

The red fox (Vulpes vulpes) as a potential invasive predator in a Critically Endangered sawfish nursery
David L. Morgan A * , Karissa O. Lear A , Jack Ingelbrecht A , Travis Fazeldean A and Patricia A. Fleming B
A * , Karissa O. Lear A , Jack Ingelbrecht A , Travis Fazeldean A and Patricia A. Fleming B
A
B
Abstract
Although the red fox (Vulpes vulpes) is considered one of the most damaging and adaptive invasive carnivorous mammals that consumes a wide variety of vertebrate and invertebrate taxa, there are surprisingly few reports of red foxes hunting fish. We observed evidence of an attempted predation event by a red fox on a neonate green sawfish (Pristis zijsron) within a deltaic island in the Ashburton River estuary, a remote desert river in Western Australia. The site is a globally important nursery where newborn sawfish arrive annually in spring. Injuries to the sawfish included paw/claw marks on the head, damage to the rostrum, which is a formidable tool that is used for both defence against predators and for detecting and attacking prey, as well as a major hole in the head and damage to the gills, which are vital for respiration, osmoregulation, nitrogenous waste excretion, pH regulation, and hormone production. A series of tracks suggests at least one fox parades the shallow tidally influenced banks, with evidence of a green mud crab (Scylla serrata) having also been predated on. There have also been reports of red fox predation of sea turtle nests nearby. This is the first record of a red fox hunting in marine waters and one of the few identifying fish as prey. We suggest that a monitoring program for foxes, and possibly a control program, is warranted prior to the annual seasonal colonisation of this habitat by neonate green sawfish and nesting turtles, which may in turn reduce predation of sympatric species.
Keywords: green sawfish, Indian Ocean, nursey habitat, Pilbara, Pristidae.
Introduction
The red fox (Vulpes vulpes) is one of the most widely distributed mesocarnivores in the world, existing across a range of habits from the artic to arid deserts and urban areas (Basuony et al. 2005). The red fox was introduced to the mainland of south-eastern Australian in the 1860s (Abbott 2011), established from around 1874 (Fairfax 2019), and has subsequently spread across ~80% of the continent and onto ~50 islands (Abbott et al. 2014; Stobo-Wilson et al. 2022). The red fox was first reported in Western Australia between 1911 and 1912 near the south-eastern border with South Australia (Crawford 1912). This species then spread rapidly throughout the southern parts of the state and was first reported in the Pilbara (~1000 km to the north) from the 1920s (see Long 1974; King and Smith 1985; Fairfax 2019). Foxes prey on an array of terrestrial animals (mammals, reptiles, birds, amphibians, and invertebrates) that are native or introduced to the Australian continent, and are considered one of the most threatening invasive species contributing to the extinction of a number of terrestrial animals (Fleming et al. 2021; Stobo-Wilson et al. 2022).
While their opportunistic diet is believed to have contributed to their success as a global invader (Bateman and Fleming 2012), very few dietary studies of foxes have identified freshwater or marine fishes as prey (Castañeda et al. 2022). In Egypt, Basuony et al. (2005) found fish remains in the stomachs of red foxes at Lake Nassar, Egypt, although these were collected near where fisherman discarded fish remains and the foxes may have been opportunistically feeding on discarded waste. A single largemouth bass (Micropterus salmoides) was found in one of 125 (0.8%) red fox guts examined in the United States (Wagnon and Serfass 2017). More recently, an adult male red fox was recorded hunting in shallow waters of a reservoir in Spain and capturing European carp (Cyprinus carpio), which were congregating and spawning in the area (Tobajas and Díaz-Ruiz 2022a, 2022b). What was interesting about that behavioural study was that the individual fox was observed capturing carp in 10 out of 12 attempts and was thought to be targeting the highly nutritional eggs of the fish. It was suggested that this may have been an opportunistic event, due to the presence of many spawning fish in shallow waters and may also have been an example of surplus killing, as a female red fox was observed caching fish (without hinderance from a male present) in a similar manner to that reported earlier by Basuony et al. (2005).
