

Ecohydraulic model for designing environmental flows supports recovery of imperilled Murray cod (Maccullochella peelii) in the Lower Darling–Baaka River following catastrophic fish kills
Ivor G. Stuart A D and Clayton P. Sharpe B C
A D and Clayton P. Sharpe B C
A Kingfisher Research, 177 Progress Road, Eltham, Vic. 3095, Australia.
B CPS Enviro, 972 Irymple Avenue, Irymple, Vic. 3498, Australia.
C Present address: New South Wales National Parks & Wildlife Service, PO Box 363, Buronga, NSW 2730, Australia.
D Corresponding author. Present address: Arthur Rylah Institute for Environmental Research, 123 Brown Street, Heidelberg, Vic. 3084, Australia. Email: ivor.stuart@delwp.vic.gov.au
Marine and Freshwater Research 73(2) 247-258 https://doi.org/10.1071/MF20377
Submitted: 23 December 2020 Accepted: 20 August 2021 Published: 20 September 2021
Journal Compilation © CSIRO 2022 Open Access CC BY-NC-ND
Abstract
Large dryland and semi-arid rivers are among the world’s most heavily modified ecosystems, and the Darling–Baaka River of eastern Australia highlights the challenges in conserving such ecosystems. Since 2000, the hydrology at the downstream end of the system (the Lower Darling River, LDR) has been transformed from a naturally near-perennial flowing system to an intermittent one by increased water abstraction, prolonged drought and climate change. This hydrological change has placed immense pressure on the native fish populations, such as the imperilled Murray cod (Maccullochella peelii), as evidenced by the 2018–19 catastrophic fish kills. Here we outline an ecohydraulic conceptual model for designing environmental flows to support spawning and recruitment of Murray cod. An environmental flow based on this model was released in 2016–17, following 524 consecutive days of continuous zero flows. The LDR flow consisted of an increased discharge in late winter–spring to promote broad-scale lotic (i.e. >0.3 m s–1) conditions, hydraulic complexity and continuous base flows to maintain connectivity and water quality. Monitoring of Murray cod during and following the flow revealed successful spawning and recruitment. This finding is significant because it provides justification for altering current water management policies that are failing to protect this nationally significant ecosystem.
Keywords: Australia, Murray–Darling Basin, recovery, recruitment, river regulation.
Introduction
Large dryland rivers are among the most heavily modified ecosystems on Earth due to resource exploitation, including flow regulation, water extraction, hydropower development and floodplain reclamation (Tockner and Stanford 2002; Vörösmarty et al. 2010). In rivers with highly variable natural flow regimes, including tropical, semi-arid and arid rivers, there is ongoing construction of new dams or the renewal of existing infrastructure to increase the reliability of water supply for human use (Nilsson et al. 2005; Grill et al. 2019). The large dryland rivers of the south-western US and Australia’s Murray–Darling Basin highlight this point, where at least 50% of flow is removed annually, mainly for agriculture (Walker et al. 1995; Gido et al. 2013; Richter et al. 2020). The natural hydrology in these rivers has become so altered due to overallocation of water that whole hydrological and hydraulic components have been lost, including perennial or near-perennial lotic base flows, hydraulic complexity, seasonal flow pulses and regular floodplain connections (Acreman et al. 2014; Mallen-Cooper and Zampatti 2020).
Alteration of flow regimes and floodplain inundation patterns can increase the frequency and intensity of fish kills, such as observed in Australia (Whitworth et al. 2012; Ellis et al. 2021), the US (Sargent and Galat 2002) and Europe (Petriki et al. 2021). After extreme kills, fish communities can be substantially altered and natural recovery can sometimes only occur after years of population augmentation by immigration or artificial stocking (Lyon and O’Connor 2008; Thiem et al. 2017).
In heavily altered river systems, one way of supporting aquatic ecosystems and preventing fish kills is through the manipulation of flow regimes using water storages or regulating structures (Richter and Thomas 2007). These managed ‘environmental flows’ aim to reinstate components of the natural flow regime that may be otherwise missing in altered rivers (Acreman et al. 2014; Moyle 2014; Webb et al. 2017). To achieve the best ecological response from environmental flow management, it is essential to understand ecohydraulic relationships so that specific flow regimes can be designed and long-term water plans developed to support fish recovery (Watts et al. 2018; New South Wales Department of Planning, Industry and Environment 2020; Yarnell et al. 2020).
The Darling River, in the Murray–Darling Basin, is 2850 km long and one of Australia’s largest semi-arid rivers. It is a good example of the environmental challenge facing many of the world’s semi-arid rivers. Nearly all (95%) run-off occurs in the headwater tributaries, where there is also gross overabstraction of flows, converting the near-perennial hydrology into a largely intermittent system, with intensification of zero-flow days, loss of continuous base flows and a much-reduced frequency of small and medium floods (Thoms and Sheldon 2000; Leblanc et al. 2012; Mallen-Cooper and Zampatti 2020). These hydrological impacts are most evident in reaches downstream of the major abstraction areas, where perennial lotic habitats are becoming increasingly rare. The species most susceptible to these hydrological changes are the lotic water specialists, including native silver perch (Bidyanus bidyanus), golden perch (Macquaria ambigua), bony herring (Nematalosa erebi) and Murray cod (Maccullochella peelii); Sharpe 2011; Tonkin et al. 2019), as well as river mussels (Alathyria jacksoni) and endemic Darling River snails (Notopala sublineata). Clear evidence of these impacts was seen in the Lower Darling River (LDR) in 2018–19 when river regulation, overabstraction, drought and extreme summer temperatures all contributed to multiple major fish deaths (Australian Academy of Science 2019; Vertessy et al. 2019). Murray cod were one species most affected by these fish kills and are classified as ‘vulnerable’ under Australia’s Environmental Protection of Biodiversity and Conservation Act (1999).
