

Islands in the sky – could complex topography help us rewild beyond the fence?
Rob Brewster A * , Tom Jameson A B , Francesca Roncolato A , Mathew S. Crowther C , Patrick B. Finnerty C * and Thomas M. Newsome C
A * , Tom Jameson A B , Francesca Roncolato A , Mathew S. Crowther C , Patrick B. Finnerty C * and Thomas M. Newsome C
A
B
C
Handling Editor: Mike Calver
Abstract
The protection of threatened species in fenced safe havens has become a vital component of conservation management in Australia. However, despite their success, fenced safe havens face several ecological and economic constraints. There is a need to explore additional approaches to restore species beyond the fence.
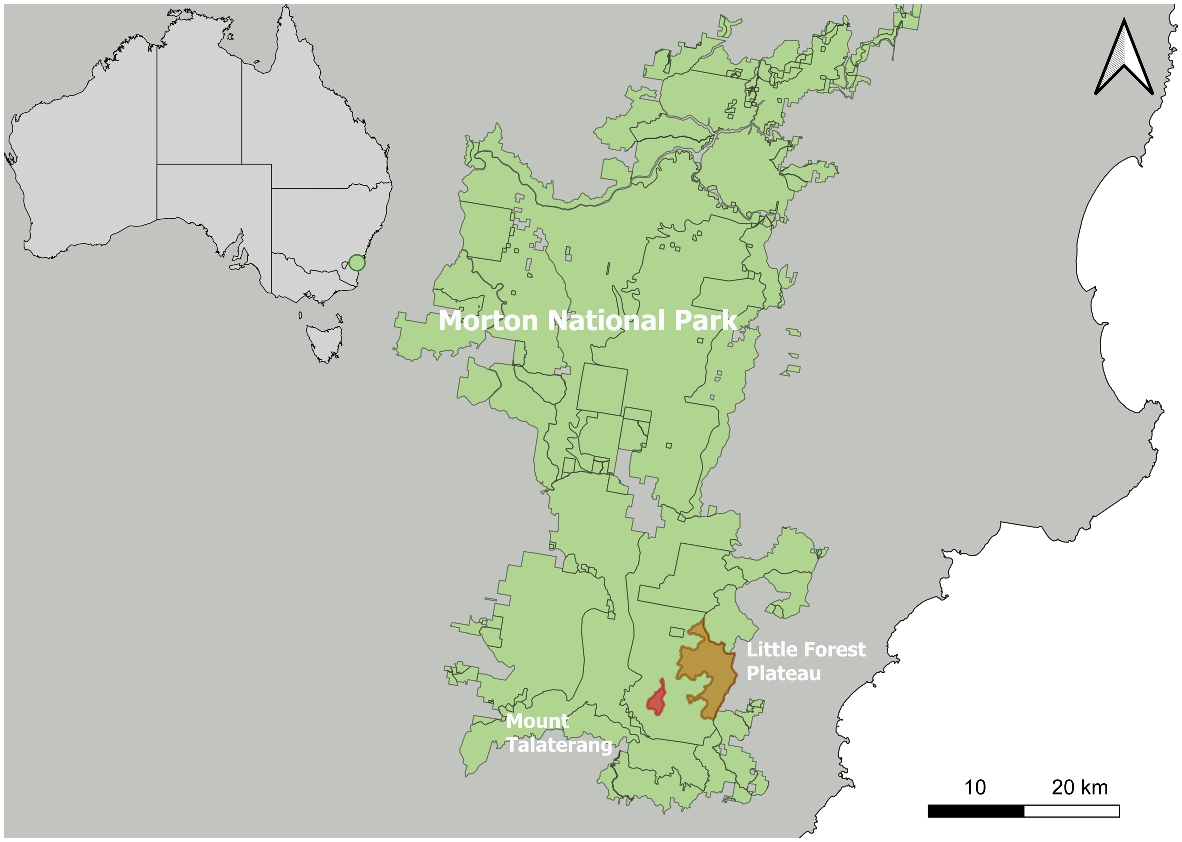
To explore naturally occurring mesas as potential ‘sky-island safe havens’, created by natural barriers in elevation and relief, which may restrict the movement of introduced predators and other mammals.
We examined species occurrences at a mesa site (Mt. Talaterang in south-east NSW, Australia) as well as a nearby lower-lying site (Little Forest Plateau). We then provide a geospatial analysis of other mesas in NSW to investigate the number of potential sky-island safe havens in the state.
Species assemblages differed between the two sites, with red foxes (Vulpes vulpes), dingoes/domestic dogs (Canis dingo/familiaris), and European rabbits (Oryctolagus cuniculus) absent from the mesa site, while Antechinus spp. were not detected from the lower-lying site. Feral cats (Felis catus) occurred at significantly lower densities on the mesa site compared to the lower-lying site. In NSW, we identified 91 other mesas of ≥10 ha with similar topology as Mt. Talaterang.
Although differences in species assemblages are expected between different habitats, the absence of red foxes and lower number of feral cat detections at the mesa site suggest the need to further explore the potential for mesas in conservation initiatives.
Our findings introduce a supplementary conservation strategy that could augment existing fenced safe haven approaches.
Keywords: conservation management, conservation translocation, invasive species, mesa, rewilding, reintroduction, safe haven, threatened species.
Introduction
A key strategy for protecting susceptible species from introduced predators is the creation of safe havens. Safe havens include islands and mainland fenced areas, where introduced predators are either naturally absent or excluded by management (Legge et al. 2018; Ringma et al. 2018). As a conservation tool, safe havens limit overdispersal of remnant or translocated populations. Overdispersal can be a contributing factor to population failure among translocated populations if fecundity and site fidelity is not sufficient to offset dispersal and mortality. For reintroduction programs, release protocols encourage individuals to remain near each other, thus ensuring that all individuals are available to contribute to genetic mixing, and to maximise access to supplementary feeding (Hardman and Moro 2006) and other support (e.g. enhanced threat management near release site).