A review of 85 dietary studies of the red fox throughout their Australian geographic range, across all seasons and over seven decades, demonstrated that foxes are opportunistic predators consuming whatever prey presents at a given space and time (Fleming et al. 2021); however only 0.22 ± 0.75% of Australian fox diet samples have contained fish (Fleming et al. 2022). Drying out of inland waters can provide an important foraging opportunities for foxes. For example, introduced European carp have been reported in fox guts in Victoria, Australia (seven of 967 (0.7%) (Coman 1973) and four of 212 (1.9%) fox stomachs collected from agricultural sites around Wentworth, New South Wales (NSW) (Lugton 1993)). Stranded carp have been noted to attract foxes to a drying lake in south-west NSW (Lugton 1993). Inland waterways also provide important foraging sites for foxes: fish remains were found in three of 112 (2.7%) fox scats collected during summer from around ski lodges in Kosciusko National Park, NSW (Bubela et al. 1998), and in two of 99 (2.0%) stomach samples from the Darling River, NSW (Ryan and Croft 1974). Coastal sites also offer opportunities for foxes, with observations suggesting opportunistic scavenging on carrion; fish were found in the scats of four of 273 (1.5%) foxes from NSW coastal reserves (Meek and Triggs 1998), 35 of 1185 (3.0%) fox scats from the Sunshine Coast, Queensland (O’Connor et al. 2020), and four of 937 (0.4%) fox scats from sites adjacent to the Mallacoota Inlet, Victoria (Triggs et al. 1984). Scavenging of fish remains from anthropogenic sources by foxes is also possible, with Stepkovitch (2017) reporting fish remains in the stomach contents of one of 108 (0.9%) foxes from Sydney, NSW, but was unable to determine whether it originated from a freshwater or anthropogenic source.
Here, we report on an interaction between a red fox and a newborn Critically Endangered green sawfish (Pristis zijsron) and hypothesise that the interaction may be related to the fox opportunistically hunting a sawfish during a time when newborn pups annually enter and aggregate in the relatively ‘safe’ shallow nursery habitats of the Ashburton River delta in the Pilbara region of Western Australia.
Materials and methods
Study area and study species
The study area is located on a remote stretch of unpopulated coastline within a small area of a deltaic island of the Ashburton River estuary in Western Australia. The Ashburton River estuary has been identified as a location where newborn green sawfish are reliably found seeking shallow water refuge from October in each year (Fig. 1) (see Morgan et al. 2015, 2017a; Lear et al. 2023). This globally important nursery for green sawfish was first reported in 2011 (Morgan et al. 2015), not long after the species, classified in 2000 as Endangered by the International Union for the Conservation of Nature (IUCN) (Compagno et al. 2000), was uplisted to Critically Endangered in 2006 (Compagno et al. 2006; Harry et al. 2022). The sawfish nursery is located on a deltaic island, which can only be colonised by foxes at low tide.

Sawfish surveys
In the period 24–29 October 2019, a nursery site identified for green sawfish (Pristis zijsron) (see Morgan et al. 2015, 2017a) was surveyed for neonate sawfish using monofilament gill nets and by deploying cast nets following visual observations. On capture, sawfish were measured for total length (TL, mm), standard rostrum length (SRL, mm) and total rostrum length (TRL, mm), and sexed, with left and right rostral teeth counted. Additionally, other variables such as scarring, missing rostral teeth, presence of a yolk sac scar, girth, parasite prevalence and intensity, as well as depth, location, method, and time of capture were recorded (see Morgan et al. 2017a; Lear et al. 2023; Ingelbrecht et al. 2024). Values are reported as means ± s.e.
