

Longitudinal trends of future suitable climate for conserving oil palm indicates refuges in tropical south-east Asia with comparisons to Africa and South America
R. Russell M. Paterson A B C
A B C
A Centre of Biological Engineering (CEB), Gualtar Campus, University of Minho, Braga 4710-057, Portugal.
B Department of Plant Protection, Faculty of Agriculture, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor DE, Malaysia.
C Corresponding author. Email: russell.paterson@deb.uminho.pt
Pacific Conservation Biology 28(1) 57-67 https://doi.org/10.1071/PC20067
Submitted: 26 August 2020 Accepted: 8 March 2021 Published: 13 April 2021
Abstract
Palms are crucial species to the flora of south-east (SE) Asia. Oil palm (OP) is a highly significant introduced species to the region and produces palm oil, an especially important commodity. OPs are grown in huge plantations in SE Asia, predominantly in Malaysia and Indonesia. These palms have detrimental effects on the environment, particularly from deforestation when creating new plantations. Future climate scenarios have indicated mostly reductions in suitable climate for growing OP throughout SE Asia and it is crucial to consider how the palms can be conserved. Climate change has been shown to stimulate poleward movements in some other species, which assist in conserving them and may permit assisted colonisation. However, poleward movement to these refuges is unlikely from the tropics. Recent research has indicated potential longitudinal refuges for OP in Africa and phased longitudinal refuges in South America, based on future suitable climate (SC) for growing OP. These data indicate how the sustainability of OP could be maintained rather than experiencing the complete decimation of the crop, which might otherwise occur. This current work was undertaken to determine trends of future SC for growing OP in SE Asia and to indicate refuges with implications for conservation and plantation management. The results are compared with those for Africa and South America. The longitudes considered in SE Asia were from 100°E to 146°E, which is an intentionally large area of land. The maps obtained from the climate modelling program CLIMEX, of climate suitability for growing OP, were employed to obtain the percentage SC in SE Asian regions and countries for current time (CT), 2050 and 2100. The average percentage SC decreased for 2050 and further for 2100. Increasing longitudinal trends in SC for growing OP were observed from CT to 2050, CT to 2100 and 2050 to 2100 from west to east. Vietnam, the Philippines, Papua New Guinea (PNG) and island Malaysia had increased SC by 2050. Large decreases in SC by 2050 for Thailand, Laos and Cambodia, which are towards the west of SE Asia, were observed. There was an increasing trend in suitable climate from CT to 2100 and a smaller trend from 2050 to 2100. Hence, OP may find suitable refuges by natural seed spreading towards the east of SE Asia, hence avoiding extinction. Similarly, new plantations could be established in the same direction, although environmental concerns are paramount. Vietnam, the Philippines, PNG and island Malaysia may become more suitable than Thailand, Laos and Cambodia. The sustainability of OP plantations is likely to decrease substantially with climate change. However, there is scope for a more sustainable situation towards the east of SE Asia. New plantations cannot be established without considering the severe effects on the environment from deforestation and increased greenhouse gases. Overall, urgent action is required to reduce the effects of climate change.
Keywords: climate maps, Elaeis guineensis, global warming, greenhouse gasses, Indonesia, longitude, moisture, Vietnam.
Introduction
Palms are iconic and highly significant to maintaining biodiversity, the conservation of which may be threatened by climate change (Cosiaux et al. 2018; Paterson 2021a). Oil palm (Elaeis guineensis) (OP) is perhaps the most economically important palm for palm oil production and huge OP plantations exist in south-east (SE) Asia. Palm oil manufacturing is dominated by Malaysia and Indonesia, which produce 85% of the world’s total (Basiron 2007; Ommelna et al. 2012), giving highly significant advantages to the economies of producing countries. However, OP cultivation is particularly disadvantageous to the environment because of deforestation during the creation of new plantations (Paterson and Lima 2018). OP, the oil and its production are associated with human health problems (Kadandale et al. 2019), including zoonotic disease in humans from conversion of natural forest (Gibb et al. 2020), adding to the overall significance of OP.
The multitude of uses of palm oil has been documented by, inter alia, the current author (Paterson et al. 2017; Paterson 2021a, 2020b, 2020c, 2020d). OP is increasingly being considered as a crop in other tropical countries as land becomes scarce (Villela et al. 2014) and climate becomes unconducive to growth (Marlier et al. 2014; Paterson 2020a, 2020b) in the countries in its original distribution in SE Asia. Many countries in SE Asia (Arrieta et al. 2007; Ohimain and Izah 2014; Paterson et al. 2015), Africa (Paterson 2021a) and South America (Paterson 2021b) have plans to increase the amount/area of OP grown (Corley and Tinker 2015). Alternative SE Asian states could, in theory, increase their market share as land in Malaysia and Indonesia becomes scarce (Paterson and Lima 2018; Paterson 2020a, 2020b).
Palms are fundamental to the tropics, providing essential ecosystem services to rural people worldwide, although many are threatened by overexploitation and habitat loss. Suitable climate for growing OP was determined in Indonesia and Malaysia using modelling techniques that employed temperature, soil moisture and wet stress (Paterson et al. 2015). In general, there was a decrease in suitable climate. That paper addressed the convention of only temperature being employed by considering additional parameters, thereby increasing the significance of Paterson et al. (2015). Paterson et al. (2017) extended this approach globally, including other countries in SE Asia. It was apparent that most of these countries would be affected detrimentally by the changes. The general trend was for a reduced level of suitable climate in 2050, further reduction by 2070 and even more by 2100. The projections indicate serious consequences to the palm oil industry generally as the climate becomes unsuitable for growing OP. Maps of the changes in suitability are provided in Fig. 1. It was anticipated that more useful, granular detail could be obtained from these data, which is the theme of the current paper.

|
SE Asia also provides excellent growth conditions for other palms such as coconut (Perera et al. 2016) and sago (Lim et al. 2020) palms. All palms (Cosiaux et al. 2018) will be affected by climate change and may not survive, although there appears to be a low extinction risk in the immediate future (Paterson et al. 2015, 2017; Sarkar et al. 2020). Paterson (2020b) described high OP mortalities in Brazil, Colombia, Ecuador, Malaysia and Indonesia, and Paterson (2020a) indicated from high to low levels of OP mortality throughout SE Asia depending on country or region. Paterson (2020c, 2020d) discovered the mortality of OP in the various regions of Malaysia and Indonesia respectively. Furthermore, Paterson (2021a, 2021b) determined longitudinal trends for suitable climate for growing OP in Africa and South America, and Paterson (2021a), in particular, emphasised that the OP data could be extrapolated to palms in Africa. It is crucial to understand the climate change threats to these plants (Cosiaux et al. 2018). Future climate change may disrupt many ecosystems in general, which is often manifested by phenology (Thackeray et al. 2010). There may be demographic changes in suitable climate for many species (Rosenzweig et al. 2008), including OP (Paterson et al. 2015, 2017; Paterson 2021a, 2021b). Extinction rates (Sinervo et al. 2010) will accelerate with the intensification of climate change (Urban 2015). Often computer models are used in climate studies, although Bellard et al. (2012) mentioned that not all relevant parameters are considered, whilst stating that socioeconomic ‘storylines‘ are also employed and, indeed, high levels of ecological changes caused by climate change are deduced by employing scientific inference (Parmesan 2006). Most studies make correlations between current climate and species distributions to novel conditions and imply that temperature-sensitive species can extend their ranges to cool refuges (Wright et al. 2011).
