

In search of sustainable livestock management in the Dry Chaco: effect of different shrub-removal practices on vegetation
L. M. Guzmán A * , P. E. Villagra B C , R. E. Quiroga D , D. I. Pereyra A , M. E. Pelliza A , A. R. Ricarte A and L. J. Blanco AA Instituto Nacional de Tecnología Agropecuaria (INTA), Estación Experimental Agropecuaria La Rioja, Chamical, La Rioja 5380, Argentina.
B Instituto Argentino de Nivología, Glaciología y Ciencias Ambientales, CCT CONICET, Mendoza 5505, Argentina.
C Facultad de Ciencias Agrarias, UNCUYO, Chacras de Coria, Mendoza 5505, Argentina.
D Instituto Nacional de Tecnología Agropecuaria (INTA), Estación Experimental Agropecuaria Catamarca, Sumalao, Catamarca 4705, Argentina.
The Rangeland Journal 44(4) 193-202 https://doi.org/10.1071/RJ22040
Submitted: 27 June 2022 Accepted: 3 January 2023 Published: 23 February 2023
© 2022 The Author(s) (or their employer(s)). Published by CSIRO Publishing on behalf of the Australian Rangeland Society. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Increasing shrub density hinders cattle raising in central Argentina rangelands. While roller-chopping and hand-cutting are used to remove shrubs and make land accessible for cattle grazing, studies on the effects of these practices on ecosystem composition, structure and functioning are scarce. We assessed the impact of shrub removal on (a) vegetation cover, composition, species richness and diversity and (b) green biomass and seasonal phenological dynamics. We used a randomised design with three treatments (degraded rangeland; roller-chopping and hand-cutting) with four replicates, and compared the treatments with the least disturbed woodland (conserved woodland). Cenchrus ciliaris L. was sown following removal treatments and grass cover increased significantly (45% roller-chopping and 84% hand-cutting vs 27% degraded rangeland, and 13% conserved woodland), and tree cover decreased (3% roller-chopping and 2% hand-cutting vs 16% degraded rangeland, and 23% conserved woodland). Shrub cover reached 45% under roller-chopping, similar to the degraded rangeland treatment (46% degraded rangeland, 60% conserved woodland), but in hand-cutting reached only 3% cover. Hand-cutting reduced species richness and vegetation diversity, whereas roller-chopping had no effect. Shrub-removal treatments decreased normalised difference vegetation index (NDVI; proxy of green biomass) and modified ecosystem phenology. Under both shrub treatments, the maximum, minimum and integral NDVI (area under the NDVI curve) were significantly lower than those of the degraded rangeland treatment. The results suggest that shrub-removal practices increase forage for livestock, but reduce functional diversity and compromise the return to less disturbed states (woodland), where tree populations are maintained.
Keywords: biomass production, disturbance, extensive grazing systems, grazing, rangeland management, shrublands, woodlands, woody-plant encroachment.
Introduction
Shrub encroachment on natural rangelands is a worldwide concern for ecosystem functioning documented in America, Australia and Africa (Archer 2010; Rejžek et al. 2017; Chandregowda et al. 2018). The main causes of encroachment include climate change, overgrazing, seed dissemination by livestock, reduced fire frequency, reduced competition with grasses and increased atmospheric CO2 (Van Auken 2000). Consequences include decreased forage grass cover and an increased erosion (Eldridge et al. 2011). Thus, in the absence of shrub treatment, thresholds could be reached where vegetation and soil changes persist (Willcox and Giuliano 2010; Bestelmeyer et al. 2015).
In North America, shrub-encroachment rates range from ~0.2% cover per year in the Sagebrush Steppe region to 1.6% in the Central Great Plains. In semi-arid regions, shrub encroachment may contribute to increased aboveground net primary production (Barger et al. 2011). In Argentina, the ‘shrub eradication paradigm’ has led to the application of various methods to eliminate shrubs, but in so doing it has also damaged tree populations. In this sense, the development of cattle ranching in the Chaco region brought with it production models designed for regions with different agro-ecological conditions (e.g. the Pampean region), where disturbances aim to create homogeneous, shrub-free grasslands (Navall 2008). Elsewhere, the increase in grasses after disturbance has been shown to suppress tree seedling establishment, but also to increase the growth of remaining large trees (Monegi et al. 2022).
In the central arid region of Argentina, shrub encroachment is a consequence of severe overgrazing and logging of woodlands, particularly in the flat region of the province of La Rioja (Dry Chaco) where the economy is now based on extensive cattle and goat farming (Blanco et al. 2005; Calella and Corzo 2006). For several decades, this region was intensely logged, causing woodland degradation (Natenzon and Olivera 1994).
