulate, rarely linear, 0.75–2.3 mm long (Figs 2D, 3A, 5B).
Distribution
Australia (NT). Endemic to the NT, this species is widely distributed across the far north of the NT, occurring mostly north of and including Nitmiluk National Park and extending to north-eastern Arnhem Land and offshore islands (Fig. 8F).
Habitat
Found in open woodlands in a variety of substrates or often along riparian or wetland margins.
Notes
This species has been previously misidentified as P. eriocephala, P. rhinanthoides and P. triflora, largely as a result of misapplication of these names. This species is superficially similar to P. pterocarpa and erect forms of P. crassitesta, but is easily distinguished from all other Polygala in Australia by having entire floral appendages, bent perpendicularly in the apical half, and a simple curved style and flat stigma.
Etymology
Latin integer (entire), referring to the undivided entire floral appendages.
Selected specimens (52 examined)
19. Polygala isingii Pedley, J. Adel. Bot. Gardens 3: 128 (1981)
Type: 5 miles [8 km] S of Mt Barkly, Barkly Station, 14.i.1971, P.K. Latz 1224; holo: BRI 118100!; iso: NT.
Annual or short-lived perennial with a well developed tap root, tufted with ascending and decumbent branches, up to 13 cm high; indumentum of curved and stiff straight hairs. Leaves narrowly oblong, oblanceolate or obovate, 5–22 mm long, 2–10 mm wide; apex usually rounded or obtuse, occasionally truncate or emarginate, rarely acute, mucro 0.3–0.4 mm long; petiole up to 1 mm long. Inflorescence axillary, a solitary flower or more or less fasciculate, rarely a short raceme, up to 4 mm long, not exceeding the leaves. Pedicel up to 3 mm long. Alae position antrorse to perpendicular in fruit; herbaceous, elliptic to spathulate, 3.5–5.0 mm long, 1.5–2.7 mm wide; with curved and straight hairs. Corolla purple; floral appendages fimbriate; upper petal ~0.6 mm wide. Stamens terminate at stigma, monadelphous; staminal flap <0.5 mm wide. Style hooked (horseshoe-shaped at apex), beaked at apex, in dried state style dorso-ventrally flattened at base and below hook, laterally compressed in middle, with flat stigma along inner surface of hook. Capsule antrorse to perpendicular, more or less aligned with alae. Capsule lacking wing, more or less symmetrical, ovoid, 4.2–5.2 mm long, 2.5–4.1 mm wide; capsule lobe apices rounded and separated. Seed elongate ovoid, 3.6–4.7 mm long, 1.2–1.7 mm wide; indumentum of fine white or ferruginous hairs, becoming shorter towards apex, or shorter for 1/2 to 2/3 of the seed. Aril head helmet-shaped with central groove, 0.9–1.0 mm long, white and brown where reduced to seed testa, with short erect hairs. Aril appendages 3, linear, 0.7–1.0 mm long (Figs 5E, 6B).
Distribution
Australia (NT, WA). This species is widely distributed across the arid zones of the NT and WA, in the Tanami, Gibson, Great Sandy and Little Sandy Deserts (Fig. 8G).
Habitat
Found growing in gravelly, sandy or shallow soils on hills in hummock grasslands.
Notes
Two specimens from western Tanami Desert, D.E. Albrecht 10887 & K.G. Brennan (NT) and D.E. Albrecht 10660 (NT) and one from the Great Sandy Desert, A.S. George 14705 (PERTH), included in the present description, are distinguished by having a thicker tap root, more robust perennial habit and a seed indumentum with short hairs extending across more of the testa (usually at least 2/3 compared with less than half). Further survey and more specimens may result in their recognition as a separate taxon in the future. However, it appears that they represent longer-lived individuals which may have escaped fire or had greater access to water.
This species can be difficult to distinguish from P. glaucifolia and P. triflora. From the former, it can be recognised by the following: indumentum, which is shorter and more hispid in P. isingii; vegetative colour, which is generally grey–green in P. glaucifolia; a more erect tufted habit; and a more developed aril head on the seed, which is reduced to where aril appendages attach in P. glaucifolia. P. triflora is geographically disjunct from P. isingii. It can generally be distinguished by its longer inflorescences. However, the only specimen east of 135°, D.E. Albrecht 9789 (NT) from the Dulcie Ranges, is difficult to place because it has longer inflorescences, approaching those of P. triflora. A depauperate specimen, it is tentatively placed under P. isingii, and further collecting in the area is required to resolve the confusion.
Selected specimens (34 examined)
20. Polygala japonica Houtt., Handl. Plant Kruidk. 10: 89, t. 62, fig. 1 (1779)
Type: none cited; from Japan n.v.; fide Adema, Blumea 14: 263 (1966).
Polygala veronicea F.Muell., Transactions and Proceedings of the Victorian Institute for the Advancement of Science 1: 117–118 (1855); Polygala veronicifolia Chodat, Monogr. Polygal. 2:355 (1893), orth. var. Type citation: in grassy or gravelly places from the King R. to the Goulburn R.; Type detail: In pratis ad flumen Goulburn juxta Wararegarwan, [F. Muell.?]; syn- MEL 2263627!, King R., [F. Muell.?]; syn- MEL 22636261!; King R., F. Muell.; syn- K (photograph DNA!); fide R.van der Meijden, Fl. Malesiana I (10): 472 (1988).
[Polygala sibirica non L. (1753), Ewart, Fl. Victoria 715 (1931).]
Perennial herb or subshrub, diffuse prostrate or branches ascending or erect, up to ~30 cm high; indumentum of curved hairs. Leaves elliptic to ovate, 5–20 mm long, 3–10 mm wide; apex acute to obtusely angled, mucro 0–0.2 mm long, mucro 0–0.2 mm; more or less sessile. Inflorescence leaf-opposed or supra-axillary, sometimes a solitary flower usually a short raceme, up to 30 mm long, exceeding the leaves or shorter than leaves. Pedicel to 2–6 mm long. Alae position ascending to perpendicular in fruit; mauve to blue, ovate to obovate, 4–6 mm long, 1.5–2.5 mm wide; glabrous. Corolla purple and cream; floral appendages much divided; upper petal claw 1 mm wide. Stamens terminate at stigma, nearly connate to apex, monadelphous; staminal flap ~0.75 mm wide. Style more or less straight becoming bent at apex, with 2 stigmas, 1 apical and globular (±sterile) and the other lateral and globular, in dried state style laterally compressed below lateral stigma, dorso-ventrally compressed above. Capsule perpendicular to deflexed, more or less aligned to alae or apex of alae diverging up to ~30° from capsule. Capsules perpendicular, more or less aligned with alae or diverging <30°. Capsule with wing, wing ~0.5 mm wide, more or less equal throughout, orbicular to broadly obcordate, 3–8 mm long, 3–8 mm wide; capsule lobe apices rounded and separated. Seed ovate, ~2.5–3.0 mm long, ~2 mm wide; indumentum of fine white or ferruginous hairs. Aril head hooked, ~0.6 mm long, white, with short hairs. Aril appendages 3, linear to oblong, ~1 mm long.
Distribution
Australia (ACT, NSW, QLD, VIC), India, Burma, Sri Lanka, Indochina, Japan, Korea, Siberia, China, Malesia, the Philippines and New Guinea. This species is widespread in eastern Australia from southern QLD to VIC (Fig. 8H).
Habitat
Found in woodlands in loam and clay loam.
Notes
This species was not revised in the present study; however, it needed to be included to provide an account of all Australian natives. The species concept of van der Meijden (1988) has been followed here because it was possible to examine only a small number of specimens. The above description has been augmented where necessary from Flora of NSW (Murray 1992), Flora of Victoria (Walsh 1999), Adema (1966), van der Meijden (1988) and Monro (2003), and the species concept sensu van der Meijden (1988) is maintained. For a full list of synonymy see van der Meijden (1988), because only Australian synonyms are listed above. Typification follows Adema (1966).
Nomenclatural notes
Question marks are recorded against Mueller collections where no collector’s name has been recorded on the specimen, and I have assumed the collector to be Mueller.
Selected specimens (5 examined)
21. Polygala kimberleyensis R.A.Kerrigan, sp. nov.
Polygalae barbatae similis, a qua arillo galeiformi differt. A P. dimorphotricha folliis latiorioribus et brevioribus, sepalis lateralibus brevioribus et angustatioribus differt. A P. obversa indumento pilis rectis differt.
Type: Kimberley, 3.8 km E of Adcock Gorge turnoff on Gibb River Rd, 13.v.2006, R.A. Kerrigan 1111; holo: DNA 176670!; iso: BRI, CANB, PERTH.
Annual herb, erect often with spreading or decumbent branches, or prostrate, up to 30 cm high; indumentum of curved and straight hairs. Leaves very widely ovate, obovate to depressed obovate or orbicular, 3–38 mm long, 4–25 mm wide; apex rounded, mucro 0.3–0.4 mm long; petiole 0–3 mm long. Inflorescence axillary or supra-axillary, a solitary flower usually a short raceme, up to 10 mm long, not exceeding the leaves. Pedicel up to 0.5 mm long. Alae position antrorse to perpendicular, sometimes deflexed in fruit; herbaceous, ovate, usually somewhat dimidiate, 3.8–5.0 mm long, 1.5–2.5 mm wide; with mostly straight and some curved hairs. Corolla purple; floral appendages fimbriate; upper petal claw ~0.5 mm wide. Stamens terminate at stigma, monadelphous; staminal flap <0.25 mm wide. Style hooked (horseshoe-shaped at apex), in dried state laterally compressed, with flat stigma along inner surface of hook. Capsule antrorse to perpendicular, sometimes deflexed, more or less aligned with alae. Capsule lacking wing, if present wing ~0.5 mm wide, more or less equal throughout, more or less symmetrical, very widely ovate to widely oblong or squarish, 3.7–5.5 mm long, 3.5–4.9 mm wide; capsule lobe apices rounded and separated. Seed ovoid, 3.15–5.00 mm long, 1.6–2.5 mm wide; indumentum of fine white or ferruginous hairs or mostly glabrous. Aril head helmet-shaped, 0.5–1.0 mm long, white or white and brown, with short hairs. Aril appendages 3, linear, 0.9–1.5 mm long (Fig. 5D).
Distribution
Australia (WA). Endemic to WA, this species is known only from a few collections scattered across the Kimberley and extending to coastal islands (Fig. 8I).
Habitat
Found in eucalypt woodlands associated with sand among sandstone.
Notes
This species is similar to P. barbata, from which it is readily distinguished by the helmet-shaped aril head and fimbriate floral appendages. This species differs from P. obversa by having an indumentum with both curved and straight hairs. Style characters from dried material need confirmation with fresh material.
Etymology
Latinised name, kimberleyensis, in reference to the geographic region where this species is found.
Selected specimens (6 examined)
22. Polygala longifolia Poir., Encyclopedie Methodique, Botanique 5: 501 (1804)
Type: Cette plante a été rapportée par Commerson de l’île de Java (v.s. in herb. Lamarck); holo: P-LA n.v.; iso: FI n.v.; fide F. Adema, Blumea 14 : 266 (1966).
Polygala leptalea DC., Prodr. 1 : 325 (1824). Type: ‘in Napaulia. Wallich’; holo: G-DC (photograph DNA!).
Polygala leptalea var. australiensis Domin, Biblioth. Bot. 89: 301 (1927); Type: Nord-Australien: Port Darwin, F. Schultz No. 276; holo: MEL37310!
Polygala abyssinica var. intercedens Domin, Biblioth. Bot. 89: 301 (1927); Type: Nord-Australien: Port Essington, Armstrong No. 365; syn- K (photograph!); Upper Victoria R., F. Muell.; syn- K (photograph!); fide A.M. Monro, Systematics of the Australian Polygalaceae and Xanthophyllaceae. Unpublished Thesis. ANU (2003).
[Polygala abyssinica [Australian material] auct. non Fresen (1837): K. Domin, Biblioth. Bot. 89 : 301 (1927)]; fide A.M. Monro, Systematics of the Australian Polygalaceae and Xanthophyllaceae. Unpublished Thesis. ANU (2003).]
Annual herb, erect occasionally with ascending branches, up to 120 cm high; indumentum of curved hairs, glabrescent. Stems and leaves often tinged red or maroon especially at base of plant. Leaves more or less caducous, plant often leafless or with only a few leaves, linear to lanceolate, sometimes oblanceolate to obovate leaves towards the very base of plant, 5–32 mm long, 0.5–7.0 mm wide; apex acute, mucro 0–0.3 mm long; petiole 0–0.5 mm long. Inflorescence terminal, a raceme, up to 300 mm long, greatly exceeding the leaves. Pedicel up to 2 mm long. Alae position perpendicular to reflexed in fruit; petaloid, pink or white with maroon veins, obovate, oblanceolate, 3.0–6.4 mm long, 1.6–3.3 mm wide; glabrous. Corolla white to pink; floral appendages fimbriate; upper petal claw ~0.6 mm wide. Stamens more or less terminate at stigma, monadelphous; staminal flap not visible. Style curved, more or less terete, with flat stigma on inner surface directly below apex. Capsule perpendicular to reflexed, more or less aligned with alae. Capsule lacking wing, if present wing up to 0.5 mm wide, generally wider at apex, more or less symmetrical, obovate, 3–5 mm long, 1.5–2.8 mm wide; capsule lobe apices rounded and separated. Seed oblong, 1.8–3.3 mm long, 0.6–1.0 mm wide; indumentum of fine white hairs. Aril head helmet-shaped, 0.3–0.5 mm long, white with brown groove where reduced to seed testa, short hairs present and often concentrated along the groove. Aril appendages 3, absent or much reduced triangular projections, 0.25 mm long.
Distribution
Australia (NT, QLD, WA), South-east Asia and Malesia. This species is very widely distributed across NT, QLD and WA, generally north of 19°S. A specimen held at CANB, which places this species in Alice Springs in 1974 (D. Verdon 778A), is treated as dubious because the species has not been re-collected from this area, which is well south of the known distribution (Fig. 8J).
Habitat
Found in a variety of habitats, including eucalypt woodlands, Acacia and Melaleuca shrublands, drainage depressions and seepage areas. Grows in a variety of substrates including laterite, sandstone, skeletal soils, sandy loam, basalt, quartzite and clay.
Notes
Readily distinguished by having petaloid, obovate alae (rounded to obtuse at apex), small flowers and erect terminal racemes.
Selected specimens (107 examined)
23. Polygala macrobotrya Domin, Biblioth. Bot. 89: 302 (1927)
Type citation: Nord-Queensland: Savannenwälden am Walsh R. nördlich von Chillagoe (Domin II. 1910); Type details: in xeryodrymiis apud Walsh R. situ septendr. ab opp. Chillagoe, K. Domin 5690; holo: PR 528251! (photograph CANB, DNA!).
Annual herb, erect with ascending branches, up to 40 cm high; indumentum of curved hairs. Leaves linear, 12–58 mm long, 1–5 mm wide; apex acute to rounded, mucro 0.2–0.3 mm long; petiole 0–1 mm long. Inflorescence supra-axillary or leaf-opposed, a raceme, up to 200 mm long, exceeding the leaves, well past the leaves at the top of the plant. Pedicel 2 mm long. Alae position perpendicular to deflexed in fruit; herbaceous, narrowly elliptic to oblanceolate, slightly dimidiate to falcate, 3.0–4.5 mm long, 1–2 mm wide; with curved hairs. Corolla purple; floral appendages fimbriate; upper petal claw 1 mm wide. Stamens terminate at stigma, monadelphous; staminal flap ~0.7 mm wide. Style distinctly hooked (horseshoe-shaped at apex), more or less terete, with flat stigma along inner surface of hook. Capsule perpendicular or deflexed (occasionally reflexed), apex of alae diverging ~30° from capsule. Capsule lacking wing, more or less symmetrical, oblong to squarish, 3.5–4.5 mm long, 2.7–3.4 mm wide; capsule lobe apices rounded and separated. Seed ovoid–oblong, 3.5–4.0 mm long, 1–2 mm wide; indumentum of fine white hairs. Aril head hooked, 0.6–0.8 mm long, white and brown with a tuft of hairs. Aril appendages 3, oblong to linear, 0.8–0.9 mm long (Fig. 13A).
