

Preliminary assessment of gidgee (Acacia cambagei) woodland thickening in the Longreach district, Queensland
R. J. Fensham A B and R. J. Fairfax AA Queensland Herbarium, Environmental Protection Agency, Mt Coot-tha Road, Toowong, Qld 4066, Australia.
B Corresponding author. Email: rod.fensham@epa.qld.gov.au
The Rangeland Journal 27(2) 159-168 https://doi.org/10.1071/RJ05013
Submitted: 3 February 2005 Accepted: 10 June 2005 Published: 21 November 2005
Abstract
Gidgee (Acacia cambagei) forms woodlands and forests on moderately fertile clay soils throughout semi-arid areas of north-eastern Australia. Analysis of soil data from Queensland indicates that gidgee’s environmental domain has a broad overlap with treeless Mitchell grassland although a preference for slightly coarser soils with higher phosphorus and lower potassium content. Mapping of woody cover change for an area of central Queensland suggests that gidgee thickening and spread has occurred between 1951 and 1994. However, this increase has been more than offset by loss of gidgee cover by clearing in other areas. Gidgee thickening and encroachment onto grassland is a problem for graziers because the trees suppress grass and are unpalatable except when very young. Landholder surveys and other information indicate that gidgee has recruited sporadically and that thickening occurred with a massive seedling recruitment in the 1950s with a smaller recruitment wave in the 1970s. These waves coincide with relatively wet times and the extreme rainfall abundance in the 1950s has a multi-century expected recurrence. There are historical records describing gidgee death during the extreme droughts in the first part of the 20th century. Young gidgee plants are fire sensitive but well-established gidgee stands are fire retardant, and the importance of fire for control of gidgee thickening is unknown. One scenario explaining gidgee dynamics emphasises the role of environment (mostly climate and soil), while another emphasises management (mostly fire). Research priorities are presented and may contribute to appropriate management of gidgee woodlands.
Additional keywords: drought, fire, grassland encroachment, vegetation thickening, woodland dynamics.
Introduction
Gidgee (Acacia cambagei1) grows on moderately fertile clay soils and is in many ways an ecological analogue of its closest botanical relative brigalow (Acacia harpophylla) from less arid climates. Gidgee and brigalow dominated similar areas of woodlands and forests throughout Queensland (about 90 000 km2) but the brigalow has been far more extensively cleared for pastoral development. Only about 11% of brigalow forests and woodlands remained in the brigalow belt biogeographic region in 1999, while at the same time gidgee forests and woodlands were about 72% intact in the Mitchell grass downs biogeographic region (Wilson et al. 2002).
A survey of graziers in western Queensland (Reynolds and Carter 1993) revealed that gidgee rather than exotic species such as prickly acacia (Acacia nilotica) and parkinsonia (Parkinsonia aculeata) was the tree causing greatest problems for stock management. Concerns include general thickening of the gidgee woodlands (i.e. increased density of trees) and encroachment of gidgee woodland into surrounding Mitchell grassland. The advance of gidgee comes at the expense of pasture production and there is a general desire to use mechanical clearance as a means of controlling gidgee. Clearing is a contentious issue and recently established legislation in Queensland (Vegetation Management and Other Legislation Amendment Act 2004) seeks to phase out broadscale clearing of remnant vegetation by 2006 (NRM 2004).
Many land managers believe that increases in gidgee density are primarily a result of lack of fire in a grazed landscape. If current land use practices are increasing the density of gidgee, then it could be argued that management through mechanical clearance provides an option for redressing the imbalance. However, if changes in gidgee density are part of the regeneration ecology of the species and the ecological limits are clearly set by soils and climatic conditions, the premise for seeking to clear gidgee is different. That is, instead of clearing to restore ecological function, clearing is being undertaken for production benefit in spite of ecological function.
This study was initiated to prepare ecological and climatic background information on the issue of structural change in gidgee woodlands, with particular emphasis on those in the Longreach area of central Queensland. The landscape setting for gidgee woodlands is reviewed and analyses of soil data provided to examine inherent differences between the soil of gidgee woodlands and adjoining Mitchell grasslands; time series mapping of gidgee woodlands and adjoining Mitchell grasslands from aerial photography is summarised; rainfall history data are analysed in relation to known gidgee recruitment events; and other aspects and observations of the ecology of gidgee woodlands provided. Two opposing scenarios both largely consistent with the available evidence are erected to explain dynamics in gidgee woodlands and recommendations for future research are provided.
Materials and methods
What is gidgee?