The transformation of rangelands into dense shrublands reduced forage supply and domestic livestock accessibility (Kunst et al. 2012). Although shrubs compete with grasses for resources, mainly water and light, and impede animal movement (Kunst et al. 2003; Blanco et al. 2005), they are fundamental in regulating systems, such as water and carbon cycles (Huxman et al. 2005; Magliano 2016; Nosetto et al. 2020), spatial heterogeneity (Villagra 2000), soil nutrient supply (Anriquez et al. 2005; Pérez Harguindeguy et al. 2022), and habitat preservation for fauna (Szymañski et al. 2021). A significant proportion of shrub species contribute to forage (Allegretti et al. 2012; Egea et al. 2014); so, strategies seeking to maximise livestock production through shrub removal can conflict with those promoting conservation of native vegetation and provision of other ecosystem services. However, new trends in sustainable woodland management aim to integrate both these approaches (Villagra and Alvarez 2019).
In the past two decades, Dry Chaco rangers have sought to increase forage production by removing shrubs mechanically with a roller-chopper, or by hand, and by sowing with exotic pasture species (Marchesini 2011; Rueda et al. 2013; Kunst et al. 2016). Roller-chopping is performed with a heavy, cylindrical machine and pulled by a tractor, which crushes and cuts vegetation (Kunst et al. 2003). It may also distribute seeds of exotic forage species such as Cenchrus ciliaris and Megathyrsus maximus (Jacq.) B.K.Simon & S.W.L.Jacobs (Anriquez et al. 2005; Marchesini 2011). Hand-cutting generally involves greater removal of woody biomass (Kunst et al. 2016), is undertaken with hand-tools such as pick, axe and shovel, and generally removes shrubs at the root. Neither practice is selective, so saplings are removed and few mature specimens retained to provide livestock shade (Nai Bregaglio et al. 2001). Such transitions reduce biodiversity, alter soil nutrient cycles and the rate of litter decomposition, and reduce carbon storage (Villarino et al. 2017; Pérez Harguindeguy et al. 2022; Lizzi and Garbulsky 2023).
In degraded rangelands of central Argentina, regional studies have focused on effects on grass productivity (Blanco et al. 2005; Kunst et al. 2016). Studies of vegetation structural and functional attributes, especially the woody stratum, are scarce (Rejžek et al. 2017). Therefore, our aim was to evaluate two types of shrub removal (mechanical = roller-chopping and hand-cutting) in degraded rangeland on (a) vegetation cover, composition, species richness and diversity, and (b) green biomass and its seasonal dynamics. We also compared these treatments with less disturbed woodlands because they are considered the desired state from a conservation and livestock production viewpoint.
We tested the following predictions: the normalised difference vegetation index (NDVI), as a proxy of green biomass, and vegetation diversity, as characterised by species cover, species richness and the Shannon diversity index, will be lower and have greater seasonal variability on sites with shrub removal than where shrubs have been retained. Furthermore, these effects are expected to be more pronounced under manual removal treatment than with mechanical removal.
Materials and methods
Study area
The study was conducted in the Southern Plains of La Rioja province of Argentina (30°30′S, 66°07′E; Calella and Corzo 2006) within the biogeographic district of the Dry Chaco (Ragonese and Castiglioni 1968; Fig. 1). The weather is monsoonal, with mean annual precipitation of 387 mm (1979–2018). The rainfall is seasonal (82% between November and March), with a high annual variability. Maximum temperatures occur between November and January, and minimums between May and July. The average annual temperature is 20°C (Blanco 2017). The landscape has three distinct vegetation strata (tree, shrub and herbaceous). Tree density is low and species form mixed stands of Aspidosperma quebracho-blanco (Schltdl), Prosopis flexuosa (DC.) and P. torquata (Cav. ex Lag.) DC. The shrub layer is dense, with dominant species including Larrea divaricata (Cav.) and Mimozyganthus carinatus (Griseb.) Burkart and the herbaceous stratum is generally sparse as a consequence of high grazing pressure. The most frequent grass species are Leptochloa crinita (Lag.) P.M. Peterson & N.W.Snow, Gouinia paraguayensis (Kuntze) Parodi, Digitaría califórnica (Benth.) Henrard, and Setaria pampeana Parodi ex Nicora (Biurrun et al. 2015).

|
Study design
Field sampling and remote-sensing analysis was conducted on sites with and without mechanical and manual shrub removal. Sites were distributed in environmentally homogeneous areas according to satellite image analysis. In addition, the region’s less disturbed woodland (‘conserved woodland’), considered a reference ecosystem from a structural and functional point of view (Villagra and Alvarez 2019), was also sampled. These sites were not logged in past decades and are currently grazed with cattle at low stocking rates. A completely randomised design with three treatments and four replicates was established. The treatments were as follows: degraded rangeland with shrub retained (hereafter ‘degraded rangeland’); degraded rangeland with mechanical shrub removal using a roller-chopper (‘roller-chopping’); and degraded rangeland with hand-cutting shrub control (‘hand-cutting’). Shrub-removal treatments were seeded with the exotic species Cenchrus ciliaris variety Texas 4464.