In Australia, the establishment of fenced or island safe havens has emerged as a crucial element in conservation management (Legge et al. 2018). Through eradication programs, these havens in Australia are primarily used to protect native species from the detrimental effects of European red foxes (Vulpes vulpes) and feral cats (Felis catus), which have had devastating impacts in causing the decline and extinction of at least 25 mammal species (Radford et al. 2018), particularly critical weight range mammals (ground dwelling species 35 g–5.5 kg) (Johnson and Isaac 2009; Woinarski et al. 2015; Doherty et al. 2017).
Although invasive species eradication programs on islands can be logistically challenging, maintenance is generally less expensive and complex than in fenced safe havens, with island perimeters not requiring repair and boundaries not being constantly challenged by invasive species (as reviewed by Legge et al. 2018). Fenced safe havens can also produce new conservation challenges including creating barriers to the migration of non-target species (Dickman 2012). Despite these constraints, fenced safe havens persist as a vital tool in safeguarding native species populations from introduced predators in areas or ecosystems where islands are not an option. Looking forward, however, there is a need to explore additional approaches to restore species beyond islands and fenced safe havens to expand the impact of conservation initiatives.
‘Refuges’, non-fenced, and non-island safe havens have been researched for well over a decade in Australia, however almost exclusively in arid regions (Pavey et al. 2017; Nano et al. 2019). Of note, extensive investigation has documented the importance of ridges and peaks within the MacDonnell Ranges of central Australia as core refuge habitats for mammalian species such as the Central Rock-rat (Zyzomys pedunculatus), ensuring protection from feral cats and fire (McDonald et al. 2013, 2014, 2015, 2020; Nano et al. 2019).
Analogous to these elevation-based refuges, similar naturally occurring mesas could act as ‘sky-island safe havens’ created by natural barriers in elevation and relief. Being isolated flat-topped areas elevated from the surrounded landscape with steep sides, mesas may offer an important supplementary conservation tool that could augment existing fenced or island safe haven approaches. Globally, the conservation value of mesas have been recognised, primarily as acting as natural movement barriers between genetically isolated populations (Ramírez-Prieto et al. 2016; Lehmberg et al. 2018; Miranda et al. 2021). In limiting species movement, we argue that the topography of mesas may also prevent the rapid incursion of invasive predators and the overdispersal by in situ mammals, two key factors crucial in any successful species recovery effort in Australia. Moreover, the topographic isolation of mesas from the surrounding landscape could also be valuable in protecting metapopulations of fire-vulnerable species. This is critical as many native Australian species are negatively affected by wildfires and the associated amplification of predation rates by fires (Hradsky et al. 2017; Ward et al. 2020).
To explore the potential of mesas as safe havens, we compared mammalian species occurrence over a short duration between a potential mesa safe haven site (Mt. Talaterang) and a nearby lower-lying site (Little Forest Plateau) in the surrounding landscape. The main goal was to compare the occurrence of introduced predators between the two areas, as well as to compare the differences in broader species assemblages. We expected to record significantly lower detections of introduced predators on the mesa-top compared with the lower-lying site and similar or higher abundances of native species. We also carried out a geospatial analysis across the state of New South Wales (NSW), Australia, to identify other mesas with the same topology as Mt. Talaterang to investigate the number of potential sky-island safe havens across the state.
Materials and methods
Study site
Mt. Talaterang (−35.2774S, 150.2709E) is a 317-ha mesa (Fig. 1). The mesa has relatively few on-foot access points; with one on its most northerly aspect, at Pallin Pass, and another on the western edge above Dummal creek, both requiring a traverse over complex terrain (boulders, slots, and hauls) to reach the plateau. We also studied the area at Little Forest plateau (−35.2387S, 150.3228E) where a camera trap site was established approximately 3 km east of the closest camera trap site on Mt. Talaterang (Fig. 1). Little Forest plateau is approximately 2954 ha. This site is geographically lower-lying and has more extensive connectivity to the surrounding landscape than Mt. Talaterang, with well-maintained public vehicle access roads (Mt. Bushwalker Road/Tianjara Fire trail, Pointers Gap Road) as well as multiple foot-passes (Egans Gap, Martins Gap, Ingold Gap, Gadara Point, among other unnamed on-foot access points).
Camera trap deployment
In April 2022, we deployed 16 Swift Enduro camera traps on Mt. Talaterang, spaced at a minimum of 250 m apart, with camera traps placed along remnant walking tracks, animal trails, or in open landscape amongst large flat rocky outcrops that characterise much of the south-western portion of this plateau (Fig. 2). Mean camera elevation for cameras deployed on the plateau was 687 m (range 619.4–761.5 m, Supplementary Table S1). On Little Forest Plateau, we deployed 20 Swift Enduro camera traps spaced a minimum of 350 m apart along a transect that used existing roads and walking trails (Fig. 2). Camera trap deployment was limited to existing roads and foot trails as the site lies within the boundaries of the former Tianjara Artillery Training Area and off-track activities carry an Unexploded Ordinance risk. Mean camera elevation on Little Forest plateau was 529 m (range 491.8–584 m; Table S1).
Camera trap sites at (a) Little Forest plateau (lower-lying site) and (b) Mt. Talaterang plateau (mesa). (c) Experimental set-up of a motioned-triggered camera facing towards a tea strainer (circled in red) that contained a universal attractant mix (mixture of oats, peanut butter, and tuna oil) fixed to the ground using a tent peg. Tea strainers were placed between 1.5 m and 5 m in front of each camera.