Gidgee is a small tree to 12 m tall. It is a species of the wattle (Acacia) genus, within a group of related species that includes A. ammophila, blackwood (A. argyrodendron), myall-gidgee (A. calcicola), georgina gidgee (A. georginae), brigalow (Acacia harpophylla), A. latzii, yarran (A. melvillei, A. omalophylla), A. machonochieana, wommel (A. maranoensis), A. microcephala, Bowyakka (A. microsperma), myall (A. pendula) and boree (A. cana, A. tephrina). The common members of this group are mostly restricted to semi-arid north-eastern Australia (excluding tropical Cape York Peninsula) and generally occupy fertile clay soils. The preference for relatively fertile soils sets these Acacia species apart from others that are common on the infertile sands and loams typical of most of the Australian continent. A stand of gidgee can be almost mono-specific or include other tree species, including boree, brigalow, false sandalwood (Eremophila mitchellii), leopardwood (Flindersia maculosa) and whitewood (Atalaya hemiglauca).
Landscape setting and soils
Landscape settings for gidgee woodland typically include landforms where generally coarse-grained highly weathered material originating from Tertiary sandstone occurs in combination with material that has weathered from fine-grained shales of Cretaceous origin. Pediments formed on Cretaceous shales that are partially mantled by residual debris of the stripped Tertiary surface and alluvium from the mixing of the Tertiary and Cretaceous source materials are examples (Turner et al. 1978, 1993; Mills et al. 1980). In the Longreach area gidgee woodland is typically associated with an intermediate position between the Mitchell grassland on the water sheds and the active alluvial areas (Fig. 1).

|
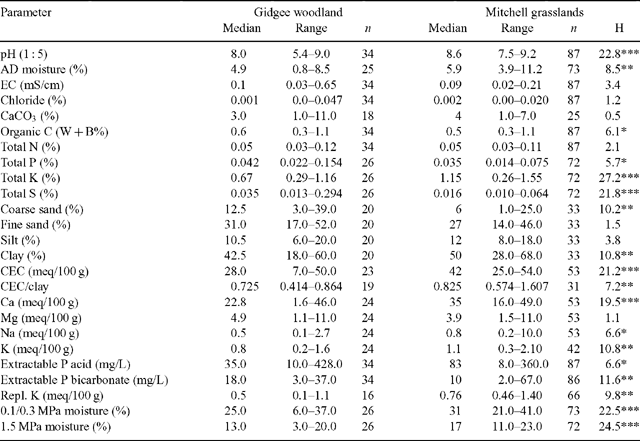
The soil data from WARLUS II (Mills et al. 1980), IV (Turner et al. 1978) and V (Turner et al. 1993) were interrogated to analyse differences between Mitchell grasslands and gidgee woodlands in central Queensland (142–147°E; 21–26°S). Sites on sedimentary substrates from Mitchell grassland land units (WARLUS II: 40, 50, 51, 55, 58, 64, 65, 67; WARLUS IV: 1, 2; WARLUS V: 1, 2, 3, 4) and gidgee woodland land units (WARLUS II: 38, 39, 40, 42, 50, 51, 59; WARLUS IV: 15, 16, 17; WARLUS V: 8, 9, 10, 57) were selected. The surface soil data from 87 Mitchell grassland sites and 34 gidgee woodland sites were compiled (Table 1). Differences in soil characteristics between gidgee woodland and Mitchell grassland were examined using Mann–Whitney U-tests and chi-squared for the soil colour categories red-yellow and brown-grey.
|
In general both gidgee woodlands and Mitchell grasslands occur on clay-rich soils, although the former have higher sand content, typically contain gravel and lower clay content. Of the gidgee woodland soils, 35% are red-yellow and 65% brown-grey, while for the Mitchell grassland soils 2% are red-yellow and 98% brown-grey (χ2 = 23.0, d.f. = 1, P < 0.001). Surface soils under gidgee woodlands are generally less alkaline, and have significantly more coarse sand and less clay than Mitchell grassland soils (Table 1). The cation exchange capacity/clay ratio is lower for gidgee woodland surface soils than Mitchell grasslands indicating a prevalence of clays with reduced concentrations of expanding minerals. The montmorillinitic clays with high proportions of expanding minerals have a tendency for considerable lateral and vertical movement that results in deep cracking on drying and tends to favour grassland. The physical and possibly chemical properties of these soils may disfavour tree establishment. During droughts gidgee soils have more available moisture as indicated by significantly lower 1.5 MPa moisture contents (Table 1).
Total phosphorus and sulfur are higher in gidgee woodlands than Mitchell grasslands and potassium and calcium show the reverse trend (Table 1). There are many exceptions to these generalisations as indicated by the overlapping ranges of all measured soil characteristics (Table 1). In the Longreach area gidgee woodland and Mitchell grasslands generally have discrete landscape settings, although there are many apparent exceptions and targeted geomorphological and soils studies are required before these relations are clearly defined.