Tropical species, such as OP, are included infrequently in climate change studies. Precipitation is often not considered, with the emphasis being on temperature: equally, latitudinal changes to range in temperate species are reported, but not longitudinal ones (Feeley et al. 2017; Fei et al. 2017), thereby underestimating the effect of climate change even in these species. However, latitudinal changes for tropical plants are often impossible (Wright et al. 2011). Successional processes were more sensitive to moisture availability than to temperature for trees, where a longitudinal effect was also reported (Fei et al. 2017): Cho and McCarl (2017) found that climate explained a longitudinal trend to favour crops. Miller et al. (2017) determined that forest responses to climate change were apparent near the western range limit of white spruce in Alaska. Nevertheless, trees were limited in their ability to track climate warming, emphasising the negative impact of climate change on forests (Sittaro et al. 2017). Paterson (2021a) determined longitudinal trends of increased suitable climate from west to east Africa, and Paterson (2021b) determined three phased longitudinal trends in South America where the trend was not from coast to coast. These refuges are extremely important to conserve OP per se and for the creation of novel plantations. They will assist in providing long term sustainability to the industry compared with the situation whereby the OP cannot survive in future climate conditions. Poleward migrations, as a survival mechanism for most equatorial species, is impossible because of the absence of latitudinal temperature gradients (Wright et al. 2011) and tropical forest species, such as OP, will be particularly sensitive to global warming. Paterson (2020a, 2020b, 2020c, 2020d) indicated a high degree of unsuitable climate for OP and mortalities of OP, from future climate change. Extension of temperature sensitive OP might extend ranges to cool refuges, although poleward distances are greatest for equatorial species and to the upper elevations of many tropical mountains. Tropical species are sensitive to global warming because they are adapted to limited geographic and seasonal variation in temperature. They live at or near the highest temperatures on Earth before global warming began and are isolated from cool refuges (Lenoir and Svenning 2015). A strong latitudinal temperature gradient averages 1°C for each 145-km change in latitude between the tropics and the polar regions, but approaches zero between the Tropics of Cancer and Capricorn. It is imperative to consider what other phenomenon could occur that would enable these species to adapt, such as the occurrence of longitudinal refuges of suitable climate.
As mentioned, climate change in the tropics is poorly studied (Ghini et al. 2011; Feeley et al. 2017) and investigations covering large areas of land are lacking. Data on thermoregulatory behaviours are non-existent for tropical lowland plants, with the exception now of Paterson (2021a, 2021b), which involve considering soil moisture and wet stress data in addition to temperature. This lack of information is especially unfortunate because tropical species will have fundamentally different responses to climate change than temperate counterparts. Prioritising tropical plants is crucial since there are no pools of plants living in a hotter climate to replace declining species in tropical lowlands, leading to tropical lowland biotic attrition (Lenoir and Svenning 2015; Feeley et al. 2017). The high levels of OP cultivation that do exist also contribute to reduced ecosystem function (Dislich et al. 2017).
The plants are susceptible to unsuitable future climate as discussed for Africa (Paterson 2021a) and South America (Paterson 2021b). African palm species will experience a decline in climatic suitability in >70% of their current ranges by 2080 (Blach-Overgaard et al. 2015). Losses of palm habitats such as tropical rain forests are increasing the threat to palm populations where OP cultivation involves deforestation (Paterson and Lima 2018) and hence the development of OP could threaten other palms. It is fundamentally important to comprehend how climate change will affect palms given their socioeconomic and ecological importance. Blach-Overgaard et al. (2015) predicted climate suitability losses across almost all terrains where palms occur in Africa, from where OP originated. Paterson (2021a, 2021b) confirmed these loses for OP and palms in general, in Africa and South America respectively. Unusually for palms, OP can grow in open habitat and in forests and will be affected in these regions. Hence, OP may be a model for palms that grow in either of the two locations. A decline was projected in climate suitability in 70% of their current predicted ranges by 2080 for all palms: many could track suitable climate space by dispersing within 100-km distance of their current range margins. Furthermore, climate change threatens other crop production (Lobell et al. 2006). Large areas for maize cultivation will be subjected to heat and dry stresses that may constrain production (Ramirez-Cabral et al. 2017). Future climate could favour the plant pathogenic fungus Fusarium oxysporum by threatening greater infection of crops (Shabani et al. 2014) and Paterson (2020a, 2020b, 2020c, 2020d) modelled increased mortality and disease of OP due to climate change. These reports indicate strongly that the sustainability of the industries that these crops represent will be negatively affected.
The current paper considers how suitable climate for the growth of OP changes across most SE Asian regions and countries (RAC) based on the information in Paterson et al. (2017) and in a manner equivalent to that for Africa (Paterson 2021a) and South America (Paterson 2021b). There is a paucity of information on how climate change affects (1) large areas such as SE Asia, (2) tropical countries, (3) other parameters in addition to temperature, and (4) longitudinal migration, all of which the current paper addresses.
Material and Methods
Most of the methods employed in this study are published in Paterson (2021a, 2021b) for Africa and South America and are reproduced herein in a modified form, but in some detail, for convenience.
Oil palm distribution
The Global Biodiversity Information Facility (http://www.gbif.org/, accessed 31 July 2020) and additional literature on the species in CAB Direct (http://www.cabdirect.org/web/about.html, accessed 31 July 2020), provided the distribution of E. guineensis (Paterson et al. 2017) and the raw data for the analysis in Paterson (2021a, 2021b). In total, 2445 records were utilised in fitting the parameters. The current distribution of palm oil is shown in Fig. 1.
Climate data and future projections
CliMond 10′ gridded climate data (Paterson et al. 2017) were employed to compare the ability of mechanistic and correlative bioclimatic modelling methodologies. Parameters of climate incorporated in the meteorological database were the mean monthly temperature maxima and minima, mean monthly precipitation level and relative humidity at 0900 hours and 1500 hours. The same parameters were also used to project the possible future climates.