Plots were not sampled prior to the application of treatments. Six years after the shrub-removal treatments had been applied, the modified point-quadrat method, was used to determine the richness and total and specific vegetation cover of trees, shrubs, grasses and forbs (Passera et al. 1983). We randomly located three 50 m line transects at each site, avoiding proximity to wire fences or cattle trails. A 2 m rod placed vertically at 0.5 m intervals along each transect (100 points) was used to assess vegetation interception. At each location and along each line, the composition and the number of species (richness) were recorded and the cover was estimated as the percentage of points intercepted by each species. Measurements were taken in March–April 2019 at the end of the growing season. Using plant species cover data, we calculated the Shannon and Weaver (1949) index using the following formula:
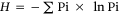
H = Shannon–Weaver diversity index,
Pi = the proportion of the total coverage represented by the coverage of each species, and
ln = log basen
Raunkiaer’s (1934) classification of bioforms was used as a criterion for grouping the recorded species into functional groups and scientific names were corroborated in the Darwinion Botanical Institute database (http://www.darwin.edu.ar/).
We used the NDVI as a proxy of green biomass (Paruelo et al. 1997). NDVI is a direct estimator of the absorbed fraction of photosynthetically active radiation and, indirectly, of aboveground net primary production (ANPP; Di Bella et al. 2004; Piñeiro et al. 2006). We estimated the following seven attributes that characterise aspects of phenology: date of the beginning of the growing season, date of the end of the growing season, duration of the growing season, date of occurrence of the maximum annual NDVI, maximum annual NDVI value, minimum annual NDVI value, annual NDVI integral (ANDVI) and amplitude. We also calculated the coefficient of inter-annual variation of NDVI (CV-NDVI) for each treatment. (Further details are presented in the Supplementary material.)
Data analysis
Analyses of botanical composition, cover, species richness and vegetation diversity among treatments were performed using one-way ANOVA. Model assumptions were explored and data transformations were used to meet these assumptions. The cover data of some species were log-transformed to meet the assumption of normality and homogeneity of variance of residuals. Seasonality parameters extracted from the fitted NDVI curves were compared by repeated-measures ANOVA over time (n = 5 years, 2013–2018), between treatments (1 pixel per 4 replicate × 3 treatment) and their interaction. The relationship between ANDVI and rainfall was analysed by simple Linear regression. In all cases, the significance level was P < 0.05 and Duncan’s test was used as a post hoc test. Data from the conserved woodland were analysed only with descriptive statistics (mean and standard deviation). Analyses were performed with the statistical software InfoStat v. 2018 (Di Rienzo et al. 2018).
Results
Cover, botanical composition and diversity of vegetation functional groups
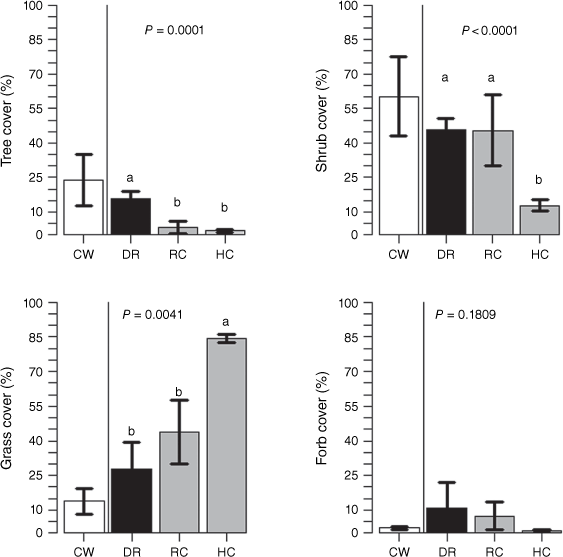
Shrub removal modified the cover of vegetation functional groups (Fig. 2). Tree cover was reduced in the roller-chopping and hand-cutting treatments (P = 0.0001), whereas the opposite was true for grass cover in the hand-cutting treatment (P = 0.0041; Fig. 2). Cover of dominant tree species, such as Prosopis torquata and Prosopis flexuosa, was lower in the cleared treatments than in degraded rangeland (Table 1). Aspidosperma quebracho-blanco, a tree species abundant in conserved woodland, was not recorded in any treatment (Table 1).
|

|
Shrub cover recovered following roller-chopping, but not hand-cutting (P < 0.001). Cover of the dominant shrub species, such as Larrea divaricata, Cordobia argentea (Griseb.) Nied. and Mimozyganthus carinatus in the roller-chopping was similar to that in degraded rangeland. These species were not recorded in the hand-cutting treatment, where dominant woody species were Vachellia aroma (Gillies ex Hook. & Arn.) Seigler & Ebinger and Prosopis flexuosa (Table 1).
Grass cover in the hand-cutting treatment was significantly higher than in degraded rangeland (Fig. 2), with Cenchrus ciliaris being the dominant species. Other dominant species were not recorded in the hand-cutting treatment (Table 1). Forb cover did not change significantly with shrub-removal treatments (P = 0.1809; Fig. 2).