Distribution
Australia (QLD). This species is known only from a restricted area around Chillagoe in far-northern QLD (Fig. 8K).
Habitat
Found in eucalypt woodlands (ironbark) associated with outcropping granite.
Notes
In Australia, this species is very similar to P. stenoclada and P. gabrielae (see notes under this species). It differs from P. stenoclada by the shape of the aril head (helmet-shaped in P. stenoclada), the centralised tuft of hairs on the aril head, the smaller capsules and narrower leaves.
Nomenclatural notes
According to Domin (1927), this species is apparently very similar to P. elongata from India. I have seen a photograph of the type of P. elongata, as determined by van der Meijden (1988) and, unlike P. macrobotrya, which has a single erect stem, the type specimen of P. elongata has spreading to ascending lateral branches. Domin (1927) indicated that compared with P. elongata, P. macrobotrya differs by its longer racemes, dull and not shiny leaves, corolla colour, capsule shape (not dilating at the apex) and by the longer aril appendages. Although van der Meijden (1988) synonymised P. elongata under P. glaucoides, the specimen B-W12975-010 at Herbarium Willdenow in Berlin (B-W), which is determined by him as the holotype of P. elongata, has a second determination slip also signed by him, labelled ‘P. elongata Willd. (non Adema 1966)’, and yet has no determination for P. glaucoides. Both van der Meijden (1988) and Bennett (1872) recorded flower colour as yellow for both P. glaucoides and P. elongata.
Specimens examined (5)
QUEENSLAND: 4 km E of Almaden on the road to Petford, 10.iii.1980, J.R. Clarkson 3039 (BRI, QRS); Walsh R., Chillagoe area, 01.ii.1910, K. Domin 5690 (PR); 26 km from Petford towards Almaden on N side of the road, 29.iii.2007, R.A. Kerrigan 1182 (DNA); Almaden Ck, picnic area on western bank of creek, N side of road, 29.iii.2007, R.A. Kerrigan 1183 (DNA).
24. Polygala obversa R.A.Kerrigan, stat. et nom. nov.
P. arvensis var. obovata Benth., Fl. Austral. 1 : 141 (1863); Polygala chinensis subvar. obovata (Benth.) Domin, Biblioth. Bot. 89: 303 (1927).
Type: Cavern I., Carpentaria, R. Brown 4972; holo: BM 929061!.
Annual herb, erect often with spreading branches or prostrate, up to 15 cm high; indumentum of curved hairs. Leaves obovate, varying to very widely obovate or depressed obovate, occasionally orbicular, 2.5–29.0 mm long, 2–24 mm wide; rounded to emarginate, mucro 0.3–0.4 mm long, petiole 0–4 mm long. Inflorescence axillary or supra-axillary, a solitary flower or raceme, up to 15 mm long, not exceeding the leaves. Pedicel to 2.5 mm long. Alae position antrorse to perpendicular in fruit; herbaceous, ovate, usually somewhat dimidiate, 2.9–6.8 mm long, 1.6–4.0 mm wide; with curved hairs or glabrescent. Corolla purple; floral appendages fimbriate and curved; upper petal ~1 mm wide. Stamens terminate at stigma, monadelphous; staminal flap 1.0–1.2 mm. Style hooked (horseshoe-shaped at apex), more or less terete in the middle and dorso-ventrally flattened at base and below hook, with flat stigma flat along inner surface of hook. Capsule antrorse to perpendicular, more or less aligned with alae. Capsule lacking wing, more or less symmetrical, very widely ovate to depressed ovate and widely elliptic, 3.0–4.7 mm long, 3.0–5.5 mm wide; capsule lobe apices rounded and separated. Seed ovoid, 2.2–4.2 mm long, 1.2–2.4 mm wide; indumentum mostly glabrous or with fine white or ferruginous hairs becoming shorter towards apex. Aril head round to hooked, 0.2–0.7 mm long, white or white and brown, with fine short hairs. Aril appendages 3, linear, 0.6–2.6 mm long (Fig. 2F).
Distribution
Australia (NT, QLD). Occurs from Kakadu National Park to Cobourg Peninsula and coastal regions of north-eastern Arnhem Land in the NT, and known from three collections around Mount Molloy and Cooktown in QLD (Fig. 8L).
Habitat
Mostly found in eucalypt woodlands in sand, sandy loam or sandstone, occasionally in coastal vine thickets on coastal sands or laterite.
Notes
This species is similar to P. barbata, P. coralliformis, P. dimorphotricha and P. kimberleyensis, and the distinguishing characters are discussed under those species. The upper petal claw in this species is not adnate to the staminal sheath throughout.
Nomenclatural notes
A sheet of P. arvensis var. obovata at K, barcoded as two entities, K279789 and K279790, has the same number (4972) as the holotype at BM929061. The specimen K279789 from Carpentaria is the same species as K279790 and may or may not be the same collection. It is unclear whether the K material is the same gathering as the BM material, and these specimens are not included as isotypes here. The epithet, obovata, could not be elevated to species rank in this instance as it is preoccupied by Polygala obovata A.St.-Hil.
Etymology
Latin obversus (broader at or towards the top than towards the base), referring to the obovate leaves of this species.
Selected specimens (37 examined)
25. Polygala orbicularis Benth., Fl. Austral. 1 : 140 (1863)
Polygala chinensis var. orbicularis (Benth.) Domin, Biblioth. Bot. 89: 302 (1927).
Lectotype (here designated): Melville I., Fraser 244; lecto: K 000279783 & K 000279784 (photograph DNA!); remaining syntypes: N. Coast, Armstrong; syn- K 000279787 (photograph CANB, DNA!); excluded syntypes: South Goulburn I., A. Cunningham K 000279786 (photograph DNA!), K 000279788! (photograph DNA!), BM 929045!
Annual herb, erect with decumbent branches, up to 10 cm high; indumentum of curved hairs, leaves mostly glabrescent. Leaves and alae often tinged deep purple, usually strongest on lower leaf surface. Leaves orbicular, depressed orbicular, very widely ovate or very widely obovate, 4–42 mm long, 3–48 mm wide; apex obtuse, rounded, truncate or emarginate, mucro 0.1–0.2 mm long; petiole up to 2.5 mm long. Inflorescence supra-axillary, rarely axillary or leaf-opposed but often appearing terminal, rarely a solitary flower usually a raceme, up to 25 mm long, not exceeding the leaves. Pedicel up to 3 mm long. Alae position antrorse to perpendicular, rarely deflexed in fruit; herbaceous, ovate, somewhat dimidiate, 4.2–7.8 mm long, 2.0–4.6 mm wide; glabrous or with curved hairs. Corolla purple; floral appendages fimbriate; upper petal claw 1–1.2 mm wide; keel petal with slight lateral pocket development or pocket absent. Stamens terminate at stigma, monadelphous; staminal flap 1–1.2 mm wide. Style hooked (horseshoe-shaped at apex), more or less terete, with flat stigma along inner surface of hook. Capsule antrorse to perpendicular, more or less aligned with alae. Capsule lacking wing, if present wing up to 0.45 mm wide, more or less equal throughout, widely ovoid, 2.8–4.0 mm long, 3.2–4.3 mm wide; capsule lobe apices rounded and separated. Seed ovoid, 2.7–3.2 mm long, 1.5–2.0 mm wide; indumentum of white or ferruginous hairs often patchily distributed across testa. Aril head round, 0.5–0.6 mm long, white, with short hairs or glabrous. Aril appendages 3, linear, 1.2–2.5 mm long.
Distribution
Australia (NT). This species is endemic to the NT and is distributed across the north-eastern part of the far north of the NT, from Litchfield to Kakadu NP, Cobourg Peninsula and the Tiwi Islands (Fig. 9A).
Habitat
Found in eucalypt woodlands or Melaleuca shrublands, mostly in lateritic or sandy soils.
Nomenclatural notes
Excluded syntypes are now determined as P. obversa. The collector of the lectotype in this instance is likely to be John Richardson because there is no record of Charles Fraser having visited Melville Island (Short 2002). There is no evidence to suggest that K279783 and K279784 are not part of the same gathering, and they are consequently treated here as one collection. A specimen, MEL2244771, from Goulburn Island, collected by Cunningham (No. 166) may be an isosyntype of the excluded syntypes at K and BM. It is apparent that an element from the MEL sheet has been cut out and removed, and the far left-hand element of K000279788 has clearly been pasted on from another source.
Selected specimens (56 examined)
26. Polygala paniculata L., Syst. Nat. 10th edn, 1154 (1759)
Type: Brown. Jam. 287 n.v.; lecto: Patrick Browne, Linn 882.9 LINN (photograph!); fide F. Fawcett & A.B. Rendle, Fl. Jamaica 4: 242 (1920).
Annual herb, erect with many ascending branches, 10–50 cm high; indumentum hispidulous on stems and/or minutely glandular hairy. Leaves narrowly elliptic, 2–5 mm long, 6–20 mm wide; apex acute, mucro ~0.3 mm long; more or less sessile. Inflorescence terminal or leaf-opposed, a raceme, up to 60 mm long, exceeding the leaves. Pedicel <1 mm long. Alae position rarely perpendicular, mostly deflexed to reflexed in fruit; white with pink flush to purple, elliptic, 0.7–2.0 mm long, ~0.5–0.75 mm wide; glabrous. Corolla white to rose-pink to purple; floral appendages bifid to much divided; upper petal claw <0.25 mm wide. Stamens terminate above stigma, nearly connate to apex, monadelphous; staminal flap not really visible, <0.25 mm wide. Style more or less straight, becoming hooked at apex, dorso-ventrally flattened, with tuft of hair in well developed pocket above globular stigma. Capsule rarely perpendicular, mostly deflexed to reflexed, more or less aligned with alae or slightly displaced. Capsule lacking wing, obovate to elliptic oblong, 2–4 mm long, 1–3 mm wide; capsule lobe apices rounded to obtuse and separated. Seed oblong, ~1.25 mm long, ~0.50 mm wide; indumentum of sparse fine white hairs. Aril head reduced, ~0.6 mm long, white, with short hairs. Aril appendages 2, linear to oblong, ~0.5 mm long.
Distribution
Australia (NSW, QLD). A native of tropical America, naturalised as a weed in Africa, South-east Asia, Malesia, Australasia and the Pacific. In Australia, found on the eastern coast in northern NSW and south-eastern QLD and in the Wet Tropics of northern QLD (Fig. 9B).
Habitat
In Australia, grows along roadsides and at disturbed sites on a variety of soils.
Notes
This species is not considered problematic and was not revised in the present treatment because only a small number of specimens were seen. The above description has been augmented where necessary from Flora of NSW (Murray 1992), Monro (2003) and van der Meijden (1988). For a full list of synonymy, see van der Meijden (1988). Type details as reported by Monro (2003) are that of Fawcett and Rendle (1920) who stated ‘The type, a specimen from Browne, is in Herb. Linn.’. It is considered a legitimate lectotype here because it is reported as such by the Linnaean Typification Project.
Specimens examined (5)
QUEENSLAND: Windsor Tableland, N of Mt Carbine, 07.vi.1992, A.R. Bean 4560 (BRI, DNA); 8 km SSW of Atherton, 19.iv.1989, J.R. Clarkson 7912 & R.J.F. Henderson (BRI, DNA, L, MBA, QRS); Mason’s property, behind Cape Tribulation shop, 29.iv.2003, L. Hucks LAH085 (BRI, CANB, DNA, MEL); along road to Mission Beach, at Carmoo Ck, 31.iii.2007, R.A. Kerrigan 1191 (BRI, DNA); Goolagan Ck and Henrietta Ck junction, Wooroonooran NP, 17.x.2003, K.R. McDonald KRM1611 (DNA).
27. Polygala parviloba R.A.Kerrigan, sp. nov.
Polygalae succulentae similis, a qua semine ovoideo, crista florali integra lobis parvis et stylo hirsuto differt. Ab congeneribus in Australia semine ovoideo, crista florali integra lobis parvis et stylo hirsuto distinguenda.
Type: Cox Peninsula, 25.iii.2003, I.D. Cowie 9691 & R.K. Harwood; holo: DNA 157571!; iso: BRI, CANB, K, MEL, PERTH.
Polygala chinensis subvar. linearifolia Domin, Biblioth. Bot. 89: 302 (1927); Type: Goulburn Is., A. Cunningham IV. 1818 No. 258; syn- BM (photograph DNA!), K (photograph DNA!), MEL 2057353!; R. Brown Iter Australiense 1802–05 No. 4973 (n.v.); Port Darwin, F. Schultz No. 786; syn- K (photograph DNA!); Type citation: halbnackte Stellen in den Savannenwäldern bei Chillagoe in Gesellschaft zahlreicher Annuellen (Domin II. 1910); Type details: locis subnudis xerodrymiis prope opp. Chillagoe, K. Domin 5692; syn- PR! (photograph BRI!).
[Polygala triflora [NT material] auct. non L. (1753), I.D. Cowie & D.A. Albrecht (eds), Checklist N. Terr. Vasc. Pl. Sp. (2004).]
[Polygala arvensis auct. non Willd. (1802), Benth. Fl. Austral. 1 140 (1863) p.p. as to Goulburn Is., A. Cunningham MEL 2057353, BM, K; A.J. Ewart & O.B. Davies, Fl. N. Terr. 160 (1917); I.D. Cowie & D.A. Albrecht (eds), Checklist N. Terr. Vasc. Pl. Sp. (2004).]
Annual herb, erect with ascending and spreading branches up to 30 cm high; indumentum of curved and straight hairs. Occasionally leaf margins and alae tinged red. Leaves linear to lanceolate, 3–46 mm long, 1–9 mm wide; apex acute, obtuse or rounded, mucro 0.1–0.4 mm long; petiole up to 1 mm long. Inflorescence supra-axillary or axillary, sometimes a solitary flower usually a raceme, up to 32 mm long, not exceeding the leaves. Pedicel up to 2 mm long. Alae position antrorse in fruit; herbaceous, ovate, often slightly dimidiate, apex often with pink mucro, 4.6–6.9 mm long, 1.6–3.8 mm wide; with curved and straight hairs. Corolla purple; floral appendages crenate; upper petal claw 1 mm wide; keel petal with lateral pocket development. Stamens terminate at stigma, monadelphous; staminal flap ≤0.5 mm wide. Style straight or curved, terete in middle becoming slightly dorso-ventrally flattened towards apex and at base, with hispid hairs just below globular stigma. Capsule antrorse rarely perpendicular, more or less aligned with alae. Capsule lacking wing, asymmetrical, widely ovoid to rhomboidal, 3.0–3.6 mm long, 2.2–3.2 mm wide; capsule lobe apices rounded and separated. Seed ovoid, 2.65–3.20 mm long, 1.0–1.8 mm wide; indumentum of white or ferruginous hairs. Aril head reduced, 0.35–0.400 mm long, white and brown or black where reduced to seed testa, with short erect hairs. Aril appendages 3, linear, 0.75–1.25 mm long (Figs 2C, 3E).
Distribution
Australia (NT, QLD, WA). This species is very widely distributed across QLD, NT and WA north of ~18°S (Fig. 9C).
Habitat
Found in a variety of habitats including eucalypt woodlands, Acacia shrublands, Melaleuca shrublands, drainage depressions and seepage areas.
Notes
The level of division of appendages and inflorescence density are quite variable across this species; however, the ascending alae and capsules, and the pink acute apex of the alae, make it readily identifiable in the field.
Etymology
Latin parvus- (small) and -lobus (lobes), in reference to the shallowly lobed floral appendages.
Selected specimens (149 examined)
28. Polygala pendulina R.A.Kerrigan, sp. nov.
A P. galeocephalae capsulis pendulinis et foliis angustatioribus, a P. praecox capsulis non alatis, a P. gabrielae capsulis pendulinis et foliis angustatioribus, et semine parvo differt. Ab congeneribus in Australia capsulis pendulinis, inflorescentiis congestis, stylo uncato, et foliis angustatioribus distinguenda.
Type: Cape York Peninsula, Bromley Station, 6.9 km along track from Bromley Homestead and Wattle Hill, 24.v.2004, R.A. Kerrigan 834 & R.K. Harwood; holo: DNA 162317!; iso: BRI, CANB, K, PERTH.