Scenario of climate change
The current study used CSIRO-Mk3·0, MIROC-H (SRES) and IPCC 2000 to model potential future distribution of OP, as described in Paterson et al. (2017), because they incorporate economic, technological and demographic factors that affect global greenhouse gas (GHG) emissions. Moderate GHG emissions were assumed by 2100. The date of 2100 was chosen since it assumes moderately increasing GHG, based on the midpoint of lower- and higher-level projections.
CLIMEX model
CLIMEX software supports ecological research by incorporating the modelling of species’ potential distributions under differing climate scenarios and assumes that climate is the paramount determining factor of plant and poikilothermic animal distributions (Paterson et al. 2017). Hence, other lesser factors are not considered in this study. CLIMEX enables the determination of geographically relevant climatic parameters describing the responses of an organism to climate. It models mechanisms that impose limitations to the geographical distribution of species and determines seasonal phenology and abundance. Species growth potential in the favourable season is denoted by the Annual Growth Index (GIA), while the impact of population reduction during an unfavourable season is established by the cold, hot, wet and dry Stress Indices and their interactions.
The product of the GIA and Stress Indices is the Ecoclimatic Index (EI), which rates the level of suitability for species’ occupation of a particular location or year. The EI is thus an annual average index, derived from weekly data of the growth and stress indices of suitability levels of climatic factors, denoted by a value on the scale 0 to 100, and a species may be established where EI > 0. CLIMEX output categorises areas according to high suitability, suitability and marginal suitability based on other studies (Paterson et al. 2017) where temperature index (limiting low temperature, lower optimal temperature, upper optimal temperature, limiting high temperature), moisture index (limiting low soil moisture, lower optimal soil moisture, upper optimal soil moisture, limiting high soil moisture), cold stress (cold stress temperature threshold, cold stress temperature rate, minimum degree-day cold stress threshold, degree-day cold stress rate), heat stress (heat stress temperature threshold, heat stress temperature rate), dry stress (dry stress threshold, dry stress rate), wet stress (wet stress threshold, wet stress rate) and degree-day threshold were fitted according to global distribution data, iteratively adjusted to achieve satisfactory agreement between known and projected species’ distributions globally. Detailed justification of these parameters, and how they were obtained, are provided in Paterson et al. (2015). Paterson et al. (2017) provided detailed parameter values used in OP modelling.
Granular map examination
Paterson et al. (2017) employed the CLIMEX model to provide scenarios of suitable future climate for growing OP in SE Asia for current time, 2050 and 2100 (Fig. 1) by magnification of the maps where necessary to determine the percentage of each climate category. Percentages of highly suitable and suitable climates were determined visually from the red and yellow areas in each map and combined to give the combined suitable climate (CSC) parameter: red represented highly suitable climate and yellow the suitable climate for growing OP. The change in CSC from future climate was determined by subtracting the CSC values from 2050 or 2100 from those of CT. This indicated the effect of climate change on the suitability for growth of OP. The percentage CSC was plotted against the longitudes of the various countries and regions and the trend line was added using Excel software.
Myanmar has a large subtropical component to its climate (Zune et al. 2020) and, as such, is different from the remainder of SE Asia, which is all tropical. It also has only small areas suitable for OP growth (Paterson et al. 2017; Paterson 2020a), again somewhat atypical of SE Asia. Hence, this country is not included in the current analysis. The longitude of each RAC was determined by using the average of the most westerly and easterly longitudes of the individual RAC. The longitudes obtained for each RAC are provided in the captions of Figs 2–9.
|
|
|
|
|
|

|
|
Results
Maps of the suitability for OP growth in SE Asia are shown in Fig. 1 and the complete set are available in Paterson et al. (2017). A general decrease in CSC was observed until 2100. However, additional trends are discussed below that are not apparent from superficial inspection of the maps.
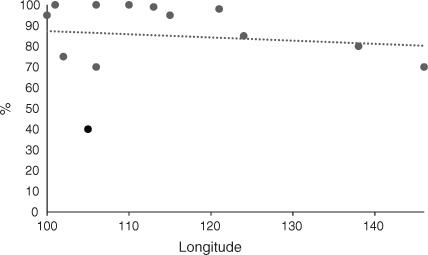
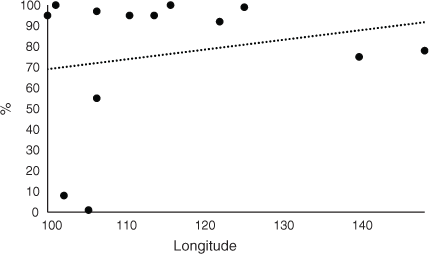
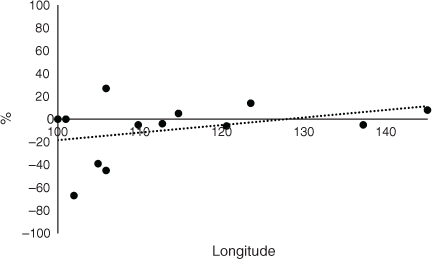
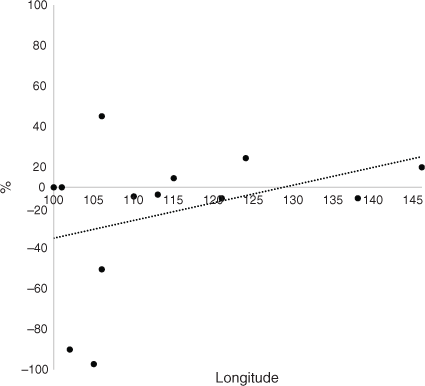
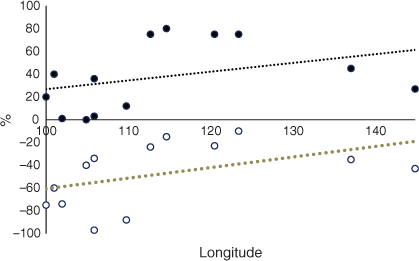
The average CSC in CT for all RAC was 85%. Fig. 2 shows that Laos had much lower levels of CSC (40%) and the other RAC had high percentages of CSC. The trend line in Fig. 2 demonstrates that there was a slight decrease in CSC from west to east. The average CSC in 2050 for all RAC was 76%. Laos and Thailand had exceptionally low levels of CSC (Fig. 3), Cambodia had 55%, and the other RAC had high levels (75% or more). The trend line for CSC in 2050 indicates a large increase from west to east from 70% to 88% at longitudes 100°E and 146°E, which contrasted with the decrease at CT (Fig. 2). The change in CSC from future climate from CT to 2050 indicated that there was no change for Sumatra and Peninsular Malaysia, but particularly large decreases for Thailand, Laos and Cambodia of –67%, –39% and –45% respectively (Fig. 4). Vietnam, island Malaysia (Sabah and Sarawak), the Philippines and Papua New Guinea (PNG) had increases in CSC by 2050 of 27%, 5%, 14% and 8% respectively. The other RAC remain little changed by 2050 compared with CT. The trend line indicated a range of CSC for this period of –20% to 9% from 100°E to 146°E and the line crosses to positive values at ~127°E. A greater rate of change was apparent when the percentage changes in the original areas of CSC at CT by 2050 are considered (Fig. 5). The trend line was from –29% to 16% from 100°E to 146°E.
