

Different responses of plant species to deferment of sheep grazing in a desert steppe of Inner Mongolia, China
Zhen Wang A , Xiangyang Hou A D , Michael P. Schellenberg C , Yan Qin A , Xiangjun Yun A D , Zhijun Wei B , Chao Jiang A and Yunfeng Wang AA Institute of Grassland Research, Chinese Academy of Agricultural Sciences, Hohhot 010010, P.R. China.
B Department of Grassland Science, College of Ecology and Environmental Science, Inner Mongolia Agricultural University, 306 Zhaowuda Road, Hohhot, Inner Mongolia 010018, P.R. China.
C Semiarid Prairie Agricultural Research Centre (SPARC), AAFC-AAC, Box 1030, Swift Current, Saskatchewan, Canada S9H 3X2.
D Corresponding authors. Email: Yunxiangjun@126.com; houxy16@126.com
The Rangeland Journal 36(6) 583-592 https://doi.org/10.1071/RJ13115
Submitted: 13 November 2013 Accepted: 4 November 2014 Published: 5 December 2014
Journal Compilation © Australian Rangeland Society 2014
Abstract
This paper reports the effects of different deferments of sheep grazing on plant variables of desert steppe vegetation in Inner Mongolia during 2010–2012. The study was initiated in May 2010 and comprised five treatments: no grazing (UG), grazing throughout the growing season (G) and grazing deferment for 40, 50 and 60 days (40UG, 50UG and 60UG) from the start of the growing season in a completely randomised block design replicated three times. The plant species were classified into two functional groups (C3 and C4 species) and the relationships among species composition, biomass and annual precipitation were examined in different deferments of sheep grazing. Treatment G significantly decreased the biomass of several species compared with some of the deferred treatments and treatment UG, because of differences in grazing preferences, and, consequently, decreased the aboveground net primary productivity (ANPP). Grazing deferment for 40, 50 and 60 days significantly increased C3-species richness and biomass compared with treatment UG, whereas grazing sheep’s preferences for C4 plants at some growth stages compared with C3 plants, resulted in lower species richness and biomass of C4 plants in treatments G, 40UG and 50UG in a dry year. Similarly, different species responses to treatment 60UG resulted in an increase in ANPP in a year with more precipitation during the growing season. The annual precipitation patterns strongly affected the temporal changes in biomass as well as the responses to grazing, indicating that the plant responses in the desert steppe were co-limited by grazing and precipitation. These findings provide important insights into the management and conservation of desert steppe vegetation in Inner Mongolia.
Additional keywords: aboveground net primary productivity, C3 species, C4 species, grazing preferences, plant functional groups.
Introduction
Changes in plant communities over time can be seen as expressions of the adaptation of the individual species to environmental change. These changes result from trade-offs between species and the different functions within individual species, thus linking environmental drivers and plant responses to ecosystem structure and function (Whalley 1994; Diaz et al. 1998, 2007; Wright et al. 2004). Climate change, including increases in atmospheric CO2 concentration, and changes in land use can result in changes in the proportions of C3 and C4 species in grassland communities (Bond 2008; Osborne 2008; Wittmer et al. 2010) but the details of these changes are often uncertain. In comparison to C4 species, lower temperatures enhance the growing conditions for C3 plants (cool season species), whereas C4 plants usually have higher temperature requirements for growth, and are generally known as warm season species (Monson et al. 1983). Elevated temperatures increase the competitiveness and expansion of warm season (C4) species over cool season (C3) species (Winslow and Hunt 2004), and further advantages may be gained through more efficient use of limited soil moisture (Wang and Wang 2001). The seasonal water availability may potentially contribute to the relative abundance of C3 and C4 species, with wet summers promoting increases in C4 species (Murphy and Bowman 2007).
Grazing can produce major changes to grassland attributes such as net primary production, plant species composition, community structure and ecosystem functioning (Anderson and Briske 1995). Different effects of grazing on C3 and C4 species result from complex interactions among forage quality, production and phenology (Ehleringer and Monson 1993). On the one hand, most C4 plants have a lower nutritive value than C3 species for ruminants when at the same stage of growth, due to lower nitrogen concentration and higher fibre content, which makes them prefer C3 plants to C4 plants. On the other hand, the biology of C4 plants could influence grazers to switch from C3 to C4 plants. For example, the higher water and nitrogen-use efficiency of C4 plants may contribute to higher productivity than C3 plants in a grassland ecosystem (Heckathorne et al. 1999). Moreover, C3 and C4 plants have different responses to environmental conditions, and plant phenology determines the length of the seasons when foliage is young and green, mature and of lower quality or dead. For instance, C3 grasses can stay green longer in regions with a cold winter, whereas C4 grasses may be young and actively growing in the late spring when C3 plants are mature and fibrous. In addition, C4 plants are typically more productive in warmer environments (Murphy and Bowman 2007). Changes in species composition in response to grazing are mostly the result of differences in preferences for the species involved and also in the plants’ responses to defoliation at different times and of different severity. Therefore, the effect of grazing on C3 and C4 plant species will depend on the interaction between quality, quantity and phenology of the plant species.