Change in gidgee structure
Mapping methodology. The study area is defined as the area of alluvia and fine grained sedimentary substrates within the Mitchell grass downs bioregion on the Longreach 1:250 000 map sheet (23–24°S; 144–145°30’ E) as determined from the mapping data of the Queensland Herbarium (Wilson et al. 2002). The sampling strategy employed the 1 km national mapping grid as the sampling unit. Cells that have more than half of their area within the study area are included. For each km2 cell, woody cover (%) was assessed from 1951 black and white (1:50 000) and 1994 colour (1:80 000) aerial photography by an experienced interpreter (R. Fairfax) with reference to pre-established black and white patterns of known cover. The cover assessments were established as a GIS coverage and used in conjunction with existing Queensland Herbarium regional ecosystem (Wilson et al. 2002) and remnant mapping coverages to perform a range of analyses quantifying change. The regional ecosystem coverage was collapsed to the km2 cell unit of this study, thus, where cells contained more than one regional ecosystem, each cell was assigned the most prevalent regional ecosystem. Crown cover will include much vegetation that is not gidgee, including a predominance of Coolibah (Eucalyptus coolabah) in regularly flooded areas and Boree (Acacia tephrina) in some other areas. The analysis includes only those 1-km2 cells that have greater than 5% crown cover in either the 1951 or 1994 photography. Data were assessed with and without clearing (1995 remnant vegetation coverage, Queensland Herbarium). The Vegetation Management Act 1999 (Queensland) defines remnant vegetation as having at least 70% of the height and 50% of the cover of the dominant stratum, relative to the undisturbed height and cover of that stratum and dominated by species characteristic of the vegetation’s undisturbed canopy (Wilson et al. 2002). Thus some areas that were cleared between 1951 and 1994 have sufficiently recovered their pre-clearing structure to be classified as remnant. The 1995 remnant coverage was used because it is closest to the 1994 aerial photography.
Calibration of aerial photography with field measurements. Fensham et al. (2002) have demonstrated that aerial cover is exaggerated as photo-scale decreases. If this effect applies to the photo-signatures of gidgee woodland this will serve to exaggerate thickening in this study given the scale difference between the initial (1:50 000) and final (1:80 000) photography. Thus there is a need to apply some field-based calibration to aerial photography of these scales within the gidgee-grassland land types to improve the accuracy of interpretation.
Two areas containing Acacia and Eucalyptus land types were captured by colour aerial photography including scales of 1:50 000 and 1:80 000 in June 2004. Each area was approximately 150 km2, one south of Barcaldine and the other south of Pentland. From these areas 22 sites from Acacia land types and 37 from Eucalyptus land types were selected for calibrating aerial cover with field cover. Each site had homogenous photo-pattern over at least 10 ha. Field survey was conducted the following month and for each site the height or absence of canopy cover (interpreted as a convex solid polygon around the live branches of a woody plant) estimated every 5 m along eight transects (each 50 m long), spaced evenly throughout the target area. Percent field woody cover was obtained from these 88 points.
Percent woody cover of each site was also visually estimated from both scales of aerial photography using the process used for mapping gidgee cover over the Longreach study area. Estimates were duplicated to represent variation in assessment and the average correlated with field woody cover using generalised linear models with a Poisson error distribution. Exploratory analysis indicated that land type was not an important explanatory variable so a combined model was employed of the form:
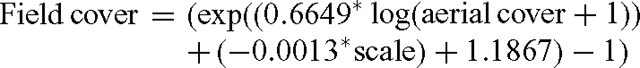
with scale represented by two digits (i.e. 1 : 80 000 as 80).
Comparison of predicted and actual field measurements using this model yielded R2 values for the Acacia land type sites of 0.87 for 1:50 000 assessments and 0.85 for 1:80 000 assessments.
The above equation was applied to the cover values estimated for the mapping.
Results
Mapping results
The study area comprised 16 292 km2 of which 85% consists of fine-grained Cretaceous sediments and ancient alluvium and 15% recent alluvium within active stream channels.