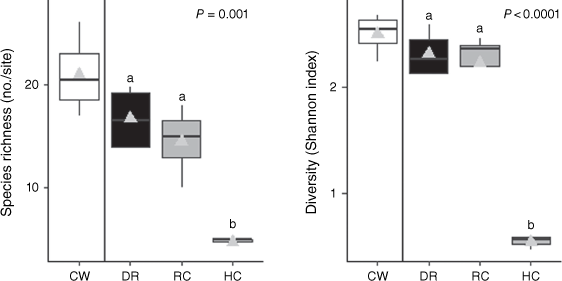
Species richness and diversity were significantly (P = 0.001) higher in conserved woodland and lower in hand-cutting treatment, with degraded rangeland and roller-chopping treatments being indistinguishable from each other at intermediate levels (Fig. 3).
|
Seasonal dynamics of NDVI and phenology
Shrub-removal treatments modified the NDVI (Fig. 4a), and reduced ANDVI and the maximum and minimum NDVI values, most evidently in the hand-cutting treatment (Table 2). CV-NDVI was higher in the roller-chopping and hand-cutting treatments than in the degraded rangeland and conserved woodland (Table 2). The seasonal variation of NDVI was associated with the seasonality of precipitation (Fig. 4b); so, the lowest NDVI values were mostly detected during 2014–2015 and 2017–2018, the least rainy periods during the 5 years of study. The analysis of the relationship between ANDVI and precipitation showed that both conserved woodland and degraded rangeland were more independent of this variable than were the roller-chopping and hand-cutting treatments (Fig. 5).

|

|

|
Discussion
Effects of shrub removal on vegetation structure and composition
Consistent with predictions, shrub removal modified vegetation functional diversity. Dominance changed from tree and shrub species to grasses and shrubs in the roller-chopping treatment, and to one dominated by more than 80% grass cover with hand-cutting. The high regeneration capacity of the shrub stratum and the low regeneration capacity of the tree stratum after the roller-chopping were consistent with results for shrubland sites in central Argentina (Steinaker et al. 2016). Roller-chopping damaged juvenile trees because it was unselective and only large, mature trees were retained (Navall 2008). In addition, the establishment of C. ciliaris probably increased competition with tree seedlings in early stages of natural regeneration (Kunst et al. 2012). Woody removal and grass augmentation can suppress tree seedling establishment (Monegi et al. 2022), and results suggest that a similar situation occurred, with tree seedlings exposed to increased grazing pressure (herbivory and trampling) as a result of intensification of grazing on disturbed areas.
Shrub cover substantially recovered following roller-chopping, but not hand-cutting. Species richness and vegetation diversity drastically declined under hand-cutting. Consequently, only species with seed banks or seedlings formed part of the secondary succession. In the Dry Chaco, basal regrowth is an important mechanism for woody species to form new aerial biomass, and could explain why shrub density was not reduced on roller-chopping compared with hand-cutting, where regeneration depends almost exclusively on the seed bank (Blanco et al. 2005; Marchesini 2011; Bravo et al. 2018). In the transition stage (5 years after shrub removal), shrub cover of species with regrowth capacity, such as L. divaricata, regained and even increased compared with woodland and degraded rangeland. However, when removed by the roots (hand-cutting), L. divaricata was replaced by V. aroma, a spinescent shrub that establishes mainly from seed.
Hand-cutting appeared more efficient than roller-chopping in reducing long-term (10 years) shrub populations. However, hand-cutting is less ecologically sustainable and could affect other essential ecosystem services beyond improving forage supply for livestock (Kunst et al. 2012). Unlike hand-cutting, roller-chopping incorporates substantial plant residues into the soil, increases soil moisture and allows more light to enter lower vegetation strata favouring grasses (Blanco et al. 2005; Kunst et al. 2012). This supports a need for strategies for selective deforestation and conservation management in the Dry Chaco to increase forage production and native woodland regeneration, as well as conservation of other ecosystem services (Boletta et al. 2006; Silberman et al. 2015).
Changes in the dynamics of NDVI (green biomass)
On the basis of the NDVI assessment, sites without shrub removal had a higher green biomass than those with shrub removal, and the conserved woodland had the highest values of green biomass. NDVI considers only the green vegetation fraction; so, by including the non-photosynthetic fraction (branches, woody stems, etc.), the differences in the biomass of these sites would be even greater (Marchesini 2011). ANDVI values observed with roller-chopping and hand-cutting treatments could be a consequence of reduction in functional diversity. Greater functional diversity has been positively related to green biomass, ANPP and carbon sequestration (Díaz and Cabido 2001; Flombaum and Sala 2008). In Dry Chaco woodland, historic tree removal, together with increased shrubs, reduced native grasses and caused ecosystem impoverishment (Marchesini 2011). Trees absorb more radiation and more efficiently convert it to primary production than do many perennial grasses (Blanco 2017). Although replacing native vegetation with grasses allows for a higher harvestable fraction of ANPP, its total value is reduced compared with natural systems, especially native woodland (Rueda et al. 2013). Our results showed that shrub-removal treatments and C. ciliaris seeding maintain total vegetation cover and increase forage supply for livestock, but do not compensate for the loss of green biomass, which could also translate into lower ANPP. In this sense, the manual clearance treatment could maintain a low proportion of woody species and a high proportion of grasses in the long term, although always with a lower green biomass than in natural sites.