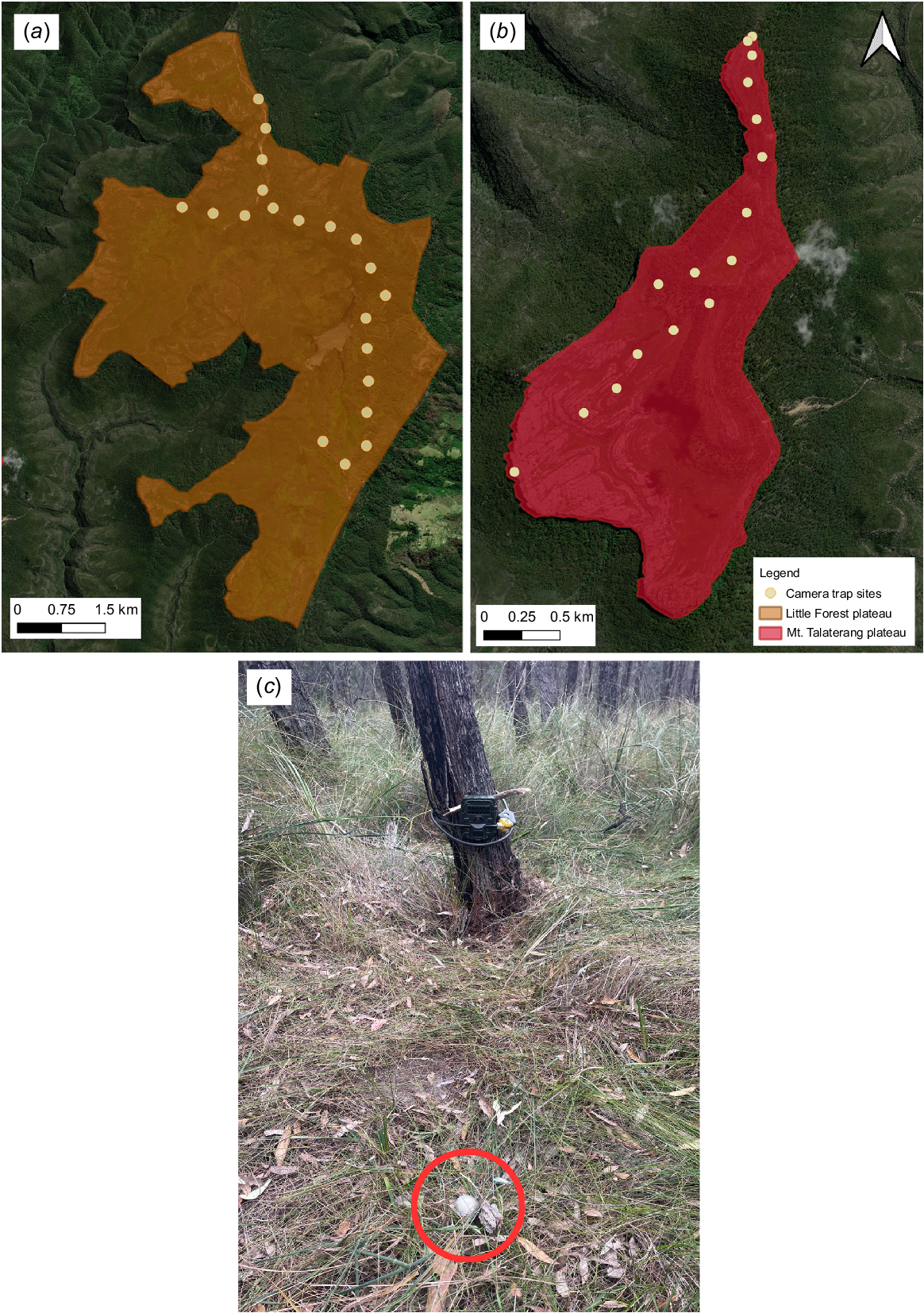
At both sites, we attached cameras to vegetation at heights of approximately 50 cm and aimed parallel to the ground. At all sites, we fixed a tea strainer containing a universal attractant mix (mixture of oats, peanut butter and tuna oil) to the ground using a tent-peg at between 1.5 m and 5 m in front of each camera. Attractants could be smelt, but not accessed by animals. Cameras were active for 24 h per day and set to take five photographs with no rest period using infra-red flash. Cameras were deployed at Mt. Talaterang for 79–118 days (giving a total survey effort of 1724 trap days), and at Little Forest Plateau for 119 days (giving a total survey effort of 2380 trap days).
We uploaded images from each camera to the Wildlife Insights image management online platform (Wildlife Insights 2022), which integrates machine learning models to assist with the identification of blank images and species. We also manually checked images and classified to the species level all mammals (except rodents, which were classified to the genus level). The catalogued data set was then downloaded from Wildlife Insights for analysis. Of the images analysed, a domestic dog (Canis familiaris) was identified as separate to a dingo (Canis dingo) by either the presence of a collar, a human walking the dog on a lead, or a human appearing in images captured immediately after those of the dog. Furthermore, based on previous genetic analysis of Cairns et al. (2021) demonstrating the rarity of feral dogs within Australia, any non-fox canid captured without the aforementioned domesticated identifiers was classed as a dingo in subsequent analysis.
Camera trap analyses
For the purposes of analyses, we collated all camera trap records for each camera trap site. We used RStudio Desktop (Posit Team 2023) to filter the dataset for 5-min intervals for each species’ records obtained from the same camera location. We used these filtered records for further analysis as independent events. We transformed all data using a 4th root transformation to avoid very common species dominating the analyses. We compared the relative abundance and diversity of species between sites using the Bray-Curtis similarity matrix, which does not use joint absences to compare sites. We visualised separation between sites using non-metric multidimensional scaling (nMDS). We compared Mt. Talaterang and Little Forest plateau sites using a Permutational Multivariate Analysis of Variance (PERMANOVA). We used a SIMPER analysis to determine which species contributed most to the separation between groups. All analyses were performed using PRIMER-E (Clarke and Gorley 2016) with PERMANOVA (Anderson et al. 2008) extension.