To capture neonate sawfish while minimising bycatch, the gill net (60 m length, 150 mm mesh) was set across the shallows in a known nursery habitat of newborn sawfish that occupy very shallow depths (see Morgan et al. 2015). Simultaneously, two researchers armed with head torches surveyed the shallow mangroves during high tide at night to spot sawfish, which were then attempted to be caught using a cast net or by hand grab. During one of these observation times (25 October 2019; ~20:35 hours), a red fox was observed in the spotlight, and quickly left the shallows where a wounded newborn green sawfish was located. The sawfish was captured using a cast net, measured as described above, and scarring, bite marks and damage to the fish recorded.
Results
Sawfish observed
Within the known neonate nursery for green sawfish during the survey period (24–29 October 2019), a total of 14 individual green sawfish pups were captured by gill net (n = 8), cast net (n = 3), or by hand grab (n = 3). The mean TL of the sawfish pups was 857 ± 14 mm TL (range 772–940 mm TL) and consisted of 10 females and four males, with water depth of the pups ranging from 0.1–0.4 m (mean depth of 0.18 ± 0.03 m). Additionally, seven older (>1 year old) sawfish (mean of 1549 ± 159 mm TL, range 1310–2232 mm TL) were captured in the vicinity during the survey period (with all but one captured by gill net and generally in deeper water; mean depth of 0.63 ± 0.29 m). Two of the larger individuals had bite marks most likely attributable to sharks (Fig. 2), while one of the sawfish pups had injuries attributed to an attempted predation event by a red fox (Fig. 3).
Injuries to green sawfish (Pristis zijsron) in the Ashburton River estuary, Western Australia, during the study period (October 2019). These include (a) secondary parasitism by a marine leech (Branchellion plicobrachus) (see Ingelbrecht et al. 2024), (a, b) suspected shark attacks.
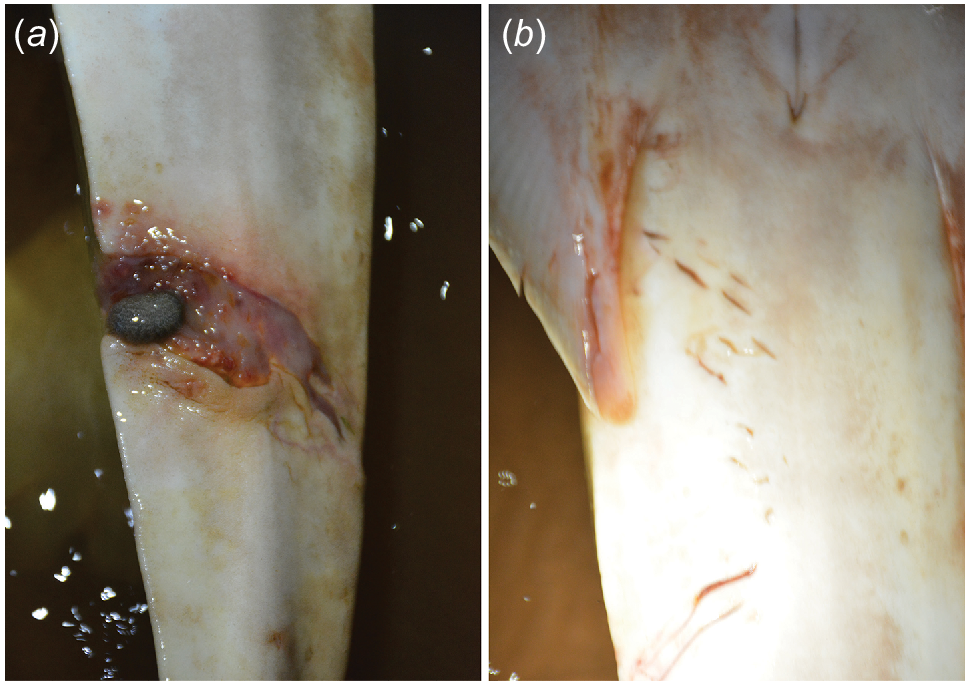
Suspected injuries incurred from a red fox (Vulpes vulpes) attack to the neonate 875 mm TL, female green sawfish (Pristis zijsron) in the Ashburton River estuary, Western Australia, with (a) damage to the rostrum as well as (b) suspected claw marks (horizontal arrows) and scrapes (vertical arrows), and a bite through the dorsal flesh exposing the ventrally located gills. (c, d) Paw prints of red fox at the nursery site.