NDVI analysis showed that sites with a higher proportion of woody species (conserved woodland and degraded woodland) had higher values of NDVI and a lower CV-NDVI than did the sites that had been roller-chopped and hand-cut. Consistent with the findings of Marchesini (2011) and Steinaker et al. (2016), shrub removal reduced the growth period, ANDVI, and maximum and minimum NDVI, probably because shrubs and trees generally have a longer growing season than do herbaceous species (Blanco 2017). The greater stability in NDVI dynamics found in the conserved woodland and the degraded rangeland may be related to a lower dependence on rainfall (Zerda and Tiedemann 2010). Woody species are generally more independent of rainfall, because they have deep roots with access to water in the lower soil horizons, whereas grasses, with shallow roots, experience growth pulses linked to rainfall and soil moisture in the upper horizons (Schwinning and Sala 2004; Villagra et al. 2011). Thus, the greater the structural difference between native vegetation (conserved woodland or degraded rangeland) and its replaced vegetation, the greater the functional changes in the ecosystem, particularly those linked to carbon and water cycling (Volante et al. 2012; Steinaker et al. 2016; Magliano et al. 2017). Ultimately, a more even distribution of green biomass throughout the year confers greater stability of forage for primary consumers, so that systems dominated by woody species could be playing a key role in ecosystem-service conservation (Volante et al. 2012).
Conclusions
Shrub removal and the seeding of C. ciliaris increased forage available for animal production, but decreased green biomass and modified seasonal dynamics. Grass-dominated systems may be less stable and more dependent on rainfall than are tree-dominated systems, and also provide a different functional diversity because of their structure. Responses also depended on the intensity of shrub-removal disturbance. Roller-chopping is a more conservative practice than is manual bush clearance, because it maintains high levels of vegetation functional diversity and forage production. However, maintaining tree regeneration is important, and because both types of disturbance affect this process, there was no evidence of transitions to desirable conservation states that make environmental conservation (ecosystem services) and livestock production compatible, as might be the case in woodlands. This study has highlighted the importance of designing selective vegetation interventions compatible with the maintenance of functional diversity, tree regeneration and increased forage production.
Supplementary material
Supplementary material is available online.
Data availability
The data that support this study will be shared upon reasonable request to the corresponding author.
Conflicts of interest
The authors declare no conflicts of interest.
Declaration of funding
The current study was funded by the Instituto Nacional de Tecnología Agropecuaria (INTA).
Acknowledgements
We thank the owners of the livestock farms evaluated, as well as all the technicians who participated in the field data collection. Thanks go to Raquel Lothringer for reviewing this paper. We also thank the referees and editors of this paper for their valuable comments.
References
Allegretti, L, Sartor, C, Paez Lama, S, Egea, V, Fucili, M, and Passera, C (2012). Effect of the physiological state of Criollo goats on the botanical composition of their diet in NE Mendoza, Argentina. Small Ruminant Research 103, 152–157.| Effect of the physiological state of Criollo goats on the botanical composition of their diet in NE Mendoza, Argentina.Crossref | GoogleScholarGoogle Scholar |
Anriquez, A, Albanesi, A, Kunst, C, Ledesma, R, López, C, Rodriguez Torresi, A, and Godoy, J (2005). Rolado de fachinales y calidad de suelos en el Chaco Occidental, Argentina. Ciencia del Suelo 23, 145–157.
Archer SR (2010) Rangeland conservation and shrub encroachment: new perspectives on an old problem. In ‘Wild rangelands: conserving wildlife while maintaining livestock in semi‐arid ecosystems’. (Eds JT du Toit, R Rock, JC Deutsch) pp. 53–97. (Wiley-Blackwell Publishing: Oxford, UK)
Barger, NN, Archer, SR, Campbell, JL, Huang, C, Morton, JA, and Knapp, AK (2011). Woody plant proliferation in North American drylands: a synthesis of impacts on ecosystem carbon balance. Journal of Geophysical Research 116, G00K07.
| Woody plant proliferation in North American drylands: a synthesis of impacts on ecosystem carbon balance.Crossref | GoogleScholarGoogle Scholar |
Bestelmeyer, BT, Okin, GS, Duniway, MC, Archer, SR, Sayre, NF, Williamson, JC, and Herrick, JE (2015). Desertification, land use, and the transformation of global drylands. Frontiers in Ecology and the Environment 13, 28–36.
| Desertification, land use, and the transformation of global drylands.Crossref | GoogleScholarGoogle Scholar |
Biurrun FN, Cabido M, Blanco LJ (2015) Consideraciones sobre la vegetación de La Rioja y su estado de conservación. In ‘El deterioro del suelo y del ambiente en la Argentina’. (Eds R Casas, G Albarracín) pp. 485–504. (FECIC-PROSA: CABA, Argentina) [In Spanish]
Blanco LJ (2017) Patrones espaciales y temporales de la productividad primaria neta aérea herbácea y leñosa en el Chaco Árido (Argentina). PhD Thesis, Universidad de Buenos Aires, Buenos Aires, Argentina. [In Spanish]
Blanco, LJ, Ferrando, CA, Biurrun, FN, Orionte, EL, Namur, P, Recalde, DJ, and Berone, GD (2005). Vegetation responses to roller chopping and buffelgrass seeding in Argentina. Rangeland Ecology and Management 58, 219–224.