Grazing is an important agricultural activity in arid and semiarid environments, and the desert steppe has a long evolutionary history of grazing from both wild and domestic herbivores. The desert steppe is a vulnerable ecosystem located in the transition zone from grassland to desert, with soils vulnerable to erosion because of their sandy texture and low water-holding capacity (23–35 g kg–1 at a soil depth of 0–40 cm) (Pei et al. 2008), and low vegetation cover (Burke et al. 1998). Because of land management practices implemented by the Chinese government (the household responsibility system) in the desert steppe, the sheep population has increased dramatically since 1988 (Li et al. 2008). Livestock production from grazing native vegetation contributes to 73% of gross agricultural production of the desert steppe, which is the major income of herdsmen’s households. The total livestock numbers reached 486 916 head in 2007 (59% sheep and 39% goats). The region (desert steppe) encompasses six Sumus (administrative units or communes) and has a population of 69 000 (Hou et al. 2012). Both the increasing population of livestock and a lack of control of livestock distribution have led to the degradation of grasslands in the desert steppe (Li et al. 2000). The improper use of grazing (e.g. overgrazing) has severely reduced grassland productivity, vegetation cover and the proportion of forage grasses, and was considered to be the greatest threat causing the continuous degradation of grasslands in Inner Mongolia. In other words, overgrazing certainly enhances desertification.
Grazing inevitably affects the growth of both C3 and C4 plants. Variations in the abundance of different species, independent of whether they are C3 or C4 species, has been recently shown to be the result of ‘sampling’ activity by livestock (Gardener et al. 2003; Auerswald et al. 2009; Van der Meulen 2014). In Inner Mongolia, livestock prefer C3 species early in the growing season because of their early growth in the spring (Wang et al. 2003), which can lead to great damage to them. Compared with C3 species, C4 species develop ~6 weeks later when temperatures are higher and more favourable for them (Liang et al. 2002) and the negative effect of grazing on C3 plant species may increase the abundance of C4 plants later in the spring. A field experiment was conducted in a desert steppe to attempt to determine if the effects of grazing by sheep on C3 species in early spring could be avoided by grazing deferment. The objectives of the study were to better understand: (1) the changes of composition of C3 and C4 species in the desert steppe with grazing deferment using sheep; (2) the preferences of grazing sheep for different species and how these preferences change with grazing deferment; and (3) whether the differences in the effects of sheep-grazing deferment on species may lead to a shift in plant-community composition and whether these changes are linked to grazing pressure and the amount of precipitation during the growing season.
Materials and methods
Study site and vegetation
The experiment was conducted in the desert steppe (41°47ʹN, 111°53ʹE, 1450 m a.s.l.) at Xisu Banner in Inner Mongolia, China. This site is ~320 km north of Hohhot and has a prototypical continental climate with a mean annual temperature of 4.9°C (1953–2012). The lowest mean annual temperature occurred in 1956 (2.6°C) and the highest in 2007 (7.1°C). The minimum average monthly air temperature occurs in January (–14.8°C) while the maximum monthly temperature occurs in July (22.5°C). The mean annual precipitation is ~214 mm, 90% of which falls between May and October. The mean temperature from May to October is 16.2°C. The frost-free period is ~120 days. The sandy loam soil of the study site is classified as Kastanozem (The Canadian System of Soil Classification). The grassland condition at the initial stage of our study reflected the past heavy use, with a low canopy height (~7 cm) and canopy cover (14–20%). The dominant species were Stipa breviflora Griseb, Cleistogenes songorica (Roshev.) Ohwi and Allium polyrhizum Turcz. ex Regel, with sub-dominant species being Convolvulus ammannii Desr. and Salsola collina Pall. We classified the species into four plant functional groups as perennial graminoids, perennial forbs, shrubs and semi-shrubs, and annual and biennials (Table 1) (Bai et al. 2008).

|
Experimental design
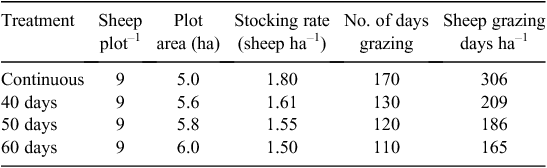
The experiment had a completely randomised block design on 85.2 ha of desert steppe. Grazing activities were performed yearly from 5 May to 31 October (a growing season of 170 days) with five treatments: continuous whole season grazing commenced on 5 May (G, plots of 5 ha each), grazing exclusion for 40 days (40UG, plots of 5.6 ha each), grazing exclusion for 50 days (50UG, plots of 5.8 ha each), grazing exclusion for 60 days (60UG, plots of 6 ha each) and no grazing (UG, plots of 6 ha each). Each treatment was replicated three times giving 15 plots in the experiment. The different grazing deferments had different stocking rates and sheep grazing days (Table 2), and the number of sheep was nine in every grazed plot. Grazing commenced on the 40UG plots on 15 June, on the 50UG plots on 25 June and on the 60UG plots on 4 July. Sheep were not randomly reassigned plots during the experimental period. The sheep were kept on the experiment for 2 years, and then replaced by 2-year-old sheep from the same flock. The sheep were of a similar age and liveweight. The sheep were given access to the plots from 0600 hours to 1800 hours each day, and then penned overnight. Water was provided twice (in early morning and evening) for sheep every day. Before the start and after the finish of the period of grazing in each year, all sheep were maintained on a confinement diet including hay, stover (wheat chaff), and some grains.
|
In each plot, 10 movable grazing exclusion cages (1.5 m × 1.5 m) were randomly established before grazing each year from 2010 to 2012. Care was taken to ensure that the cages were not placed in a location that had been sampled in the previous year. The peak biomass was sampled for individual species in mid-August within a quadrat (1 m × 1 m) inside each cage.
Attributes measured
Aboveground net primary production (ANPP) was determined as the biomass harvested in mid-August of each year from 2010 to 2012 inside the grazing exclusion cages set up each year at the beginning of the growing season (5 May). The biomass was cut to the ground surface (including living aboveground biomass and litter) by individual species inside a quadrat 1 × 1 m within each cage. A total of 150 quadrats were cut each year, 10 for each plot. Harvested biomass for each species was oven-dried at 65°C for 48 h and then weighed and ANPP was calculated as the sum of aboveground biomass for all plant species. Species richness (species number per quadrat) was used to describe the overall plant community composition and C3 community richness was the number of C3 species per quadrat and similarly for C4 community richness. Therefore, the data collected each August reflected the rainfall and temperature conditions of that growing season but the effects of the grazing treatments that had been applied in the growing season of the previous year. The data collected in 2010 from the different grazing treatments were a measure of the uniformity across the experimental area.