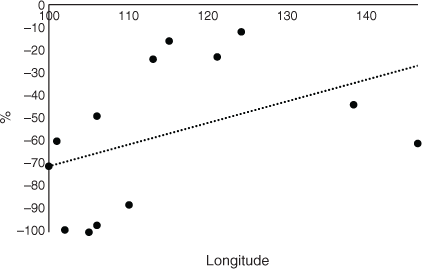
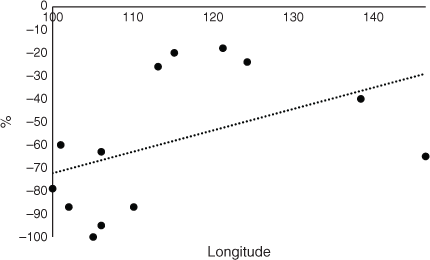
The CSC for the 2100 data are provided in Fig. 6 (upper data) and the average CSC for 2100 was 38%, which is much lower than for 2050 or CT. Kalimantan, island Malaysia, Sulawesi, and the Philippines maintained a high level of CSC of ~78% for 2100. The change in CSC from CT to 2100 is also represented in Fig. 6 (lower data). As for 2050, an increasing trend was observed from west to east, although larger decreases were observed for Papua and PNG in the most easterly regions, somewhat against trend. All the differences in CSC between CT and 2100 had negative values. There was a very steep increase from west to east, again indicating a strong longitudinal trend by 2100 when the percentage change in CSC areas from current time was considered (Fig. 7). There was a small increasing trend from west to east when the CSC for 2050 was compared with that for 2100 and all the values were negative by 2100 (Fig. 8). When the change in areas from 2050 to 2100 was considered, a steep trend was observed with increasing CSC from west to east consistent with the previous data (Fig. 9).
Discussion
In general, the maps revealed a decrease in CSC from current time to 2100 with some indication that the suitable climate decreased most towards the west of SE Asia (Fig. 1). However, it was impossible to determine more detail without resorting to the methods described herein and represented in Figs 2–9.
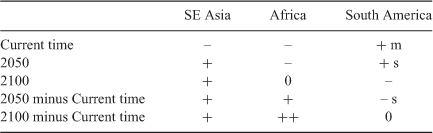
The CSC throughout SE Asia was high in current time (Fig. 2), indicating the general suitability of climate for OP in terms of the parameters employed in the model. These parameters are the predominant factors that determine whether OP will survive, as mentioned in the Materials and Methods. Other factors such as soil type or slope of the land for cultivation are not considered. There was a slight decreasing west to east trend from Sumatra (100°E longitude) to PNG (146°E longitude). There were rapidly decreasing CSC for longitudes 102°E, 105°E and 106°E, representing Thailand, Laos and Vietnam, which was specific for these countries. Overall, there was a decreasing trend (Table 1). To compare these data with Africa, high percentages of CSC were observed for current time in west Africa, which decreased rapidly in the countries further east (Paterson 2021a), representing a large decreasing trend from west to east. The equivalent data for South America revealed a moderately increasing trend from west to east although there was a large decrease in CSC towards the centre of the continent. Hence, the decreasing trends for SE Asia and Africa were reversed for South America (Table 1).
|
The longitudinal trend of CSC for 2050 in SE Asia revealed the opposite trend from current time (Fig. 3), with an increase from west to east (Table 1). Particularly obvious decreases were observed for Thailand and Laos, whereas the other countries did not change greatly. The trend for Africa was a decrease from west to east in 2050, although the slope was less than for current time (Paterson 2021a). In the case of South America, a slight increasing trend from west to east was observed, which was the same for CT. Hence the change in trend in 2050 for SE Asia was increase from west to east, which was similar to South America whereas that for Africa was a decrease from west to east (Table 1), representing another difference between CT and 2050.
When the change in CSC from climate in 2050 was considered there was no change for Sumatra and Peninsular Malaysia, but large decreases in the SE Asian peninsula where Thailand, Laos and Cambodia were observed (Fig. 4). These countries are more towards the west of SE Asia than most of the other RAC and strongly influence the increasing longitudinal trend. However, Vietnam, which is also on the peninsula, had a large increase in CSC (27%). An increasing trend of CSC from west to east on the mainland can be observed (Fig. 4) and the exceptionally high figure for Vietnam may relate to the influences of the South China Sea and the Gulf of Tonkin off the coast of the country. The large increase in CSC in 2050 compared with CT for Vietnam indicates clearly how climate change could increase the suitability for growing OP, depending on other factors such as land degradation, deforestation, soil erosion and unsustainable ground water extractions (Blach-Overgaard et al. 2015). Island Malaysia (Sabah and Sarawak), the Philippines and PNG also had increases in CSC by 2050 and were towards the east of SE Asia and so may represent refuges for OP.
The trend line for the difference in CSC between 2050 and CT (Fig. 4) demonstrates an increasing trend in suitable climate from west to east (Table 1) and indicates how migration of OP to refuges could occur naturally within this period by, for example, the spread of seeds. The industry may be more sustainable if plantations are concentrated on the more easterly RAC until 2050, in terms of OP management. Also, similar events could occur with the other species of palms within the region insofar as OP represents a model for other palms. OP may find refuges towards the east, especially in RAC where there was positive change by 2050. The trend line for the difference in CSC between 2050 and CT for Africa (Paterson 2021a) was similar to that for SE Asia (Fig. 4). The trend line for Africa was from –22% in the west to ~9% in the east. For SE Asia, the trend line was from –20% in the west to ~10% in the east. However, the trend line for South America had only a slight decreasing trend from approximately –20% in the west to –23% in the east Paterson (2021b). Hence, the effect of climate change on suitable climate is similar in SE Asia and Africa and quite different to that for South America (Table 1).
Interestingly, the islands that lie beyond the Philippines but before Papua, such as Halmahera and Ceram, could experience an increase for CSC by 2050, because their average longitude is 128°E. These islands may be refuges for OPs, or form the basis of new plantations. It cannot be overemphasised that the effect on the environment of new plantations is of paramount importance and must be taken into consideration well before development of new plantations. In addition, suitable soil and slope of cultivation are likely important, as are socioeconomic factors when establishing novel plantations. It is interesting that when the percentage of original area that has changed is considered (Fig. 5) there is a greater increasing trend due to climate change compared with simply the difference between 2050 and CT (Fig. 4).