Identifying other mesas in NSW
We used topographic maps of NSW to identify other mesas across the state with similar topology to Mt. Talaterang, with the goal of identifying other sky-island safe havens. We carried out all spatial data modification and analysis in QGIS ver. 3.8 (QGIS.org 2023). We used a Topographic Positioning Index based landform classification (Weiss 2001; Jenness 2008) of a 3-s Digital Elevation Model of Australia (Gallant et al. 2009) clipped to NSW and reprojected to EPSG: 32356 – WGS 72/UTM Zone 56S. We used a small neighbour (400 m) and long neighbour (2000 m) combination that distinguished Mt. Talaterang from the surrounding landscape as a collection of points classified as mesa surrounded on all sides by points classified as high ridges. We kept all other parameters as default. We used this parameter combination as it allowed Mt. Talaterang to be distinguished from the surrounding topology and hence allowed us to identify other mesas with the same topological characteristics. We counted all mesas in NSW with the same landform classification as Mt. Talaterang and measured the ellipsoidal area in hectares of all points classified as a flat mesa top for each mesa. We recorded all mesas of 10 ha or greater as potential sites for further investigation as sky-island safe havens.
Results
Camera trap data
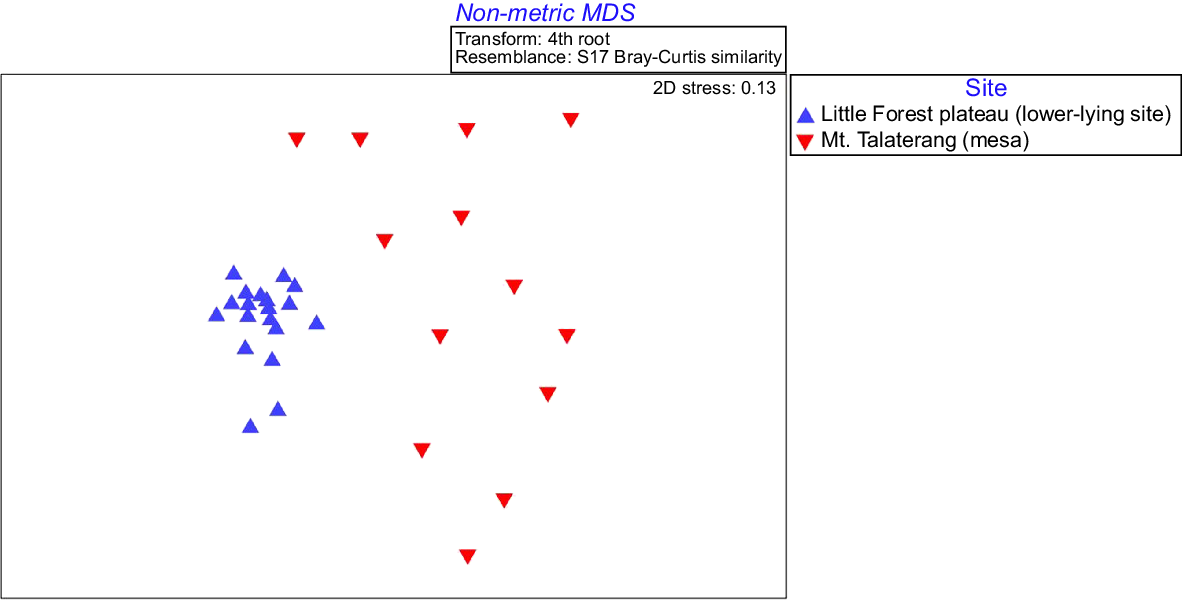
We found mammal abundances at Mt. Talaterang and Little Forest Plateau differed significantly (Pseudo-F = 14.621; d.f. = 1, 30; P < 0.001) and there was clear separation between the two sites in the nMDS (Fig. 3, Table 1). Importantly, red foxes were absent from Mt. Talaterang and feral cats were detected significantly less on the mesa-top than the surrounding landscape (Table 1). Two other invasive species identified at the lower-lying Little Forest Plateau site were absent on Mt. Talaterang: (1) the European rabbit (Oryctolagus cuniculus); and (2) the domestic dog, (Table 1).
nMDS results on mammal abundances at Mt. Talaterang (mesa) and Little Forest Plateau (lower-lying site).
| Species | Mt. Talaterang (mesa) | Little Forest plateau (lower-lying site) | Average dissimilarity | Dissimilarity/s.d. | Contrib.% | Cum.% | |
|---|---|---|---|---|---|---|---|
| Average abundance | Average abundance | ||||||
| European Red Fox (Vulpes vulpes) | 0 | 3 | 10.74 | 4.45 | 13.81 | 13.81 | |
| Bare-nosed Wombat (Vombat ursinus) | 0.68 | 3.07 | 8.96 | 2.19 | 11.52 | 25.33 | |
| Domestic Cat (Felis catus) | 0.55 | 2.53 | 6.94 | 2.32 | 8.92 | 34.24 | |
| Swamp Wallaby (Wallabia bicolor) | 1.31 | 2.01 | 6.71 | 1.45 | 8.62 | 42.86 | |
| Eastern Grey Kangaroo (Macropus giganteus) | 0 | 1.81 | 6.31 | 1.29 | 8.11 | 50.97 | |
| Long-nosed Bandicoot (Perameles nasuta) | 1.36 | 1.5 | 4.37 | 1.44 | 5.62 | 56.59 | |
| Rodent (Rattus spp.) | 1.32 | 0.29 | 4.28 | 1.08 | 5.5 | 62.1 | |
| Domestic Dog (Canis familiaris) | 0 | 1.11 | 4.19 | 1.19 | 5.38 | 67.48 | |
| Dingo (Canis dingo) | 0 | 1.04 | 3.6 | 1.28 | 4.63 | 72.11 | |
| Spotted-tailed Quoll (Dasyurus maculatus) | 0.68 | 0.05 | 2.38 | 0.83 | 3.06 | 79.19 | |
| European Rabbit (Orycotlagus cuniculus) | 0 | 0.5 | 1.77 | 0.65 | 2.28 | 81.47 | |
| Antechinus (Antechinus spp.) | 0.51 | 0 | 1.77 | 0.62 | 2.27 | 83.74 | |
| Ringtail Possum (Pseudocheirus peregrinus) | 0.38 | 0.09 | 1.5 | 0.68 | 1.93 | 87.71 |
The species that contributed most to the separation between sites were red foxes (percent contribution 13.83%), Bare-nosed Wombats (Vombatus ursinus) (percent contribution 11.5%), feral cats (percent contribution 8.92%), Swamp Wallabies (Wallabia bicolor) (percent contribution 8.65%), and Eastern Grey Kangaroos (Macropus giganteaus) (percent contribution 8.18%) (Table 1). As with red foxes, rabbits, and dogs, Eastern Grey Kangaroos and dingoes were absent from Mt. Talaterang. Antechinus (likely Antechinus stuartii) were not detected on Little Forest Plateau (Table 1).