In this paper we describe the development of an ecohydraulic conceptual model that can be used to design environmental flows to support spawning and recruitment of Murray cod in the LDR. The model was developed using a hydrological dataset spanning the period 1974 and 2020 and used to design the first ever LDR environmental flow, which was delivered in 2016–17. Here we report the results of fish sampling during and following the environmental flow and draw conclusions regarding its effectiveness in supporting Murray cod spawning and recruitment. Following this study, extended cease-to-flow conditions returned to the LDR, culminating in the catastrophic fish kills of the summer 2018–19 (Australian Academy of Science 2019). We discuss how our findings helped inform a ‘river restart’ following the fish kills and how they can inform future planning to restore perennial flows in the LDR to protect fish communities.
Materials and methods
Site description
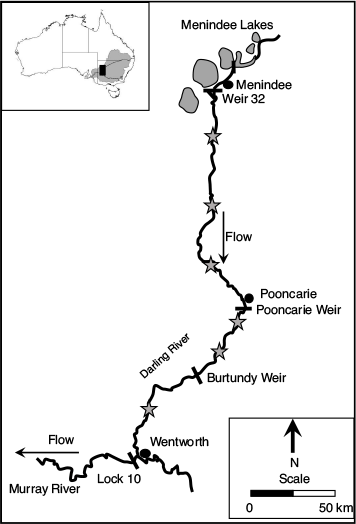
The LDR drains a semi-arid region of NSW and constitutes the 510 km downstream reach of the Darling River between the Menindee Lakes and the Murray River at Wentworth (Fig. 1). Baaka is the Aboriginal name for the LDR that has been used by the local Barkandji people for tens of thousands of years. There are no significant tributaries within this reach, with river flows originating from the upper catchment upstream of the town of Bourke (Thoms and Sheldon 2000). Small and medium flows upstream of Bourke can be entirely captured, and there is also major floodplain water harvesting with levees and block banks (Kingsford 2000).
|
Flows to the LDR are entirely regulated by the Menindee Lakes, which were converted into a floodplain storage scheme in the early 1960s (Thoms and Sheldon 2000) and now control flows into the LDR from the >1800 km upstream reach. The natural seasonality of flows to the LDR has not changed markedly, but the discharge magnitude has reduced, with fewer small and medium floods (10 000–50 000 ML day–1) and a greater frequency and duration of low flows (<500 ML day–1; Thoms and Sheldon 2000). The LDR occupies an incised low-gradient channel with abundant instream habitat and a terraced flood plain fringed by open desert plains and semi-arid woodlands (Sharpe 2011).
There are three low-level (<3 m high) weirs on the LDR that re-regulate flows for stock and domestic purposes, with each contributing to a cumulative total of ~100 km of lentic habitat. Naturally, the LDR was a near-perennial lotic habitat (Mallen-Cooper and Zampatti 2020), but since 2000 there has been gross overextraction in the upper catchment and an increasing frequency of drying and shorter water retention times in the Menindee Lakes (Kingsford 2000). Consequently, there has been a 56% decrease in average annual discharge and an increasing frequency of zero-flow sequences in the LDR (Mallen-Cooper and Zampatti 2020; New South Wales Department of Planning, Industry and Environment 2020; Stuart and Sharpe 2020).
Murray cod: ecohydraulic conceptual model
From a global perspective, freshwater megafauna (i.e. body mass >30 kg) are particularly prone to extinction (He et al. 2019). Murray cod underscore this point, being a nationally vulnerable freshwater fish species, growing to 1.5 m long and 50 kg, with an estimated maximum life span of 48 years (Anderson et al. 1992; Koehn et al. 2020). In the late 20th century, Murray cod populations declined dramatically due to overfishing, habitat destruction and river regulation contributing to broad-scale loss of perennial lotic conditions, hydraulic complexity and continuous base flows (Rowland 2004, 2020). There are conservation efforts throughout the range of Murray cod, mostly targeting physical habitat restoration and the stocking of hatchery-reared fingerlings (Thiem et al. 2017). The lower 500-km reach of the Darling River is a population stronghold for Murray cod and other native species, supported by lotic conditions and historic base flows (Stuart and Sharpe 2020).
Murray cod preferentially select nest sites in lotic habitats where they spawn annually in spring and early summer (October–December), when water temperature exceeds ~18°C (Koehn and Harrington 2005, 2006). Eggs are laid on hard surfaces, such as clay ledges, undercut banks, rocks and hollow logs (Rowland 2020), and males guard the nest for up to 18 days (Rowland 1998). Evidence from closely related species (the eastern freshwater cod Maccullochella ikei) suggests that spawning occurs in shallow (0.3–1.0 m) water (Butler and Rowland 2009). Hatching occurs 4–13 days after fertilisation depending on water temperature (Rowland 1988), with larvae commonly drifting downstream from the nest but also possibly dispersing locally (Koehn and Harrington 2006; King et al. 2009; Stuart et al. 2019).
Young-of-the-year (YOY) Murray cod occupy large and small woody debris, often in shallow near-riverbank habitats (Koehn 2009; Lieschke et al. 2016). Juvenile recruitment (i.e. survival to 1-year of age) appears to only occur in habitats characterised by: (1) hydraulic complexity (faster- and slow-flowing water), which is largely perennial or with short periods of zero flow outside the spawning season; (2) an increase in discharge in spring, with no rapid or major declines in discharge during the core nesting season so as to maintain the viability of eggs in nests in shallow areas; (3) a continuous summer base flow for juveniles to access productive littoral habitats for rapid growth and enhanced survival (Tonkin et al. 2017; Stuart et al. 2019); (4) base flows in winter to maintain lotic habitats, hydraulic complexity, connectivity and water quality, and thus reduce density-dependent pressure to improve survival of juvenile fish; (5) abundant large structural habitat; (6) a natural temperature regime; and (7) good water quality with few anoxic blackwater events (Koehn 2004; Stuart et al. 2019; Koehn et al. 2020; Tonkin et al. 2021).