The fox interaction
On 25 October 2019 at ~20:35 hours during a large tide (high tide of 2.11 m at 21:00 hours for nearby town of Onslow), we disturbed a red fox that scampered from the shallows of the green sawfish nursery hotspot in the Ashburton River mouth. At the location where the fox exited the water, we discovered a neonate female green sawfish of 875 mm TL with severe wounds to its rostrum (SRL = 223 mm) including loss of rostral teeth, as well as paw/claw marks behind the head and a flesh wound through the body from the dorsal surface to the ventrally located gills (Fig. 3). The individual appeared in poor body condition compared to most other neonate sawfish observed at the same time, and although the bite marks on the dorsal surface of the rostrum were fresh, it was not clear if the lateral bites out of the rostral blade were wounds from a previous predation attempt, noting that neonate sawfish spend several months in the area (Morgan et al. 2017a). The attack occurred in mangroves in a depth of ~0.1 m within several metres from the tide line. Six other neonate sawfish were captured adjacent to the attack site between 18:45 hours and 21:40 hours, none of which had markings to suggest that they had been attacked. Fox tracks were observed surrounding the nursery site (Fig. 3), where remains of a green mud crab (Scylla serrata) were found amongst fox paw prints.
Discussion
The distributional ranges of red fox and green sawfish barely overlap on the west coast of Australia, presenting limited opportunity for interactions, which would perhaps be more common should their ranges more widely intersect. There is some distributional overlap between the species in the Red Sea and Persian Gulf, although green sawfish presence is uncertain and less common there (Moore 2015; Dulvy et al. 2016). As an opportunistic invasive predator, it is not surprising that a red fox discovered a location that is visited each year in spring by newborn green sawfish. It is surprising, however, that there are few records relating to red fox predation upon fishes, with none relating to elasmobranchs in marine waters. While previous studies have generally assumed that foxes have consumed fish through scavenging carrion or feeding on stranded animals (Coman 1973; Lugton 1993; O’Connor et al. 2020), there are few records of active predation by foxes on live fish (Tobajas and Díaz-Ruiz 2022a, 2022b).
Although the rostrum of the sawfish was damaged, it is not clear if that was caused by the fox or another predator previously (Fig. 3). The rostrum is a morphological marvel that includes a blade of elongated cranial cartilage that is adorned by small but sharp rostral teeth protruding laterally along the saw (Morgan et al. 2016). The sawfish rostrum, which is vital for prey detection, possesses a well-developed electroreceptive system with ampullary pores located on the dorsal and ventral surface (Wueringer et al. 2011). Damage to the rostrum and loss of rostral teeth is likely to impact not only the ability to detect, capture, and manipulate prey, but also the individual’s capacity to defend itself from predators, such as sharks, which are known to predate on sawfish (Wueringer et al. 2012; Morgan et al. 2017b). Further impact to the individual included paw/claw marks and scrapes behind the head and a hole created from the end of the marks through to the gills (Fig. 3). The degree to which damage was inflicted to the gills, which are critical for respiration, osmoregulation, nitrogenous waste excretion, pH regulation, and hormone production is unknown (Evans et al. 2005). The wound also presents an area for both infection and possible parasitism by marine leeches (Fig. 2) (Ingelbrecht et al. 2024).