Annual herb, erect with ascending branches or with spreading to decumbent branches, up to 45 cm high; indumentum of curved and straight hairs or very rarely with curved hairs only. Leaves linear, oblanceolate, 7–35 mm long, 1–3 mm wide; apex acuminate or acute; mucro ~0.3 mm long; sessile. Inflorescence supra-axillary, a raceme up to 125 mm long, usually not exceeding the leaves, sometimes longer. Pedicel up to 4 mm long. Alae position deflexed to reflexed in fruit; herbaceous, ovate, elliptic, dimidiate to falcate, 5.0–7.8 mm long, 1.4–3.3 mm wide; glabrous or with scattered minute curved hairs or a few marginal cilia or glabrous. Corolla purple; floral appendages fimbriate; upper petal claw 0.5–0.6 mm wide. Stamens terminate at stigma, monadelphous; staminal flap <0.5 mm wide. Style hooked (horseshoe-shaped at apex), more or less terete becoming dorso-ventrally flattened below hook and twisting in apical half, with flat stigma flat along inner surface of hook. Capsule deflexed to reflexed, more or less aligned with alae. Capsule lacking wing, if present wing only up to 0.25 mm, more or less equal throughout, more or less symmetrical, obovate 3.0–5.5 mm long, 2.00–2.75 mm wide; capsule lobe apices rounded and separated. Seed ovoid-oblong, 2.50–2.85 mm long, ~1 mm wide; indumentum of fine white hairs. Aril head hooked to helmet-shaped, 0.5–0.6 mm long, white or white and brown, with hairs. Aril appendages 3, linear, ~0.5 mm long.
Distribution
Australia (QLD). This species is known only from Cape York Peninsula, QLD, currently recorded between Strathmore and Bromley Station (Fig. 9D).
Habitat
Often found in sandy soil in Melaleuca shrublands or eucalypt woodlands.
Notes
This species is most similar to P. gabrielae, P. macrobotrya, P. scorpioides and P. stenoclada. It is distinguished from all four species by the mostly smaller seed and the very congested infructescences and pendulous capsules. P. gabrielae, P. macrobotrya and P. scorpioides tend to have wider leaves, and P. stenoclada has an indumentum of curved hairs only.
Etymology
Latin pendulinus (hanging down), referring to the pendulous nature of the capsules.
Selected specimens (9 examined)
29. Polygala persicariifolia DC., Prodromus 1: 326 (1824)
Type: in Nepauliâ. Wallich; holo?: G-DC (photograph!), K, PR; fide A.M. Monro, Systematics of the Australian Polygalaceae and Xanthophyllaceae. Unpublished Thesis, ANU (2003).
Annual herb, erect with ascending branches, up to 50 cm high; indumentum of curved hairs on stems, rarely straight on stems and inflorescence, and scaberulous on leaves, particularly veins, midrib and leaf margins. Leaves strongly discolorous. Leaves narrowly elliptic, lanceolate to ovate, 14–60 mm long, 4–14 mm wide; apex acute, mucro 0–0.3 mm long; petiole 0–1.5 mm long. Inflorescence supra-axillary or axillary in leaf or branch axils, rarely a solitary flower usually a raceme, up to 90 mm long, usually exceeding the leaves. Pedicel up to 5 mm long. Alae perpendicular to reflexed in fruit; petaloid, white to pale green, widely elliptic, 4.0–6.7 mm long, 2.4–5.5 mm wide; glabrous or margin minutely ciliate. Corolla pink to purple; floral appendages fimbriate; upper petal claw 1 mm wide. Stamens terminate at apical stigma, monadelphous; staminal flap ~0.7 mm wide. Style straight at base and bent in apical half, in dried state dorso-ventrally flattened, with 2 globular stigmas, 1 apical and 1 lateral, both apparently receptive. Capsule deflexed to reflexed, more or less aligned with alae. Capsule lacking wing, if present, wing up to 0.4 mm wide at apex, widely ovoid to obovoid, 4.8–5.0 mm long, 4.0–4.3 mm wide; capsule lobe apices rounded and separated. Seed ovoid–oblong, 3.0–3.6 mm long, 1.3–1.7 mm wide; indumentum of fine white or ferruginous hairs. Aril head helmet-shaped, 0.50–0.65 mm long, white, glabrous or with hairs in groove where aril reduced to seed testa. Aril appendages 3, short, more or less triangular, 0.25–0.35 mm long.
Distribution
Australia (QLD), Africa, South-east Asia and Malesia. In Australia, recorded only from a few isolated collections around Mareeba, Mount Molloy and the Endeavour R. (Fig. 9E).
Habitat
In Australia, found in eucalypt woodlands associated with granite, often close to creeks.
Notes
This species is very distinct and is readily distinguished from all other northern species, except P. longifolia, by the distinct helmet-shaped aril and petaloid alae. From P. longifolia it is easily distinguished by the size of the flowers and the axillary versus terminal inflorescences. Style characters from dried material need confirmation with fresh material.
According to Domin (1927), this species was collected from the Endeavour River by A. Cunningham (No. 288) in 1819. I have not seen this specimen; however, considering its relatively broad natural distribution in Australia and throughout Indonesia and Papua New Guinea, I do not see any evidence to suggest it is secondarily introduced as reported by Monro (2003).
Nomenclatural notes
This species was not revised in the present study and is included here to provide a comprehensive Australian account. Typification of this species remains unresolved. Adema (1966) cited the type for this species as Wallich 4185 (L!, G!, CGE n.v.), whereas Monro (2003) cited a specimen from G-DC. The only L specimen catalogued and available to view online, labelled 4185, is also separately labelled as a Hooker JD and Thomson T collection. Annotations on the 4185 label are indecipherable, and although it cannot be excluded as a Wallich specimen, there is no Adema determinavit slip indicating its type status. The specimen cited by Monro (2003) could refer to one of two specimens at G-DC, one with an annotated typed label, ‘in Napaulia, Mr Wallich’, matching de Candolle’s citation, and the other with a handwritten label annotated, ‘Mr Wallich’, but with no locality details. Again, there are no determinavit slips from Adema, and neither may be the G specimen that Adema (1966) cited. Also, the ABLO confirmed that a Wallich specimen matching the type details, ‘in Napulia’, is housed in Wallich’s collection at K. Apparently, a full set of his collections is deposited in the Central National Herbarium of the Botanical Survey of India in Calcutta (CAL). I agree with Monro (2003) that a G-DC specimen is a more likely candidate for the holotype because of access for the author, but one of the two specimens there requires lectotypification, and the status of potential isolectotypes needs verification.
Selected specimens (9 examined)
30. Polygala petrophila R.A.Kerrigan, sp. nov.
Polygalae succulentae similis, a qua semine ovoideo et stylo canaliculato differt. Ab congeneribus in Australia crista florali succulenta distinguenda.
Type: Kimberley, 7.6 km E along Gibb River Road from Beverley Springs turnoff, 12.v.2006, R.A. Kerrigan 1107; holo: DNA 176666!; iso: BRI, CANB, K, PERTH.
Annual herb, erect with ascending branches, up to 30 cm high; indumentum of curved and straight, pubescent to hispidulous hairs. Leaves linear to oblanceolate, elliptic or obovate, 6–55 mm long, 1.5–11.0 mm wide; apex acute to rounded, mucro 0.2–0.6 mm long; petiole 0–1 mm long. Inflorescence supra-axillary or rarely axillary, a raceme, sometimes with a solitary flower on internode preceding raceme, up to 40 mm long, usually not exceeding the leaves, sometimes longer. Pedicel up to 2.5 mm long. Alae position antrorse to perpendicular in fruit; herbaceous, lanceolate, falcate, 5.0–7.5 mm long, 1.3–2.4 mm wide; with curved and straight hairs or straight hairs only. Corolla purple; floral appendages succulent, bifid to 3–4 times forked; upper petal claw 1–1.5 mm wide. Stamens terminate at stigma, monadelphous; staminal flap ~0.5 mm wide. Style straight to slightly curved, dorso-ventrally flattened, often hairy at base and always prominently grooved above globular stigma. Capsule perpendicular to slightly deflexed, more or less aligned with alae. Capsule lacking wing, orbicular or widely ovate, 3.0–4.0 mm long, 3.0–3.8 mm wide; capsule lobe apices obtuse to rounded and separated. Seed ovoid, 2.7–3.3 mm long, 1.6–1.9 mm wide; indumentum of fine white or ferruginous hairs. Aril head reduced, 0.3–0.6 mm long, brown or black where reduced to seed testa, or brown and white, with short hairs. Aril appendages 3, linear, 1–2 mm long (Figs 3I, 6D).
There are two varieties, which can be distinguished as follows:
|
1 Leaves 6–11 mm wide, elliptic, obovate or oblanceolate, linear |
Distribution
Australia (NT, WA). Occurs between 15°S and 18°S from the Kimberley region of WA to Roper River in the NT (Fig. 9F).
Habitat
Found mostly in eucalypt woodlands among rubble on rocky sandstone scree slopes and rises.
Notes
Polygala petrophila is quite variable in habit and hairiness. P. petrophila var. angustifolia is easily distinguished from P. succulenta by its ovoid seed, but can be difficult to distinguish when mature seeds are not available. A flowering specimen from around Roper River, J.A. Risler 2146 (DNA), is easily confused with P. succulenta, but is recognised as P. petrophila by the presence of a groove in the style above the stigma.
Etymology
Greek petra- (rock) and -philus (loving), rock lover because this species is always found among sandstone/laterite rubble on scree slopes.
Polygala petrophilavar. angustifolia R.A.Kerrigan, var. nov.
A var. petrophila foliis angustatioribus differt.
Type: Jalboi R. catchment, 16 km E by N of Crown Hill, 18.iii.2003, I.D. Cowie 9674; holo: DNA 160389!; iso: BRI, CANB, K, MEL, PERTH.
Plants usually erect up to 30 cm high. Leaves 6–55 mm long, 1.5–7.0 mm wide, narrowly elliptic, lanceolate to linear. Inflorescence up to 40 mm long.
Distribution
Australia (NT). From the Roper River to the Victoria River (Fig. 9F).
Notes
Intermediates between the two varieties, R.A. Kerrigan 916 (DNA, PERTH) and R.A. Kerrigan 918 (DNA), are placed with var. angustifolia because of their leaf dimensions. However, they share the stature and shorter inflorescence of var. petrophila.
Etymology
Latin angustus- (narrow) and -folium (leaf), in reference to the fact that this variety has narrow leaves.
Selected specimens (12 examined)
P. petrophila R.A.Kerrigan var. petrophila
Plants erect usually up to 9(–21) cm high. Leaves 22–30 mm long, 6–11 mm wide, elliptic, obovate or oblanceolate, linear. Inflorescence up to 35 mm long.
Distribution
Australia (NT, WA). Distributed around 18°S in NT and WA from Victoria River to Windjana National Park (Fig. 9F).
Notes
The variety, petrophila, tends to have a more congested and compact inflorescence than var. angustifolia, although both would be considered relatively congested. The most easterly specimen of this variety, K.G. Brennan 5920 & K. Metcalfe (DNA), is approaching the leaf dimension of var. angustifolia, but has the stature and compressed inflorescence of var. petrophila.
Selected specimens (9 examined)
31. Polygala polifolia C.Presl, Reliquiae Haenkeanae. ii. 101. (1831–1835)
Type: ‘Habitat in insula Luzon.’; holo: PR 375755 (photograph DNA!).
[Polygala chinensis [Australian material] auct. non L. (1753), Adema, Blumea 14 (2) (1966), 269.]
Annual herb, erect with decumbent, ascending or spreading branches, up to 20 cm high; indumentum of curved hairs. Leaves oblanceolate, narrowly elliptic or narrowly oblong, 4–22 mm long, 2–8 mm wide; apex obtuse or rounded, rarely some leaves acute, mucro 0.1–0.3 mm long; petiole 0–1 mm long. Inflorescence supra-axillary, a raceme, up to 2 mm long, not exceeding the leaves. Pedicel up to 0.5 mm long. Alae position perpendicular or deflexed in fruit; herbaceous, ovate to lanceolate, falcate or dimidiate, 2.40–2.95 mm long, 1.0–1.3 mm wide; with curved hairs or glabrescent. Corolla purple, white or yellow (from dried material only); floral appendages fimbriate; upper petal claw ~0.25 mm wide. Stamens terminate at stigma, triadelphous (fused as 3, 2, 3); staminal flap ~0.25 mm wide. Style curved in apical half, truncate at apex with sterile tip distal to more or less globular stigma, style with narrow groove at apex; in dried state dorso-ventrally flattened. Capsule deflexed, becoming reflexed, more or less aligned with alae or apex of alae diverging at ~45° from capsule. Capsule lacking wing, more or less symmetrical, widely ovoid to orbicular, 2.1–2.5 mm long, 1.9–2.2 mm wide; capsule lobe apices rounded and separated. Seed ovoid to elliptic, 1.90–2.10 mm long, 0.90–1.15 mm wide; indumentum of fine white. Aril head hooked to round, 0.2–0.4 mm long, white, with short erect hairs. Aril appendages 3, linear, 0.25–0.6 mm long.
Distribution
A very widespread species distributed from Pakistan to China and from the Philippines to Australia (QLD). In Australia, it is known from only a few collections from Rockingham Bay near Cardwell, Uningan Nature Reserve at Weipa, and a dubious collection from central QLD. Attempts to relocate the populations near Cardwell and Weipa in March 2007 failed. A BRI collection from Carmoo Creek (N.B. Byrnes 209), on QLD’s eastern coastline, is not mapped or listed. It seems it may be mislabelled because I found only P. paniculata at the precise location (Fig. 9G).
Habitat
With so few collections, it is difficult to determine the habitat of this species in Australia. Van der Meijden (1988) indicated that this species occupies roadsides, grasslands and waste places. The few collections of this species may reflect the fact that much of the accessible coastal lowland savanna is under cultivation. This species is considered Data Deficient because there has been inadequate targeted survey of this species, and further survey should be given priority.
Notes
This species is very similar to P. glaucoides, but is easily distinguished from it by the fusion of staminal filaments into three bundles (3, 2, 3), rather than one bundle of eight, and by having a style with an apical point rather than a hook.
Nomenclatural notes
Although van der Meijden (1988) discussed difficulties with the synonymy of this species, the application of the name P. polifolia to these Australian specimens is relatively straightforward. Flower colour for this species needs to be treated with caution; Forster 6484 (BRI, DNA) records flower colour as white, and Mueller records flower colour as yellow and blue, and yet van der Meijden (1988) described the species as having light to deep violet flowers. It has been my experience in the field that dark blue flowers fade to white with age. Both the Mueller and Forster collections are of mature fruiting specimens, and it is likely that any flowers present were faded at the time of collection.
Selected specimens (4 examined)
32. Polygala praecox R.A.Kerrigan, sp. nov.
Polygalae pendulinae affinis, a qua capsulis alatis differt; a speciebus aliis alatis capsulis asymmetricis, et sepalis lateralibus angustatioribus distinguitur.
Type: Kalumburu road, 85.6 km by road N from junction with Gibb R. and Ellenbrae Rd, 29.iv.1985, T.E.H. Aplin 715 & R.J. Cranfield; holo: PERTH 1854550!
Annual herb, erect with ascending branches, up to 55 cm high; indumentum of curved hairs only. Leaves linear, narrowly elliptic, 10–55 mm long, 1.5–3.5 mm wide; apex acute, mucro up to 0.2 mm long; sessile. Inflorescence supra-axillary or axillary, a raceme, up to 50 mm long, not exceeding the leaves. Pedicel up to ~3 mm long. Alae position perpendicular to deflexed in fruit; herbaceous, lanceolate, narrowly elliptic, slightly dimidiate to falcate, 7–9 mm long, ~2.4–3.0 mm wide; with minute more or less appressed to curved hairs or glabrescent, sometimes with straight hairs along margin. Corolla purple; floral appendages reduced, more or less entire. Stamens terminate below stigma, monadelphous. Style shallowly hooked with small beak or protrusion on outer edge of bend, with somewhat flat stigma. Capsule perpendicular to deflexed, more or less aligned with alae. Capsule lacking wing or with uneven wing, but winged capsules always present, wing ~0.5 mm wide on 1 locule, 1–2 mm wide on the other; strongly asymmetrical, obovate or rhomboidal, 5–6 mm long, 2–3 mm wide; capsule lobe apices rounded and strongly unequal. Seed oblong, 2.0–3.5 mm long, ~1.5 mm wide; indumentum of fine white hairs. Aril shape domed or round, ~0.6 mm long, white, with hairs. Aril appendages 3, linear to narrowly triangular, ~0.75 mm long.