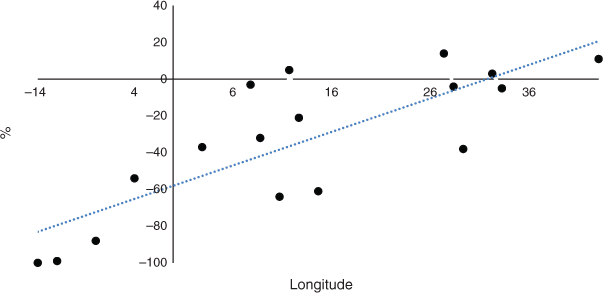
Kalimantan, island Malaysia, Sulawesi, and the Philippines maintained a high level of CSC of ~78% in 2100 and may be able to maintain an OP industry until 2100 based on this data (Fig. 6, upper data). However, the overall reduction in CSC was large. Other factors such as disease may affect the palms (Paterson 2019a, 2019b, 2020a, 2020b, 2020c, 2020d). There were also increasing west to east trends in CSC. In Africa, there was no trend in change of CSC (Paterson 2021a) and for South America there was a decrease in CSC from west to east by 2100 (Paterson 2021b) (Table 1). All the values were negative when the change in CSC from future climate by 2100 was considered (Fig. 6, lower data). There remained a positive trend from west to east and there might be more scope for sustainable OP towards the east of the continent even by 2100. In fact, only small decreases were observed after Kalimantan (113°E) to Papua (138°E), providing scope for OP survival and maintenance of the industry. There was virtually no longitudinal trend for South America for the equivalent data (Paterson 2021b) although there was a large increase in CSC toward the centre of the continent. This parameter was not determined for Africa in Paterson (2021a) but is presented here in Fig. 10 for comparison purposes and is derived from the results in Paterson (2021a). This demonstrates that there was a sharp increasing trend from west to east and to a greater extent than for SE Asia (Table 1).
|
The results presented herein are based only on suitable climate for OP although this is the predominant factor for determining successful survival (see Material and Methods). Other factors will affect whether OP can be established in novel locations. These may include (1) soil type, (2) the land may be being used for another purpose, (3) lack of other resources, such as workers or appropriate infrastructure, (4) extreme slope of the land on which the OP could not grow and/or (5) the high occurrence of diseases and pests of OP (Paterson et al. 2015, 2017; Paterson 2020a, 2020b, 2020c, 2020d).
Blach-Overgaard et al. (2015) suggested assisted colonisation to promote species tracking climate change and the results presented herein indicate that the more easterly RAC may be appropriate for this purpose in SE Asia. The range limits of OP distribution are defined by climate by (1) exerting ecophysiological constraints and (2) soil property and biotic interactions (Paterson et al. 2017). Cold, heat and dry stresses were highly responsible for the changes in climatic suitability for OP cultivation, whereas wet stress was unimportant (Paterson et al. 2017). A significant negative relationship was determined between annual average temperature, sea level rise and OP production in Malaysia (Sarkar et al. 2020) where temperature rises of 1–4°C could decrease OP production by 10–41%.
The information provided herein indicates how OP could survive climate change by the creation of refuges through tracking beneficial climate changes towards the east of SE Asia. Equally, novel areas for plantations may become available and areas to avoid can be determined. Before plantations are initiated the environmental issues raised require careful consideration. The effects of climate change on natural systems must be understood to predict or mitigate changes in diversity and ecosystem function (Feeley et al. 2017). Anthropogenic climate change can impact all species and ecosystems and large scale conversion of tropical forest to OP plantations has detrimental effects on biodiversity. Fitzherbert et al. (2008) determined that OP plantations support many fewer species than forests and some other tree crops: habitat fragmentation and increased pollution occur, including GHG emissions. OP cultivation affects climate by increasing GHG emissions and consequently climate change (Paterson and Lima 2018). Koh and Wilcove (2008) suggested that OP expansion occurs at the expense of forests, which may result in biodiversity loss. Dislich et al. (2017) determined that 11 of 14 ecosystem functions decreased in levels of function by the introduction of OP plantations. Hence, countries that experience less severe change in climate, or positive effects, cannot develop plantations at the expense of the environment. Latitudinal refuges may not be accessible in the tropics, as mentioned above (Wright et al. 2011). For example, OPs could not be displaced to countries such as Laos and Cambodia but could be displaced to the Philippines and Vietnam.
OP are a marker species that indicate how other palms could adapt, especially as OP grows in rainforest and open plain, hence representing all types of palms which normally grow in only one of these locations (Blach-Overgaard et al. 2015). The results are important because there are few data otherwise concerning climate change effects on (1) tropical species and (2) longitudinal effects and there are few reports that take into account other parameters apart from temperature (Paterson et al. 2017).
In terms of refuges and novel plantations, it is useful to consider where there are stresses within SE Asia at higher and lower latitudes and longitudes (Paterson et al. 2017). Central (1) Papua and (2) PNG will have regions of cold stress for OP by 2050 within SE Asia, otherwise the region is free from cold, dry and heat stress in the future. This stress in Papua and PNG may partially explain the atypically low CSC values described above for the region and country. There is cold stress apparent in China to the north of SE Asia and so OP could not readily migrate there. A specific region in the north of Australia appears suitable for OP but further south there are large regions with dry stress in 2050. There are smaller areas of hot stress toward the west of Australia by 2050.
Furthermore, there are areas of heat stress within SE Asia in countries such as Thailand and especially Cambodia by 2100 (Paterson et al. 2017) and the cold stress in Papua and PNG becomes greatly reduced by 2100. A small amount of heat stress was determined in Java. Myanmar and southern China have less cold stress than in 2050 and these may become more suitable for OP. However, in northern Australia a large area of heat stress was apparent. Overall, there are few areas that offer refuges in poleward directions. Correlating these stresses with climate suitability is not absolute for complications from, inter alia, (1) growing season length, (2) vernalisation and (3) other stress combinations, e.g. cold–dry, hot–dry, cold–wet and hot–wet. Also, although some regions develop fewer stress factors in the future, such as the lack of cold stress in southern China by 2100, these regions may not become suitable for OP growth for other reasons, as previously mentioned. There are some indications that southern China will become moderately more suitable (Paterson et al. 2017). On the other hand, there is no indication that northern Australia will become suitable for growing OP, as indicated by the maps, despite there being no stress in northern Australia in 2050. OP would not find a refuge in this location. Wright et al. (2011) indicate that distances to latitudinal cool refuges are particularly large in Africa, which may also apply to SE Asia.