Although predation events by red foxes on marine organisms appear to be rare, Grassi (2022) found evidence of marine crabs in red fox scats in the Pilbara (~350 km from our study site), and red fox predation of flatback turtle (Natator depressus) nests along the Pilbara coastline is also common (Grassi 2022; King et al. 2023). Flatback turtles nest adjacent to the sawfish nursery site (between October and February), which attracts foxes to the beaches and estuary mouth during this time. It is plausible that red foxes hunting along the coastline shift their diet to optimise their foraging efficiency during annual aggregations of neonate sawfish and nesting flatback turtles, with increased predation of sawfish focussed during annual aggregations of neonates and flatback turtle nesting. There is further potential for foxes to target other fish species, including elasmobranchs, which use the estuaries and shallow coastal mangroves. For example, neonate giant shovelnose rays (Glaucostegus typus), which are listed as Critically Endangered by the IUCN (Kyne et al. 2019), are prolific along this coast and in the estuary (Bateman et al. 2024; Ingelbrecht et al. 2024). They are often sympatric in the shallows with neonate sawfish. Although morphologically similar and often mistaken as sawfishes, giant shovelnose rays are not afforded the protection of the rostral weaponry of sawfish. It is assumed that larger sawfish, which have larger rostrums, may inflict considerable damage to foxes if subjected to attack (Morgan et al. 2017b). The rostrum has been described as a stealthy weapon with its lateral swipe virtually undetectable to potential prey (and possibly predators) (Bradney et al. 2017). It is suspected that the fox was initially undeterred by the sharp-toothed rostrum and pinned the head and rostrum, presumably to minimise risk of being struck by the rostrum. Alternatively, the sawfish may have been mistaken for a juvenile giant shovelnose ray, which are abundant at the site. It is possible that our presence led the fox to retreat from the sawfish.
In contrast to the spread of the red fox in Australia, green sawfish populations are estimated to have undergone an almost 40% decline in global Extent of Occurrence since the start of the 20th century (Dulvy et al. 2016). Sawfish are invariably threatened by loss of coastal habitat, including mangroves, over-fishing, death as net bycatch from rostrum entanglement, and removal of the rostrum as curios (e.g. Morgan et al. 2016). As the Ashburton River estuary is a globally important nursery for green sawfish (Morgan et al. 2017a), and contains natural sawfish predators, the impact of this additional novel predator should be investigated further. This could involve a monitoring program and appropriate management actions, such as fox control, which may also benefit other native fauna in the area.
Acknowledgements
We acknowledge Paul de Lestang and Steve Moore (Chevron Australia) who assisted with field work and project coordination.
References
Abbott I (2011) The importation, release, establishment, spread, and early impact on prey animals of the red fox Vulpes vulpes in Victoria and adjoining parts of south-eastern Australia. Australian Zoologist 35, 463-533.
| Crossref | Google Scholar |
Basuony M, Saleh M, Riad A, Fathy W (2005) Food composition and feeding ecology of the red fox Vulpes vulpes (Linnaeus, 1758) in Egypt. Egyptian Journal of Biology 7, 96-102.
| Google Scholar |
Bateman PW, Fleming PA (2012) Big city life: carnivores in urban environments. Journal of Zoology, London 287, 1-23.
| Crossref | Google Scholar |
Bateman RL, Morgan DL, Wueringer BE, McDavitt M, Lear KO (2024) Collaborative methods identify a remote global diversity hotspot of threatened, large-bodied rhino rays. Aquatic Conservation: Marine and Freshwater Ecosystems 34, e4047.
| Crossref | Google Scholar |
Bradney DR, Davidson A, Evans SP, Wueringer BE, Morgan DL, Clausen PD (2017) Sawfishes stealth revealed using computational fluid dynamics. Journal of Fish Biology 90, 1584-1596.
| Crossref | Google Scholar | PubMed |
Bubela TM, Dickman CR, Newsome AE (1998) Diet and winter foraging behaviour of the red fox (Vulpes vulpes) in alpine and subalpine New South Wales. Australian Mammalogy 20, 321-330.
| Crossref | Google Scholar |
Castañeda I, Doherty TS, Fleming PA, Stobo-Wilson AM, Woinarski JCZ, Newsome TM (2022) Variation in red fox Vulpes vulpes diet in five continents. Mammal Review 52, 328-342.
| Crossref | Google Scholar |
Coman BJ (1973) The diet of red foxes, Vulpes vulpes L., in Victoria. Australian Journal of Zoology 21, 391-401.