In lowland rivers, flooding can enhance productivity, adult condition and the survival and growth of early life stages for Murray cod (Rowland 2004; Tonkin et al. 2017; Stoffels et al. 2020; Lyon et al. 2021). Despite evidence of spawning, there is very little recruitment to Murray cod populations in lentic riverine habitats or impoundments (Rowland 2004; Forbes et al. 2016), which appears related to both a low proportion of eggs hatching and insufficient suitable zooplankton to support larval feeding and survival (Matt McLellan, NSW DPI Fisheries, pers. comm.).
Antecedent hydrology
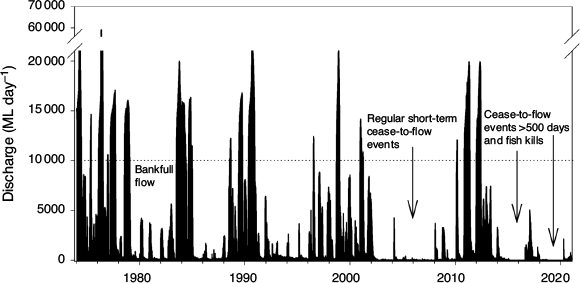
Although some aspects of this ecohydraulic conceptual model are generic, there are elements that are context specific in rivers due to climate, hydrological and hydraulic characteristics (Tonkin et al. 2017). The key to planning the LDR environmental flow was to examine the local hydrological data, which, before our study, were characterised by remarkable contrasts. Specifically, between 1974 and 2000 there were no zero- or near-zero (i.e. <15 ML day–1)-flow days at Burtundy gauge (No. 425007). However, between January 2000 and June 2020 there were six separate zero-flow events and a total of 1733 zero- or near-zero-flow days, including two separate events of 524 (March 2015–August 2016) and 555 (October 2018–April 2020) consecutive zero- or near-zero-flow days (Fig. 2). Major fish deaths occurred in 2004, 2018–19 and 2019–20, during these long, continuous, zero or near-zero flows, coupled with extreme hot weather and intense blue–green algal blooms, or at the recommencement of flows, when oxygen-stratified refuge pools became mixed and hypoxic (Ellis and Meredith 2004; Vertessy et al. 2019).
|
Environmental flow
By applying the ecohydraulic conceptual model, we can conclude that since 2000 the prevailing hydrology of the LDR was unsuitable for Murray cod. Specifically, there were: (1) periodic severe fish death events due to poor water quality caused by long periods of zero flow and extremely hot temperatures (Ellis and Meredith 2004; Vertessy et al. 2019); (2) variable regulated flows each spring with major discharge reductions interrupting Murray cod nesting during the spring spawning period; (3) losses of preferred perennial lotic conditions for nesting due to regular and long-duration continuous zero-flow events (Mallen-Cooper and Zampatti 2020); and (4) losses of end-of-system connection flows and fish movement opportunities to the perennial Murray River.
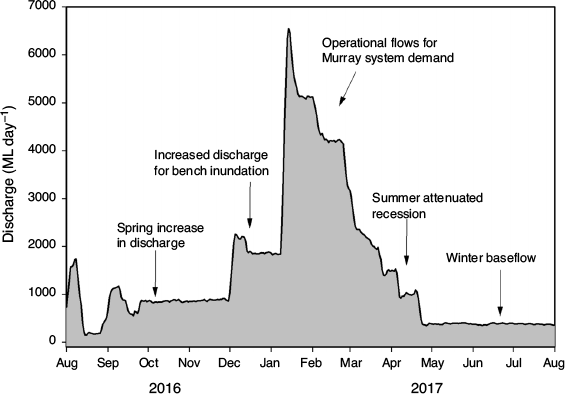
A potentially effective way to address these issues was through delivery of an environmental flow regime that applied the ecohydraulic principals, with four key functional components (Table 1). The environmental flow was delivered in spring–summer 2016–17 and adaptively managed and optimised in real time as monitoring data were collected (Fig. 2, 3) and included:
-
a late winter–spring increase in discharge to promote broad-scale lotic (i.e. >0.3 m s–1) conditions and to inundate low-lying benches and woody habitats to maximise nesting habitat availability, hydrodynamic complexity, depth, productivity and food availability to enhance larval survival (Tonkin et al. 2017)
-
a spring and early summer increase in discharge to inundate low-lying benches and woody habitats to maximise habitat complexity, hydrodynamic complexity, depth, productivity, water quality and feeding conditions to enhance larval survival, with no major water level drops (i.e. <0.30 m total reduction) during the core spring and early summer Murray cod nesting season to enable fish to complete their nesting and for larval retention (Stuart et al. 2019)
-
an annual summer continuous base flow to enable juvenile fish to access productive littoral habitats for rapid growth and enhanced survival (Tonkin et al. 2017; Stuart et al. 2019; Stoffels et al. 2020) and to maintain water quality conditions
-
an annual overwinter continuous base flow to maintain lotic habitats, hydraulic complexity, connectivity and water quality to reduce density-dependent pressure to improve survival of juvenile fish and promote adult conditioning (Tonkin et al. 2017; Stuart et al. 2019).

|
|
Sampling for larval Murray cod
Throughout the delivery of the environmental flow, sampling of larval Murray cod was undertaken six times in the LDR. At fortnightly intervals, Murray cod larvae were sampled during the breeding and post-spawning period in Austral spring and early summer (September 2016–January 2017). Six larval sampling sites, approximately 80 km apart, were selected along the 510-km length of the LDR, from ~35 km upstream of the Darling–Murray river confluence to ~50 km below Menindee Weir (Table 2). Five sites were in lotic habitats, with one in a slow-flowing weir pool environment in the LDR, some 5 km above the weir pool influence of Lock 10 on the Murray River.