As well as future climate, diseases will also affect growth of OP in the future. Paterson et al. (2013) considered it likely that fungal and fungal-like diseases of OP would increase under climate change. Paterson (2019b) assessed that basal stem rot (BSR) of OP in Malaysia would be a considerable threat to sustainability in Peninsular Malaysia, but less so in Sabah and Sarawak (i.e. island Malaysia) after 2050. Hence, island Malaysia may have a sustainable OP industry, as suggested also by the results in the current paper. Although the CSC of Sumatra does not change a great deal by 2050 in the results presented herein, small increases in BSR were observed in most parts of the island (Paterson 2019a). Large increases in BSR were determined after 2050 until 2100, coinciding with large decreases in suitable climate for growing OP by 2100, as determined in the current paper. Kalimantan and the Philippines had low incidences of BSR (Paterson 2020b), indicating that they may be sustainable compared with the other RAC considered in the current paper. However, PNG, and particularly Thailand, had large incidences of BSR with further severe problems for these countries. The potential for the acute form of Phytophthora palmivora disease was very high in Malaysia and Indonesia even by 2050 (Paterson 2020a) and the levels of the acute and chronic forms were extremely high by 2100. Hence, the potential for this disease is high even in Indonesia and Malaysia, which had high CSC, as reported in the present paper. Paterson (2020c) discussed quantitatively the effect of future climate on BSR in Malaysia. Low levels of BSR were determined for Sabah by 2100 which represented the most sustainable region in Malaysia for PO production followed by Sarawak. Climate change will affect the incidence of BSR particularly after 2050. A similar treatment was undertaken for Indonesia in Paterson (2020d). Sumatra and Java were especially affected by BSR and plantations may be unsustainable quickly, whereas Papua and Sulawesi may be able to sustain the crop longer.
Furthermore, only Thailand had significant OP mortality by 2050 (Paterson 2020b) of the RAC discussed in the present paper, again indicating how the industry might be unsustainable soon in this country. Kalimantan and the Philippines were considered to have mortalities of ~10% by 2100, which is quite low. A total of 10% of the OP in Kalimantan would represent many OP, because there are many plantations in that region. PNG was assessed as having a large percentage OP mortality by 2100 of ~40% (Paterson 2020b), indicting problems with sustainability, which is consistent with the large reduction in CSC in the present paper. Thailand had an extremely high level of mortality of 70% by 2100, consistent with the severe reduction in CSC in the current paper. There were low OP mortalities by 2050 for Malaysia and Indonesia when considered as whole countries, which increased to a level of 10% by 2100 (Paterson 2020a), although 10% of the OP would represent a very large number given the high number of OP in these countries. Paterson (2020c) discussed the effect of future climate on OP mortality in Malaysia. High levels of OP mortality were provided by the model in Peninsular Malaysia by 2100, which was considered as being the least sustainable region in Malaysia for palm oil production. A similar treatment was undertaken for Indonesia (Paterson 2020d). Mortality of OP from unsuitable climate was high for Sumatra and Java and low for Sulawesi, Kalimantan and Papua.
The accuracy of the scenarios described in the current paper can be assessed with time. For example, it may be determinable by 2035 whether the trends are consistent with the derived schemes for 2050. However, procedures for (1) reducing climate change and (2) the effect of climate change on OP may increase average CSC. Climate change may be more severe than determined herein and predicted generally, and overall CSC may decrease even more than described in this report.
The development of OP cultivars resistant to climate change, or environmental stress, may overcome the less favourable growth conditions imposed by climate change (Rival 2017). This approach has yet to result in resistant palms, and a problem may be not knowing which climate stress(es) and levels will be important in the future. Furthermore, Paterson and Lima (2018) described some key areas of research and development which may reduce the effect on OP cultivation of climate change and the effect of climate change on OP agronomy. These areas include (1) interplanting leguminous crops to increase nitrogen in the soil, (2) using empty OP bunches to enhance soil, (3) adding earthworms to improve soil, (4) employing arbuscular mycorrhizal fungi, and (5) biochar to assist nutrition of OP. In addition, plantation management will be assisted by utilising the results presented in the present paper to indicate where OP will be most and least likely to survive. The most effective manner of addressing climate change is to adhere to the policies devised at the recent climate meeting in Spain (COP25), by reducing greenhouse gases and keeping temperature rise to more manageable levels (https://unfccc.int/cop25, accessed 2 April 2020). Conservation scientists, managers and environmental policy makers need to adapt their guidelines and policies accordingly (Brooke 2008) to mitigate the impact of climate change and contribute to future COP meetings.
In conclusion, there was a longitudinal trend for more favourable climate for growing OP from west to east. This applies to (1) current time to 2050, (2) current time to 2100, and (3) 2050 to 2100. Hence, the more easterly RAC in SE Asia may provide refuges for OP and offer scope for growing OP commercially into the future. SE Asia is more similar to Africa than to South America in terms of response to future suitable climate. The results indicate that the OP will not be totally decimated by future climate and the industry will have options to develop in countries with higher CSC or where the decreases are not as great and this appears to occur longitudinally. The environmental issues must be given priority. The general trend is for suitable climate to decrease severely in the future. The methods described herein offer the possibility of providing much more useful information from climate maps than is obtained by more superficial visual estimations. The results indicate how other plants in the tropics, about which there is little information, and given that latitudinal spread is unlikely, could adapt to climate change.
Declaration of funding
This research did not receive any funding.
Conflicts of interest
The author declares no conflict of interest.
Acknowledgements
The author is grateful for the IOI Professorial Chair at University Putra Malaysia in 2018–2019.
References
Arrieta, F., Teixeira, F., Yanez, E., Lora, E., and Castillo, E. (2007). Cogeneration potential in the Columbian palm oil industry: three case studies. Biomass and Bioenergy 31, 503–511.| Cogeneration potential in the Columbian palm oil industry: three case studies.Crossref | GoogleScholarGoogle Scholar |
Basiron, Y. (2007). Palm oil production through sustainable plantations. European Journal of Lipid Science and Technology 109, 289–295.
| Palm oil production through sustainable plantations.Crossref | GoogleScholarGoogle Scholar |
Bellard, C., Bertelsmeier, C., Leadley, P., Thuiller, W., and Courchamp, F. (2012). Impacts of climate change on the future of biodiversity. Ecology Letters 15, 365–377.
| Impacts of climate change on the future of biodiversity.Crossref | GoogleScholarGoogle Scholar | 22257223PubMed |
Blach-Overgaard, A., Balslev, H., Dransfield, J., Normand, S., and Svenning, J. (2015). Global-change vulnerability of a key plant resource, the African palms. Scientific Reports 5, 12611.
| Global-change vulnerability of a key plant resource, the African palms.Crossref | GoogleScholarGoogle Scholar | 26211732PubMed |
Brooke, C. (2008). Conservation and adaptation to climate change. Conservation Biology 22, 1471–1476.
| Conservation and adaptation to climate change.Crossref | GoogleScholarGoogle Scholar | 18759775PubMed |
Cho, S. J., and McCarl, B. A. (2017). Climate change influences on crop mix shifts in the United States. Scientific Reports 7, 40845.
| Climate change influences on crop mix shifts in the United States.Crossref | GoogleScholarGoogle Scholar | 28098198PubMed |
Corley, R. H. V., and Tinker, P. B. (2015). ‘The Oil Palm.’ (Wiley Blackwell: Chichester.)