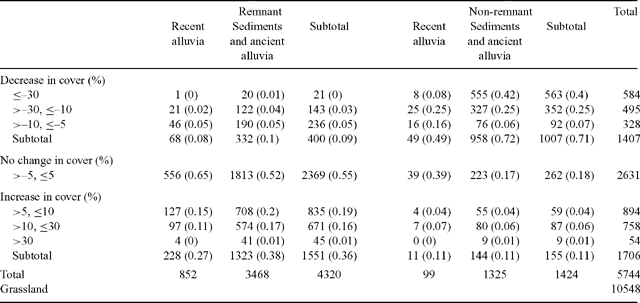
The data in Table 2 indicate that 10 548 km2 of grassland had less than 5% crown cover at both sampling times. For the 1424 km2 of wooded cells mapped as non-remnant (i.e. mechanically cleared), 71% decreased in cover, 18% exhibited little change (≤5% absolute cover change in either direction) and 11% increased in cover (Table 2). For the remainder included as remnant with more than 5% crown cover at either sampling time, 2369 km2 (55%) exhibited very little change; 400 km2 (9%) had reduced cover (>5%), and 1551 km2 (36%) had increased cover (>5%)(Table 2). Some areas mapped as having undergone cover decline may have been cleared at some time between 1951 and 1994, but have regrown sufficiently to be mapped in 1995 as remnant vegetation. The trend to increased cover is greater on sediments and ancient alluvia relative to recent alluvia (Table 2).
|
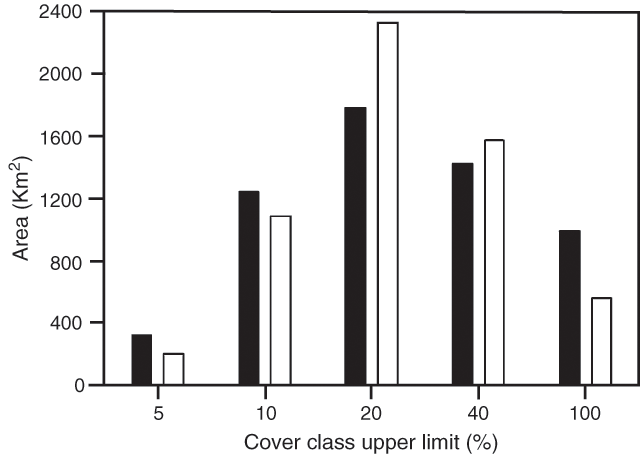
The average cover of all wooded cells (>5% at either sampling time) in 1951 was 21.9% and in 1994 was 19.1%. This indicates that overall clearing and the relatively rare instances of self-thinning have resulted in a greater loss of gidgee cover than increases resulting from thickening and encroachment. The overall effects of the changes by cover class are indicated in Fig. 2.
|
At the coarse scale of our mapping we are not able to separate the encroachment of gidgee on to grassland and thickening of gidgee woodland. Our impressions from conducting the mapping is that the vast majority of the increased cover of gidgee woodland is a result of the thickening of existing woodland, but more detailed mapping is required to quantify the extent of these different phenomena.
Causes of change in gidgee structure
Grazier survey. Long-term graziers with sizeable areas of remnant gidgee woodland in the Longreach district were interviewed in April 2003 in order to canvass observations on gidgee ecology and dynamics. Seventeen graziers were interviewed in relation to 38 gidgee paddocks on 19 properties. The interviews confirmed that there had been little mechanical disturbance within the remnant gidgee woodland included in the survey, including no broad-scale ringbarking or tree poisoning, or mechanical clearance. Respondents indicated that only three of 38 gidgee paddocks had been extensively cut to provide fuel for steam engines (2 sites before the 1960s) and generally only limited areas had been cut for fence-posts. Cutting of fence posts and firewood may have been more substantial in the early part of the twentieth century.
Gidgee structural change and rainfall. For areas where gidgee has increased most graziers were able to identify cohorts of regeneration. The most common cohort described was the wave of regeneration that occurred during the successive wet years of the 1950s. Davidson (1954) verifies this regeneration event,
‘Since the heavy rain of 1950 there has been an amazing regeneration of gidgee. Seedlings appeared in enormous numbers—sometimes 60 per square yard. They were mostly very close to parent trees and many have since died’.
Some graziers also identified a cohort that established (Fig. 3) after the extreme wet months early in 1974 (Fig. 4). One grazier also identified some trees that established in the early 1990s. Reynolds et al. (1992) concluded that there had been no major recruitment of gidgee on their study sites in the Longreach–Blackall area at least since 1983.