|
At each site, three drift nets (500-µm mesh, opening diameter 0.5 m, length 1.5 m) and four light traps (clear Perspex with a yellow light stick) were set immediately before dusk and retrieved early the following morning (i.e. ~14-h soak time). The drift nets were suspended from fallen trees, in the central river channel, to filter the top 0.5 m of the water column. Each light trap had 3-mm mesh across the opening to exclude small predatory fish and was set in slow-flowing littoral habitats (e.g. <0.5 m deep, partially inundated sedges, saplings, macrophytes and fallen branches) where fish larvae may accumulate (Vilizzi et al. 2008). To provide an estimate of the volume of water filtered by each drift net, a measurement of water velocity was taken with a water velocity meter (FlowMate, Hong Kong, China) in the mouth of each net upon deployment and retrieval. All larval samples were preserved in >70% ethanol and returned to the laboratory for identification. Larval catch from the sites and different time periods is presented as a raw number.
Electrofishing and netting for YOY Murray cod
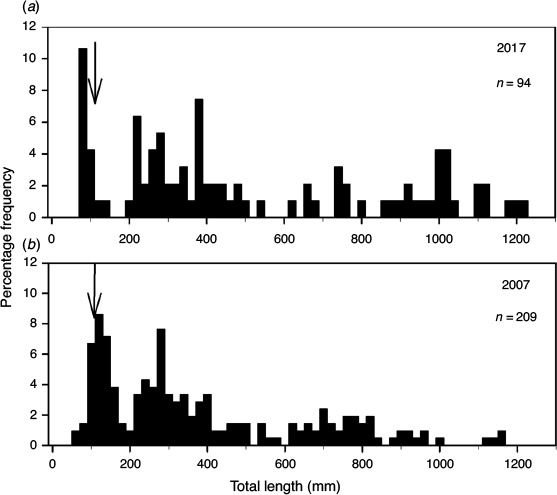
In spring 2016 and winter 2017, field surveys determined the relative abundance of YOY Murray cod before and after the environmental flow was implemented at the same six sites as for the larval netting. The 5.2-m electrofishing boat was equipped with an on-board 5.0-kVA Smith–Root Model GPP 5.0 H/L electrofishing system that usually operated at 1000 V DC, 7.5 A pulsed at 120 Hz and a 35% duty cycle. All boat electrofishing followed Sustainable River Audit sampling protocols with twelve 90-s ‘power-on’ replicates per site and one dip-netter at the front of the boat to collect stunned fish (Davies et al. 2010). All fish captured were weighed and measured (total length, TL) and then released. For broad comparison, the historic size structure of the Murray cod population is presented from the same region in 2007 (Wallace et al. 2008).
This work was conducted under Griffith University animal care and ethics committee permit ENV0414AEC.
Results
Environmental flow
Environmental flows from the Menindee Lakes into the LDR began in August 2016 and broadly matched the ecohydraulic conceptual model for Murray cod. Environmental water investments totalling 119.2 GL were contributed by various state and commonwealth agencies, with delivery aligned to the ecohydraulic conceptual model (Fig. 3). During September–December 2016, changes in water level at Weir 32 gauge (number 425012) on the LDR were relatively minor (i.e. maximum fall <0.05 m day–1) and matched the regime prescribed by the ecohydraulic conceptual model to support Murray cod nesting.
Leading up to and for the duration of the Murray cod nesting period (September–December 2016), discharge increased from ~400 ML day–1 (historically exceeded 50% of the time since 1974 at Weir 32 gauge) to a steady flow of ~850 ML day–1 (historically exceeded 39% of the time since 1974 at Weir 32 gauge) using NSW and Commonwealth environmental water. Between 1 September and 31 December 2016, water level stability was maintained, varying up or down by 0.01 m day–1 (s.d. 0.018 m day–1) with a maximum daily drop of <0.05 m and a maximum cumulative drop of 0.21 m.
From early December 2016, when monitoring had indicated Murray cod spawning was complete, the environmental flow was adaptively managed to include a rapid increase in discharge from 860 to 2253 ML day–1 in early December 2016 to stimulate downstream out-migration of golden perch from the Menindee Lakes and spawning in the LDR (Stuart and Sharpe 2020). To this point, the implemented flow regime closely matched the ecohydraulic conceptual model for Murray cod. In mid-January 2017, there was a second and larger rapid increase in releases of water from the Menindee Lakes to a maximum discharge of 6544 ML day–1 as part of operational demands in the lower Murray River. From January to April 2017, there was an attenuated flow recession in the LDR, after which there was a return to zero flows from mid-February 2019 to late March 2020.
Larval occurrence
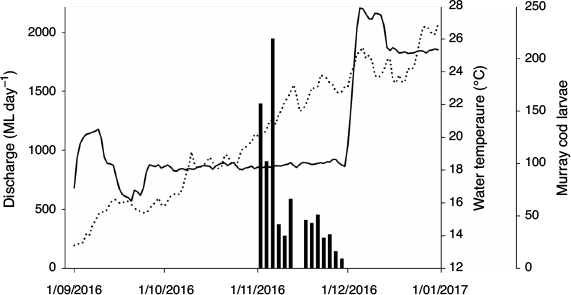
Seven species of fish eggs or larvae were collected from six trips at the six monitoring sites. Murray cod represented 23% of total catch and were present at all sites (Table 3). Bony herring, carp gudgeons (Hypseleotris spp.) and Australian smelt (Retropinna semoni) were also common. Murray cod were first detected on 2 November 2016 when the water temperature was 20.1°C (Fig. 4). No Murray cod larvae were collected after mid-December 2016, but golden perch were collected at this time during a rapid increase in river discharge. Most Murray cod larvae were collected from the drift nets set in the central part of the river channel.