Cosiaux, A., Gardiner, L. M., Stau, F. W., Bachman, S. P., Sonké, B., Baker, W. J., and Couvreur, T. L. P. (2018). Low extinction risk for an important plant resource: conservation assessments of continental African palms (Arecaceae/Palmae). Biological Conservation 221, 323–333.
| Low extinction risk for an important plant resource: conservation assessments of continental African palms (Arecaceae/Palmae).Crossref | GoogleScholarGoogle Scholar |
Dislich, C., Keyel, A. C., Salecker, J., Kisel, Y., Meyer, K. M., Auliya, M., et al. (2017). A review of the ecosystem functions in oil palm plantations, using forests as a reference system. Biological Reviews of the Cambridge Philosophical Society 49, 1539–1569.
Feeley, K. J., Stroud, J. T., and Perez, T. M. (2017). Most ‘global’ reviews of species’ responses to climate change are not truly global. Diversity and Distributions 23, 231–234.
| Most ‘global’ reviews of species’ responses to climate change are not truly global.Crossref | GoogleScholarGoogle Scholar |
Fei, S., Desprez, J. M., Potter, K. M., Jo, I., Knott, J. A., and Oswalt, C. M. (2017). Divergence of species responses to climate change. Science Advances 3, e1630355.
Fitzherbert, E. B., Struebig, M. J., Morel, A., Danielsen, F., Brühl, C. A., Donald, P. F., and Phalan, B. (2008). How will oil palm expansion affect biodiversity? Trends in Ecology & Evolution 23, 538–545.
| How will oil palm expansion affect biodiversity?Crossref | GoogleScholarGoogle Scholar |
Ghini, R., Bettiol, W., and Hamada, E. (2011). Diseases in tropical and plantation crops as affected by climate changes: current knowledge and perspectives. Plant Pathology 60, 122–132.
| Diseases in tropical and plantation crops as affected by climate changes: current knowledge and perspectives.Crossref | GoogleScholarGoogle Scholar |
Gibb, R., Redding, D. W., Chin, K. Q., Donnelly, C. A., Blackburn, T. M., Newbold, T., and Jones, K. E. (2020). Zoonotic host diversity increases in human-dominated ecosystems. Nature 584, 398–402.
| Zoonotic host diversity increases in human-dominated ecosystems.Crossref | GoogleScholarGoogle Scholar | 32759999PubMed |
Kadandale, S., Martin, R., and Smith, R. (2019). The palm oil industry and noncommunicable diseases. Bulletin of the World Health Organization 97, 118–128.
| 30728618PubMed |
Koh, L. P., and Wilcove, D. S. (2008). Is oil palm agriculture really destroying tropical biodiversity? Conservation Letters 1, 60–64.
Lenoir, J., and Svenning, J. (2015). Climate-related range shifts – a global multidimensional synthesis and new research directions. Ecography 38, 15–28.
| Climate-related range shifts – a global multidimensional synthesis and new research directions.Crossref | GoogleScholarGoogle Scholar |
Lim, L. W. K., Chung, H. H., and Hussain, H. (2020). Complete chloroplast genome sequencing of sago palm (Metroxylon sagu Rottb.): molecular structures, comparative analysis and evolutionary significance. Gene Reports 19, 100662.
| Complete chloroplast genome sequencing of sago palm (Metroxylon sagu Rottb.): molecular structures, comparative analysis and evolutionary significance.Crossref | GoogleScholarGoogle Scholar |
Lobell, D. B., Field, C. B., Cahill, K. N., and Bonfils, C. (2006). Impacts of future climate change on California perennial crop yields: model projections with climate and crop uncertainties. Agricultural and Forest Meteorology 141, 208–218.
| Impacts of future climate change on California perennial crop yields: model projections with climate and crop uncertainties.Crossref | GoogleScholarGoogle Scholar |
Marlier, M. E., Defries, R., Pennington, D., Nelson, E., Ordway, E. M., Lewis, J., Koplitz, S. N., and Mickley, L. J. (2014). Future fire emissions associated with projected land use change in Sumatra. Global Change Biology 21, 345–362.
| 25044917PubMed |
Miller, A. E., Wilson, T. L., Sherriff, R. L., and Walton, J. (2017). Warming drives a front of white spruce establishment near western treeline, Alaska. Global Change Biology 23, 5509–5522.
| Warming drives a front of white spruce establishment near western treeline, Alaska.Crossref | GoogleScholarGoogle Scholar | 28712139PubMed |
Ohimain, E. I., and Izah, S. C. (2014). Energy self-sufficiency of smallholder oil palm processing in Nigeria. Renewable Energy 63, 426–431.
| Energy self-sufficiency of smallholder oil palm processing in Nigeria.Crossref | GoogleScholarGoogle Scholar |
Ommelna, B. G., Jennifer, A. N., and Chong, K. (2012). The potential of chitosan in suppressing Ganoderma boninense infection in oil-palm seedlings. Journal of Sustainable Science and Management 7, 186–192.
Parmesan, C. (2006). Ecological and evolutionary responses to recent climate change. Annual Review of Ecology, Evolution, and Systematics 37, 637–669.
| Ecological and evolutionary responses to recent climate change.Crossref | GoogleScholarGoogle Scholar |
Paterson, R. R. M. (2019a). Ganoderma boninense disease of oil palm to significantly reduce production after 2050 in Sumatra if projected climate change occurs. Microorganisms 7, 4–6.
| Ganoderma boninense disease of oil palm to significantly reduce production after 2050 in Sumatra if projected climate change occurs.Crossref | GoogleScholarGoogle Scholar |
Paterson, R. R. M. (2019b). Ganoderma boninense disease deduced from simulation modelling with large data sets of future Malaysian oil palm climate. Phytoparasitica 47, 255–262.
| Ganoderma boninense disease deduced from simulation modelling with large data sets of future Malaysian oil palm climate.Crossref | GoogleScholarGoogle Scholar |
Paterson, R. R. M. (2020a). Future scenarios for oil palm mortality and infection by Phytophthora palmivora in Colombia, Ecuador and Brazil, extrapolated to Malaysia and Indonesia. Phytoparasitica 48, 513–523.
| Future scenarios for oil palm mortality and infection by Phytophthora palmivora in Colombia, Ecuador and Brazil, extrapolated to Malaysia and Indonesia.Crossref | GoogleScholarGoogle Scholar |
Paterson, R. R. M. (2020b). Oil palm survival under climate change in Kalimantan and alternative SE Asian palm oil countries with future basal stem rot assessments. Forest Pathology 50, e12604.