Mesas across NSW
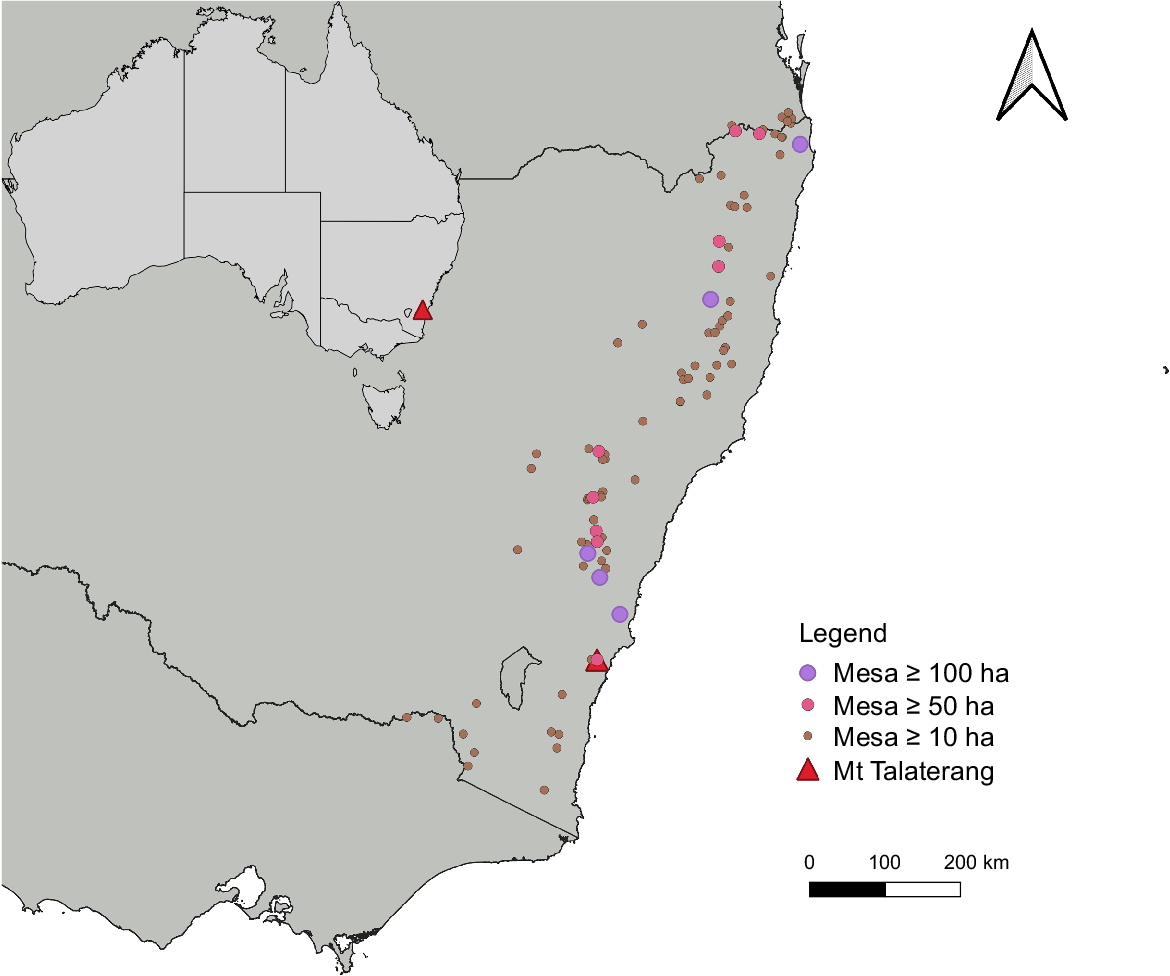
We identified 91 mesas of ≥10 ha or greater with similar landform identity as Mt. Talaterang (Table S2). Of the other 91 NSW mesas, 14 were 50 ha or larger, and 5 were 100 ha or larger, with the 2 largest, greater than 300 ha in area (Fig. 4).
Discussion
We found that Mt. Talaterang, a potential sky-island safe haven with geographic features that may limit the movement of some terrestrial mammals, was statistically different from the lower-lying site survey, in terms of the presence and detections of invasive and native mammals. Most importantly, significantly lower feral cat densities and no red foxes were detected on the mesa during the camera trap survey.