Statistical analyses
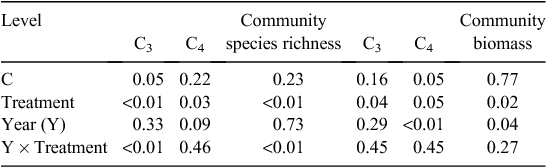
The effects of grazing treatments on biomass of dominant and sub-dominant species (S. breviflora, A. polyrhizum, C. songorica, S. collina, C. ammannii), C3 and C4 plant group richness and biomass, total plant community richness and ANPP for 2011 and 2012 were analysed using repeated-measures analysis of covariance (ANCOVA) with MIXED model (PROC MIXED, SAS Institute 2008) with sampling year as a repeated effect. Pre-treatment values of plant species biomass and richness in mid-August 2010 were used as covariates for those variables. If the interaction between sampling year and treatments was significant, the significance of treatments were tested within sampling year using the BY option. Multiple comparisons were conducted by Tukey’s HSD adjustment in SAS. Regression with correction for autocorrelation and stepwise multiple linear analyses were used to examine the relationships of plant community richness and productivity with grazing treatments and annual precipitation.
Results
Weather conditions
The mean temperatures in May–August were 21.1°C, 20.1°C and 19.9°C for 2010, 2011 and 2012, respectively, and ranged from 1.7°C, 0.7°C to 0.5°C above the long-term average (19.4°C) (Fig. 1). The mean annual precipitation in 2010 (204 mm) and 2011 (142 mm) were lower than the long-term mean (214 mm) by 4.7% and 33.6%, respectively. The total precipitation in 2012 (329 mm) was 53.7% higher than the long-term mean. During the growing period, the accumulated precipitation in May–August were 128 mm, 82 mm and 245 mm for the years from 2010, 2011 and 2012, respectively, which were 82.3%, 52.7% and 157.5% of the long-term average in May–August (156 mm).

|
Effect of grazing on vegetation characteristics
Twenty-four species were found in the plots, including 15 C3 and 9 C4 species (Table 1). Among the 15 C3 plants, most were perennials, whereas only two species were annual or biennial (Table 1). Among the nine C4 plants, most were annual or biennials, including Amaranthus retroflexus L., Enneapogon boreali (Griseb.) Honda., Eragrostis pilosa (L.) Beauv., Portulaca oleracea L., Salsola collina Pall., Setaria viridis (L.) Beauv. and Tragus berteronianus Schult. (Table 1).
Effect of grazing treatments on important plant species
Stipa breviflora Griseb. (C3-dominant perennial bunch grass). The grazing treatments (P < 0.01), sampling year (P = 0.03) and their interaction (P = 0.03) significantly affected the peak biomass of this species (Table 3). The biomass was higher in the wet 2012 than in the dry 2011 (Table 3). The biomass of S. breviflora was the highest in the 60UG treatment (23.3 g m–2) in the wet 2012, and the lowest in the G, 40UG and 60UG treatments (~0.31 g m–2) in the dry 2011 (P < 0.05, Fig. 2a).

|

|
Allium polyrhizum Turcz. ex Regel. (C3-dominant perennial forb). The grazing treatments (P < 0.01) significantly affected the biomass of A. polyrhizum, whereas there were no effects of sampling year (P = 0.45) or its interaction with grazing treatment (P = 0.86) (Table 3). There were no significant differences among the grazing treatments for biomass of A. polyrhizum in the dry 2011 (Fig. 2b). However, in the wet 2012, compared with treatments UG and G, both treatments 40UG and 50UG produced significantly more biomass (P < 0.05, Fig. 2b).
Convolvulus ammannii Desr. (C3-sub-dominant perennial forb). None of the grazing treatments, sampling year, or their interaction affected the biomass of C. ammanni (Table 3, Fig. 2c).
Cleistogenes songorica (Roshev.) Ohwi. (C4-dominant perennial bunch grass). The grazing treatments significantly affected the biomass of C. songorica (P < 0.01, Table 3). The biomass of C. songorica was higher in the dry 2011 than in the wet 2012 (P = 0.05, Table 3). Compared with treatment UG, the other grazing treatments significantly reduced the biomass of C. songorica in 2012 (P < 0.05, Fig. 2d).
Salsola collina Pall. (C4-sub-dominant annual herb). The biomass was higher in the wet 2012 than in the dry 2011 (P < 0.01, Table 2; Fig. 2e). The biomass was lower for treatment 40UG than treatments UG and 60UG in the wet 2012 (P < 0.05), whereas no differences were detected in the dry 2011 (Fig. 2e). As a weedy annual, S. collina accounted for 4.5% of ANPP in (2011), and for 71.5% in 2012 (Fig. 2e).