| Oil palm survival under climate change in Kalimantan and alternative SE Asian palm oil countries with future basal stem rot assessments.Crossref | GoogleScholarGoogle Scholar |
Paterson, R. R. M. (2020c). Oil palm survival under climate change in Malaysia with future basal stem rot assessments. Forest Pathology 50, e12641.
| Oil palm survival under climate change in Malaysia with future basal stem rot assessments.Crossref | GoogleScholarGoogle Scholar |
Paterson, R. R. M. (2020d). Depletion of Indonesian oil palm plantations implied from modeling oil palm mortality and Ganoderma boninense rot under future climate. AIMS Environmental Science 7, 366–379.
| Depletion of Indonesian oil palm plantations implied from modeling oil palm mortality and Ganoderma boninense rot under future climate.Crossref | GoogleScholarGoogle Scholar |
Paterson, R. R. M. (2021a). Longitudinal trends of future climate change and oil palm growth: empirical evidence for tropical Africa. Environmental Science and Pollution Research , .
| Longitudinal trends of future climate change and oil palm growth: empirical evidence for tropical Africa.Crossref | GoogleScholarGoogle Scholar |
Paterson, R. R. M. (2021b). Modelling longitudinal trends to oil palm refuges and oil palm mortalities in South America under future climate. SF Journal of Agriculture and Crop Management 2, 1011.
Paterson, R. R. M., and Lima, N. (2018). Climate change affecting oil palm agronomy, and oil palm cultivation increasing climate change, require amelioration. Ecology and Evolution 8, 452–461.
Paterson, R. R. M., Sariah, M., and Lima, N. (2013). How will climate change affect oil palm fungal diseases? Crop Protection 46, 113–120.
| How will climate change affect oil palm fungal diseases?Crossref | GoogleScholarGoogle Scholar |
Paterson, R. R. M., Kumar, L., Taylor, S., and Lima, N. (2015). Future climate effects on suitability for growth of oil palms in Malaysia and Indonesia. Scientific Reports 5, 14457.
| Future climate effects on suitability for growth of oil palms in Malaysia and Indonesia.Crossref | GoogleScholarGoogle Scholar |
Paterson, R. R. M., Kumar, L., Shabani, F., and Lima, N. (2017). World climate suitability projections to 2050 and 2100 for growing oil palm. Journal of Agricultural Science 155, 689–702.
| World climate suitability projections to 2050 and 2100 for growing oil palm.Crossref | GoogleScholarGoogle Scholar |
Perera, L., Baudouin, L., and Mackay, I. (2016). SSR markers indicate a common origin of self-pollinating dwarf coconut in South-East Asia under domestication. Scientia Horticulturae 211, 255–262.
| SSR markers indicate a common origin of self-pollinating dwarf coconut in South-East Asia under domestication.Crossref | GoogleScholarGoogle Scholar |
Ramirez-Cabral, N. Y. Z., Kumar, L., and Shabani, F. (2017). Global alterations in areas of suitability for maize production from climate change and using a mechanistic species distribution model (CLIMEX). Scientific Reports 7, 5910.
| Global alterations in areas of suitability for maize production from climate change and using a mechanistic species distribution model (CLIMEX).Crossref | GoogleScholarGoogle Scholar |
Rival, A. (2017). Breeding the oil palm (Elaeis guineensis Jacq.) for climate change. OCL 24, D107.
| Breeding the oil palm (Elaeis guineensis Jacq.) for climate change.Crossref | GoogleScholarGoogle Scholar |
Rosenzweig, C., Karoly, D., Vicarelli, M., Neofotis, P., Wu, Q., Casassa, G., et al. (2008). Attributing physical and biological impacts to anthropogenic climate change. Nature 453, 353–357.
| Attributing physical and biological impacts to anthropogenic climate change.Crossref | GoogleScholarGoogle Scholar | 18480817PubMed |
Sarkar, S. K., Begum, R. A., and Pereira, J. J. (2020). Impacts of climate change on oil palm production in Malaysia. Environmental Science and Pollution Research 27, 9760–9770.
Shabani, F., Kumar, L., and Esmaeili, A. (2014). Future distributions of Fusarium oxysporum f. spp. in European, Middle Eastern and North African agricultural regions under climate change. Agriculture, Ecosystems and Environment 197, 96–105.
| Future distributions of Fusarium oxysporum f. spp. in European, Middle Eastern and North African agricultural regions under climate change.Crossref | GoogleScholarGoogle Scholar |
Sinervo, B., Méndez-de-la-Cruz, F., Miles, D. B., Heulin, B., Bastiaans, E., Cruz, M. V. S., et al. (2010). Erosion of lizard diversity by climate change and altered thermal niches. Science 328, 894–899.
| Erosion of lizard diversity by climate change and altered thermal niches.Crossref | GoogleScholarGoogle Scholar | 20466932PubMed |
Sittaro, F., Paquette, A., Messier, C., and Nock, C. (2017). Tree range expansion in eastern North America fails to keep pace with climate warming at northern range limits. Global Change Biology 23, 3292–3301.
| Tree range expansion in eastern North America fails to keep pace with climate warming at northern range limits.Crossref | GoogleScholarGoogle Scholar | 28165187PubMed |
Thackeray, S. J., Sparks, T. H., Frederiksen, M., Burthe, S., Bacon, P. J., Bell, J. R., et al. (2010). Trophic level asynchrony in rates of phenological change for marine, freshwater and terrestrial environments. Global Change Biology 16, 3304–3313.
| Trophic level asynchrony in rates of phenological change for marine, freshwater and terrestrial environments.Crossref | GoogleScholarGoogle Scholar |
Urban, M. (2015). Accelerating extinction risk from climate change. Science 348, 571–573.
| 25931559PubMed |
Villela, A. A., Jaccoud, D. B., Rosa, L. P., and Freitas, M. V. (2014). Status and prospects of oil palm in the Brazilian Amazon. Biomass and Bioenergy 67, 270–278.
| Status and prospects of oil palm in the Brazilian Amazon.Crossref | GoogleScholarGoogle Scholar |
Wright, S. J., Muller-Landau, H. C., and Schipper, J. A. N. (2011). The future of tropical species on a warmer planet. Conservation Biology 23, 1418–1426.
| The future of tropical species on a warmer planet.Crossref | GoogleScholarGoogle Scholar |
Zune, M., Rodrigues, L., and Gillott, M. (2020). Vernacular passive design in Myanmar housing for thermal comfort. Sustainable Cities and Society 54, 101992.
| Vernacular passive design in Myanmar housing for thermal comfort.Crossref | GoogleScholarGoogle Scholar |