Although differences in species assemblages are expected between the different habitats surveyed, the absence of red foxes and lower number of feral cat detections at Mt. Talaterang suggest the need to further explore the potential for mesas to be used as safe havens in landscape-scale conservation initiatives. For example, Morton National Park, where Mt. Talaterang is located, is within the range of several locally threatened or regionally extinct species including the Southern Brown Bandicoot (Isoodon obesulus obesulus), Long-nosed Potoroo (Potorous tridactylus), Parma Wallaby (Notamacropus parma), Brush-tailed Rock Wallaby (Petrogale penicillata), and Eastern Quoll (Dasyurus viverrinus). If suitable habitat and foraging resources exists for these species, Mt. Talaterang may provide a candidate release site for targeted conservation translocations aimed at increasing and protecting locally threatened species, or reintroducing species that have become regionally extinct. However, it should be noted that these species may have never inhabited the site, despite once being found in the vicinity. This may be a function of access (i.e. the reason that perhaps foxes and other invasive species have also not been detected in this study), or due to sub-optimal habitat at the site, or surrounding the site, which provided a barrier to access. Alternatively, these species may have persisted at the site at one stage or multiple stages in the past, but stochastic events drove them off the plateau (i.e. catastrophic bushfire, drought, disease epidemic, or predation by another species). Further research is therefore warranted, to ascertain whether this, or other mesas, are suitable for the proposed species over the long-term.
It is also important to consider that differences in camera trap placement between the mesa and lower-lying site may have influenced overall predator detection rates. Predators, such as foxes and cats have a propensity to use formed roads to move through the landscape (Raiter et al. 2018; Hill et al. 2021). As such, the deployment of camera traps on fire trails at the lower-lying site, and the absence of fire trails on the mesa site where cameras were instead placed on hiking and animal trails, could have affected the detectability of these species.
More broadly, we identified 91 mesas of ≥10 ha with a similar landform identity as Mt. Talaterang in NSW alone, suggesting that other areas could be explored for similar efforts, and we discuss this further below.
Getting beyond the fence
There is a growing need to identify additional strategies that can be integrated with fenced safe haven initiatives to restore mammal populations in Australia. Land procurement for fenced sites in suitable habitats is a continual barrier to safe haven development, along with logistical challenges inherent with fence construction and maintenance costs. For example, a Population Viability Analysis for the Eastern Quoll indicated an additional occupancy area of 437 km2 is required to remove the species from the IUCN Red List within 10 years (Wilson et. al. 2022). This required land area is equivalent to 90 fenced reserves similar to the 485 ha Eastern Quoll safe haven, Mulligans Flat Woodland Sanctuary, developed in the ACT (not accounting for edge effects) (Wilson et. al. 2022). Due to the inherent difficulty in securing large parcels of land with suitable habitat for species recovery, geographical features such as mesas, functioning as sky-island refuges that may naturally restrict the incursion of red foxes and feral cats, could serve as viable supplementary translocation sites to aid conservation initiatives. These sites have the potential to bolster the extensive network of protected areas and aid in the management of species such as the Eastern Quoll.
We recognise that the geography of all mesas may not provide total protection from introduced predators. Variation in topography, such as fissures in rocks, waterfalls, and boulder fields, may provide the ability for many species, not only red foxes and feral cats, to move across mesa boundaries, albeit in a restricted manner. Indeed, even very low densities of feral cats can exert significant predation pressure on native species (Moseby et al. 2015, 2021). Similarly, even under intense management regimes, foxes have been known to extirpate reintroduced populations of critical weight range mammals including the Eastern Bettong (Bettongia gaimardi) (Evans et. al. 2021). Therefore, sky-island safe havens may not be suitable for species that are particularly vulnerable to cat and fox predation, or additional management may be required to address potential predator incursion points via the strategic placement of gates and fences. However, some level of ‘leakiness’ may be an advantage of mesas over entirely fenced safe havens that can disrupt long-term movement and migratory routes of non-target species.
As mesas may also be naturally semi-permeable to target taxa, sky-island safe havens could function under a ‘leaky fence model’ facilitating a halo or spill-over effect. In doing so, mesas may reduce the potential negative effects of overpopulation and genetic isolation associated with fenced or island safe havens (Agarwal and Bode 2019; Moseby et al. 2020a). Many mesas in NSW occur in relatively close clusters. For example, there are eight mesas of 10 ha or greater within a 20-km radius in Wollemi National Park. A ‘sky archipelago’ approach could therefore be developed to potentially form networks of protected core areas. However, to ensure the best chances of success individual mesas will need to be assessed on a case-by-case basis to evaluate for habitat suitability, access to appropriate foraging resources, and assess ‘leakiness’ to target species. Furthermore, the surrounding landscape will need to be managed to limit populations of introduced predators to low enough levels for persistence of target species beyond safe haven boundaries (Agarwal and Bode 2019; Moseby et al. 2020a).
The potential remoteness and limited accessibility of mesas may present management hurdles and lead to delays in emergency response times. For example, swift responses required to address red fox and feral cat incursion, or when a bushfire threatens the site may be delayed due to the topographic complexities limiting vehicle or even on-foot access. These limitations may be addressed in part by technological advances; including the deployment of remote ‘Felixer’ grooming traps that have been successfully used to selectively control invasive predators (Moseby et al. 2020b). However, the exposure to invasive predators in low numbers may potentially address another negative consequence of fenced safe havens – that of predator naivety. Exposing naïve mammals to real predation risk may provide individuals within target populations a degree of predator awareness (Moseby et al. 2015), which is key in increasing the likelihood or survival if interactions occur in future. This ‘Goldilocks Zone’, which is the ‘just right’ levels of predation needed to drive selection for predator-resistant native species, while ensuring population viability has been proposed recently (Evans et. al. 2021), and some of the sites identified within this study may provide suitable locations for developing this concept further.
Although sky-island safe havens may hold several advantages over fenced havens or islands, we recognise that mesa-tops may not be suitable for the release and survival of all species. Species that occur at lower elevations, are highly vulnerable to stochastic or extreme weather events, require extensive ranges, or require permanent water sources may not be candidates for release atop particular mesas. However, mesas may provide particularly valuable safe havens for montane species, including some critical weight range mammals within the Great Dividing Range of eastern Australia.