| Vegetation responses to roller chopping and buffelgrass seeding in Argentina.Crossref | GoogleScholarGoogle Scholar |
Boletta, PE, Ravelo, AC, Planchuelo, AM, and Grilli, M (2006). Assessing deforestation in the Argentine Chaco. Forest Ecology and Management 228, 108–114.
| Assessing deforestation in the Argentine Chaco.Crossref | GoogleScholarGoogle Scholar |
Bravo S, Abdala R, del Corro F, Ibáñez-Moro V, Santacruz-García AC, Loto D, Ojeda F (2018) Regeneración en especies de leñosas nativas del Chaco de Argentina y su respuesta a disturbios. In ‘Los Bosques y el futuro consolidando un vinculo permanente en educacion forestal’. (Eds AM Gimenez, GI Bolzón de Muñiz) pp.127–150. (UNSE-UFPR: Santiago del Estero, Argentina) [In Spanish]
Calella HF, Corzo RR (2006) ‘El Chaco Árido de La Rioja: Vegetación y suelos. Pastizales naturales.’ (Ediciones INTA: Buenos Aires, Argentina) [In Spanish]
Chandregowda, MH, Murthy, K, and Bagchi, S (2018). Woody shrubs increase soil microbial functions and multifunctionality in a tropical semi-arid grazing ecosystem. Journal of Arid Environments 155, 65–72.
| Woody shrubs increase soil microbial functions and multifunctionality in a tropical semi-arid grazing ecosystem.Crossref | GoogleScholarGoogle Scholar |
Di Bella, CM, Paruelo, JM, Becerra, JE, Bacour, C, and Baret, F (2004). Effect of senescent leaves on NDVI-based estimates of fAPAR: experimental and modelling evidences. International Journal of Remote Sensing 25, 5415–5427.
| Effect of senescent leaves on NDVI-based estimates of fAPAR: experimental and modelling evidences.Crossref | GoogleScholarGoogle Scholar |
Di Rienzo JA, Casanoves F, Balzarini MG, González L, Tablada M, Robledo CW (2018) InfoStat version 2018. Centro de transferencia InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. Available at https://www.infostat.com.ar/ [accessed 13 July 2022]
Díaz, S, and Cabido, M (2001). Vive la différence: plant functional diversity matters to ecosystem processes. Trends in Ecology & Evolution 16, 646–655.
| Vive la différence: plant functional diversity matters to ecosystem processes.Crossref | GoogleScholarGoogle Scholar |
Egea, AV, Allegretti, L, Paez Lama, S, Grilli, D, Sartor, C, Fucili, M, Guevara, JC, and Passera, C (2014). Selective behavior of Creole goats in response to the functional heterogeneity of native forage species in the central Monte desert, Argentina. Small Ruminant Research 120, 90–99.
| Selective behavior of Creole goats in response to the functional heterogeneity of native forage species in the central Monte desert, Argentina.Crossref | GoogleScholarGoogle Scholar |
Eldridge, DJ, Bowker, MA, Maestre, FT, Roger, E, Reynolds, JF, and Whitford, WG (2011). Impacts of shrub encroachment on ecosystem structure and functioning: towards a global synthesis. Ecology Letters 14, 709–722.
| Impacts of shrub encroachment on ecosystem structure and functioning: towards a global synthesis.Crossref | GoogleScholarGoogle Scholar |
Flombaum, P, and Sala, OE (2008). Higher effect of plant species diversity on productivity in natural than artificial ecosystems. Proceedings of the National Academy of Sciences 105, 6087–6090.
| Higher effect of plant species diversity on productivity in natural than artificial ecosystems.Crossref | GoogleScholarGoogle Scholar |
Huxman, TE, Wilcox, BP, Breshears, DD, Scott, RL, Snyder, KA, Small, EE, Hultine, K, Pockman, WT, and Jackson, RB (2005). Ecohydrological implications of woody plant encroachment. Ecology 86, 308–319.
| Ecohydrological implications of woody plant encroachment.Crossref | GoogleScholarGoogle Scholar |
Kunst, C, Ledesma, R, Basan Nickish, M, Angella, G, Prieto, D, and Godoy, J (2003). Rolado de fachinales e infiltración de agua en el suelo en el Chaco occidental argentino. Revista de investigaciones Agropecuarias 32, 105–122.