Distribution
Australia (WA). This species is known only from the Kimberley region of WA, along the Kalumburu Road (Fig. 9H). Two unvouchered specimens listed below were intact and rooted in the ground at the time of survey, but had completely senesced and were too brittle to collect.
Habitat
Apparently restricted to a specific habitat of Eucalyptus tectifica F.Muell. and Corymbia grandifolia (R.Br ex Benth.) K.D.Hill & L.A.S.Johnson woodlands growing on red clay loam, with Heteropogon Pers. and Sehima Forssk., possibly associated with basalt.
Notes
This species is similar to P. pendulina but can be distinguished by the asymmetrical capsule and the presence of capsule wings. Capsules on an individual are irregularly winged, i.e. capsules sometimes lack wings or have wings only on one locule. Flower characteristics are described from one depauperate specimen, and further material is required. Staminal flap, upper petal claw width and style shape in cross-section are not described here as there was not enough flowering material available.
Etymology
Latin praecox (developing early), referring to the senescence of this species well before other species in the area.
Selected specimens (5 examined)
33. Polygala pterocarpa R.A.Kerrigan, sp. nov.
Polygalae crassitestae simillimus a qua testa non incrassata differt. Ab congeneribus in Australia capsulis alatis symmetricis conspicuis, et sepalis lateralibus vel aequalis vel angustatioribus quam capsulis distinguenda.
Type: Gregory National Park, 3.8 km from Bullita road turnoff, on Limestone Gorge Rd, 11.iv.2003, R.A. Kerrigan 621; holo: DNA 169062!; iso: BRI, CANB, K, PERTH.
[Polygala rhinanthoides auct. non Sol. ex Benth. (1863), A.J. Ewart & O.B. Davies, Fl. of the N. Terr. 160 (1917) p.p. as to NT specimens; J.R. Wheeler, Fl. Kimberley 643 (1992); I.D. Cowie & D.A. Albrecht (eds), Checklist N. Terr. Vasc. Pl. Sp. (2004) p.p. as to western NT specimens.]
Annual herb, erect with ascending or spreading branches, up to 90 cm high; indumentum of curved and straight hairs. Leaves linear, narrowly elliptic to oblanceolate, elliptic or obovate, 3–77 mm long, 1–11 mm wide; apex rounded, truncate, retuse or emarginate, mucro 0–0.3 mm long; petiole 0–2.5 mm long. Inflorescence axillary or supra-axillary, rarely a solitary flower usually a raceme, up to 53 mm long, not exceeding the leaves. Pedicel up to 3 mm long. Alae position perpendicular, deflexed to occasionally reflexed in fruit; herbaceous, very widely to widely ovate and elliptic, 3.6–7.6 mm long, 2.1–6.2 mm wide; margins ciliate, outer surface with scattered minute curved hairs and straight hairs, or straight hairs only or curved hairs only. Corolla purple; floral appendages fimbriate; upper petal claw 0.6–1 mm wide. Stamens terminate at stigma, monadelphous; staminal flap 1 mm wide. Style hooked (horseshoe-shaped at apex), style often with beak on outer edge of bend visible in dried material, more or less terete, becoming dorso-ventrally flattened below hook and twisting in apical half, with flat stigma along inner surface of hook. Capsule perpendicular, deflexed to occasionally reflexed, more or less aligned with alae. Capsule with wing, 1.0–3.9 mm wide, more or less equal throughout, symmetrical, widely elliptic, very widely ovate, orbicular to depressed orbicular, 3.5–5.5 mm long, 2.2–4.1 mm wide; capsule lobe apices rounded and separated. Seed ovoid-oblong, 3.06–4.70 mm long, 1.31–2.10 mm wide; indumentum of fine white hairs. Aril head round to hooked, 0.4–0.8 mm long, white and brown or black, with short hairs. Aril appendages 3, linear, 1.1–3.3 mm long (Fig. 6G).
Distribution
Australia (NT, QLD, WA). In the NT, it occurs as far north as Katherine, south to the Tanami Desert and east to the Barkly Tablelands, and occurs as far east as Chillagoe in QLD and west to the Napier Range in WA (Fig. 9I).
Habitat
Found mostly in open woodland habitats with a grassy understory, typically with Eucalyptus patellaris F.Muell., E. pruinosa, Corymbia terminalis or Acacia spp., occasionally, in vine thickets or bulwaddy thickets (Macropteranthes kekwickii) over Aristida L., Astrebla or Triodia spp. Found growing in a variety of substrates, including limestone, sandy soil and clay loam.
Notes
This species is very similar to P. crassitesta and P. rhinanthoides. It is distinguished from the former by the absence of the thickened testa and from the latter by the size of the winged capsule relative to the alae.
Etymology
Greek pterus- (winged), -carpo (fruit), in reference to the prominently winged capsules.
Selected specimens (73 examined)
34. Polygala pycnantha R.A.Kerrigan, nom. nov.
Polygala pycnophylla Domin, Biblioth. Bot. 89: 304 (1927), nom. illeg.
Type citation: Queensland: Savannenwälder bei Pentland (Domin III. 1910); Type detail: in xerodrymiis prope opp. Pentland, K. Domin 5695; holo: PR 528255! (photograph CANB, DNA!).
Annual herb, erect with ascending or spreading branches, up to 15 cm high; indumentum of short curved and long straight hairs. Leaves linear, narrowly elliptic, oblanceolate or occasionally with some obovate leaves, sometimes with linear and obovate leaves on the same plant, 5–46 mm long, 2–10 mm wide; apex acute or rounded, mucro ~0.2 mm long, petiole 0–1 mm long. Inflorescence supra-axillary or axillary, a raceme, up to 30 mm long, not exceeding the leaves. Pedicel up to 2 mm long. Alae position mostly antrorse and perpendicular in fruit; herbaceous, ovate, usually falcate to dimidiate, 5–8 mm long, 2.5–3.0 mm wide; densely hairy, with mostly long straight hairs and scattered minute curved hairs. Corolla poorly known, yellow in bud with purple tinges in material seen; floral appendages spathulate; upper petal claw ~0.5 mm wide. Stamens terminate at stigma, monadelphous; staminal flap ~0.25 mm wide. Style hooked (horseshoe-shaped at apex), in dried state more or less terete but dorso-ventrally flattened at base, with flat stigma along inner surface of hook. Capsule antrorse and perpendicular, rarely reflexed, more or less aligned with alae. Capsule lacking wing, oblong, 4–5 mm long, 2.8–3.6 mm wide; capsule lobe apices rounded and separated. Seed ovoid-oblong, 3.70–4.25 mm long, 1.35–1.50 mm wide; indumentum of fine white sometimes ferruginous hairs. Aril head helmet-shaped, 0.70–0.85 mm long, white or white and brown, with fine short hairs in central groove. Aril appendages 3, oblong, 0.8–1 mm long.
Distribution
Australia (QLD). This species is known only from a small area around Charters Towers and Pentland and a few collections near the Gulf of Carpentaria east of Karumba (Fig. 9J).
Habitat
Usually found in a variety of eucalypt woodlands including Eucalyptus crebra and Corymbia leichhardtii on sandy soil and loam, occasionally in seasonally inundated situations with Melaleuca spp.
Notes
This species is very similar to P. difficilis. It is distinguished by the densely hairy alae with long spreading hairs, the restriction of hairs to the groove of the aril head, rather than across the whole aril head, and the absence of strigose hairs on the back of the keel petal.
Nomenclatural notes
Polygala pycnophylla Domin is an illegitimate later homonym of P. pycnophylla Brandegee (1917). I have used Domin’s type for pycnophylla and the epithet pycnantha because the flowers are congested in the inflorescence. Although a collection number is not cited in the protologue for P. pycnophylla Domin, there seems to be little doubt that K. Domin 5695 is the holotype. There is a close match to the type citation in locality details, the specimen is part of Domin’s Herbarium and was clearly seen by him, it is a good fit to Domin’s original description and the ‘n. sp.’ (new species?) annotation on the label provides an additional clue. In line with recommendation 9A.4 and Art. 37.3 of the International Code of Botanical Nomenclature (McNeill et al. 2006), the specimen is considered the type.
Etymology
Greek pycno- (congested) and -anthos (flowers), referring to the congested flowers of this species.
Selected specimens (8 examined)
35. Polygala rhinanthoides Sol. ex. Benth., Fl. Austral. 1 : 140 (1863)
Lectotype (here designated): Endeavour R., R. Brown; lecto: BM 566299! (photograph DNA!); isolectotype: BM 566300!; excluded syntypes: Upper Victoria R., F. Muell.; syn- MEL 226369 a! and b!; Upper Victoria R., F. Muell.; syn- K 00279781!
Annual herb, erect rarely with short spreading to ascending lateral branches, up to 60 cm high; indumentum of curved and straight hairs, sometimes with curved hairs only. Stems occasionally tinged red. Leaves linear to lanceolate, 6–83 mm long, 1.5–7.0 mm wide; apex acute, obtuse or rounded, mucro 0.3–0.4 mm long; petiole 0–2 mm long. Inflorescence supra-axillary, a raceme, up to 110 mm long, exceeding the leaves or shorter than the leaves. Pedicel up to 5 mm long. Alae position sometimes perpendicular, mostly deflexed to reflexed in fruit; thinly herbaceous, ovate to widely ovate, 4.8–7.7 mm long, 2.5–7.1 mm wide; usually with curved hairs, sometimes straight hairs present especially on the margin or glabrescent. Corolla purple; floral appendages fimbriate; upper petal claw 1.2–1.5 mm wide. Stamens terminate at stigma, monadelphous; staminal flap 1.5 mm wide. Style hooked (horseshoe-shaped at apex), with beak on curve of bend, terete in middle and dorso-ventrally flattened at base and below hook, twisting in apical half, with flat stigma along inner surface of hook. Capsule sometimes perpendicular, mostly deflexed to reflexed, more or less aligned with alae. Capsule with wing, 0.7–3.0 mm wide, more or less equal throughout, widely ovoid to orbicular, 3.3–5.5 mm long, 3.0–3.5 mm wide; capsule lobe apices rounded and separated. Seed ovoid-oblong 2.7–4.0 mm long, 1.3–1.8 mm wide; indumentum of fine white or ferruginous hairs. Aril head round, 0.5–0.75 mm long, white or white and brown, with short erect hairs. Aril appendages 3, narrowly oblong, 1.0–2.3 mm long.
Distribution
Australia (QLD), Papua New Guinea. This species is known, in Australia, from Cooktown on Cape York Peninsula to Thursday Island in the Torres Strait (Fig. 9K).
Habitat
Found in eucalypt woodlands in sandy soils or loam and in seasonally inundated situations.
Notes
This species is very similar to P. pterocarpa. It is distinguished by habit, inflorescence and alae size. P. rhinanthoides is usually a single-stemmed erect herb, as opposed to the more branching and spreading habit in P. pterocarpa. P. rhinanthoides generally has a longer inflorescence, which is less congested, and has a finer-textured alae. The capsule and capsule wing in P. rhinanthoides tend to be covered by the alae, whereas the capsule is usually prominently exserted past the alae in P. pterocarpa.
Nomenclatural notes
There are four sheets that could be considered syntypes of P. rhinanthoides, two from the ‘Endeavour R., R. Brown’ housed at BM (BM 566299 and BM566300), and two from ‘Upper Victoria R., F. Muell.’ housed at MEL (MEL226369 and K (K00279781). See note under P. exsquarrosa regarding Brown specimens from Endeavour River, one of which is here chosen as the lectotype. The two collections from the Upper Victoria River are excluded because they are a different species, and K00279781 appears to be one part of a mixed gathering. The Melbourne material bears Bentham’s initial (B) and is separated into two subcollections, ‘a’ and ‘b’. Specimen ‘a’ can be identified as P. pterocarpa, but Specimen ‘b’ is too incomplete to allow identification, although the shrivelled area below the aril may have developed into the thickened collar seen in P. crassitesta.
Selected specimens (25 examined)
36. Polygala rhynchocarpa R.A.Kerrigan, sp. nov.
Polygalae canaliculatae affinis, a qua lobis capsulae apicibus acutis differt. Ab congeneribus in Australia stylo curvo et torto, apice styli truncato, et lobis capsulae apicibus acutis distinguenda.
Type: Kimberley, George’s Jump-up, 125.3 km E of Kalumburu turnoff on Gibb River Rd, 15.v.2006. R.A. Kerrigan 1129; holo: DNA 176688!; iso: BRI, CANB, PERTH.
Annual herb, erect rarely with ascending branches, up to 55 cm high; indumentum of curved hairs. Leaves sparse and caducous, linear, 22–75 mm long, 0.75–1.50 mm wide; apex acute, mucro 0–0.2 mm long; more or less sessile. Inflorescence supra-axillary or axillary, a raceme, up to 300 mm long, exceeding the leaves. Pedicel up to 5 mm long. Alae position perpendicular to deflexed, occasionally reflexed or ascending in fruit; herbaceous; lanceolate, narrowly elliptic or oblanceolate; dimidiate to falcate, 4.9–7.0 mm long, 1–2 mm wide; with minute curved hairs or glabrescent. Corolla purple; floral appendages bifid, each appendage shortly divided; upper petal claw 1.25 mm wide. Stamens terminate at stigma, monadelphous; staminal flap 1 mm wide. Style slightly curved and twisted, more or less terete, truncated into a narrow hammer head above stigma, with somewhat flattened stigma. Capsule reflexed or perpendicular, more or less aligned with alae. Capsule lacking wing, symmetrical, obovate or squarish, emarginate with acute apex to capsule lobes, 5.8–7.0 mm long, 3.1–3.4 mm wide; capsule lobe apices acute or sub-acute and separated. Seed oblong, 4.0–4.8 mm long, ~1.5 mm wide; indumentum of fine white hairs. Aril head an elongated helmet shape, 1.0–1.3 mm long, white, sometimes pale brown, hairy, often obscurely so. Aril appendages 3, more or less linear, 0.5–0.9 mm long (Fig. 3G).
Distribution
Australia (WA). This species is known only from the Kimberley Region in WA, north of and including the Gibb River Road (Fig. 9L).
Habitat
Usually growing in sandy soil often associated with sandstone.
Notes
This species is very similar to P. stenoclada, P. macrobotrya, P. pendulina and P. canaliculata. It is distinguished from the first three by the curved style, rather than hooked, and by the size of the leaves, and from the the fourth species by the absence of the groove on the end of the broad truncated style, and the acute apex of the capsule.
Etymology
Greek rhyncho- (beak) and -carpo (fruit), because of the acute or beaked apex of the lobes of the capsule.
Selected specimens (7 examined)
37. Polygala saccopetala R.A.Kerrigan, sp. nov.
Polygalae scorpioides similis, a qua carina saccatiore indumento et pilis curvis non nisi differt. Ab congeneribus in Australia carina saccata distinguenda.
Type: Bullo River Station, N from Bullo River Homestead, 12.iii.2006, R.A. Kerrigan 1083; holo: DNA 176651!; iso: BRI, CANB, K, L, MEL, PERTH.