Effect of grazing treatments on C3 and C4 communities
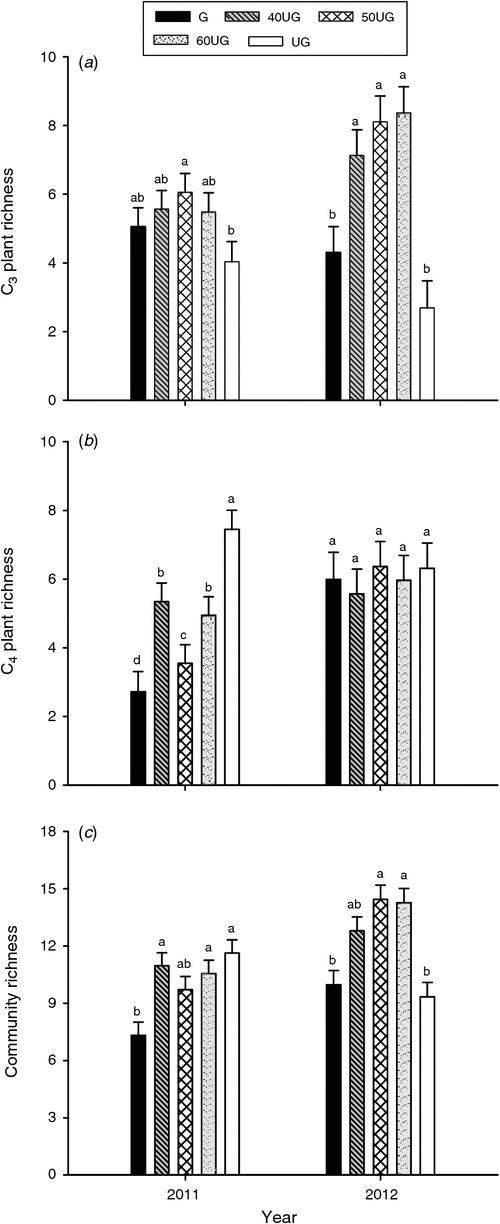
C3 plant community. Grazing treatments significantly affected the C3 plant community richness (P < 0.01), whereas no effects of the sampling year were detected (P = 0.33, Table 4). The C3 plant community richness was higher in treatment 50UG than in treatment UG in the dry 2011 (P < 0.05, Fig. 3a). Compared with treatments UG and G, grazing deferment (treatments 40UG, 50UG and 60UG) significantly increased C3 plant community richness in the wet 2012 (P < 0.05, Fig. 3a). Grazing deferment significantly increased the C3 plant community biomass averaged over both years (P = 0.04, Table 3) but there were no differences among the treatments in the dry 2011 (Fig. 4a). In the wet 2012, grazing deferment (treatments 40UG, 50UG and 60UG) significantly increased C3 plant community biomass compared with the G and UG treatments (P < 0.05, Fig. 4a).
|
|
|
C4 plant community. The grazing treatments significantly affected C4 plant richness (P = 0.03, Table 4). The C4 plant richness was higher in treatment UG than in the other grazing treatments (treatments G, 40UG, 50UG and 60UG) in the dry 2011 with treatment G the lowest of all treatments (P < 0.05, Fig. 3b). Similar to C4 plant richness, grazing deferment significantly decreased the C4 plant biomass (P = 0.05, Table 4) overall. The C4 plant community biomass was lower in 2011 than in 2012 (P < 0.01, Table 4; Fig. 4b). Compared with treatment UG, grazing deferment decreased C4 plant biomass in the dry 2011, and treatments G, 40UG and 50UG significantly decreased C4 plant biomass, with treatment 40UG the lowest of all the treatments, in the wet 2012 (P < 0.05, Fig. 4b).
Effect of grazing treatments on plant communities
The grazing treatments significantly affected plant community richness (P < 0.01, Table 4). The plant community richness was lower in treatment G than in treatments UG, 40UG and 60UG in 2011 (P < 0.05), but the plant community richness was higher in treatments 50UG and 60UG than in treatments G and UG in the wet 2012 (P < 0.05, Fig. 3c). A similar pattern occurred with the plant community biomass (P = 0.02, Table 4). The community ANPP was higher in the wet 2012 than in the dry 2011 (P = 0.04, Table 4). Compared with treatment UG, the grazing treatments (treatments G, 40UG, 50UG and 60UG) had a significantly lower plant community ANPP in the dry 2011, whereas treatment 60UG had a significantly higher community ANPP in the wet 2012 compared with all other treatments (P < 0.05, Fig. 4c).
Impacts of grazing treatments and annual precipitation on vegetation
Whereas both C3 plant richness (R2 = 0.52, P < 0.001) and total plant community richness (R2 = 0.32, P = 0.005) showed quadratic relationships with increasing grazing deferment (Fig. 5a, c), C4 plant richness increased linearly (R2 = 0.33, P < 0.001) (Fig. 5b). The C3 plant biomass and grazing deferment also showed a quadratic relationship (R2 = 0.30, P = 0.008; Fig. 5d). However, there was no significant relationship between the grazing treatments and the C4 plant species biomass and the ANPP (Fig. 5e, f).

|
Discussion
Stocking rates, grazing days and grazing deferment
Moderate grazing pressure generally increases biodiversity and productivity of grassland ecosystems (McNaughton 1979), whereas heavy grazing severely decreases these attributes (Noy-Meir et al. 1989). In this study, the stocking rates and total number of sheep grazing-days per growing season were confounded with the grazing deferment (Table 2). The G treatment had a stocking rate of 1.80 sheep ha–1 for the whole of the growing season resulting in 306 sheep grazing-days. However, the 60UG treatment had a stocking rate of 1.50 sheep ha–1 for the last 110 days of the growing season giving only 165 sheep grazing days with the other grazing deferment treatments being intermediate (Table 2). Therefore, the stocking rates of the longer deferment treatments were lower that of the continuous stocking treatment and would perhaps have decreased the impact on the effects of the deferment. In addition, the reduction of the total sheep grazing-days for the growing season by nearly half would have a major impact on the returns from the rangeland to the landholder, which would have to be balanced against the changes in species composition.
The vegetation community structure and composition were strongly associated with grazing deferment, and the community plant species richness showed a hump-shaped relationship with the decreasing of grazing deferment (Fig. 5c), with high species richness in treatment 50UG and low richness in treatment G. In addition, compared with treatment G, the community ANPP was higher under the deferred grazing (treatments 50UG and 60UG) due to reduced grazing damage in the early spring. Both the reduced grazing damage for plant species at the start of grazing and the lower number of sheep grazing-days resulted in the high ANPP and species richness at the end of grazing in these two deferred grazing treatments.