Kunst, C, Ledesma, R, Bravo, S, Albanesi, A, Anriquez, A, van Meer, H, and Godoy, J (2012). Disrupting woody steady states in the Chaco region (Argentina): responses to combined disturbance treatments. Ecological Engineering 42, 42–53.
| Disrupting woody steady states in the Chaco region (Argentina): responses to combined disturbance treatments.Crossref | GoogleScholarGoogle Scholar |
Kunst C, Navall M, Ledesma R, Silberman J, Anríquez A, Coria D, Bravo S, Gomez A, Albanesi A, Grasso D, Dominguez Nuñez JA, Gonzalez A, Tomsic P, Godoy J (2016) Silvopastoral Systems in the Western Chaco Region, Argentina. In ‘Silvopastoral Systems in Southern South America’. (Eds PL Peri, F Dubé, A Varella) pp. 63–87. (Springer International Publishing: Switzerland)
Lizzi, JM, and Garbulsky, MF (2023). Precipitation controls the annual net primary productivity in sown pastures across a precipitation gradient in dry subtropical Chaco. Journal of Arid Environments 208, 104863.
| Precipitation controls the annual net primary productivity in sown pastures across a precipitation gradient in dry subtropical Chaco.Crossref | GoogleScholarGoogle Scholar |
Magliano PN (2016) Dinámica superficial del agua en el Chaco Seco: el papel de las precipitaciones y de la cobertura vegetal. PhD Thesis, Universidad de Bueno Aires, Buenos Aires, Argentina. [In Spanish]
Magliano, PN, Fernández, RJ, Florio, EL, Murray, F, and Jobbágy, EG (2017). Soil physical changes after conversion of woodlands to pastures in dry chaco rangelands (Argentina). Rangeland Ecology & Management 70, 225–229.
| Soil physical changes after conversion of woodlands to pastures in dry chaco rangelands (Argentina).Crossref | GoogleScholarGoogle Scholar |
Marchesini VA (2011) Cambios en el uso de la tierra y el balance de agua en ecosistemas semiáridos: el desmonte selectivo en el Chaco árido analizado a diferentes escalas espaciales. PhD Thesis, Universidad de Buenos Aires, Buenos Aires, Argentina. [In Spanish]
Monegi, P, Mkhize, NR, Tjelele, TJ, Ward, D, and Tsvuura, Z (2022). The impact of tree removal on standing grass biomass, seedling establishment and growth of woody species. The Rangeland Journal 44, 25–32.
| The impact of tree removal on standing grass biomass, seedling establishment and growth of woody species.Crossref | GoogleScholarGoogle Scholar |
Nai Bregaglio, M, Karlin, U, and Coirini, R (2001). Efecto del desmonte selectivo sobre la regeneración de la masa forestal y la producción de pasturas, en el Chaco Árido de la provincia de Córdoba, Argentina. Multequina 010, 17–24.
Natenzon, CE, and Olivera, G (1994). La tala del bosque en los Llanos de La Rioja (1900-1960). Desarrollo Económico 34, 263–284.
| La tala del bosque en los Llanos de La Rioja (1900-1960).Crossref | GoogleScholarGoogle Scholar |
Navall M (2008) Rolados y Manejo Forestal. In ‘Rolado selectivo de baja intensidad’. (Eds C Kunst, R Ledesma, M Navall) pp. 71–85. (Ediciones INTA: Santiago del Estero, Argentina) [In Spanish]
Nosetto, MD, Luna Toledo, E, Magliano, PN, Figuerola, P, Blanco, LJ, and Jobbágy, EG (2020). Contrasting CO2 and water vapour fluxes in dry forest and pasture sites of central Argentina. Ecohydrology 13, e2244.
| Contrasting CO2 and water vapour fluxes in dry forest and pasture sites of central Argentina.Crossref | GoogleScholarGoogle Scholar |
Paruelo, JM, Epstein, HE, Lauenroth, WK, and Burke, IC (1997). ANPP Estimates from NDVI for the Central Grassland Region of the United States. Ecology 78, 953–958.
| ANPP Estimates from NDVI for the Central Grassland Region of the United States.Crossref | GoogleScholarGoogle Scholar |
Passera CB, Dalmaso AD, Borsetto O (1983) Método del Point Quadrat Modificado. In ‘Taller de arbustos forrajeros. Actas del II taller de arbustos forrajeros’. (Eds RJ Candia, RH Braun) pp. 135–152 (FAO-IADIZA: Mendoza, Argentina) [In Spanish]
Pérez Harguindeguy, N, Cingolani, AM, Enrico, L, Vaieretti, MV, Giorgis, MA, Moreno, ML, Falczuk, V, Gurvich, DE, Bertone, GA, Díaz, SM, and Cabido, MR (2022). How human-induced transitions from forest to treeless ecosystems affect litter decomposition. Ecología Austral 32, 716–733.
| How human-induced transitions from forest to treeless ecosystems affect litter decomposition.Crossref | GoogleScholarGoogle Scholar |
Piñeiro, G, Oesterheld, M, and Paruelo, JM (2006). Seasonal variation in aboveground production and radiation-use efficiency of temperate rangelands estimated through remote sensing. Ecosystems 9, 357–373.
| Seasonal variation in aboveground production and radiation-use efficiency of temperate rangelands estimated through remote sensing.Crossref | GoogleScholarGoogle Scholar |
Ragonese, A, and Castiglioni, C (1968). La Vegetación del Parque Chaqueño. Boletín de La Sociedad Argentina de Botánica 11, 133–160.