|
|
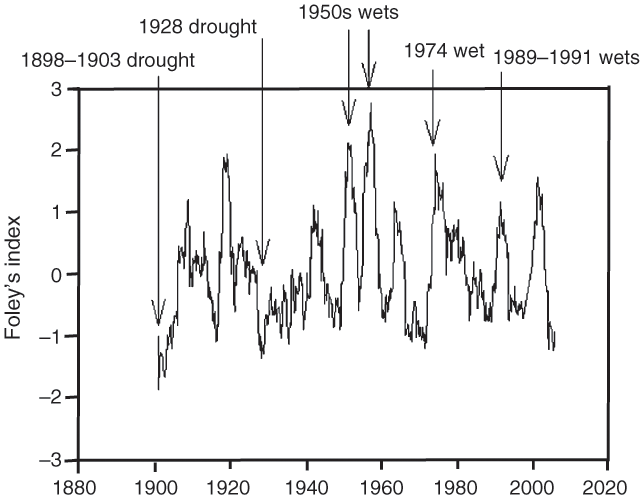
The rainfall data for Ilfracombe (central to the study area) provides some indications of the conditions required for gidgee establishment (Fig. 4). For this analysis we have used a 3-year Foleys index, which is computed as 3-year running totals of standardised variations in annual rainfall (actual annual rainfall less mean annual rainfall divided by the mean annual rainfall) prior to and including each month (i.e. for September 1970 the running total is calculated from October 1967–September 1970) (Foley 1957). This provides a measure of sustained wet and dry periods, and has proved useful for relating rainfall patterns to tree dynamics in other studies (Fensham and Holman 1999).
Gidgee seed germinates rapidly after seed-fall and does not accumulate in the seed-store (Reynolds et al. 1992). Seed-set is sporadic. One grazier indicated that from his observations seeding occurred after abundant spring rain. Establishment occurs only in wet years. Thus, it is likely that sequences of wet years are required for successful gidgee recruitment events. These prolonged wet times need to be sufficient to trigger mass flowering and seeding and then allow for seedling survival and establishment.
The trend in the 3-year Foleys index indicates the significance of the wet periods in the 1950s (Fig. 4). Twice during the 1950s decade, Foley’s index had values over +2 and a maximum value of +2.7 in 1956. This deviation from average is far greater than any other in recorded history in either negative or positive direction indicating that this may have been an extremely rare event (i.e. multi-century scale). The 1974 wet year was a less extreme event (peak Foley’s index value +1.9) and equally as wet as a previous event ending about 1919 (Fig. 4).
In the areas mapped as stable, open-structured gidgee woodlands with large trees prevail. The open understories contain few grasses, but have sufficient herbage (mostly chenopods) to provide useful forage for sheep. In areas that have thickened long-term graziers remember a pre-1950s structure akin to the open gidgee woodlands and large dead trees are testament to their former structure (Fig. 3). The dense regenerating gidgee forms multi-stemmed low trees (‘whipstick gidgee’) dating from the regeneration events primarily of the 1950s, to a lesser extent the 1970s and rarely from the 1990s. Herbage is virtually non-existent and the thickened areas are virtually worthless for grazing.
The worst drought on record was between 1898 and 1903 where Foley’s index fell to –1.7 (Fig. 4). There are historical records of this drought causing extensive tree dieback including gidgee (Fensham and Holman 1999):
‘Ever since the black soil downs of the far West have been occupied the question as to what cause brought about the large areas of dead timber to be seen in the mulga, gydia, and boree scrubs of the interior has been a matter of controversy amongst bushmen. Some attributed it to the action of fire, others to a cold snap, possibly accompanied by snow; again others to a plague of caterpillars, which ate all the leaves and so destroyed the trees: but seeing that timber is now dying in all districts of western Queensland, it would seem not unreasonable to conclude that drought was the cause of thousands of square miles of country in the ‘Never never’ being denuded of scrub.’ (Anon. 1902).
Another less severe drought culminated in 1928 (minimum Foley’s index –1.4), with the drought conditions persisting into the 1930s (Fig. 4). Blake (1938) records that large areas of dead gidgee were evident in parts of the central west after this period. The two droughts prior to 1930 were both more severe than any of the droughts that have followed (Fig. 4).
Foley’s index at Ilfracombe in March 2005 dropped to –1.24, the lowest value since 1928 (Fig. 5). During this recent period large patches of thickened gidgee have died on at least one property in the Longreach district (Fig. 5) and drought would seem to be the likely cause.

|
Fire. No graziers confirmed any fire within the gidgee paddocks since the 1970s. One grazier thought that parts of one paddock could have been burnt from the railway line in the late 1960s. A second grazier remembers a fire in the late 1950s that burnt through the crowns of boree trees but did not have a specific memory of fire in any of the gidgee paddocks. A third grazier remembers a hot fire burning some gidgee areas in one paddock around 1956. Fire scars are evident on the 1951 aerial photography, although these relatively small fires were restricted to grassland and clearly did not burn into gidgee woodlands. In summary fire has been extremely rare throughout the district over the last 50 years at least. Most graziers were of the view that fire would not carry through gidgee woodlands because of the suppressive effect of gidgee trees on understorey fuel loads.
There is quantified evidence that gidgee is fire sensitive (Butler and Fairfax 2003), although there was no direct confirmation of this from the land-holders interviewed because of the rarity of fire events. However, all graziers agree that the dense whipstick gidgee that currently dominates much of the Longreach district cannot be thinned using fire because the absence of ground fuel means that the areas simply could not carry a fire, even under extreme conditions.