|
|
The sampling sites commonly had abundant large woody debris and fast lotic water (mean 0.32 m s–1 in early November 2016; range 0.15–0.53 m s–1; discharge ~850 ML day–1). The size range of Murray cod larvae was 8–25 mm (mean ± s.d., 12.01 ± 2.86 mm) and, based on their developmental stage (Serafini and Humphries 2004), spawning was estimated to have occurred between 2 October and 28 November 2016 when flows were 833–918 day–1 and water level varied by <0.1 m.
In June 2017, YOY Murray cod were sampled with boat electrofishing at six sites in the LDR along with four other native fish species. In all, 117 Murray cod were collected, ranging in size from 70 to 1210 mm TL (mean ± s.d., 503 ± 350 mm TL) with YOY (based on a size of <115 mm TL) accounting for 14% of the fish sampled (Table 4; Fig. 5). For broad comparison, the size of fish from 2007, well before fish kills, is also provided (Wallace et al. 2008).

|
|
Post-fish kill restart flow
The LDR ceased to flow and major fish kills occurred between February 2019 and March 2020 (Australian Academy of Science 2019; Vertessy et al. 2019). When water returned to the Menindee Lakes in February 2020 from flooding in the northern Darling River catchments, there was an opportunity to restart the disconnected LDR. This was achieved with a fast, 7-day ramp-up of flows by 2 April 2020 to 3484 ML day–1 (i.e. ~40% bankfull; historically exceeded 25% of the time since 1974 at Weir 32 gauge) with a slow, 30-day recession back to a base flow of 200 ML day–1 by 3 May 2020. The rapid flow ramp-up was designed to push water through refuge pools as quickly as possible to restore connectivity and water quality. There was no evidence of further fish mortalities occurring during or immediately following the restart flow.
Discussion
Large semi-arid rivers present some of the most complex challenges for recovery, with flow regimes characterised by increasing water extraction, intermittency and highly stressed ecosystems (King and Brown 2006; Archdeacon and Reale 2020). Our case study of the first environmental flow delivered into the LDR in 2016–17 demonstrated spawning and recruitment of Murray cod and other native fish species. The present study also helped inform how recovery of a vulnerable native fish can be achieved by adaptively managing environmental flows, especially to support recovery after the 2018–20 catastrophic fish kills. For Murray cod recovery in the LDR, the priority was for a perennial flow regime, without cease-to-flow events, especially during the Murray cod spawning season, from which additional within-channel flow components can be added (Stuart et al. 2019).
Environmental flow outcomes
We hypothesise that the YOY Murray cod collected in the LDR can be explained by the 2016–17 environmental flow and were not present due to other causes (e.g. immigration) because the LDR was disconnected from the Murray River for 524 consecutive days; hence, prior migration into the 510-km study reach would likely be impossible, and no augmentation of the population by hatchery stocking had occurred since 2010–11 (New South Wales Department of Primary Industries Fisheries 2020). Downstream migration into the LDR was also an unlikely explanation of the occurrence of YOY in the study reach because there are few Murray cod in the ephemeral Menindee Lakes (Sharpe 2011). Otolith microchemistry also provides evidence of a self-sustaining local Murray cod population with few immigrants (Thiem et al. 2021). Hence, for Murray cod, local-scale interventions, such as environmental flows, are likely to be influential in post-fish kill recovery and long-term local population maintenance.
Spatial context of hydrodynamics and habitat
In the LDR, the environmental flows were applied over a large within-river spatial scale of 510 km, with lotic water dominating (>400 km) even at the lowest flows observed during this study (<100 ML day–1; historically exceeded 90% of the time since 1974 at Weir 32 gauge). Applying a perennial lotic flow regime, including continuous base flows, over relatively long river distances builds on previous work for Murray cod recovery at smaller (i.e. ~50 km) spatial scales (Stuart et al. 2019). Murray cod provide an instructive case study of life history flexibility, where populations may be able to be recovered at a broad range of spatial scales (i.e. tens to hundreds of kilometres). This insight suggests a range of transferable management opportunities for recovering specific reaches of the Murray–Darling Basin (e.g. the lower Goulburn River) or entire valleys (e.g. the Edward or Murrumbidgee rivers), particularly where Murray cod populations have declined. In these cases, discharge is the scalable tool that can be used to create perennial lotic habitats and hydraulic complexity, and there is potential to embed the designed ecohydraulic conceptual model for Murray cod within current consumptive flow management without affecting service to other river users.
Saving Murray cod
Our present understanding of the life history of Murray cod enabled development of an ecohydraulic conceptual model with clear relationships to four important functional elements of the hydrological and hydraulic flow regime of the LDR. During the design of the environmental flow regime, it was especially important to examine prior recruitment and hydrological data to optimise environmental flow conditions during the short (3-month) Murray cod breeding window in 2016. This included reviewing knowledge of flow strategies used elsewhere to support other nesting fish species with complex breeding strategies (Grabowski and Isely 2007; Butler and Rowland 2009; Couch et al. 2020).
One functional element of the flow regime that needed to be reinstated was a spring discharge rise to provide adults access to preferred spawning habitats, including lotic zones to build nests and brood young and for larval fish to disperse from the nest. A second element was an elevated summer discharge to inundate productive littoral habitats to support the rapid growth of young fish (Tonkin et al. 2017; Rowland 2020) followed by gradual summer flow recession. Continuous winter base flows likely also supported juvenile survival by increasing habitat availability and productivity and reducing predation rates (Stuart et al. 2019; Bestgen et al. 2020). Some key ecological knowledge gaps remain that could further inform designed flow hydrographs and recovery for Murray cod, including the preferred depth of nest sites, thresholds of water level variability to prevent nests being abandoned and minimum summer and winter base flows to improve juvenile survival.