Annual herb, erect with ascending, spreading or decumbent branches, up to 40 cm high; indumentum of curved hairs, leaves often glabrescent. Leaves linear, narrowly elliptic to oblanceolate, elliptic to ovate, rarely obovate, 2–53 mm long, 1.4–12.0 mm wide; apex acute, obtuse, rounded or truncate, rarely retuse or emarginate, mucro 0.2–0.3 mm long; petiole 0–1.5 mm long. Inflorescence supra-axillary, a raceme, up to 150 mm long, exceeding the leaves or sometimes shorter than the leaves and often scorpioid. Pedicel up to 3 mm long. Alae position perpendicular to deflexed in fruit; herbaceous, ovate, elliptic or obovate, slightly dimidiate, 3.7–5.7 mm long, 2.0–2.8 mm wide; with curved hairs or glabrous. Corolla purple; keel petal with well developed pockets on both lower flaps; floral appendages fimbriate; upper petal claw ~0.5 mm wide. Stamens terminate at stigma, monadelphous; staminal flap ~0.5 mm wide. Style very hooked (horseshoe-shaped at apex), more or less terete, thickened in middle, becoming dorso-ventrally flattened below hook and twisting in apical half, with flat stigma along inner surface of hook. Capsule occasionally perpendicular, mostly deflexed to reflexed, more or less aligned with alae. Capsule lacking wing, more or less symmetrical, widely ovoid, 4.2–5.0 mm long, 3.3–4.3 mm wide; capsule lobe apices rounded and separated. Seed ovoid–oblong, 3.5–4.4 mm long, 1.5–1.8 mm wide; indumentum of fine white hairs. Aril head hooked or helmet-shaped, 0.6–0.9 mm long, white or brown and white, with minute short erect hairs, inconspicuous even under magnification. Aril appendages 3, shortly linear to narrowly oblong, 0.8–1.5 mm long (Figs 4C, 5C).
Distribution
Australia (NT, WA). This species is found west of the Victoria River in the NT and extends to King Leopold National Park in WA (Fig. 10A).
Habitat
Found in woodlands growing amongst sandstone rubble.
Notes
This species is very similar to P. scorpioides and is distinguished by the absence of straight hairs and by having much more developed pockets on the keel petal.
Etymology
Latin saccatus- (pouched) and -petalum (petal), referring to the well developed pockets on the keel petal.
Selected specimens (15 examined)
38. Polygala scorpioides R.A.Kerrigan, sp. nov.
Polygalae saccopetalae affinis, a qua pilis rectis indumentorum et carina parvulis saccata differt. Ab congeneribus in Australia carina saccata distinguenda.
Type: Rookwood Station, near old mango farm on N bank of Walsh R., 29.iii.2007, R.A. Kerrigan 1185; holo: DNA 180131!; iso: BRI, CANB, PERTH.
Annual herb, erect with ascending lateral branches rarely sprawling, up to 30 cm high; indumentum of curved and straight hairs. Stems and leaves occasionally tinged maroon. Leaves linear, narrowly elliptic, elliptic or ovate, 4.5–6.5 mm long, 2.5–9.0 mm wide; apex acute or obtuse, mucro 0.2–0.3 mm long; petiole 0–2 mm long. Inflorescence supra-axillary or axillary, a raceme, up to 160 mm long, not exceeding the leaves and scorpioid. Pedicel up to 3.5 mm long (4.5 mm in capsule). Alae position antrorse to perpendicular in fruit; herbaceous, lanceolate or narrowly elliptic to elliptic, usually dimidiate, 3.6–6.4 mm long, 1.2–2.6 mm wide; often with straight hairs on margin cilia and curved hairs on outer surface. Corolla purple; keel petal with slightly developed pocket; floral appendages fimbriate; upper petal claw 0.6–1.0 mm wide. Stamens terminate at stigma, monadelphous; staminal flap ~0.6 mm wide. Style hooked (horseshoe-shaped at apex), terete at base, thickened in middle and becoming dorso-ventrally flattened below hook, twisting in apical half, with flat stigma along inner surface of hook. Capsule antrorse to perpendicular, rarely becoming reflexed, more or less aligned with alae. Capsule lacking wing, widely elliptic to widely oblong, 4.4–5.5 mm long, 2.5–3.5 mm wide; capsule lobe apices rounded and separated. Seed ovoid–oblong, 3.25–4.20 mm long, 1.1–1.8 mm wide; indumentum of fine white becoming ferruginous hairs. Aril head hooked to helmet-shaped, 0.5–1.0 mm long, white or white and brown, with fine short hairs. Aril appendages 3, oblong, 0.5–1.0 mm long (Fig. 3C).
Distribution
Australia (QLD). This species is found in Cape York Peninsula from Somerset in the north to the Lamb Range and the Walsh River near Chillagoe in the south (Fig. 10B).
Habitat
Usually found in a variety of eucalypt woodlands on sandy soil and loam.
Notes
This species is very similar to P. saccopetala, P. gabrielae (see notes under that species) and P. triflora. From P. saccopetala, it is immediately distinguished by an indumentum including straight hairs and by much reduced pocket development on the keel. P. triflora is generally distinguished by a more thickened taproot, an often multi-stemmed, sometimes woody habit and by the absence of pocket development on the keel petal. P. scorpioides tends to have bigger seeds and bigger alae than does P. triflora. P. scorpioides is a variable species, varying from slender, low spreading to ascending, few-flowered specimens, e.g. RW Johnson 5078 (BRI, DNA, L), to more robust, erect specimens with scorpioid inflorescences, e.g. RA Kerrigan 1185 (BRI, DNA, PERTH). Although these extremes are easily distinguished, many specimens share characters from both extremes, e.g. RA Kerrigan 1174 (BRI, DNA, PERTH) and RA Kerrigan 822 (BRI, DNA), making it difficult to further divide this species taxonomically.
Nomenclatural notes
Specimens in this group have generally been identified in the past as P. linariifolia, following Adema (1969). Adema recognised that specimens attributed to this name in Australia differed from Malesian material in flower colour and size and in shape of seed. The naming and synonymy of this taxon are quite confused and are discussed under P. triflora.
Etymology
Latin scorpioideus, referring to the inflorescence with the main axis coiled like the tail of a scorpion.
Selected specimens (24 examined)
39. Polygala stenoclada Benth., Fl. Austral. 1 : 141 (1863)
Polygala stenoclada var. typica Domin nom. inval., Biblioth. Bot. 89: 303 (1927).
Lectotype (here designated): Upper Victoria R., F. Muell. 5/1856; lecto: MEL 37322!; excluded syntype, Upper Victoria R. F. Muell.; syn- K 279792!
Annual herb, erect with spreading to ascending branches, up to 60 cm high; indumentum of curved hairs only. Leaves linear, 10–70 mm long, 0.5–2.5(4.0) mm wide; apex acute, mucro ~0.2 mm long; more or less sessile. Inflorescence supra-axillary or rarely leaf-opposed, a raceme, up to 200 mm long, exceeding the leaves. Pedicel up to 4.5 mm long. Alae position sometimes perpendicular, mostly deflexed becoming reflexed in fruit; herbaceous, lanceolate, narrowly elliptic and oblanceolate, often dimidiate to falcate, 4.3–6.3 mm long, 1.0–2.6 mm wide; with minute appressed hairs. Corolla purple; floral appendages spathulate; upper petal claw 1 mm wide. Stamens terminate at stigma, monadelphous; staminal flap <0.5 mm wide. Style hooked (horseshoe-shaped at apex), laterally compressed at base, terete and thickened in middle, becoming dorso-ventrally flattened below hook and twisting in apical half, with a flat stigma along inner surface of hook. Capsule sometimes perpendicular, mostly deflexed becoming reflexed, more or less aligned with alae. Capsule lacking wing, somewhat asymmetrical, obovate, 4.7–6.0 mm long, 2.5–3.3 mm wide; capsule lobe apices rounded and separated. Seed oblong, 4.0–5.5 mm long, 1.0–1.8 mm wide; indumentum of fine white hairs. Aril head helmet-shaped, 0.65–1.20 mm long, white, with hairs. Aril appendages 3, linear often shortly so, 0.45–1.00 mm long (Fig. 6E).
Distribution
Australia (NT, QLD). This species is mainly distributed from the Gulf of Carpentaria in QLD to the western coast of the NT (Fig. 10C).
Habitat
Usually growing in sandy soil in savanna, on sandy plains or among sandstone outcrops.
Notes
This species is very similar to P. macrobotrya and P. stenosepala. From P. macrobotrya, it is distinguished by the helmet-shaped aril head and the presence of hairs over most of the aril head, rather than just a central tuft. Compared with P. stenosepala, P. stenoclada has broader ovate to obovate outer sepals, usually with darkened tips, a more erect, taller habit, and only curved hairs. P. stenosepala also has a more candelabra-like habit, lanceolate to subulate outer sepals and mostly curved hairs with a few straight hairs. None of these characters, however, is exclusive, and P. stenoclada is best distinguished from P. stenosepala by the size of the seeds and by the absence of thick hairs below the aril head.
Nomenclatural notes
The specimen chosen as the lectotype is labelled with two different names, apparently in Mueller’s hand, as follows: P. arvensis var. linearis and P. stenoclada ‘typical’. The top left-hand corner of the label, ‘P. arvensis var. linearis’, is folded over and annotated with a ‘B’, accepted as a symbol that it was sighted by Bentham.
The excluded syntype, Upper Victoria R., F. Muell., K279792, has no capsules and as such cannot be unequivocally assigned to this species or P. stenosepala. Both can exhibit similar habits, reach similar lengths and have compatible distributions.
Selected specimens (25 examined)
40. Polygala stenosepala (Benth.) R.A.Kerrigan, comb. et stat. nov.
Polygala stenoclada var. stenosepala Benth. Fl. Austral., 1: 141 (1863).
Type: Carpentaria, Point T and Point S Arnhem Bay [Cape Shield and Point Blane], 26.i.1803, R. Brown s.n.; holo: BM 566304!
[Polygala stenoclada auct. non Benth. (1863), I.D. Cowie & D.A. Albrecht (eds), Checklist N. Terr. Vasc. Pl. Sp. (2004).]
Annual herb, erect often with arching ascending branches, rarely decumbent or prostrate, up to 35 cm high; indumentum of mostly curved and rarely straight hairs. Stems often tinged maroon. Leaves linear, occasionally narrowly elliptic or lanceolate, 2.5–58.0 mm long, 0.5–4.0 mm wide; apex acute or acuminate, mucro ~0.2 mm long; petiole 0–1 mm long. Inflorescence supra-axillary or axillary, a raceme, up to 170 mm long, exceeding the leaves. Pedicel up to 8 mm long. Alae position perpendicular, deflexed and sometimes antrose in fruit; herbaceous, occasionally with red stripe along outer surface, lanceolate, narrowly elliptic, oblanceolate, slightly dimidiate to falcate, 3.6–7.6 mm long, 1.0–2.4 mm wide; with curved hairs, ±ciliate on margins, or glabrescent. Corolla purple; floral appendages fimbriate; upper petal claw 1.2–1.5 mm wide. Stamens terminate at stigma, monadelphous; staminal flap 0.25–0.6 mm wide. Style hooked (horseshoe-shaped at apex), terete from base and dorso-ventrally flattened below hook, with flat stigma along inner surface of hook. Capsule sometimes antrorse, mostly perpendicular or deflexed, more or less aligned with alae. Capsule lacking wing, usually strongly asymmetrical, rhomboid in shape, rarely squarish, 2.5–5.0 mm long, 2.0–4.5 mm wide; capsule lobe apices rounded, separated and unequal. Seed ovoid-oblong, 2.2–3.5 mm long, 1.1–2.0 mm wide; indumentum with two hair types present, fine white hairs throughout and thick clearly hollow hairs spreading to strongly recurved below aril. Aril round to hooked, 0.3–0.8 mm long, white, with hairs, rarely glabrous. Aril appendages 3, linear, 0.3–1.5 mm long.
Distribution
Australia (NT, WA). This species is mainly distributed across the far north of the NT from north-eastern Arnhem Land to the Victoria River. Only one specimen has been collected from WA, M.D. Barrett 188, 10.ii.1993 (PERTH), on Beverley Springs Station, and although it is curious that no other specimens have been collected for this species in WA, there are no reasons to doubt the locality. M. D. Barrett lived for some time on Beverley Springs Station and made several valuable collections from the area (Fig. 10D).
Habitat
Usually found in seasonally inundated situations, often in Melaleuca woodlands, sedge or herb fields, on sand, silt or clay loam.
Notes
This species is very similar to P. bifoliata, P. stenoclada and P. scorpioides; distinguishing characters are discussed under those species. Three outlier specimens, C.R. Dunlop 9464 & I.D. Cowie (DNA), I.D. Cowie 9490 (DNA) and K.G. Brennan 6962 (DNA), found in north-eastern Arnhem Land, are treated here under P. stenosepala, but are possibly hybrids with another species. They tend to have longer inflorescences, leaves, flowers and seeds than typical.
Typification
There are two specimens labelled R. Brown 4971, namely BM566304 and K279794. The BM Specimen 566304 is the only specimen with a hand-written label matching that of Flora Australiensis ‘Carpentaria Point and Arnhem Bays’. Specimen K279794 is simply labelled ‘Carpentaria’, and it is difficult to establish whether it is part of the same gathering. The number is known to have been added some time later (Orchard 1999). Because it does not match the full location description cited in Bentham (1863), it is not considered a syntype here, and therefore lectotypification is not required.
Selected specimens (36 examined)
41. Polygala succulenta R.A.Kerrigan, sp. nov.
Ab congeneribus in Australia haec species crista florali succulenta, semine oblongo, stylo recto planato, arillo appendiculato luteo et albo distinguenda.
Type: 780 m NE along Green Ant Ck towards East Springs, off Adelaide River Scenic Hwy, 02.ii.2004. R.A. Kerrigan 727 & R.K. Harwood; holo: DNA 161693!; iso: BRI, CANB, PERTH.
[Polygala arvensis auct. non Willd. (1802), Benth. Fl. Austral. 1 140 (1863) p.p. as to Upper Victoria R. F. Muell. MEL 2244589; A.J. Ewart & O.B. Davies, The Fl. of the N. Terr.: 160 (1917); I.D. Cowie & D.A. Albrecht (eds), Checklist N. Terr. Vasc. Pl. Sp. (2004).]
Annual herb, erect with ascending branches, up to 40 cm high; indumentum of curved and straight hairs. Plant occasionally tinged maroon on stems and alae. Leaves linear, narrowly elliptic to oblanceolate, 10–77 mm long, 1.5–10.0 mm wide; apex acute, obtuse or rounded, mucro 0.3–0.5 mm long; petiole up to 1.5 mm long. Inflorescence axillary or supra-axillary, a raceme, rarely a solitary flower on internode preceding raceme, up to 80 mm long, not exceeding the leaves, rarely longer than the leaves. Pedicel up to 3 mm long. Alae position antrorse to perpendicular or deflexed to reflexed in fruit; herbaceous, lanceolate, straight to falcate or narrowly dimidiate, 4.1–9.4 mm long, 1.1–2.9 mm wide; with straight and sometimes curved hairs. Corolla colour recorded as purple; floral appendages, succulent, bifid to three to four times forked; upper petal claw 1.5 mm wide. Stamens terminate at stigma, monadelphous; staminal flap <0.5 mm wide. Style straight to slightly curved and extended beyond globular stigma, broadly dorso-ventrally flattened. Capsule perpendicular to deflexed, rarely reflexed, apex of alae diverging at ~30° from capsule or capsule aligned with alae. Capsule lacking wing, asymmetrical, oblanceolate, 3.5–4.5 mm long, 2.0–2.8 mm wide; capsule lobe apices rounded and separated. Seed ovoid–oblong, 2.75–3.75 mm long, 0.9–1.5 mm wide; indumentum of fine white or ferruginous hairs. Aril head domed, 0.4–0.6 mm long, white, with short hairs. Aril appendages 3, linear, 0.3–1.0 mm long (Figs 2A, 3H, 5A).
There are two varieties, which can be distinguished as follows:
|
1 Inflorescence not congested in leaf axils, up to 80 mm long, flowers generally not overlapping |
Distribution
Australia (NT, WA). In the NT, distributed between Kakadu National Park and Nitmiluk National Park, the Victoria River District and Barkly Tablelands. In WA, several specimens are recorded along the Gibb River Road (Fig. 10E).
Habitat
Found mostly in woodland habitats in a variety of soils.
Notes
Two varieties are apparent, P. succulenta var. succulenta and P. succulenta var. congesta. P. succulenta var. succulenta is generally larger than P. succulenta var. congesta, has longer, less congested inflorescences and bigger flowers, and occurs mostly in Kakadu and Nitmiluk National Park. Similarities to P. petrophila are discussed under that species.
Etymology
Latin succulentus (succulent), referring to the succulent floral appendages.
Polygala succulenta var. congesta R.A.Kerrigan, var. nov.
A var. succulentae inflorescentiis congestis differt.