The effect of grazing deferment on plant species and plant functional groups
The original steppe vegetation in this region was dominated by C3 perennial grasses including Leymus chinensis (Trin.) Tzvel, Stipa grandis P. Smirn., Achnatherum sibiricum (L.) Keng and Agropyron cristatum (L.) P. Beauv. (Wang et al. 2014), but these species are no longer present in the degraded desert steppe used in this study. The sole remaining dominant from this functional group was S. breviflora. Nevertheless, the same responses to grazing deferment by these original dominants would perhaps be a reasonable assumption.
The grazing preferences for different species of forage were related to the changes in grassland productivity in this study. The continuous grazing for the whole growing season (treatment G) decreased the biomass of the C3-dominant S. breviflora because this species was preferentially grazed early in the growing season. The 60UG treatment for 2010 and 2011 meant that this important species was able to recover and produce the largest biomass of any of the grazing treatments in 2012. In addition, the variation in the proportion of the C3/C4 species contributing to the biomass was determined by the preferential grazing activity of sheep due to the different grazing periods. In Inner Mongolia, sheep graze the C3 species in early spring because of their early development (Wang et al. 2003), which leads to overgrazing and damage to these species. Compared with the C3 species, C4 species develop about 6 weeks later when the temperatures are higher and more favourable for their growth (Liang et al. 2002). In the present study, the C4 plants were at different phenological stages due to their later growth initiation in the grazing deferment treatments. The livestock seek out young C4 plants due to a preference for fresh succulent feed (Williams 1972). The C4 species were at a young phenological stage when the sheep were introduced into the 40UG treatment and they suffered the heaviest grazing. With the increasing time of deferment (50UG and 60UG treatments), the plants were at a later stage when the sheep were introduced and they were less preferred. The result was an increase in C4 biomass with increasing grazing deferment in 2012 (Fig. 4b). These differences were not apparent in the 2011 results (Fig. 4b) because the average rainfall in 2010 would have meant that the greater total feed availability would have meant that there was less selectivity by the sheep. However, the amount of biomass of the C4 plants produced in 2011 was very low (Fig. 4b) and so the preferences of the sheep for the young plants of the C4 species would have been much higher than in 2010. The results in 2012 (a very wet year) reflected the results of the grazing selectivity in the previous dry year. This result emphasises that the rangeland degradation that occurs in a dry year may not be apparent until later, wet years (Yan et al. 2013).
The difference in biomass for the annual S. collina between the dry year and the wet year was greater than for any other single species or functional group. It is suggested that this annual species was able to colonise the gaps produced by overgrazing between the perennial species in 2011, in the wet year of 2012. The lowest biomass of S. collina produced in 2012 was in the 40UG deferment treatment. Perhaps the seedlings of this species were at the growth stage where they were most preferred when the sheep were introduced into these plots in 2011 and so were heavily grazed, reducing seed production in this treatment compared with that in the other treatments.
The quadratic relationships for C3 plant richness and biomass, and grazing deferment (Fig. 5a, d) suggest that grazing offers a management tool to maintain production, biodiversity and habitat structure (McNaughton 1979). The high biomass produced by C3 plants in the 50UG and 60UG treatments (Fig. 5d) may be attributed to the earlier growth in spring of the perennials S. breviflora and A. polyrhizum. The young C3 plants suffer little damage when the grazing is deferred in early spring. Therefore, C3 plant take advantage of the good growing conditions and have a high biomass with increasing grazing deferment up to nearly 60 days. These observations suggest delaying grazing maintains C3 dominance in the desert steppe.
The effects of grazing deferment on the plant communities
The grazing deferment treatments (treatments 40UG, 50UG and 60UG) significantly increased C3 plant community biomass compared with the continuously grazed treatment but there were no differences among these three treatments (Fig. 4a). However, treatment 40UG produced less C4 plant biomass than the G and the 50UG treatments and the C4 plant biomass in the 60UG treatment was the highest of all the grazing treatments. Thus, the compensatory growth (Bai et al. 2004; Fanselow et al. 2011) between the C3 and C4 plants meant that only the 60UG treatment had a higher total plant community productivity (ANPP) than all the other grazing treatments (Fig. 4c). Therefore, the results of this study support the grazing optimisation hypothesis which predicts that appropriate grazing can maintain or improve primary production (McNaughton 1979). Our results are also in agreement with previous studies in China (Su et al. 2005; Han et al. 2008).
Roles of temporal variations in precipitation
The frequency and amounts of precipitation have important impacts on plant biomass production (Knapp et al. 2002; Nippert et al. 2009). The higher production in the wet year than in the dry year in our study (Table 4; Fig. 4c) is consistent with previous studies (Xia et al. 2009; Zheng et al. 2011). The different responses among plant functional groups to varying precipitation suggest that there are trade-offs among plant species in dry versus wet years. Precipitation plays an important role in the amount of C4 community biomass produced, and a wet growing season promotes C4 plant expansion (Murphy and Bowman 2007). For example, the precipitation increased the absolute and relative abundance of opportunistic and unpreferred annuals with shallow roots, and it has been shown that annuals have a competitive advantage in depleting soil available nutrients and water (Bai et al. 2012). In our study, the increase of S. collina (annual C4 species) could have depended on the seasonal variations of precipitation. Salsola collina became even more dominant at the end of the 2 years’ study (Table 3; Fig. 2e). Nevertheless, the shift to an annual-dominated community under high precipitation could have profound impacts on ecosystem functioning and services, as these annual-biennial species were less preferred by domestic grazers than the perennial forbs and grasses in this ecosystem (Han et al. 2004). Moreover, compared with treatment UG, all the grazing treatments applied in an average year (2010), significantly decreased community ANPP in the following dry year (2011), whereas only treatment 40UG, applied in a dry year, decreased community ANPP in the following wet year (2012) (Fig. 4c). These findings indicate the importance of changes in precipitation in mediating the negative effects of heavy grazing in the desert steppe in northern China and also that damage from overgrazing or incorrect grazing in a dry year is very apparent in a following wet year. The length of time that these effects continue cannot be determined from this study.