Hydrological challenges
During regulated conditions at the Menindee Lakes, there were major interannual and interseasonal variations in discharge to the LDR, including two separate zero- or near-zero-flow events of 524 and 555 days. Catastrophic fish kills followed these zero-flow events and highlight that during dry conditions there is presently not enough water retained in the Darling system to avoid loss of Murray cod populations. Murray cod do not naturally inhabit ephemeral rivers (Rowland 2020) and so, in spring 2016, our primary objective was to provide an increase in discharge with continuous reach-scale (i.e. 500 km) lotic habitats. To sustain these vulnerable fish populations, protection of a perennial flow regime in the LDR is needed to improve survival of early life stages of Murray cod. We also support recent calls to maintain lotic hydraulics and hydraulic complexity (Mallen-Cooper and Zampatti 2020) and add the need for setting end-of-system flow and hydraulic targets at Burtundy to maintain the health of the LDR (New South Wales Department of Planning, Industry and Environment 2020; see Table 1).
An emergency element of the planned adaptive environmental flow regime in the LDR was required in March 2020 after a very long period (555 days) of zero, or near-zero, flows and major fish deaths (Vertessy et al. 2019). Following rainfall in the upper Darling catchment in 2020 and despite mass upstream extraction of water for agriculture, the Menindee Lakes partially refilled and the planned flow included a rapid ramp-up and relatively high discharge (i.e. ~40% bankfull) in the LDR. This restart flow enabled restoration of connectivity and improved water quality conditions to sustain fish that had survived the protracted cease-to-flow event and catastrophic fish kills that occurred in the LDR without further fish death events (Ellis et al. 2021). No further fish deaths were observed during the restart flow. Nevertheless, the restart flow was considered ‘high risk’ compared with preventing cease-to-flow and disconnected conditions by operating the LDR as perennial or near perennial, as per the natural flow regime (Mallen-Cooper and Zampatti 2020).
Fish deaths and the future of the LDR
The science for preserving large river ecosystems is clear: important elements of the natural flow and hydraulic regime must be protected, otherwise there will be severe damage to ecosystems (Poff et al. 1997; Ward et al. 2001). Unfortunately, this warning manifested, when the LDR was returned to zero flows and ‘artificial drought’ conditions (Boulton 2003) despite flow events occurring in upstream tributaries (Mallen-Cooper and Zampatti 2020). These zero flows, together with high daily air temperatures and toxic blue–green algae were major causal factors in catastrophic fish deaths (Vertessy et al. 2019; Jackson and Head 2020) and significant social impacts (Ellis et al. 2021). After the fish kills, to help recover Murray cod, an environmental flow was implemented in spring–summer 2020–21, informed by the present case study.
Can the natural character of the Darling River ecosystem be maintained while gross overallocation of water continues upstream? Our case study underscores a difficulty in answering this question. To redress overextraction of river flows and collapsing riverine ecosystems, governments must continue to provide economic compensation to acquire water from irrigators as willing sellers, with scientists and river managers designing flows with the goal of optimising ecological outcomes from reallocations of water for environmental purposes (Sisto 2009; Wheeler et al. 2014; Yarnell et al. 2015). In practice, large semi-arid rivers still present major restoration challenges because water resources are often fully utilised and there has been human-made intensification of droughts, with added stress from declining rainfall patterns (Propst et al. 2008; Arthington and Balcombe 2011). Aquatic ecosystems now require protection from these ‘artificial droughts’ (Boulton 2003). For recovery of semi-arid rivers, a key is to apply both environmental and operational water in a manner that sustains ecosystems while supporting regional community needs (Bestgen et al. 2020).
New directions
Ecosystem recovery is a common shared objective of environmental flows and, on the LDR, the designed and adaptively implemented flow regime appeared to support the life history of Murray cod. The most likely cause for enhanced Murray cod survival was implementation of the annual environmental flow, including promotion of perennial lotic habitats. Further multiyear experimental work is currently being conducted to explore: (1) juvenile survival rates at much reduced spring discharge (e.g. 400 ML day–1), which appears a more common future scenario; (2) tolerances of adult fish to river level variation during nesting; (3) the value of summer and winter continuous base flows to improve connectivity, water quality and fish survival rates; and (4) the inundation of flood plains, which may be important for enhancing productivity. Our case study addresses only a few of these aspects but may be instructive for future planning for other perennial lotic specialists, such as silver perch, river mussel and Darling River snail.
Conclusion
The challenges facing conservation of imperilled Murray cod in the LDR are emblematic of global losses of arid and semi-arid river fishes, commonly as a direct result of human overallocation of flows, exacerbated by climate change (Archdeacon and Reale 2020). In particular, serious overabstraction of water in the upper Darling catchment has contributed to an increased frequency and duration of zero-flow events in the LDR resulting in major fish kills (Vertessy et al. 2019), compromising major environmental assets (Kingsford 2000; Jackson and Head 2020). Our study highlights that Murray cod populations can potentially be protected and recovered with environmental flows that adaptively promote base flows, flow pulses, perennial lotic conditions, hydraulic complexity and prevention of cease-to-flow conditions. For the LDR, these flow components must be incorporated into planning for a long-term flow regime. Failing to provide perennial lotic habitats for Murray cod by maintaining current water diversion policies will ultimately fail to protect this nationally significant species and other aquatic biota. Across the Tasman Sea, the Whanganui River of New Zealand was recently granted legal ‘personhood’ asserting the rights of Nature (Brierley et al. 2019). For the Darling River, such a profoundly different approach may be needed to protect the long-term health of a nationally significant native fish and its ecosystem.
Conflicts of interest
The authors declare that they have no conflicts of interest.
Declaration of funding
Funding was provided by the Commonwealth Environmental Water Office and the Murray–Darling Basin Authority.
Acknowledgements
Sincere thanks to Paula D’Santos (NSW DPIE), Iain Ellis (NSW DPI Fisheries) and Vince Kelly (WaterNSW) for local support in planning and implementing the environmental flows. The authors thank Steve Campbell-Brown for field assistance. John Koehn, from the Arthur Rylah Institute, Rob Rolls, an anonymous referee and the handling editor, Craig Boys, all provided valuable comments to improve the manuscript.