Type: Bullo River Station, on road to homestead, 08.iii.2006, R.A. Kerrigan 989; holo: DNA 177026!; iso: BRI, CANB, PERTH.
Inflorescence congested in leaf axils, up to 20 mm long, flowers overlapping.
Etymology
Latin congestus (crowded together), referring to the congested nature of the inflorescence.
Selected specimens (39 examined)
Polygala succulenta R.A.Kerrigan var. succulenta
Inflorescence not congested in leaf axils, up to 80 mm long, flowers generally not overlapping.
Selected specimens (21 examined)
42. Polygala tepperi F.Muell., The Victorian Naturalist 7: 38(1890)
Type: Roebuck-Bay; W. Tepper s.n.; holo: MEL 2263628! iso: K! (photograph DNA!), PERTH!.
Polygala arvensis var. stenosepala Benth., Fl. Austral. 1: 141 (1863). Polygala chinensis var. stenosepala (Benth.) Domin, Biblioth. Bot. 89: 303 (1927): Lectotype (here designated): Upper Victoria R., F. Muell.; lecto- K279791!; excluded syntypes: Arnhem Bay, R. Brown.
Robust annual herb, erect with ascending branches, up to 65 cm high; indumentum of curved hairs throughout, very rarely with short straight appressed to spreading hairs or glabrescent. Leaves more or less caducous, plant often leafless or with only a few leaves, linear, 5–35 mm long, 1.0–2.5 mm wide; apex acuminate or acute, mucro ~0.3 mm long; petiole 0–0.5 mm long. Inflorescence axillary or supra-axillary, a solitary flower or a short few-flowered raceme, up to 5(–10) mm long, not exceeding the leaves. Pedicel up to 3 mm long. Alae position antrorse in fruit; herbaceous, oblanceolate, 3.6–6.2 mm long, 1.2–2.0 mm wide; often with margins ciliate, outer surface with appressed hairs. Corolla purple; floral appendages fimbriate; upper petal claw 1 mm wide. Stamens terminate at stigma, monadelphous; staminal flap ~0.75 mm wide. Style hooked (horseshoe-shaped at apex), more or less terete, with flat stigma along inner surface of hook. Capsule antrorse, more or less aligned with alae. Capsule lacking wing, more or less symmetrical, oblong, locule apex acute, rarely obtuse, 4.0–6.5 mm long, 2.3–3.5 mm wide; capsule lobe apices acute or less often rounded and separated. Seed oblong, 3.0–4.5 mm long, 1.0–1.8 mm wide; indumentum of fine white or ferruginous hairs. Aril head helmet-shaped, 0.8–1.0 mm long, white, brown where reduced to seed testa, with short hairs, often concentrated along groove. Aril appendages absent or 3 much reduced triangular projections, 0.25–0.9 mm long.
Distribution
Australia (NT, WA). Widely distributed across WA and the NT mostly between 20°S and 15°S (Fig. 10F).
Habitat
Found in eucalypt woodlands and Acacia shrublands mostly with a Triodia understorey in sandy soils.
Nomenclatural notes
As a Mueller name, the MEL specimen is considered the holotype as it was the author’s host institution, in line with Recommendation 9A.4 of the International Code of Botanical Nomenclature (McNeill et al. 2006). The excluded syntype for P. arvensis var. stenosepala Benth. is a different species, namely P. parviloba.
Selected specimens (33 examined)
43. Polygala triflora L., Sp. Pl. 2: 705 (1753)
Type: ‘in Zeylona’ lecto: Hermann Herbarium, Mus. Zeyl. 31: ‘linaria minima seylanica’ fol. 10 of volume 3. 269; lecto: BM 621828 & BM 621829 (photographs DNA!); fide D.B.Sumithraarachchi, Rev. Hand. Fl. Ceylon 6 : 310 (1988).
Polygala chinensis var. dissitiflora Domin, Biblioth. Bot. 89: 303 (1927). Type citation: Queensland: Sandsteinhügel der Dividing Range bei Pentland (Domin II. 1910); Type detail: in collibus arenaceis Dividing Range dictis pr. opp. Pentland, K. Domin 5693; holo: PR 528253! (photograph BRI!).
[Polygala linariifolia [Australian material] auct. non Willd. (1802): Adema, Proc. R. Soc. Qld. 80 (9) 126 (1969); fide Van der Meijden, Fl. Males. I: 10: 481(1988).]
Annual herb or more or less perennial, often with thickened tap root, erect with ascending or spreading branches, up to 30 cm high, sometimes woody at base; indumentum of curved hairs only or with straight and curved hairs. Leaves linear to oblanceolate or obovate, 7–48 mm long, 2–7 mm wide; apex acute, rounded, truncate or retuse, mucro 0.3–0.5 mm long; petiole 0–1 mm long. Inflorescence supra-axillary or axillary, a raceme, up to 90 mm long, exceeding the leaves, but not past leaves at the top of plant. Pedicel up to 2 mm long. Alae position perpendicular to deflexed in fruit; herbaceous, lanceolate, usually falcate, 3.3–4.4 mm long, 1.0–1.8 mm wide; variable, as per whole plant. Corolla not seen fresh, colour recorded as purple, light purple to yellow, or white; floral appendages fimbriate; upper petal claw 1 mm wide. Stamens terminate at stigma, monadelphous; staminal flap ~0.6 mm wide. Style hooked (horseshoe-shaped at apex), in dried state dorso-ventrally flattened, with flat stigma along inner surface of hook. Capsule perpendicular to deflexed, becoming reflexed, more or less aligned with capsule or apex of alae sometimes diverging ~30° from capsule. Capsule lacking wing, widely ovate to widely oblong to elliptic, 3.2–4.5 mm long, 2.5–3.3 mm wide; capsule lobe apices rounded and separated. Seed ovoid–oblong, 3–4 mm long, 1.0–1.4 mm wide; indumentum of fine white hairs becoming ferruginous. Aril head helmet-shaped, 0.5–0.8 mm long, white, with brown groove where reduced to testa, fine short hairs along groove. Aril appendages 3, oblong, 0.30–0.75 mm long.
Distribution
Australia (NSW, QLD, SA), Sri Lanka, South-east Asia, northern Sumatra, northern Borneo, Celebes, Molluccas, Philippines, New Guinea. This species is widely distributed in QLD, extending from Mount Fox, near Ingham, to as far south as Warialda in NSW, with the bulk of the distribution extending inland along the lower western slopes of the Great Dividing Range. A few collections are also known further west near Cunnamulla, Currawinya National Park, Lake Eyre in north-eastern SA and the Diamantina National Park. The sole collection of Mueller’s from NT is treated as dubious, with no specimens of this species collected from the NT or WA since (Fig. 10G).
Habitat
Usually found in a variety of eucalypt woodlands in sandy soil and loam.
Notes
This species is widespread and variable with annual and short-lived perennial forms. Although two variants exist, including those with curved hairs only and those with curved and straight hairs, they have overlapping distributions, and no additional evidence supports treating them as separate taxa. This species is very similar to P. glaucoides and is distinguished in the present treatment by an erect and ascending habit, compared with a spreading or creeping habit, and by purple flowers versus yellow flowers. This species is also very similar to P. scorpioides; differences are discussed under that species. Some specimens approaching P. scorpioides may be intergrades, e.g. I.G. Champion 1284 (BRI) and P.I. Forster 14881 (BRI).
Nomenclatural notes
The application of the name for this taxon remains difficult because it relates to a group of taxa that have a difficult nomenclatural history, as per P. glaucoides. This taxon is quite variable in Australia and has generally been treated under the name P. linariifolia Willd., following Adema (1969), who indicated that the Australian material differed in flower colour and capsule shape from the Malesian material. Van der Meijden (1988) synonymised P. linariifolia Willd. (non Adema) under P. triflora and P. linariifolia non Willd. sensu Adema (1966) under P. glaucoides in his Flora Malesiana treatment. However, Australian material previously identified as P. linariifolia non Willd. sensu Adema were determined as P. triflora by van der Meijden, including the type specimen (B-W12958) cited by Adema (1966). This is confounded further in that the illustrations used are replicated from Adema and do not match the descriptions, i.e. P. triflora is described as having long hairs on the capsule margin, and yet the illustration shows curved hairs and is the same as that used by Adema to illustrate his concept for P. linariifolia.
The synonymy, types and circumscription of P. glaucoides and P. triflora need further examination before the assignation of this name to Australian material can be verified. I have seen a photograph of the type of P. triflora and it is not clearly attributable to Australian material. However, it is a relatively poor specimen. In the present treatment, I have deferred to van der Meijden’s (1988) description, treatment and annotated specimens, because he is likely to have seen the type and is likely to have had access to more material throughout the taxon’s global distribution.
Within the Australian group, two inconsistencies with van der Meijden’s treatment can be found, namely, flower colour and indumentum. Flower colour recorded from dry material in this instance is problematic and unreliable. Polygala flowers that are yellow with purple tinges have been recorded by collectors as either yellow or purple only. Also, purple flowers often fade to white with age or can fade to yellow with drying. Van der Meijden (1988) described this taxon as having yellow flowers. I have only seen one fresh flower, which was light purple in colour, and yet in Australia, collections record white, yellow and purple flower colour. The indumentum is equally unreliable; van der Meijden (1988) described P. triflora as having curved hairs throughout, with straight and curved hairs on the capsule margin. Pendry (2001), however, recorded P. triflora as having curved or rarely straight hairs throughout, with curved and straight hairs on the capsule margin. It is difficult with Australian material to separate specimens with straight and curved hairs on any other character from specimens with curved hairs only.
The nomenclatural and taxonomic issues relating to this group are clearly unresolved. To avoid complicating things further, the present treatment follows van der Meijden (1988), but acknowledges the need for much more work across the global distribution of this taxon and P. glaucoides before the status of the Australian material can be verified.
There appears to be no formal lectotypification in Sumithraarachchi (1987). The lectotype is considered legitimate here because it is reported as such by the Linnaean Typification Project. The lectotype consists of two individuals on a page of several different species. The barcode number BM 595652, as reported in Linnaean Database available online (see http://www.nhm.ac.uk/research-curation/research/projects/linnaean-typification/, accessed 25 March 2010; record last updated 26 June 2006), refers to the barcode given to the whole page. The barcode numbers BM 621828 and BM 621829 cited under the lectotype in my description correspond to the individual numbers given to both specimens of Polygala on this page. Presumably, the numbers were added after the image was loaded.
Selected specimens (46 examined)
44. Polygala validiflora R.A.Kerrigan, sp. nov.
Ab congeneribus in Australia haec species sepalis late ovatis conspicuis and sepalis lateralibus latioribus quam capsulis distinguenda.
Type: 250 m NE along Green Ant Ck towards East Springs, off Adelaide River Scenic Hwy, 02.ii.2004, R.A. Kerrigan 726 & R.K. Harwood; holo: DNA 161694!; iso: BRI, CANB, L, PERTH.
Annual herb, erect with spreading or ascending branches or decumbent, up to 20 cm high; indumentum of curved and straight hairs. Leaf margins and alae occasionally tinged red. Leaves lanceolate, elliptic to obovate 9–95 mm long, 3–23 mm wide; apex rounded, obtuse or truncate, lower leaves often acute, mucro 0.5–0.6 mm long; petiole 0–2 mm long. Inflorescence supra-axillary or axillary, a raceme, up to 27(–40) mm long, not exceeding the leaves. Pedicel lacking up to 1.5 mm long. Alae position antrorse to perpendicular in fruit; herbaceous, ovate, 7.7–12.0 mm long, (2.8–)4.5–9.0 mm wide; margins ciliate, otherwise glabrous or glabrescent. Corolla pale purple; floral appendages spathulate; upper petal claw <0.5 mm wide. Stamens terminating at stigma, monadelphous; staminal flap <0.5 mm wide. Style hooked (horseshoe-shaped at apex), in dried state dorso-ventrally flattened and twisted, with flat stigma along inner surface of hook. Capsule antrorse to perpendicular, more or less aligned with alae. Capsule with wing, 0.3–0.4(–1.5) mm wide, more or less equal throughout, more or less symmetrical, oblong or obovate, 4.5–5.3 mm long, 2.7–3.7 mm wide; capsule lobe apices rounded and separated. Seed oblong, 3.5–4.4 mm long, 1.3–2.0 mm wide; indumentum of fine white hairs. Aril head round to hooked, 0.5–0.7 mm long, white or brown and white, with hairs. Aril appendages 3, linear, 0.8–1.8 mm long.
Distribution
Australia (NT). Endemic to the NT, distributed from Tipperary Waters in the Daly Region to Katherine and southern Kakadu (Fig. 10H).
Habitat
Found in woodlands on a variety of substrates, often on drainage flats or seasonally wet areas.
Notes
This species is easily distinguished by its large alae and generally more robust form.
Etymology
Latin validus- (robust) and -flos (flower), referring to the large alae.
Selected specimens (25 examined)
45. Polygala wightiana Wight & Arn., Prod. Fl. Penin. Ind. Orient. 1: 38 (1834)
Type citation: Dindygul; Type details: [no location details given] Wallich 4190; syn- CGE, E, L 16576! (photograph), NYx2 (photograph!), K; Wight cat n. 134; syn- CGE, E 0017413 (photograph!).
Polygala stenoclada var. queenslandica Domin, Biblo. Bot. 89: 859 (1927). Type citation: Savannenwälder bei Chillagoe, Mareeba, Pentland und auf dem Castle Hill bei Townsville (Domin II-III. 1910); Type details: in xerodrymiis apud opp. Chillagoe K. Domin 5699, Pentland K.R. Domin 5698, Mareeba K.R. Domin 5700, in collibus Castle Hill prope opp. Townsville K.R. Domin 5701; syn- PR 528258 to 528261! (photograph BRI!) Type citation: ohne nähere Standortsangabe [English translation = without closer indication of location], BOWMAN; Type detail: Cape R., BOWMAN 274; syn- MEL 37324!?
Annual herb, erect with ascending branches, up to 1 m high; glabrous throughout. Stems and leaves glaucous. Leaves linear or narrowly oblong, 7–27 mm long, 1–3 mm wide; apex acute, mucro ~0.3 mm long; sessile. Inflorescence supra-axillary, rarely axillary or leaf-opposed, appearing terminal, usually a raceme, rarely with solitary flower on internode preceding raceme, up to 290 mm long, greatly exceeding the leaves. Pedicel up to 2 mm long. Alae position mostly antrorse sometimes perpendicular in fruit, herbaceous, ovate to narrowly dimidiate, 3.0–4.8 mm long, 1.1–2.1 mm wide; glabrous. Corolla yellow; floral appendages fimbriate; upper petal claw ~0.6 mm wide. Stamens terminate at stigma, triadelphous, (3, 2, 3); staminal flap ~0.8 mm wide. Style hooked (horseshoe-shaped at apex), more or less terete, with flat stigma along inner surface of hook, inner surface of style enlarged into flat fin-like membrane just below end of hook. Capsule mostly antrorse, sometimes perpendicular, more or less aligned with alae. Capsule lacking wing, more or less symmetrical, oblong, 3.2–4.0 mm long, 2.2–2.6 mm wide; capsule lobe apices rounded and separated. Seed oblong, 3.0–3.4 mm long, 1.0–2.4 mm wide; indumentum of fine white or ferruginous hairs. Aril head helmet-shaped, 0.6–0.9 mm long, white, with short hairs. Aril appendages 3, very shortly linear to oblong, 0.25–0.50 mm long (Fig. 6C).
Distribution
Australia (NT, QLD, WA), India, South-east Asia and Malesia. In Australia, it is distributed across the Kimberley in WA, at the far north of the NT and on Cape York Peninsula in QLD (Fig. 10I).
Habitat
Found in open woodlands in sandy, silt or clay soils.
Nomenclatural notes
Wight and Arnott (1834) cited two specimens in their protologue. The lower right-hand element on Sheet E 0017413 (E) labelled ‘Wight propr. 134’ is the syntype cited by Wight and Arnott (1834). Both Wallich 4190 and Wight 134 appear to fit the protologue and represent the same species. There is no clear evidence that Wight 134 is a paratype as stated by Adema (1966), and both are treated here as syntypes.
The question mark (above) after MEL 37324! indicates the uncertainty regarding the identification of this specimen as a syntype of P. stenoclada var. queenslandica Domin. It is the only candidate available, but with such vague type details it is difficult to be authoritative.