Conclusions and future directions
Further research is required to confirm and test the utility of mesas for rewilding species beyond the fence. We acknowledge that sky-island safe havens may be limited to species who are able to survive and reproduce on isolated mesa-tops. To be successful, ground truthing of mesa sites to assess habitat suitability and appropriate foraging resource availability for target taxa must be undertaken. Furthermore, assessing the infrastructure needs necessary for accessing and potentially reinforcing sites to optimise their suitability for safeguarding locally threatened or translocated species is a crucial step. Additionally, movement studies of both native and introduced mammals atop selected mesas and in the proximity around them would provide an important ‘leakiness’ score that may also inform site selection and future management decisions.
Optimum geographic barriers are those that totally exclude introduced predators and are only partially permeable to target native species, to limit over-dispersal but to provide the potential to facilitate some dispersal into the broader landscape, particularly when populations reach high densities. Sites selected for pilot translocations should therefore ensure that ongoing invasive species monitoring is undertaken, that translocation outcomes are monitored including population size and reproductive success of target species, and that genetic analysis is undertaken to monitor the impact of limiting movement of a target species and to inform future admixing required to maintain genetic health of the population. Thus, mesas will be most likely successful when used as part of an integrated strategy with other landscape-scale conservation initiatives.
Declaration of funding
This research was funded by the WWF-Australia Australian Wildlife and Nature Recovery Fund and (partially) by the Australian Government through the Australian Research Council LP210301265.
Acknowledgements
The authors acknowledge the Yuin people, the Traditional Owners of the land upon which the Morton National Park study sites are located. We thank NSW National Parks and Wildlife Service staff, Michael Phelan, Steine Lofts, Mike Jarman, Libby Shields, Nathan Cattel, Samantha Walker, Meghan Kempster, Ron Rutter, and Jackson Rutter, WWF-Australia staff Jemma Everingham, Scott Norford, Leah Hunt, Leonie Sii, Prishani Vengetas, Kita Ashman, Patrick Giumelli, Annie Yan, Caitlin Chalhoub, Jessica Sommers, and Kaz Hing, and volunteers Erika Schauberger, Stefan Briggs, Nathan Quigley, Jana Bartsch, and Leigh Sage for their assistance with logistics, fieldwork, and camera deployment. Our research was carried out under a NSW Department of Planning, Industry and Environment Scientific Licence (SL 102575) and NPWS Animal Ethics Committee Approval No. RVF21/2394.
References
Agarwal K, Bode M (2019) Modelling the effects of leaky predator-exclusion fences and their surrounding halo [Preprint]. bioRxiv
| Crossref | Google Scholar |
Cairns KM, Crowther MS, Nesbitt B, Letnic M (2021) The myth of wild dogs in Australia: are there any out there? Australian Mammalogy 44, 67-75.
| Crossref | Google Scholar |
Dickman CR (2012) Fences or ferals? Benefits and costs of conservation fencing in Australia. In ‘Fencing for conservation: restriction of evolutionary potential or a riposte to threatening processes?’. (Eds M Somers, M Hayward) pp. 43–63. (Springer: New York) doi:10.1007/978-1-4614-0902-1
Doherty TS, Dickman CR, Johnson CN, Legge SM, Ritchie EG, Woinarski JCZ (2017) Impacts and management of feral cats Felis catus in Australia. Mammal Review 47, 83-97.
| Crossref | Google Scholar |
Evans MJ, Batson WG, Gordon IJ, Belton E, Chaseling T, Fletcher D, Harrison M, McElroy T, Mungoven A, Newport J, Pierson J, Portas T, Swain S, Wimpenny C, Manning AD (2021) The ‘Goldilocks Zone’ of predation: the level of fox control needed to select predator resistance in a reintroduced mammal in Australia. Biodiversity and Conservation 30, 1731-1752.
| Crossref | Google Scholar |
Gallant J, Wilson N, Tickle PK, Dowling T, Read A (2009) 3 second SRTM derived Digital Elevation Model (DEM). Available at https://dev.ecat.ga.gov.au/geonetwork/srv/api/records/a05f7892-ef04-7506-e044-00144fdd4fa6 [Accessed 1 September 2023]
Hardman B, Moro D (2006) Optimising reintroduction success by delayed dispersal: Is the release protocol important for hare-wallabies? Biological Conservation 128, 403-411.
| Crossref | Google Scholar |
Hill JE, DeVault TL, Belant JL (2021) A review of ecological factors promoting road use by mammals. Mammal Review 51(2), 214-227.
| Crossref | Google Scholar |
Hradsky BA, Mildwaters C, Ritchie EG, Christie F, Di Stefano J (2017) Responses of invasive predators and native prey to a prescribed forest fire. Journal of Mammalogy 98, 835-847.
| Crossref | Google Scholar |
Jenness J (2008) Topographic Position Index (TPI) an ArcView 3.X tool for analyzing the shape of the landscape. Available at https://www.jennessent.com/arcview/tpi.htm [Accessed 1 September 2023]
Johnson CN, Isaac JL (2009) Body mass and extinction risk in Australian marsupials: the ‘Critical Weight Range’ revisited. Austral Ecology 34, 35-40.
| Crossref | Google Scholar |
Legge S, Woinarski JCZ, Burbidge AA, Palmer R, Ringma J, Radford JQ, Mitchell N, Bode M, Wintle B, Baseler M, Bentley J, Copley P, Dexter N, Dickman CR, Gillespie GR, Hill B, Johnson CN, Latch P, Letnic M, Manning A, McCreless EE, Menkhorst P, Morris K, Moseby K, Page M, Pannell D, Tuft K (2018) Havens for threatened Australian mammals: The contributions of fenced areas and offshore islands to the protection of mammal species susceptible to introduced predators. Wildlife Research 45, 627-644.
| Crossref | Google Scholar |
Lehmberg ES, Elbassiouny AA, Bloom DD, López-Fernández H, Crampton WGR, Lovejoy NR (2018) Fish biogeography in the “Lost World” of the Guiana Shield: phylogeography of the weakly electric knifefish Gymnotus carapo (Teleostei: Gymnotidae). Journal of Biogeography 45, 815-825.