Grazing. Sheep grazing has been the dominant land-use over the study area through the period of land settlement. Only one paddock included in the survey had a long history of cattle grazing and that was a paddock that had been used for milking cows since 1937. Cattle have exclusively grazed another paddock since 1990. Quoted stocking rates vary between 1 sheep to 3 acres to 1 sheep to 8 acres. Destocking generally occurs only intermittently.
All graziers agreed that sheep do not eat live adult gidgee foliage. However, several graziers had directly witnessed sheep eating young gidgee seedlings. Davidson (1954) verifies this within the study area.
‘In paddocks that are constantly stocked, the leaves [of seedlings] have been stripped off by the sheep and there are comparatively few seedlings left. Where the country has been unstocked for considerable periods the seedlings are now a foot or two high and are still present in large numbers.’
One grazier was of the view that the considerable gidgee advance on his property may be partly attributed to his father’s conservative stocking after the regeneration event of the 1950s that provided the opportunity for the gidgee to get away. The role of sheep in eliminating gidgee regeneration was occasionally evident in situations where fence-lines contrasted open structure gidgee woodland with dense whipstick gidgee. The open areas are recognised as places favoured by sheep (see Fig. 6).
|
Hail damage. Two graziers verified that hail storms can kill gidgee trees. One of these graziers has witnessed gidgee death on at least two occasions over the last 50 years and the other witnessed the death of several hundred hectares of gidgee following a recent hail storm.
Synthesis
The mapping identified substantial gidgee thickening in the Longreach area over the last 50 years. Some areas have been stable, but there are very few areas that have thinned without mechanical clearance. The amount of gidgee clearing in the same area has been substantial and exceeds woody cover increases. Large areas of gidgee thickening on single properties are a major impost for management and are not reversible by burning because of the lack of fuels within the dense stands. Establishment events seem to occur about 1–4 times a century, although the massive establishment event(s) of the 1950s may be a multi-century phenomenon.
Two divergent scenarios could potentially describe gidgee dynamics and are broadly consistent with the background evidence presented here. The first scenario emphasises the influence of management effects. In the pre-pastoral landscape extensive fires fuelled by large bodies of grass fuel eliminated most of the regenerating gidgee that established after occasional periods of rainfall excess, limiting gidgee encroachment onto grasslands and maintaining an open structure in gidgee woodlands. With pastoral management burning is infrequent not only because of the general lack of fuel but also because burning is discouraged because of the limited means of control and the potential for lost forage. Fires have become extremely rare events. Even if fire were reintroduced, the reduction in fuel loads brought about by sheep grazing would ensure that ground fuels would not be sufficient to support the fire regimes required to eliminate gidgee after the recruitment events. Without fire in the landscape gidgee will eventually invade Mitchell grasslands where soil type differences are not sufficient to limit spread. Sheep grazing can eliminate recruiting gidgee if applied in sufficient densities. However, in the bountiful seasons that trigger gidgee establishment, sheep grazing is insufficient to effectively eliminate the massive recruitment of gidgee. Pastoral management has resulted in thickening and encroachment of gidgee that would not have occurred in the pre-pastoral landscape. The system is ‘out of balance’ and the cost is reduced herbaceous forage over large areas that were formerly productive. Mechanical clearance is the only practical way to restore the balance.
The second scenario emphasises the importance of climate and soils. Gidgee is generally limited by soil structural characteristics, although in favourable times it can spread on to the heavier grassland clay soils. Extreme drought will eliminate gidgee from these unfavourable sites where the clays restrict rooting depth, have extremely low available moisture during drought, and undergo shrinking and cracking unfavourable to woody plants. Gidgee woodlands are favoured on lighter textured soils than those that support Mitchell grassland. Within these woodlands, the cycle of gidgee regeneration can be described, starting with big old trees forming an open-structured woodland. During occasional wet events gidgee regenerates en masse. The gidgee regeneration rapidly precludes adequate fuels for burning under and beyond its canopy. Fire frequency and coverage are insufficient to eliminate the gidgee recruitment during the narrow window of opportunity before the stands of young gidgee trees become fire resistant. The recruiting gidgee trees develop into a dense stand beneath the original old gidgee trees. The competitive effect of the dense gidgee eventually kills the preceding generation of old trees through competition for moisture. Gradually as the stand continues to develop, self-thinning occurs until the structure eventually resembles the original open woodland. The self-thinning is accelerated during extreme droughts that cause substantial gidgee dieback. The cycle takes place over about a century or more. Mechanical thinning effectively shortens the ‘thickened’ periods and prolongs the more open periods.