Specimens examined (20)
WESTERN AUSTRALIA: 250 m NE of Beverley Springs Station Homestead, W Kimberley, WA, 01.ii.1996, R.L. Barrett 785 (PERTH); Lennard R., 10 miles [16 km] above the junction of Baker R., 1.v.1905, W.V. Fitzgerald 576 (PERTH); Kimberley, 6.3 km E of Barnett R. crossing on Gibb R. Rd, 14.v.2006, R.A. Kerrigan 1116 (DNA, PERTH); Kimberley, 62.2 km E of Barnett R. Gorge turnoff on the Gibb R. Rd, 14.v.2006, R.A. Kerrigan 1125 (DNA, PERTH); Kimberley, 26.4 km N of Gibb R. Rd and Kalumburu intersection on Kalumburu road, 14.v.2006, R.A. Kerrigan 1126 (DNA, PERTH); Kimberley, 7.5 km S of Drysdale R. Homestead on Kalumburu road, 15.v.2006, R.A. Kerrigan 1127 (BRI, CANB, DNA, L, MEL, PERTH); Kimberley District, 1887, C.W. Nyulasy s.n. (MEL). NORTHERN TERRITORY: ~75 miles [121 km] NE of Maranboy Police Station, 05.iii.1965, M. Lazarides 88 & L.G. Adams (BRI, CANB, K, L, NSW, NT, US); Flora R., 08.iv.1997, C.R. Michell 756 & D.S. Calliss (DNA). QUEENSLAND: Don R., Edgecomb Bay, 1886, C.M. Birch s.n. (MEL); Cape R., E.M. Bowman 274 (MEL); Castle Hill, Townsville, 24.vi.1990, R.J. Cumming 10264 (BRI); Herald I., NW of Townsville, 13.iii.1998, R.J. Cumming 16996 (DNA, BRI); Chadshunt, Gilbert R., 05.v.1954, S.L. Everist 5408 (BRI); Strathmore, 06.v.1954, S.L. Everist 5422 (BRI, CANB); Great Basalt Wall, 03.iv.1995, R.J. Fensham 2132 (BRI, DNA); Burke Development Rd, 8.2 km E of Chillagoe, on S side of road, 30.iii.2007, R.A. Kerrigan 1188 (BRI, DNA); Burke Development Rd, at Mt Garnet and Mt Surprise turnoff, SE side of intersection, 30.iii.2007, R.A. Kerrigan 1189 (BRI, DNA); ~89 km N of Hughenden, 01.iv.1998, E.J. Thompson & G. Turpin 526 (BRI, DNA).
Notes on informal, misapplied, excluded and doubtful names
Polygala arvensis Willd., Sp. Pl. Edn. 3: 876 (1802)
Type: in India orientali; holo: B (photograph DNA!).
A synonym of Polygala chinensis L. sensu van der Meijden, Flora Malesiana I vol. 10: 478 (1988). Two of the three specimens cited by Bentham (1863) under P. arvensis Willd. are now referred to P. parviloba and P. succulenta. The third specimen, P. arvensis auct. non Willd: Benth., Fl. Austral. 1 : 140 (1863). North Coast, R.Brown, was not located. A potential candidate for this specimen is a sheet at K of a mixed gathering with different elements identified with three different labels. The left-hand element is labelled on white paper P. arvensis var. [no epithet recorded] ‘Broadsound, Bowman’, the middle element is labelled on blue paper ‘R. Brown, Iter Australiense, 1802–5’, Polygala arvensis, No. 4973?, Rec 2.6/78’, and the right-hand element is labelled on white paper ‘Port Darwin North Australia from R. Schomburgk Oct 1869 [?apparently], 130. Polygala arvensis’. This third element is likely to be an F. Schultz specimen, as Schomburgk sent specimens to K after the publication of Vol. 1 of Flora Australiensis (Bentham 1863). The middle element now identified as P. parviloba R.A.Kerrigan may be the specimen recorded by Bentham (1863) for P. arvensis auct. non Willd.
On the basis of specimens seen, P. arvensis Willd. has been widely misapplied to many different taxa.
Polygala chinensis L., Sp. Pl. 2: 704 (1753)
Type: habitat in India; lecto: LINN (photograph DNA!).
In her treatment of Australian Polygalaceae, Monro (2003) cited the lectotype of P. chinensis L. as Herb. Linn. 882.26, according to F. Fawcett & A. B. Rendle, Fl. Jamaica, Dicot. pt. 2, 4 : 244 (1920). Because there is no clear lectotypification in Fawcett and Rendle, the typification of this specimen remains unresolved. Burtt (1973), however, stated that ‘there is no reason to suppose that the specimen labelled P. chinensis in the Linnaean Herbarium (Sheet 882.26 of the Savage catalogue) is not the type specimen’, and reported that the type material consists of two lateral branches of a plant.
Assuming that Herb. Linn. 882.26 is the holotype, the name P. chinensis has been widely misapplied according to Burtt (1973) and van der Meijden (1988), and I am unable to assign the name to any Australian material. On the basis of photographs of putative type material of P. chinensis, I support van der Meijden’s (1988) concept and exclude his concept from Australian material.
Polygala crotalarioides DC., Prodromus 1: 327 (1824)
Type: in Nepauliâ... (v.s.); (microfiche DNA!). Lectotype: Wallich 4176a, 1829; lecto: G00076242
The author, de Candolle, stated vidi siccam (v.s.) in the protologue, meaning ‘I have it in dried state’ (Stearn 1992) and I assume the image of a dried specimen on microfiche in de Candolle (1824) is an image of the type. The Geneva Herbarium online catalogue cites Wallich 4176a as the lectotype (Catalogue des herbiers de Genève (CHG), Conservatoire & Jardin Botaniques de la Ville de Genève, available at http://www.ville-ge.ch/musinfo/bd/cjb/chg, accessed 05 August 2011). This species appears to be a woody multi-stemmed perennial, with showy alae and large obovate leaves with an obtuse apex. It bears no resemblance to any of the northern taxa treated here or the more southern native, P. japonica. It has been misapplied by Hooker (1859), perhaps to the introduced species P. myrtifolia, which was recorded from Victoria in 1880 (Walsh 1999). Also, it may be misapplied to P. triflora sensu R.A.Kerrigan, because a Mueller label (MEL 2244745) determined here as P. triflora sensu R.A.Kerrigan is annotated syn. P. crotalarioides and syn. P. rosmarinifolia, apparently in Mueller’s hand.
Polygala linariifolia [QLD Material] auct. non Willd.: Adema, Blumea 14(2): 268 (1966)
A synonym of P. triflora L. sensu van der Meijden, Flora Malesiana I vol. 10: 478 (1988).
Polygala linariifolia [WA material] auct. non Willd.: J.R. Wheeler, Fl. Kimberley 643 (1992)
According to specimens determined as P. linariifolia from WA, this name has been misapplied to several new species described here, including P. bifoliata, P. dimorphotricha, P. galeocephala and P. saccopetala.
Polygala rosmarinifolia Wight & Arn., Prodromus Florae Peninsulae Indiae Orientalis: 37 (1834)
Type: Wight! Cat. 124; 133. b (a younger state); 140, Asewalendan Hills and Cunnewaddy near Dindygul, Wallich L. n. 4169; syn- E (photographs of syntypes available from online catalogueat http://elmer.rbge.org.uk/bgbase/vherb/bgbasevherb.php!, accessed February 2007).
Like P. crotalarioides, this name is likely to be misapplied to P. triflora sensu R.A.Kerrigan, although the types bear little resemblance.
Informal names published in regional floras and checklists or the Australian Plant Census (APC) are listed in Table 3, along with the corresponding names accepted by this treatment.

|
Unlocated types
The type for the following epithet was not located at BM, BRI, CANB, DNA, K, MEL, NT, PERTH, PR or SA:
-
Polygala stenoclada var. clementii Domin, Biblioth. Bot. 89: 305 (1927)
Type: Nordwest-Australien: zwischen Ashburton – und Yule R., E. CLEMENT.
Types for the following epithets were not pursued. Non-native taxa were not treated here, apart from their inclusion in the key.
-
P. duarteana A.St.-Hil.
-
P. monspeliaca L.
-
P. myrtifolia L.
-
P. virgata Thunb.
-
P. vulgaris L.
Index to taxa
Current names are in bold, and new names, combinations and synonyms (including informal and misapplied names) are not bold.
-
P. abyssinica auct. non R.Br. ex Fresen.………………114
-
P. abyssinica var. intercedens Domin………………114
-
P. arvensis Willd. ………………132
-
P. arvensis auct. non Willd. ………………100
-
P. arvensis var. obovata Benth. ………………115
-
P. arvensis var. squarrosa Benth. ………………106
-
P. arvensis var. stenosepala Benth. ………………129
-
P. barbata R.A.Kerrigan, sp. nov. ………………99
-
P. barklyensis R.A.Kerrigan, sp. nov. ………………99
-
P. bifoliata R.A.Kerrigan, sp. nov.………………100
-
P. canaliculata R.A.Kerrigan, sp. nov.………………101
-
P. chinensis L. ………………133
-
P. chinensis auct. non L. ………………121
-
P. chinensis subvar. linearifolia Domin………………117
-
P. chinensis subvar. obovata Domin………………115
-
P. chinensis var. dissitiflora Domin………………130
-
P. chinensis var. orbicularis (Benth.) Domin………………116
-
P. chinensis var. squarrosa (Benth.) Domin………………106
-
P. chinensis var. stenosepala (Benth.) Domin………………129
-
P. clavistyla R.A.Kerrigan, sp. nov. ………………102
-
P. coralliformis R.A.Kerrigan, sp. nov. ………………102
-
P. crassitesta R.A.Kerrigan, sp. nov. ………………104
-
P. crotalarioides DC. ………………134
-
P. dependens R.A.Kerrigan, sp. nov. ………………104
-
P. difficilis R.A.Kerrigan, sp. nov. ………………104
-
P. dimorphotricha R.A.Kerrigan, sp. nov. ………………106
-
P. duarteana A.St.-Hil. ………………96
-
P. eriocephala F.Muell. ex. Benth. ………………106
-
P. eriocephala auct. non F.Muell. ex. Benth. ………………111
-
P. exsquarrosa Adema ………………106
-
P. gabrielae Domin ………………107
-
P. gabrielae auct. non Domin ………………103
-
P. galeocephala R.A.Kerrigan, sp. nov. ………………109
-
P. geniculata R.A.Kerrigan, sp. nov. ………………109
-
P. glaucifolia R.A.Kerrigan, sp. nov. ………………110
-
P. glaucoides L. ………………111
-
P. glaucoides auct. non L. ………………110
-
P. integra R.A.Kerrigan, sp. nov. ………………111
-
P. isingii Pedley………………112
-
P. japonica Houtt. ………………113
-
P. kimberleyensis R.A.Kerrigan, sp. nov. ………………114
-
P. leptalea DC. ………………114
-
P. leptalea var. australiensis Domin………………114
-
P. linariifolia auct. non Willd. ………………130
-
P. longifolia Poir. ………………114
-
P. macrobotrya Domin………………115
-
P. monspeliaca L. ………………97
-
P. myrtifolia L. ………………97
-
P. obversa nom. nov. ………………115
-
P. orbicularis Benth. ………………116
-
P. paniculata L. ………………117
-
P. parviloba nom. nov. ………………117
-
P. pendulina R.A.Kerrigan, sp. nov. ………………118
-
P. persicariifolia DC. ………………119
-
P. petrophila R.A.Kerrigan, sp. nov. ………………120
-
P. petrophila var. angustifolia R.A.Kerrigan, var. nov. ………………120
-
P. petrophila var. petrophila………………120
-
P. polifolia C.Presl………………121
-
P. praecox R.A.Kerrigan, sp. nov. ………………121
-
P. pterocarpa R.A.Kerrigan, sp. nov. ………………122
-
P. pycnantha, nom. nov. ………………123
-
P. pycnophylla Domin………………123
-
P. pycnophylla auct. non Domin………………100
-
P. rhinanthoides Sol. ex Benth. ………………123
-
P. rhinanthoides auct. non Sol. ex Benth. ………………122
-
P. rhinanthoides var. minor Benth. ………………109
-
P. rhynchocarpa R.A.Kerrigan, sp. nov. ………………124
-
P. rosmarinifolia Wight & Arn. ………………134
-
P. saccopetala R.A.Kerrigan, sp. nov. ………………125
-
P. scorpioides R.A.Kerrigan, sp. nov. ………………126
-
P. sibirica auct. non L. ………………113
-
P. stenoclada Benth. ………………126
-
P. stenoclada var. clementii Domin………………134
-
P. stenoclada var. queenslandica Domin………………132
-
P. stenoclada var. stenosepala Benth. ………………127
-
P. stenoclada var. typica Domin, nom. inval. ………………126
-
P. stenosepala, comb. et stat. nov. ………………127
-
P. succulenta R.A.Kerrigan, sp. nov. ………………128
-
P. succulenta var. congesta R.A.Kerrigan, var. nov. ………………129
-
P. succulenta var. succulenta………………129
-
P. tepperi F.Muell. ………………129
-
P. triflora L. ………………130
-
P. triflora auct. non L. ………………118
-
P. validiflora R.A.Kerrigan, sp. nov. ………………131
-
P. veronicea F.Muell. ………………113
-
P. virgata Thunb. ………………97
-
P. vulgaris L. ………………96
-
P. wightiana Wight & Arn. ………………132
Acknowledgements
I thank the staff at Northern Territory herbaria for their collection efforts, useful discussion and identification skill, which helped refine this treatment. I am also greatly indebted to the staff of Australian and overseas herbaria, in particular B, BM, BRI, CANB, CGE, DNA, E, G, GDC, K, L, LINN, MEL, P, PR, SA and SING, for supplying specimens and images. Australian Botanical Liaison Officers Alex George, Jenny Tonkin, Jeremy Bruhl and Juliet Wege provided invaluable assistance with locating, imaging and sending specimens. The Department of Natural Resources, Environment, the Arts and Sport (NRETAS) and James Cook University (JCU) provided support that allowed for the collection of specimens. I thank the library staff at NRETAs and JCU for their efforts in retrieving protologues. Monika Madsen Osterkamp provided botanical illustrations and assistance with German translation, Ellie Johnston scanned illustrations, and Emma Short provided comment on some Latin diagnoses. Ian Cowie, Bob Harwood, John Kerrigan, Anna Monro, Anne Penfold, Phil Short, Zita Stokes, Gloria Webb and anonymous referees and examiners provided valued comments on this manuscript. Donna Lewis, Andrea Hope and Ben Stuckey assisted with various and I’m sure inconvenient requests for references, data and images. Collection of herbarium specimens was undertaken with the following scientific research permits: WA SW010459, QLD WISP01882804. Field support was generously donated by Rory Ryan and John, Mhornin and Mark Kerrigan. This work is the result of a Master of Science Candidature with JCU and the support and supervision of Professor Paul Gadek (JCU) and Dr Dale Dixon (DNA). Special mention must be given to Ian Cowie for helping to refine the manuscript through the review process. Special mention also must be given to J. R. Abbott who provided rigorous feedback, proofreading and advice as a referee. Although at times this caused us both great angst, I am grateful for his contributions and commitment to seeing this manuscript presented in a more polished form. Both of these people never failed to respond swiftly and comprehensively to all enquiries, and their ongoing support enabled me to persevere with this manuscript.
References
Abbott JR (2011) Notes on the disintegration of Polygala (Polygalaceae) with four new genera for the Flora of North America. Journal of the Botanical Research Institute of Texas 5, 125–137.Adema F (1966) A review of the herbaceous species of Polygala in Malesia (Polygalaceae). Blumea 14, 253–356.
Adema F (1969) Identities of the herbaceous Australian species of Polygala represented in the Brisbane Herbarium. Proceedings of the Royal Society of Queensland 80, 125–130.