| Crossref | Google Scholar |
McDonald PJ, Pavey CR, Knights K, Grantham D, Ward SJ, Nano CEM (2013) Extant population of the critically endangered central rock-rat Zyzomys pedunculatus located in the Northern Territory, Australia. Oryx 47, 303-306.
| Crossref | Google Scholar |
McDonald PJ, Brittingham R, Nano C, Paltridge R (2014) A new population of the critically endangered central rock-rat (Zyzomys pedunculatus) discovered in the Northern Territory. Australian Mammalogy 37, 97-100.
| Crossref | Google Scholar |
McDonald PJ, Griffiths AD, Nano CEM, Dickman CR, Ward SJ, Luck GW (2015) Landscape-scale factors determine occupancy of the critically endangered central rock-rat in arid Australia: the utility of camera trapping. Biological Conservation 191, 93-100.
| Crossref | Google Scholar |
McDonald PJ, Stewart A, Jensen MA, McGregor HW (2020) Topographic complexity potentially mediates cat predation risk for a critically endangered rodent. Wildlife Research 47(8), 643-648.
| Crossref | Google Scholar |
Miranda CV, Schwartsburd PB, Labiak PH, Prado J (2021) Three new species of Oleandra (Oleandraceae, Polypodiopsida) from the Neotropics, and notes on the morphological groups among the Neotropical species. Brittonia 73, 143-151.
| Crossref | Google Scholar |
Moseby KE, Peacock DE, Read JL (2015) Catastrophic cat predation: A call for predator profiling in wildlife protection programs. Biological Conservation 191, 331-340.
| Crossref | Google Scholar |
Moseby KE, McGregor H, Hill BM, Read JL (2020a) Exploring the internal and external wildlife gradients created by conservation fences. Conservation Biology 34, 220-231.
| Crossref | Google Scholar | PubMed |
Moseby KE, McGregor H, Read JL (2020b) Effectiveness of the Felixer grooming trap for the control of feral cats: a field trial in arid South Australia. Wildlife Research 47, 599-609.
| Crossref | Google Scholar |
Moseby KE, McGregor H, Read JL (2021) The lethal 23%: predator demography influences predation risk for threatened prey. Animal Conservation 24, 217-229.
| Crossref | Google Scholar |
Nano CEM, Randall DJ, Stewart AJ, Pavey CR, McDonald PJ (2019) Spatio-temporal gradients in food supply help explain the short-term colonisation dynamics of the critically endangered central rock-rat (Zyzomys pedunculatus). Austral Ecology 44(5), 838-849.
| Crossref | Google Scholar |
Pavey CR, Addison J, Brandle R, Dickman CR, McDonald PJ, Moseby KE, Young LI (2017) The role of refuges in the persistence of Australian dryland mammals. Biological Reviews 92, 647-664.
| Crossref | Google Scholar |
Posit Team (2023) RStudio: integrated development environment for R. Available at http://www.posit.co/ [Accessed 1 March 2023]
QGIS.org (2023) QGIS geographic information system. Available at QGIS.org [Accessed 1 June 2023]
Radford JQ, Woinarski JCZ, Legge S, Baseler M, Bentley J, Burbidge AA, Bode M, Copley P, Dexter N, Dickman CR, Gillespie G, Hill B, Johnson CN, Kanowski J, Latch P, Letnic M, Manning A, Menkhorst P, Mitchell N, Morris K, Moseby K, Page M, Ringma J (2018) Degrees of population-level susceptibility of Australian terrestrial non-volant mammal species to predation by the introduced red fox (Vulpes vulpes) and feral cat (Felis catus). Wildlife Research 45, 645-657.
| Crossref | Google Scholar |
Raiter KG, Hobbs RJ, Possingham HP, Valentine LE, Prober SM (2018) Vehicle tracks are predator highways in intact landscapes. Biological Conservation 228, 281-290.
| Crossref | Google Scholar |
Ramírez-Prieto J, Koch-Olt S, de Jesús Balleza-Cadengo J, Adame-González M, Romero-Nápoles J (2016) Flora on summit of the Mesa Alta, Jerez, Zacatecas, México. Botanical Sciences 94, 357-375.
| Crossref | Google Scholar |
Ringma J, Legge S, Woinarski J, Radford J, Wintle B, Bode M (2018) Australia’s mammal fauna requires a strategic and enhanced network of predator-free havens. Nature Ecology and Evolution 2, 410-411.
| Crossref | Google Scholar | PubMed |
Ward M, Tulloch AIT, Radford JQ, Williams BA, Reside AE, Macdonald SL, Mayfield HJ, Maron M, Possingham HP, Vine SJ, O’Connor JL, Massingham EJ, Greenville AC, Woinarski JCZ, Garnett ST, Lintermans M, Scheele BC, Carwardine J, Nimmo DG, Lindenmayer DB, Kooyman RM, Simmonds JS, Sonter LJ, Watson JEM (2020) Impact of 2019–2020 mega-fires on Australian fauna habitat. Nature Ecology & Evolution 4, 1321-1326.
| Crossref | Google Scholar | PubMed |
Weiss AD (2001) Topographic position and landforms analysis. Available at https://www.jennessent.com/arcview/tpi.htm [Accessed 1 September 2023]
Wildlife Insights (2022) WildlifeInsights.org. Available at https://www.wildlifeinsights.org/ [Accessed 1 April 2022]
Wilson BA, Evans MJ, Gordon IJ, Banks SC, Batson WG, Wimpenny C, Newport J, Manning AD (2022) Personality and plasticity predict postrelease performance in a reintroduced mesopredator. Animal Behaviour 187, 177-189.
| Crossref | Google Scholar |
Woinarski JCZ, Burbidge AA, Harrison PL (2015) Ongoing unraveling of a continental fauna: Decline and extinction of Australian mammals since European settlement. Proceedings of the National Academy of Sciences of the United States of America 112, 4531-4540.
| Crossref | Google Scholar |