| Crossref | Google Scholar |
Dulvy NK, Davidson LNK, Kyne PM, Simpfendorfer CA, Harrison LR, Carlson JK, Fordham SV (2016) Ghosts of the coast: global extinction risk and conservation of sawfishes. Aquatic Conservation: Marine and Freshwater Ecosystems 26, 134-153.
| Crossref | Google Scholar |
Evans DH, Piermarini PM, Choe KP (2005) The multifunctional fish gill: dominant site of gas exchange, osmoregulation, acid-base regulation, and excretion of nitrogenous waste. Physiological Reviews 85, 97-177.
| Crossref | Google Scholar | PubMed |
Fairfax RJ (2019) Dispersal of the introduced red fox (Vulpes vulpes) across Australia. Biological Invasions 21, 1259-1268.
| Crossref | Google Scholar |
Fleming PA, Crawford HM, Stobo-Wilson AM, Dawson SJ, Dickman CR, Dundas SJ, Newsome TM, O’Connor J, Palmer R, Riley J, Ritchie EG, Speed J, Gentle M, Saunders G, Stuart J-MD, Thompson E, Turpin JM, Woinarski JCZ (2021) Diet of the introduced red fox Vulpes vulpes in Australia: analysis of temporal and spatial patterns. Mammal Review 51, 508-527.
| Crossref | Google Scholar |
Fleming PA, Stobo-Wilson AM, Crawford HM, Dawson SJ, Dickman CR, Doherty TS, Fleming PJS, Newsome TM, Palmer R, Thompson JA, Woinarski JCZ (2022) Distinctive diets of eutherian predators in Australia. Royal Society Open Science 9, 220792.
| Crossref | Google Scholar | PubMed |
Harry AV, Everett B, Faria V, Fordham S, Grant MI, Haque AB, Ho H, Jabado RW, Jones GCA, Lear KO, Morgan DL, Phillips NM, Spaet JLY, Tanna A, Wueringer BE (2022) Pristis zijsron. The IUCN Red List of Threatened Species 2022: e.T39393A58304631. Available at https://dx.doi.org/10.2305/IUCN.UK.2022-2.RLTS.T39393A58304631.en [Accessed 20 December 2024]
Ingelbrecht J, Lear KO, Martin SB, Lymbery AJ, Norman BM, Boxshall GA, Morgan DL (2024) Ectoparasites of the Critically Endangered green sawfish Pristis zijsron and sympatric elasmobranchs in Western Australia. Parasitology International 101, 102900.
| Crossref | Google Scholar | PubMed |
King DR, Smith LA (1985) The distribution of the European red fox (Vulpes vulpes) in Western Australia. Records of the Western Australian Museum 12, 197-205.
| Google Scholar |
King J, Whiting SD, Adams PJ, Bateman PW, Fleming PA (2023) Camera traps show foxes are the major predator of flatback turtle nests at the most important mainland western Australian rookery. Wildlife Research 51, WR22109.
| Crossref | Google Scholar |
Kyne PM, Rigby CL, Dharmadi, Gutteridge AN, Jabado RW (2019) Glaucostegus typus. The IUCN Red List of Threatened Species 2019: e.T104061138A68623995. Available at https://dx.doi.org/10.2305/IUCN.UK.2019-2.RLTS.T104061138A68623995.en [Accessed 20 December 2024]
Lear KO, Fazeldean T, Bateman RL, Inglebrecht J, Morgan DL (2023) Growth and morphology of Critically Endangered green sawfish Pristis zijsron in globally important nursery habitats. Marine Biology 170, 70.
| Crossref | Google Scholar |
Lugton IW (1993) Diet of red foxes (Vulpes vulpes) in south-west New South Wales, with relevance to lamb predation. Rangeland Journal 15, 39-47.
| Crossref | Google Scholar |
Meek PD, Triggs B (1998) The food of foxes, dogs and cats on two peninsulas in Jervis Bay, New South Wales. Proceedings of the Linnean Society of New South Wales 120, 117-128.