These scenarios are not mutually exclusive and there are probably at least elements of truth in both. There may also be related but different processes affecting gidgee encroachment than thickening. For example, there may be a positive feedback where the invasion of grassland during wet seasons renders soils more hospitable for tree growth. Eldridge and Freudenberger (2005) have demonstrated that in fine-textured soils supporting eucalypts the presence of trees increase macropore space, which increases sorptivity (the early phase of wetting up a dry soil) and steady-state infiltration (water passing through a wet soil). If gidgee trees that have pioneered grassland invasion modify their environments in this way then the presence of trees may sufficiently improve soil characteristics to increase the likelihood of persistence through drought. Deciphering the ecology of gidgee provides an exciting research opportunity of great relevance to contemporary land management, vegetation management policy and biodiversity conservation.
Recommendations for future research
Functional models describing the causal mechanisms of structural change have been informed by studies relating management attributes to vegetation change (Roques et al. 2001; Asner et al. 2003; Fensham et al. 2005). The evidence from the Longreach district would suggest that such studies are not likely to be fruitful in relation to dynamics in gidgee woodlands. Examples of burning throughout much of gidgee’s range do not exist. Grazing history information is not available for the rare gidgee germination events in the 1950s and 1970s. Furthermore because of vegetation patterns, availability of water and sheep behaviour there is temporal and spatial variation in sheep grazing intensity within a paddock.
Land survey property plans have been successfully utilised to assess structural change in other areas of Australian rangeland (Lunt 1997; Fensham and Holman 1998). Land survey plans from the Longreach area dating from about 1890 to 1920 include descriptions of vegetation structure and composition. These plans locate areas described variously as ‘very thickly timbered’, ‘dense gidya scrub’, ‘open with clumps of gidya’ and ‘open gidya scrub’. These records require more detailed assessment but could be compared with the aerial photography evidence to provide a century-scale assessment of structural change.
More detailed mapping of woodland-grassland boundaries from aerial photography would also distinguish between gidgee thickening and encroachment on to Mitchell grassland. These studies should be extended to fieldwork designed to further identify the association of gidgee woodland and Mitchell grassland with soils and landforms and to address the hypothesis that gidgee encroachment may be limited by soil type. The coincidence of recent drought and areas of dead gidgee may provide opportunities to study the effects of moisture stress on gidgee thickening and encroachment.
Burning trials are required to further our understanding of gidgee fire ecology. There has been only limited fire research on gidgee and efforts to date (Butler and Fairfax 2003) have not examined fire intensity or size class interactions. Three National Parks in western Queensland (Moorinya, Forest Den and Bladensburg) may provide suitable sites to conduct such trials. On these conservation reserves domestic stock grazing does not reduce fuels and the opportunities to trial hot burns may be available.
Fires would have to be extensive to eliminate gidgee seedlings during the narrow window of opportunity before the woodlands develop fire retardance. The FARSITE model (Finney 1998) is an example of a cellular model that predicts the spread and extent of fires using parameters such as fuel type, terrain and weather conditions. Fires probably extinguish during certain weather conditions (i.e. heavy dew) and at topographic barriers such as major watercourses. Rates of ignition in the pre-pastoral landscape could be developed from lightning incidence and probable aboriginal burning patterns (see Fensham 1997). Spatial modelling of fire spread using informed parameters could address the question: What proportion of gidgee recruitment would be killed by fire in a 5-year period after a recruitment event in a pre-pastoral landscape?
Tropical grasses (C4 plants) and trees (C3 plants) have different photosynthetic pathways and their plant material yields different proportions of 2 of the stable isotopes of carbon. The relative proportions of these carbon isotopes (13C and 12C) in soil carbon is the δ13C signature, which has been used to interpret changes in the proportions of grass and trees at a particular site over time. Soil carbon processes are complex, and the technique can yield results that are difficult to interpret. However, in many landscapes it represents the only technique available that may provide insights into past vegetation patterns before the photographic and written record. Krull et al. (2005) have analysed soil organic matter for δ13C along a gradient from thickened gidgee woodland, within invaded grassland, and in open Mitchell grassland near Longreach. The data show that the gidgee woodlands have δ13C carbon signatures that are consistent with the thickening of those woodlands since the gidgee recruitment events of the 1950s. Interpreting changes that might have occurred at the site in the more distant past becomes increasingly difficult. However, multiple sampling of the thickened gidgee woodlands throughout the region may yield consistent results that could be more clearly interpreted and may provide signals indicating whether or not the thickening of gidgee has occurred in the past.