References
Acreman, M., Arthington, A. H., Colloff, M. J., Couch, C., Crossman, N. D., Dyer, F., Overton, I., Pollino, C. A., Stewardson, M. J., and Young, W. (2014). Environmental flows for natural, hybrid, and novel riverine ecosystems in a changing world. Frontiers in Ecology and the Environment 12, 466–473.| Environmental flows for natural, hybrid, and novel riverine ecosystems in a changing world.Crossref | GoogleScholarGoogle Scholar |
Anderson, J. R., Morison, A. K., and Ray, D. J. (1992). Age and growth of Murray cod, Maccullochella peelii (Perciformes: Percichthyidae), in the lower Murray–Darling Basin, Australia, from thin-sectioned otoliths. Marine and Freshwater Research 43, 983–1013.
| Age and growth of Murray cod, Maccullochella peelii (Perciformes: Percichthyidae), in the lower Murray–Darling Basin, Australia, from thin-sectioned otoliths.Crossref | GoogleScholarGoogle Scholar |
Archdeacon, T. P., and Reale, J. K. (2020). No quarter: lack of refuge during flow intermittency results in catastrophic mortality of an imperiled minnow. Freshwater Biology 65, 2108–2123.
| No quarter: lack of refuge during flow intermittency results in catastrophic mortality of an imperiled minnow.Crossref | GoogleScholarGoogle Scholar |
Arthington, A. H., and Balcombe, S. R. (2011). Extreme flow variability and the ‘boom and bust’ ecology of fish in arid-zone floodplain rivers: a case history with implications for environmental flows, conservation and management. Ecohydrology 4, 708–720.
| Extreme flow variability and the ‘boom and bust’ ecology of fish in arid-zone floodplain rivers: a case history with implications for environmental flows, conservation and management.Crossref | GoogleScholarGoogle Scholar |
Australian Academy of Science (2019). Investigation of the causes of mass fish kills in the Menindee Region NSW over the summer of 2018–2019. (AAS: Canberra, ACT, Australia.) Available at https://www.science.org.au/supporting-science/science-policy-and-sector-analysis/reports-and publications/fish-kills-report
Bestgen, K. R., Poff, N. L., Baker, D. W., Bledsoe, B. P., Merritt, D. M., Lorie, M., Auble, G. T., Sanderson, J. S., and Kondratieff, B. C. (2020). Designing flows to enhance ecosystem functioning in heavily altered rivers. Ecological Applications 30, e02005.
| Designing flows to enhance ecosystem functioning in heavily altered rivers.Crossref | GoogleScholarGoogle Scholar | 31532056PubMed |
Boulton, A. J. (2003). Parallels and contrasts in the effects of drought on stream macroinvertebrate assemblages. Freshwater Biology 48, 1173–1185.
| Parallels and contrasts in the effects of drought on stream macroinvertebrate assemblages.Crossref | GoogleScholarGoogle Scholar |
Brierley, G., Tadaki, M., Hikuroa, D., Blue, B., Šunde, C., Tunnicliffe, J., and Salmond, A. (2019). A geomorphic perspective on the rights of the river in Aotearoa New Zealand. River Research and Applications 35, 1640–1651.
| A geomorphic perspective on the rights of the river in Aotearoa New Zealand.Crossref | GoogleScholarGoogle Scholar |
Butler, G. L., and Rowland, S. J. (2009). Using underwater cameras to describe the reproductive behaviour of the endangered eastern freshwater cod Maccullochella ikei. Ecology Freshwater Fish 18, 337–349.
| Using underwater cameras to describe the reproductive behaviour of the endangered eastern freshwater cod Maccullochella ikei.Crossref | GoogleScholarGoogle Scholar |
Couch, A. J., Dyer, F., and Lintermans, M. (2020). Multi-year pair-bonding in Murray cod (Maccullochella peelii). PeerJ 8, e10460.
| Multi-year pair-bonding in Murray cod (Maccullochella peelii).Crossref | GoogleScholarGoogle Scholar | 33354425PubMed |
Davies, P. E., Harris, J. H., Hillman, T. J., and Walker, K. F. (2010). The Sustainable Rivers Audit: assessing river ecosystem health in the Murray–Darling Basin, Australia. Marine and Freshwater Research 61, 764–777.
| The Sustainable Rivers Audit: assessing river ecosystem health in the Murray–Darling Basin, Australia.Crossref | GoogleScholarGoogle Scholar |
Ellis, I., and Meredith, S. (2004). Guidelines for future release effects on Lower Darling River fish deaths. Consultancy report for NSW Department of Infrastructure, Planning and Natural Resources. Murray–Darling Freshwater Research Centre, Mildura, Vic., Australia.
Ellis, I., Bates, W., Martin, S., McCrabb, G., Koehn, J., Heath, P., and Hardman, D. (2021). How fish kills affected traditional (Baakandji) and non-traditional communities on the Lower Darling–Baaka River. Marine and Freshwater Research , .
| How fish kills affected traditional (Baakandji) and non-traditional communities on the Lower Darling–Baaka River.Crossref | GoogleScholarGoogle Scholar |
Forbes, J., Watts, R. J., Robinson, W. A., Baumgartner, L. J., McGuffie, P., Cameron, L. M., and Crook, D. A. (2016). Assessment of stocking effectiveness for Murray cod (Maccullochella peelii) and golden perch (Macquaria ambigua) in rivers and impoundments of south-eastern Australia. Marine and Freshwater Research 67, 1410–1419.
| Assessment of stocking effectiveness for Murray cod (Maccullochella peelii) and golden perch (Macquaria ambigua) in rivers and impoundments of south-eastern Australia.Crossref | GoogleScholarGoogle Scholar |
Gido, K. B., Propst, D. L., Olden, J. D., and Bestgen, K. R. (2013). Multidecadal responses of native and introduced fishes to natural and altered flow regimes in the American Southwest. Canadian Journal of Fisheries and Aquatic Sciences 70, 554–564.