Raunkiaer C (1934) ‘The life forms of plants and statistical plant geography.’ (Clarendon Press: Oxford, UK)
Rejžek, M, Coria, RD, Kunst, C, Svátek, M, Kvasnica, J, Navall, M, Ledesma, R, Gómez, A, and Matula, R (2017). To chop or not to chop? Tackling shrub encroachment by roller-chopping preserves woody plant diversity and composition in a dry subtropical forest. Forest Ecology and Management 402, 29–36.
| To chop or not to chop? Tackling shrub encroachment by roller-chopping preserves woody plant diversity and composition in a dry subtropical forest.Crossref | GoogleScholarGoogle Scholar |
Rueda, CV, Baldi, G, Verón, SR, and Jobbágy, EG (2013). Apropiación humana de la producción primaria en el Chaco Seco. Ecología Austral 23, 44–54.
| Apropiación humana de la producción primaria en el Chaco Seco.Crossref | GoogleScholarGoogle Scholar |
Schwinning, S, and Sala, OE (2004). Hierarchy of responses to resource pulses in arid and semi-arid ecosystems. Oecologia 141, 211–220.
| Hierarchy of responses to resource pulses in arid and semi-arid ecosystems.Crossref | GoogleScholarGoogle Scholar |
Shannon CE, Weaver W (1949) ‘The mathematical theory of communication.’ (University of Illinois Press: Urbana, IL, USA)
Silberman, JE, Anriquez, AL, Dominguez Nuñez, JA, Kunst, CG, and Albanesi, AS (2015). La cobertura arbórea en un sistema silvopastoril del Chaco y su contribución diferencial al suelo. Ciencia del Suelo 33, 19–29.
Steinaker, DF, Jobbágy, EG, Martini, JP, Arroyo, DN, Pacheco, JL, and Marchesini, VA (2016). Vegetation composition and structure changes following roller-chopping deforestation in central Argentina woodlands. Journal of Arid Environments 133, 19–24.
| Vegetation composition and structure changes following roller-chopping deforestation in central Argentina woodlands.Crossref | GoogleScholarGoogle Scholar |
Szymañski, CR, Tabeni, S, Alvarez, JA, and Campos, CM (2021). Diversity of plants and mammals as indicators of the effects of land management types in woodlands. Forest Ecosystems 8, 74.
| Diversity of plants and mammals as indicators of the effects of land management types in woodlands.Crossref | GoogleScholarGoogle Scholar |
Van Auken, OW (2000). Shrub invasions of North American semiarid grasslands. Annual Review of Ecology and Systematics 31, 197–215.
| Shrub invasions of North American semiarid grasslands.Crossref | GoogleScholarGoogle Scholar |
Villagra, PE (2000). Aspectos ecológicos de los algarrobales Argentinos. Multequina 9, 35–51.
Villagra, PE, and Alvarez, JA (2019). Determinantes ambientales y desafíos para el ordenamiento forestal sustentable en los algarrobales del Monte, Argentina. Ecología Austral 29, 146–155.
| Determinantes ambientales y desafíos para el ordenamiento forestal sustentable en los algarrobales del Monte, Argentina.Crossref | GoogleScholarGoogle Scholar |
Villagra, PE, Giordano, C, Alvarez, JA, Cavagnaro, JB, Guevara, A, Sartor, C, Passera, CB, and Greco, S (2011). Ser planta en el desierto: estrategias de uso de agua y resistencia al estrés hídrico en el Monte Central de Argentina. Ecología Austral 21, 29–42.
Villarino, SH, Studdert, GA, Baldassini, P, Cendoya, MG, Ciuffoli, L, Mastrángelo, M, and Piñeiro, G (2017). Deforestation impacts on soil organic carbon stocks in the Semiarid Chaco Region, Argentina. Science of the Total Environment 575, 1056–1065.
| Deforestation impacts on soil organic carbon stocks in the Semiarid Chaco Region, Argentina.Crossref | GoogleScholarGoogle Scholar |
Volante, JN, Alcaraz-Segura, D, Mosciaro, MJ, Viglizzo, EF, and Paruelo, JM (2012). Ecosystem functional changes associated with land clearing in NW Argentina. Agriculture, Ecosystems & Environment 154, 12–22.
| Ecosystem functional changes associated with land clearing in NW Argentina.Crossref | GoogleScholarGoogle Scholar |
Willcox, EV, and Giuliano, WM (2010). Seasonal effects of prescribed burning and roller chopping on saw palmetto in flatwoods. Forest Ecology and Management 259, 1580–1585.
| Seasonal effects of prescribed burning and roller chopping on saw palmetto in flatwoods.Crossref | GoogleScholarGoogle Scholar |
Zerda, HR, and Tiedemann, JL (2010). Dinámica temporal del NDVI del bosque y pastizal natural en el Chaco de la Provincia de Santiago del Estero, Argentina. Ambiência 6, 13–24.