Conclusion
This study produced a better understanding of the effects of grazing pressure, grazing deferment and growing season rainfall on desert steppe vegetation as proposed in the objectives. The continuous season grazing (treatment G) decreased biomass of the C3-dominant perennial grass S. breviflora, compared with 60-days grazing deferment, due to sheep’s preferences for this species during the early growing season. Compared with ungrazed plots, continuous whole season grazing also decreased the biomass of C4 plants. Delayed grazing (treatments 50UG and 60UG) increased species richness and biomass of C3 species. These results emphasise that different grazing deferment periods can control plant-community composition in the desert steppe. The rainfall differences in the growing seasons (dry year: 2011; wet year: 2012) provided a unique opportunity to understand how inter-annual variation in precipitation can affect both C3 and C4 species and plant communities, and their responses to grazing deferment in the desert steppe. These results also illustrate the importance of very dry years in the degradation of vegetation under grazing in the desert steppe in Inner Mongolia.
Acknowledgements
We thank Shixian Sun, Yuanheng Li and Xiliang Li for their help in setting up the experiment and for field data measurements. This study was financially supported by the National Key Basic Research Program of China (2014CB138806), the Important National Natural Science Foundation of China (70933004), the National Important Research Program of Inner Mongolia (2010ZD08), China Postdoctoral Science Foundation (2013M541096), National Natural Science Foundation of Inner Mongolia (2014BS0329) and Central Non-profit Research Institutes Fundamental Research Funds (1610332014010).
References
Anderson, V. J., and Briske, D. D. (1995). Herbivore-induced species replacement in grasslands – is it driven by herbivory tolerance or avoidance. Ecological Applications 5, 1014–1024.| Herbivore-induced species replacement in grasslands – is it driven by herbivory tolerance or avoidance.Crossref | GoogleScholarGoogle Scholar |
Auerswald, K., Wittmer, M. H. O. M., Männel, T. T., Bai, Y. F., Schäufele, R., and Schnyder, H. (2009). Large regional-scale variation in C3/C4 distribution pattern of Inner Mongolia steppe is revealed by grazer wool carbon isotope composition. Biogeosciences 6, 795–805.
| Large regional-scale variation in C3/C4 distribution pattern of Inner Mongolia steppe is revealed by grazer wool carbon isotope composition.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXpvFyrtbY%3D&md5=026c3ef37c13fafc328a26f2eff00740CAS |
Bai, Y. F., Han, X. G., Wu, J. G., Chen, Z. Z., and Li, L. H. (2004). Ecosystem stability and compensatory effects in the Inner Mongolia grassland. Nature 431, 181–184.
| Ecosystem stability and compensatory effects in the Inner Mongolia grassland.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXntlGks7w%3D&md5=4521d03d8ef508af376edf636aff5f41CAS |
Bai, Y. F., Wu, J. G., Xing, Q., Pan, Q. M., Huang, J. H., Yang, D. L., and Han, X. G. (2008). Primary production and rain-use efficiency across a precipitation gradient on the Mongolia plateau. Ecology 89, 2140–2153.
| Primary production and rain-use efficiency across a precipitation gradient on the Mongolia plateau.Crossref | GoogleScholarGoogle Scholar |
Bai, Y. F., Wu, J. G., Clark, M. C., Pan, Q., Zhang, L., Chen, S., Wang, Q. B., and Han, X. G. (2012). Grazing alters ecosystem functioning and C:N:P stoichiometry of grasslands along a regional precipitation gradient. Journal of Applied Ecology 49, 1204–1215.
| Grazing alters ecosystem functioning and C:N:P stoichiometry of grasslands along a regional precipitation gradient.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3sXislSmt7k%3D&md5=43c8bcb788cc850737201d9f8cd3a81cCAS |
Bond, W. J. (2008). What limits trees in C4 grasslands and savannas? Annual Review of Ecology, Evolution and Systematics 39, 641–659.
| What limits trees in C4 grasslands and savannas?Crossref | GoogleScholarGoogle Scholar |
Burke, I. C., Lauenroth, W. K., Vinton, M. A., Hook, P. B., Kelly, R. H., Epstein, H. E., Aguiar, M. R., Robles, M. D., Aguilera, M. O., Murphy, K. L., and Gill, R. A. (1998). Plant-soil interactions in temperate grasslands. Biogeochemistry 42, 121–143.
| Plant-soil interactions in temperate grasslands.Crossref | GoogleScholarGoogle Scholar |
Diaz, S., Cabido, M., and Casanoves, F. (1998). Plant functional traits and environmental filters at a regional scale. Journal of Vegetation Science 9, 113–122.
| Plant functional traits and environmental filters at a regional scale.Crossref | GoogleScholarGoogle Scholar |
Diaz, S., Lavorel, S., de Bello, F., Quetier, F., Grigulis, K., and Robson, M. (2007). Incorporating plant functional diversity effects in ecosystem service assessments. Proceedings of the National Academy of Sciences of the United States of America 104, 20 684–20 689.
| Incorporating plant functional diversity effects in ecosystem service assessments.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXkt1ejsA%3D%3D&md5=dd15b35f1c3dc8cf2a29867e77607a81CAS |
Ehleringer, J. R., and Monson, R. K. (1993). Evolutionary and ecoloigcal aspects of photosynthetic pathway variation. Annual Review of Ecology and Systematics 24, 411–439.