| Google Scholar |
Moore ABM (2015) A review of sawfishes (Pristidae) in the Arabian region: diversity, distribution, and functional extinction of large and historically abundant marine vertebrates. Aquatic Conservation: Marine and Freshwater Ecosystems 25, 656-677.
| Crossref | Google Scholar |
Morgan DL, Allen MG, Ebner BC, Whitty JM, Beatty SJ (2015) Discovery of a pupping site and nursery for critically endangered green sawfish Pristis zijsron. Journal of Fish Biology 86, 1658-1663.
| Crossref | Google Scholar | PubMed |
Morgan DL, Wueringer BE, Allen MG, Ebner BC, Whitty JM, Gleiss AC, Beatty SJ (2016) What is the fate of amputee sawfish? Fisheries 41, 71-73.
| Crossref | Google Scholar |
Morgan DL, Ebner BC, Allen MG, Gleiss AC, Beatty SJ, Whitty JM (2017a) Habitat use and site fidelity of neonate and juvenile green sawfish Pristis zijsron in a nursery area in Western Australia. Endangered Species Research 34, 235-249.
| Crossref | Google Scholar |
Morgan DL, Somaweera R, Gleiss AC, Beatty SJ, Whitty JM (2017b) An upstream migration fought with danger: freshwater sawfish fending off sharks and crocodiles. Ecology 98, 1465-1467.
| Crossref | Google Scholar | PubMed |
O’Connor JM, Srivastava SK, Tindale NW, Burnett SE (2020) From carrion to Christmas beetles: the broad dietary niche of the red fox in a hybrid coastal ecosystem in south-eastern Queensland. Australian Journal of Zoology 67, 82-93.
| Crossref | Google Scholar |
Ryan GE, Croft JD (1974) Observations on the food of the fox, Vulpes vulpes (L.), in Kinchega National Park, Menindee, NSW. Wildlife Research 1, 89-94.
| Crossref | Google Scholar |
Stobo-Wilson AM, Murphy BP, Legge SM, Caceres-Escobar H, Chapple DG, Crawford HM, Dawson SJ, Dickman CR, Doherty TS, Fleming PA, Garnett ST, Gentle M, Newsome TM, Palmer R, Rees MW, Ritchie EG, Speed J, Stuart J-M, Suarez-Castro AF, Thompson E, Tulloch A, Turpin JM, Woinarski JCZ (2022) Counting the bodies: estimating the numbers and spatial variation of Australian reptiles, birds and mammals killed by two invasive mesopredators. Diversity and Distributions 28, 976-991.
| Crossref | Google Scholar |
Tobajas J, Díaz-Ruiz F (2022a) Fishing behavior in the red fox: opportunistic-caching behavior or surplus killing? Ecology 103, e3814.
| Crossref | Google Scholar | PubMed |
Tobajas J, Díaz-Ruiz F (2022b) Unrevealing the opportunistic fishing behavior of the red fox. Bulletin of the Ecological Society of America 103, e2021.
| Crossref | Google Scholar |
Triggs B, Brunner H, Cullen JM (1984) The food of fox, dog and cat in Croajingalong National Park, south-eastern Victoria. Wildlife Research 11, 491-499.
| Crossref | Google Scholar |
Wagnon CJ, Serfass TL (2017) Use of camera traps provides insight into the feeding ecology of red foxes (Vulpes vulpes). The Canadian Field-Naturalist 131, 19-22.
| Crossref | Google Scholar |
Wueringer BE, Peverell SC, Seymour J, Squire L, Jr, Kajiura SM, Collin SP (2011) Sensory systems in sawfishes. 1. The ampullae of Lorenzini. Brain Behavior and Evolution 78, 139-149.
| Crossref | Google Scholar | PubMed |
Wueringer BE, Squire L, Kajiura SM, Hart NS, Collin SP (2012) The function of the sawfish’s saw. Current Biology 22, R150-R151.
| Crossref | Google Scholar | PubMed |