Anon. (2002) ‘Conservation action plan – public version. Polygala smallii.’ (Fairchild Tropical Botanic Garden.) Available at http://www.ftg.org/research/conservation/CAP%20Polygala%202003.doc [accessed 24 July 2006, last updated 10 May 2006]
Anon. (1998) ‘FloraBase – the Western Australian flora.’ (Western Australian Herbarium.) Available at http://florabase.calm.wa.gov.au/ [accessed 6 January 2005]
Bennett AW (1872) Polygala. In ‘The flora of British India. Part 1’. (Ed. JD Hooker) pp. 200–206. (L. Reeve & Co.: London)
Bentham G (1863) Polygala. In ‘Flora Australiensis. Vol. 1’. pp. 138–141. (Lovell Reeve & Co.: London)
Berg RY (1975) Myrmecochorous plants in Australia. Australian Journal of Botany 23, 475–508.
| Myrmecochorous plants in Australia.Crossref | GoogleScholarGoogle Scholar |
Bernardi LF (2000) Consideraciones taxonómicas y fitogeográficas acerca de 101 Polygalae Americanas. Cavanillesia Altera 1, 1–456.
Brantjes NBM (1982) Pollen placement and reproductive isolation between two Brazilian Polygala species (Polygalaceae). Plant Systematics and Evolution 141, 41–52.
| Pollen placement and reproductive isolation between two Brazilian Polygala species (Polygalaceae).Crossref | GoogleScholarGoogle Scholar |
Burbidge NT (1953) The genus Triodia R.Br. (Gramineae). Australian Journal of Botany 1, 121–184.
| The genus Triodia R.Br. (Gramineae).Crossref | GoogleScholarGoogle Scholar |
Burtt BL (1973) Polygala arvensis, chinensis and glomerata. Notes from the Royal Botanic Garden Edinburgh 32, 403–404.
Carolin R (1990) Nomenclatural notes and new taxa in the genus Goodenia (Goodeniaceae). Telopea 3, 517–570.
Castro S, Silvera P, Navarro L (2008) How does secondary pollen presentation affect the fitness of Polygala vayredae (Polygalaceae). American Journal of Botany 95, 706–712.
| How does secondary pollen presentation affect the fitness of Polygala vayredae (Polygalaceae).Crossref | GoogleScholarGoogle Scholar |
Chodat R (1893) Monographia Polygalacearum. Mémoires de la Société de Physique et d’Histoire. Naturelle de Genève 31, 1–500.
Chodat R (1896) Polygalaceae. In ‘Die Natürlichen Pflanzenfamilien. Vol. 3’. (Eds A Engler, K Prantl) pp. 323–345. (Engelman: Leipzig, Germany)
Clarkson JR, Kenneally KF (1988) The floras of Cape York and the Kimberley: a preliminary analysis. Proceedings of the Ecological Society of Australia 15, 259–266.
Cowie ID, Albrecht DA (2004) ‘Checklist of NT vascular plants’. (NT Herbarium, Department of Infrastructure Planning and Environment: Darwin)
Craven LA (1996) A taxonomic revision of Heliotropium (Boraginaceae) in Australia. Australian Systematic Botany 9, 521–657.
| A taxonomic revision of Heliotropium (Boraginaceae) in Australia.Crossref | GoogleScholarGoogle Scholar |
Cronquist A (1981) Polygalaceae. In ‘An integrated system of classification of flowering plants’. pp. 775–778. (Columbia University Press: New York)
Davis PH, Heywood VH (1963) ‘Principles of angiosperm taxonomy.’ (Oliver and Boyd: London)
de Candolle AP (1824) Polygalae. In ‘Prodomus systematis naturalis regni vegetabilis Vol. 1’. Microfiche edition. (Ed. AP de Candole) pp. 321–342. (Treuttel et Wurtz: Strasbourg)
Domin K (1927) Beiträge zur Flora und Pflanzengeographie Australiens. Bibliotheca Botanica 89, 301–305.
Dube VP, Awasthi AK (1985) Morphological and anatomical studies of floral nectaries in Polygalaceae. Journal of the Indian Botanical Society 64, 231–235.
Dunlop CR (1996) Mitrasacme. Flora of Australia 28, 29–57.
Dunlop CR, Webb LJ (1991) Flora and vegetation. In ‘Monsoonal Australia. Landscape, ecology and man in the northern lowlands’. (Eds CD Haynes, MG Ridpath, MAJ Williams) pp. 41–60. (Balkema: Rotterdam, The Netherlands)
Duretto MF (1997) Taxonomic notes on Boronia species of north-western Australia, including a revision of the Boronia lanuginosa group (Boronia section Valvatae: Rutaceae). Nuytsia 11, 301–346.
Duretto MF (1999) Boronia sect. Valvatae (Benth.) Engl. (Rutaceae) in Queensland, Australia. Austrobaileyea 5, 263–298.
Duretto MF, Ladiges PY (1997) Morphological variation within the Boronia grandisepala group (Rutaceae) and the description of nine taxa endemic to the Northern Territory, Australia. Australian Systematic Botany 10, 249–302.
| Morphological variation within the Boronia grandisepala group (Rutaceae) and the description of nine taxa endemic to the Northern Territory, Australia.Crossref | GoogleScholarGoogle Scholar |
Eriksen B (1993a) Floral anatomy and morphology in the Polygalaceae. Plant Systematics and Evolution 186, 17–32.
| Floral anatomy and morphology in the Polygalaceae.Crossref | GoogleScholarGoogle Scholar |
Eriksen B (1993b) Phylogeny of the Polygalaceae and its taxonomic implications. Plant Systematics and Evolution 186, 33–55.
| Phylogeny of the Polygalaceae and its taxonomic implications.Crossref | GoogleScholarGoogle Scholar |
Eriksen B, Persson C (2007) Polygalaceae. In ‘The families and genera of vascular plants, vol. 9’. (Ed K Kubitzki) pp. 345–363. (Springer: Berlin)
Fawcett F, Rendle AB (1920) Polygalaceae. Flora of Jamaica 4, 242–245.
Forest F, Chase MW, Persson C, Crane PR, Hawkins JA (2007) The role of biotic and abiotic factors in evolution of ant dispersal in the milkwort family (Polygalaceae). Evolution 61, 1675–1694.
| The role of biotic and abiotic factors in evolution of ant dispersal in the milkwort family (Polygalaceae).Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXltFCltb8%3D&md5=d5ee8e8cfdb5bd00a43c2324ec7ae7c7CAS |
Halford DA (1997) Notes on Tiliaceae in Australia, 3: a revision of the genus Triumfetta L. Austrobaileya 4, 495–587.
Harwood R, Dessein S (2005) Australian Spermacoce (Rubiaceae: Spermacoceae). 1. Northern Territory. Australian Systematic Botany 18, 297–365.
| Australian Spermacoce (Rubiaceae: Spermacoceae). 1. Northern Territory.Crossref | GoogleScholarGoogle Scholar |
Hedberg O (1958) The taxonomic treatment of vicarious taxa. Uppsala Univeritet Arsskrift 6, 186–195.
Henderson RJF (1984) Bibliography of Karel Domin’s Beiträge on Australian plants. Taxon 33, 673–679.
| Bibliography of Karel Domin’s Beiträge on Australian plants.Crossref | GoogleScholarGoogle Scholar |
Henderson RJF (2002) ‘Names and distributions of Queensland plants, algae and lichens. Queensland Herbarium. 16.’ (Queensland Herbarium, Department of Environment and Heritage: Indooroopilly, Qld)
Hooker JD (1859) The botany of the Antarctic voyage of H.M. discovery ships Erebus and Terror. III. Flora Tasmaniae 2, xiii
IUCN (2001) ‘IUCN Red List categories and criteria: version 3.1.’ Prepared by the IUCN Species Survival Commission. (IUCN: Gland, Switzerland)
Kerrigan RA (2008) ‘A taxonomic revision of Polygala in Australia.’ MSc Thesis, James Cook University, Cairns.
Kerrigan RA, Albrecht DA (2007) ‘Checklist of NT vascular plants.’ (NT Herbarium, Department of Infrastructure Planning and Environment: Darwin)
Kruger H, Pretorius WE (1997) Notes on the structure of the stigma of Polygala virgata var. virgata (Polygalaceae). South African Journal of Botany 63, 261–266.
Lack AJ, Kay QON (1987) Genetic structure, gene flow and reproductive ecology in sand-dune populations of Polygala vulgaris. Journal of Ecology 75, 259–276.
| Genetic structure, gene flow and reproductive ecology in sand-dune populations of Polygala vulgaris.Crossref | GoogleScholarGoogle Scholar |
Ladd PG (1994) Pollen presenters in the flowering plants, form and function. Botanical Journal of the Linnean Society 115, 165–195.
Ladd PG, Donaldson JS (1993) Pollen presenters in the South African flora. South African Journal of Botany 59, 465–477.
Lazarides M (1997) A revision of Triodia including Plectrachne (Poaceae, Eragrostideae, Triodiinae). Australian Systematic Botany 10, 381–489.
| A revision of Triodia including Plectrachne (Poaceae, Eragrostideae, Triodiinae).Crossref | GoogleScholarGoogle Scholar |
Leach GJ (2000) Notes and new species of Eriocaulon (Eriocaulaceae) from Australia. Australian Systematic Botany 13, 755–772.
| Notes and new species of Eriocaulon (Eriocaulaceae) from Australia.Crossref | GoogleScholarGoogle Scholar |
Linnaeus C (1753) Polygala. In ‘Species plantarum. Vol. 2’. pp. 701–706. (Impensis Laurentii Salvii: Stockholm)
Mallet J (1995a) A species definition for the modern synthesis. Trends in Ecology & Evolution 10, 294–299.
| A species definition for the modern synthesis.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BC3M7itFaksA%3D%3D&md5=6b90926311645b5f5fe127f22a3f5077CAS |
Mallet J (1995b) A species definition: a functional approach. Trends in Ecology & Evolution 10, 490–491.
| A species definition: a functional approach.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BC3M7itFensA%3D%3D&md5=ed20fa0d3922a1bdca116d908b648186CAS | [Reply]
Mauseth JD (1988) ‘Plant anatomy.’ (The Benjamin/Cummings Publishing Company: Menlo Park, CA)
Mayr E (1993) Fifty year of progress in research on species and speciation. Proceedings of the California Academy of Sciences 48, 131–140.
McDade LA (1995) Species concepts and problems in practice: insights from botanical monographs. Systematic Botany 20, 606–622.
| Species concepts and problems in practice: insights from botanical monographs.Crossref | GoogleScholarGoogle Scholar |
McNeill J, Barrie R, Burdet HM, Demoulin V, Hawksworth L, Marhold K, Nicolson DH, Prado J, Silva PC, Skog JE, Wiersema JH, Turland NJ (2006) ‘International code of botanical nomenclature (Vienna Code).’ Regnum Vegetabile 146 [A.R.G. Gantner Verlag KG]. Available at http://ibot.sav.sk/icbn/main.htm [verified March 2012]
Monro A (2003) Systematics of the Australian Polygalaceae and Xanthophyllaceae. PhD Thesis, Australian National University, Canberra.
Munir AA (1979) A taxonomic revision of the genus Pityrodia (Chloanthaceae). Journal of the Adelaide Botanical Gardens 2, 1–138.
Murray L (1992) Polygalaceae. In ‘Flora of NSW. Vol. 3’. (Ed. GJ Harden) pp. 3–7. (University Press: Sydney)
Oostermeijer JGB (1989) Myrmecochory in Polygala vulgaris L., Luzula campestris (L.) DC., and Viola curtisii Forster in a Dutch dune area. Oecologia 78, 302–311.
| Myrmecochory in Polygala vulgaris L., Luzula campestris (L.) DC., and Viola curtisii Forster in a Dutch dune area.Crossref | GoogleScholarGoogle Scholar |
Orchard AE (1999) A history of systematic botany in Australia. In ‘Flora of Australia. Introduction. Vol. I’. (Eds AE Orchard, HS Thompson) pp. 11–103. (Australian Biological Resources Study/CSIRO Publishing: Melbourne)
Paiva JAR (1998) Polygalarum Africanarum et Madagascariensium prodromus atque gerontogaei generis Heterosamara Kuntze, a genere Polygala L. segregati et a nobis denuo recepti, synopsis monographica. Fontqueria 50, 1–346.
Pendry CA (1999) A new combination of Polygala (Polygalaceae) for Southeast Asia. Novon 9, 545
| A new combination of Polygala (Polygalaceae) for Southeast Asia.Crossref | GoogleScholarGoogle Scholar |
Pendry CA (2001) Polygalaceae. Flora of Thailand 7, 498–520.
Persson C (2001) Phylogenetic relationships in Polygalaceae based on plastid DNA sequences from the trnL–F region. Taxon 50, 763–779.
| Phylogenetic relationships in Polygalaceae based on plastid DNA sequences from the trnL–F region.Crossref | GoogleScholarGoogle Scholar |
Radford AE (1986) Plant description. In ‘Fundamentals of plant systematics’. pp. 107–145. (Harper & Row Publishers Inc.: New York)
Short PS (2002) John Richardson, an early collector from Melville Is., Northern Territory. Australian Systematic Botany Society Newsletter 112, 3–7.
Sneath PH, Sokal RR (1973) ‘Numerical taxonomy. The principles and practice of numerical classification.’ (W.H. Freeman and Company: San Francisco, CA)
Specht RL (1958) The history of botanical exploration in the Arnhem Land Aboriginal Reserve. In ‘Records of the American–Australian scientific expedition to Arnhem Land. 3. Botany and plant ecology’. (Eds RL Specht, CP Mountford) pp. 1–7. (Melbourne University Press: Melbourne)
Stearn WT (1992) ‘Botanical Latin.’ 4th edn. (David & Charles: Newton Abbott, UK)
Stevens PF (2001–). Angiosperm phylogeny website. Version 8, June 2007 [and more or less continuously updated since]. Available at http://www.mobot.org/mobot/research/apweb/ [accessed 26 March 2010].
Stuessy TF (1990) ‘Plant taxonomy: the systematic evaluation of comparative data.’ (Columbia University Press: New York)
Sumithraarachchi DB (1987) Polygalaceae. In ‘A revised handbook to the flora of Ceylon. Vol. VI’. (Eds MD Dassanayake, FR Fosberg) pp. 301–317. (A.A. Balkema: New Delhi, India)
Toelken HR (2010) Notes on Hibbertia (Dilleniaceae) 5. H. melhanioides and H. tomentosa groups from tropical Australia. Journal of the Adelaide Botanic Gardens 23, 1–117.
van der Meijden R (1988) Polygalaceae. Flora Malesiana 10, 455–482.
Verkerke W (1985) Ovules and seeds of the Polygalaceae. Journal of the Arnold Arboretum. Arnold Arboretum 66, 353–394.
Verkerke W, Bouman F (1980) Ovule ontogeny and its relation to seed-coat structure in some species of Polygala (Polygalaceae). Botanical Gazette 141, 277–282.
| Ovule ontogeny and its relation to seed-coat structure in some species of Polygala (Polygalaceae).Crossref | GoogleScholarGoogle Scholar |
Walsh N (1999) Polygalaceae. In ‘Flora of Victoria. Vol. 4’. (Eds NG Walsh, TJ Entwisle). pp. 130–137. (Inkata Press: Melbourne)
Westerkamp C, Weber A (1997a) Keel flowers of Polygalaceae and Fabaceae: how far do similarities reach. In ‘13th Symposium Morphology, Anatomy & Systematics, programmes & abstracts’. (Eds E Smets, LP Ronse Decraene, E Robbrecht) pp. 165–166. Scripta Botanica Belgica 15.
Westerkamp C, Weber A (1997b) Secondary and tertiary pollen presentation in Polygala myrtifolia and allies (Polygalaceae, South Africa). South African Journal of Botany 63, 254–258.
Wheeler JR (1992) Polygalaceae. In ‘Flora of the Kimberley Region’. (Ed. JR Wheeler) pp. 638–644. (Department of Conservation & Land Management: Perth)
Wight R, Arnott GAW (1834) Polygala. In ‘Prodromus Florae Peninsulae Indiae Orientalis’. pp. 36–39. (Pardury, Allen and Co.: London)
Woinarski JC, Hempel C, Cowie I, Brennan K, Kerrigan R, Leach G, Russell-Smith JR (2006) Distributional patterning of restricted range plant species in the Northern Territory, Australia. Australian Journal of Botany 54, 627–640.
| Distributional patterning of restricted range plant species in the Northern Territory, Australia.Crossref | GoogleScholarGoogle Scholar |