| Evolutionary and ecoloigcal aspects of photosynthetic pathway variation.Crossref | GoogleScholarGoogle Scholar |
Fanselow, N., Schoenbach, P., Gong, X. Y., Lin, S., Taube, F., Loges, R., Pan, Q., and Dittert, K. (2011). Short-term regrowth responses of four steppe grassland species to grazing intensity, water and nitrogen in Inner Mongolia. Plant and Soil 340, 279–289.
| Short-term regrowth responses of four steppe grassland species to grazing intensity, water and nitrogen in Inner Mongolia.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXitFGkt7w%3D&md5=4c50f27791d5c3c10ccd8e6b29b4621dCAS |
Gardener, M. R., Whalley, R. D. B., and Sindel, B. M. (2003). Ecology of Nassella neesiana, Chilean needle grass in pastures on the Northern Tablelands of New South Wales. I. Seed production and dispersal. Australian Journal of Agricultural Research 54, 613–619.
| Ecology of Nassella neesiana, Chilean needle grass in pastures on the Northern Tablelands of New South Wales. I. Seed production and dispersal.Crossref | GoogleScholarGoogle Scholar |
Han, G. D., Biligetu, , and Gao, A. D. (2004). Comparison study on selective intake behavior of sheep at different stocking rates in Stipa breviflora desert steppe. Pratacultural Science 21, 95–98.
Han, G. D., Hao, X. Y., Zhao, M. L., Wang, M. J., Ellert, B. H., Willms, W., and Wang, M. J. (2008). Effect of grazing intensity on carbon and nitrogen in soil and vegetation in a meadow steppe in Inner Mongolia. Agriculture, Ecosystems & Environment 125, 21–32.
| Effect of grazing intensity on carbon and nitrogen in soil and vegetation in a meadow steppe in Inner Mongolia.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXjtlSrsr8%3D&md5=e6da2b0eb1ff60f442b10cafc89e84b8CAS |
Heckathorne, S. A., McNaughton, S. J., and Coleman, J. S. (1999). C4 plants and herbivory. C4 Plant Biology 9, 285–312.
Hou, X. Y., Han, Y., and Li, F. Y. (2012). The perception and adaptation of herdsmen to climate change and climate variability in the desert steppe region of northern China. The Rangeland Journal 34, 349–357.
| The perception and adaptation of herdsmen to climate change and climate variability in the desert steppe region of northern China.Crossref | GoogleScholarGoogle Scholar |
Knapp, A. K., Fay, P. A., Blair, J. M., Collins, S. L., Smith, M. D., Carlisle, J. D., Harper, C. W., Danner, B. T., Lett, M. S., and McCarron, J. K. (2002). Rainfall variability, carbon cycling, and plant species diversity in a mesic grassland. Science 298, 2202–2205.
| Rainfall variability, carbon cycling, and plant species diversity in a mesic grassland.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38XpsVSktLg%3D&md5=2d8b35ddbe81d75a2962418c5bd9d129CAS | 12481139PubMed |
Li, S. G, Harazono, Y., Oikawa, T., Zhao, H. L., He, Z. Y., and Chang, X. L. (2000). Grassland desertification by grazing and the resulting micrometeorological changes in Inner Mongolia. Agricultural and Forest Meteorology 102, 125–137.
| Grassland desertification by grazing and the resulting micrometeorological changes in Inner Mongolia.Crossref | GoogleScholarGoogle Scholar |
Li, C. L., Hao, X. Y., Zhao, M. L., Han, G. D., and Willms, W. D. (2008). Influence of historic sheep grazing on vegetation and soil properties of a Desert Steppe in Inner Mongolia. Agriculture, Ecosystems & Environment 128, 109–116.
| Influence of historic sheep grazing on vegetation and soil properties of a Desert Steppe in Inner Mongolia.Crossref | GoogleScholarGoogle Scholar |
Liang, C., Michalk, D. L., and Millar, G. D. (2002). The ecology and growth patterns of Cleistogenes species in degraded grasslands of eastern Inner Mongolia, China. Journal of Applied Ecology 39, 584–594.
| The ecology and growth patterns of Cleistogenes species in degraded grasslands of eastern Inner Mongolia, China.Crossref | GoogleScholarGoogle Scholar |
McNaughton, S. J. (1979). Grazing as an optimization process: grass-ungulate relationships in the serengeti. American Naturalist 113, 691–703.
| Grazing as an optimization process: grass-ungulate relationships in the serengeti.Crossref | GoogleScholarGoogle Scholar |
Monson, R. K., Littlejohn, R. O., and Williams, G. J., III (1983). Photosynthetic adaptation to temperature in four species from the Colorado shortgrass steppe: a physiological model for coexistence. Oecologia 58, 43–51.
| Photosynthetic adaptation to temperature in four species from the Colorado shortgrass steppe: a physiological model for coexistence.Crossref | GoogleScholarGoogle Scholar |
Murphy, B. P., and Bowman, D. M. J. S. (2007). Seasonal water availability predicts the relative abundance of C3 and C4 grasses in Australia. Global Ecology and Biogeography 16, 160–169.
| Seasonal water availability predicts the relative abundance of C3 and C4 grasses in Australia.Crossref | GoogleScholarGoogle Scholar |
Nippert, J. B., Fay, P. A., Carlisle, J. D., Knapp, A. K., and Smith, M. D. (2009). Ecophysiological responses of two dominant grasses to altered temperature and precipitation regimes. Acta Oecologica 35, 400–408.