Acknowledgments
This project was partly funded by the Tropical Savannas Management CRC and the support of Jill Landsberg and Gordon Duff is greatly appreciated. The Longreach Office of the EPA is thanked for logistic support during the questionnaire survey, and in particular David Akers and Vanessa Bailey. Don Butler and Rosemary Niehus provided GIS and analytical support. Evelyn Krull took the time to discuss the complex processes behind her carbon isotope study. The enthusiasm, time and hospitality of all the graziers involved is greatly appreciated. Don Alexander is especially thanked for alerting me to the drought-killed gidgee on ‘Willoughby’ after the initial grazier survey had been completed. The reviewer’s and editor’s comments were useful for improving the manuscript.
Anon.
(1902) The drought. The Queenslander 24 May, 1168.

Asner G. P.,
Archer S.,
Hughes R. F.,
Ansley R. J., Wessman C. A.
(2003) Net changes in regional woody vegetation cover and carbon storage in Texas drylands, 1937–1999. Global Change Biology 9, 316–355.
| Crossref | GoogleScholarGoogle Scholar |
Blake S. T.
(1938) The plant communities of western Queensland and their relationships, with special reference to the grazing industry. Proceedings of the Royal Society of Queensland 49, 156–205.
Butler D. W., Fairfax R. J.
(2003) Buffel grass and fire in a gidgee and brigalow woodland: a case study from central Queensland. Ecological Management and Restoration 4, 120–125.
| Crossref | GoogleScholarGoogle Scholar |
Davidson D.
(1954) The Mitchell grass associations of the Longreach district with notes of gidgee and coolibah communities. University of Queensland Department of Botany Occasional Papers 3, 45–59.
Eldridge D. J., Freudenberger D.
(2005) Ecosystem wicks: woodland trees enhance water infiltration in a fragmented agricultural landscape in eastern Australia. Austral Ecology 30, 336–347.
| Crossref | GoogleScholarGoogle Scholar |
Fensham R. J.
(1997) Aboriginal fire regimes in Queensland, Australia: analysis of the explorers’ record. Journal of Biogeography 24, 11–22.
| Crossref | GoogleScholarGoogle Scholar |
Fensham R. J.,
Fairfax R. J., Archer S.
(2005) Rainfall, land-use and woody vegetation cover change in semi-arid Australian savanna. Journal of Ecology 93, 596–606.
| Crossref | GoogleScholarGoogle Scholar |
Fensham R. J.,
Fairfax R. J.,
Holman J. E., Whitehead P. D.
(2002) Quantitative assessment of vegetation structural attributes from aerial photography. International Journal of Remote Sensing 23, 2293–2317.
| Crossref | GoogleScholarGoogle Scholar |
Fensham R. J., Holman J. E.
(1998) The use of the land survey record to assess changes in vegetation structure. A case study from the Darling Downs, Queensland, Australia. The Rangeland Journal 20, 132–142.
Fensham R. J., Holman J. E.
(1999) Temporal and spatial patterns in drought related tree dieback in Australian savanna. Journal of Applied Ecology 36, 1035–1050.
| Crossref | GoogleScholarGoogle Scholar |
Finney, M. A. (1998).
Foley, J. C. (1957).
Henderson, R. J. F. (2002).
Krull E. S.,
Skjemstad J. O.,
Burrows W. H.,
Bray S. G.,
Wynn J. G.,
Bol R.,
Spouncer L., Harms B.
(2005) Recent vegetation changes in central Queensland, Australia: evidence from δ13C and 14C analyses of soil organic matter. Geoderma 126, 241–259.
| Crossref | GoogleScholarGoogle Scholar |
Lunt I. D.
(1997) Tree densities last century on the lowland Gippsland plain, Victoria. Australian Geographical Studies 35, 342–348.
| Crossref | GoogleScholarGoogle Scholar |
Mills, J. R. ,
Boyland, D. E. ,
and
Ahern, C. R. (1980).
Reynolds, J. A. ,
Muller, K. W. ,
and
Muller, S. L. (1992).
Roques K. G.,
O’Connor T. G., Watkinson A. R.
(2001) Dynamics of shrub encroachment in an African savanna: relative influences of fire, herbivory, rainfall and density dependence. Journal of Applied Ecology 38, 268–280.
| Crossref | GoogleScholarGoogle Scholar |
Turner, E. J. ,
Lee, A.N. ,
Hughes, K.K. ,
Ahern, C.R. ,
and
Beeston, G.R. (1978).
Turner, E. J. ,
McDonald, W. J. F. ,
Ahern, C. R. ,
and
Thomas, M. B. (1993).
Wilson B. A.,
Neldner V. J., Accad A.
(2002) The extent and status of remnant vegetation in Queensland and its implications for statewide vegetation management and legislation. Rangeland Journal 24, 6–35.
1Nomenclature according to Henderson (2002).