| Ecophysiological responses of two dominant grasses to altered temperature and precipitation regimes.Crossref | GoogleScholarGoogle Scholar |
Noy-Meir, I., Gutman, M., and Kaplan, Y. (1989). Responses of Mediterranean grassland plants to grazing and protection. Journal of Ecology 77, 290–310.
| Responses of Mediterranean grassland plants to grazing and protection.Crossref | GoogleScholarGoogle Scholar |
Osborne, C. P. (2008). Atmosphere, ecology and evolution: what drove the Miocene expansion of C4 grasslands? Journal of Ecology 96, 35–45.
| 18784799PubMed |
Pei, S. F., Fu, H., and Wan, C. G. (2008). Changes in soil properties and vegetation following exclosure and grazing in degraded Alxa desert steppe of Inner Mongolia, China. Agriculture, Ecosystems & Environment 124, 33–39.
| Changes in soil properties and vegetation following exclosure and grazing in degraded Alxa desert steppe of Inner Mongolia, China.Crossref | GoogleScholarGoogle Scholar |
SAS Institute (2008). ‘SAS OnlineDoc 9.1.3.’ (SAS Institute Inc.: Cary, NC.)
Su, Y.-Z., Li, Y.-L., Cui, H.-Y., and Zhao, W.- Z. (2005). Influences of continuous grazing and livestock exclusion on soil properties in a degraded sandy grassland, Inner Mongolia, northern China. Catena 59, 267–278.
| Influences of continuous grazing and livestock exclusion on soil properties in a degraded sandy grassland, Inner Mongolia, northern China.Crossref | GoogleScholarGoogle Scholar |
Van der Meulen, A. (2014). Invasion of the Northern Tablelands of New South Wales by Nassella trichotoma (Nees) Hack. ex Arechav.: landholder perceptions and seed ecology. PhD Thesis, University of New England, Armidale, NSW, Australia.
Wang, S. P., and Wang, Y. F. (2001). Study on over-compensation growth of Cleistogenes squarrosa population in Inner Mongolia Steppe. Acta Botanica Sinica 43, 413–418.
Wang, S. P., Wang, Y. F., and Chen, Z. Z. (2003). Effect of climate change and grazing on populations of Cleistogenes squarrosa in Inner Mongolia steppe. Acta Phytoecologica Sinica 27, 337–343.
Wang, Z., Zhang, Q., Xin, X. P., Ding, Y., Hou, X. G., Sarula, , Li, X. L., Chen, H. J., Yin, Y. T., Hu, J., and Liu, Z. G. (2014). Response of the annual biomass production of a typical steppe plant community to precipitation fluctuations. The Rangeland Journal 36, 527–534.
Whalley, R. D. B. (1994). Successional theory and vegetation change. Tropical Grasslands 28, 195–205.
Williams, D. G. (1972). Ecological studies on shrub-steppe of the Western Riverina. New South Wales. PhD Thesis, Australian National University, Canberra, Australia.
Winslow, J. C., and Hunt, E. R. (2004). Using seasonal water availability to predict percentages of cool and warm season grasses. In: ‘57th Annual Meeting of the Society for Range Management’. Abstract, p. 223. (Society for Range Management: Littleton, CO.)
Wittmer, M., Auerswald, K., Bai, Y. F., Schaufele, R., and Schnyder, H. (2010). Changes in the abundance of C3/C4 species of Inner Mongolia grassland: evidence from isotopic composition of soil and vegetation. Global Change Biology 16, 605–616.
| Changes in the abundance of C3/C4 species of Inner Mongolia grassland: evidence from isotopic composition of soil and vegetation.Crossref | GoogleScholarGoogle Scholar |
Wright, I. J., Reich, P. B., Westoby, M., Ackerly, D. D., Baruch, Z., Bongers, F., Cavender-Bares, J., Chapin, T., Cornelissen, J. H. C., Diemer, M., Flexas, J., Garnier, E., Groom, P. K., Gulias, J., Hikosaka, K., Lamont, B. B., Lee, T., Lee, W., Lusk, C., Midgley, J. J., Navas, M. L., Niinemets, U., Oleksyn, J., Osada, N., Poorter, H., Poot, P., Prior, L., Pyankov, V. I., Roumet, C., Thomas, S. C., Tjoelker, M. G., Veneklaas, E. J., and Villar, R. (2004). The worldwide leaf economics spectrum. Nature 428, 821–827.
| The worldwide leaf economics spectrum.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXjt1Crt74%3D&md5=73a55078facbe298e4afe033a510f5ceCAS | 15103368PubMed |
Xia, J. Y., Niu, S. L., and Wan, S. (2009). Response of ecosystem carbon exchange to warming and nitrogen addition during two hydrologically contrasting growing seasons in a temperate steppe. Global Change Biology 15, 1544–1556.
| Response of ecosystem carbon exchange to warming and nitrogen addition during two hydrologically contrasting growing seasons in a temperate steppe.Crossref | GoogleScholarGoogle Scholar |
Yan, L., Zhou, G., and Zhang, F. (2013). Effects of different grazing intensities on grassland production in China: a meta-analysis. PLoS ONE 8, e81466.
| Effects of different grazing intensities on grassland production in China: a meta-analysis.Crossref | GoogleScholarGoogle Scholar | 24324694PubMed |
Zheng, S. X., Lan, Z. C., Li, W. H., Shao, R. X., Shan, Y. M., Wan, H. W., Taube, F., and Bai, Y. F. (2011). Differential responses of plant functional trait to grazing between two contrasting dominant C3 and C4 species in a typical steppe of Inner Mongolia, China. Plant and Soil 340, 141–155.
| Differential responses of plant functional trait to grazing between two contrasting dominant C3 and C4 species in a typical steppe of Inner Mongolia, China.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXitFGks7k%3D&md5=a7e393afb0fff8ef7011a67e1d67cef9CAS |