| Multidecadal responses of native and introduced fishes to natural and altered flow regimes in the American Southwest.Crossref | GoogleScholarGoogle Scholar |
Grabowski, T. B., and Isely, J. J. (2007). Effects of flow fluctuations on the spawning habitat of a riverine fish. Southeastern Naturalist 6, 471–478.
| Effects of flow fluctuations on the spawning habitat of a riverine fish.Crossref | GoogleScholarGoogle Scholar |
Grill, G., Lehner, B., Thieme, M., Geenen, B., Tickner, D., Antonelli, F., Babu, S., Borrelli, P., Cheng, L., Crochetiere, H., and Macedo, H. E. (2019). Mapping the world’s free-flowing rivers. Nature 569, 215–221.
| Mapping the world’s free-flowing rivers.Crossref | GoogleScholarGoogle Scholar | 31068722PubMed |
He, F., Zarfl, C., Bremerich, V., David, J. N., Hogan, Z., Kalinkat, G., Tockner, K., and Jähnig, S. C. (2019). The global decline of freshwater megafauna. Global Change Biology 25, 3883–3892.
| The global decline of freshwater megafauna.Crossref | GoogleScholarGoogle Scholar | 31393076PubMed |
Jackson, S., and Head, L. (2020). Australia’s mass fish kills as a crisis of modern water: understanding hydrosocial change in the Murray–Darling Basin. Geoforum 109, 44–56.
| Australia’s mass fish kills as a crisis of modern water: understanding hydrosocial change in the Murray–Darling Basin.Crossref | GoogleScholarGoogle Scholar |
Jenkins, K. M., and Boulton, A. J. (2003). Connectivity in a dryland river: short-term aquatic microinvertebrate recruitment following floodplain inundation. Ecology 84, 2708–2723.
| Connectivity in a dryland river: short-term aquatic microinvertebrate recruitment following floodplain inundation.Crossref | GoogleScholarGoogle Scholar |
Jones, M. J., and Stuart, I. G. (2007). Movements and habitat use of common carp (Cyprinus carpio) and Murray cod (Maccullochella peelii peelii) juveniles in a large lowland Australian river. Ecology Freshwater Fish 16, 210–220.
| Movements and habitat use of common carp (Cyprinus carpio) and Murray cod (Maccullochella peelii peelii) juveniles in a large lowland Australian river.Crossref | GoogleScholarGoogle Scholar |
King, J., and Brown, C. (2006). Environmental flows: striking the balance between development and resource protection. Ecology and Society 11, art26.
| Environmental flows: striking the balance between development and resource protection.Crossref | GoogleScholarGoogle Scholar |
King, A. J., Tonkin, Z., and Mahoney, J. (2009). Environmental flows enhance native fish spawning and recruitment in the Murray River, Australia. River Research and Applications 25, 1205–1218.
| Environmental flows enhance native fish spawning and recruitment in the Murray River, Australia.Crossref | GoogleScholarGoogle Scholar |
Kingsford, R. T. (2000). Ecological impacts of dams, water diversions and river management on floodplain wetlands in Australia. Austral Ecology 25, 109–127.
| Ecological impacts of dams, water diversions and river management on floodplain wetlands in Australia.Crossref | GoogleScholarGoogle Scholar |
Koehn, J. D. (2004). The loss of valuable Murray cod in fish kills: a science and management perspective. In ‘Management of Murray Cod in the Murray–Darling Basin, Statement, Recommendations and Supporting Papers Proceedings of a Workshop’, 3–4 June 2004, Canberra, ACT, Australia. (Eds M. Lintermans and B. Phillips.) pp. 73–82. (Murray–Darling Basin Commission: Canberra, ACT, Australia.)
Koehn, J. D. (2009). Multi-scale habitat selection by Murray cod Maccullochella peelii peelii in two lowland rivers Journal of Fish Biology 75, 113–129.
| Multi-scale habitat selection by Murray cod Maccullochella peelii peelii in two lowland riversCrossref | GoogleScholarGoogle Scholar | 20738486PubMed |
Koehn, J. D., and Harrington, D. J. (2005). Collection and distribution of the early life stages of the Murray cod (Maccullochella peelii peelii) in a regulated river. Australian Journal of Zoology 53, 137–144.
| Collection and distribution of the early life stages of the Murray cod (Maccullochella peelii peelii) in a regulated river.Crossref | GoogleScholarGoogle Scholar |
Koehn, J. D., and Harrington, D. J. (2006). Environmental conditions and timing for the spawning of Murray cod (Maccullochella peelii peellii) and the endangered trout cod (M. macquariensis) in southeastern Australian rivers. River Research and Applications 22, 327–342.
| Environmental conditions and timing for the spawning of Murray cod (Maccullochella peelii peellii) and the endangered trout cod (M. macquariensis) in southeastern Australian rivers.Crossref | GoogleScholarGoogle Scholar |
Koehn, J. D., Raymond, S. A., Stuart, I., Todd, C. R., Balcombe, S. R., Zampatti, B. P., Bamford, H., Ingram, B. A., Bice, C., Burndred, K., Butler, G., Baumgartner, L., Clunie, P., Ellis, I., Forbes, J., Hutchison, M., Koster, W., Lintermans, M., Lyon, J. P., Mallen-Cooper, M., McLellan, M., Pearce, L., Ryall, J., Sharpe, C., Stoessel, D. J., Thiem, J. D., Tonkin, Z., Townsend, A., and Ye, Q. (2020). A compendium of ecological knowledge for restoration of freshwater fishes in the Murray–Darling Basin. Marine and Freshwater Research 71, 1391–1463.
| A compendium of ecological knowledge for restoration of freshwater fishes in the Murray–Darling Basin.Crossref | GoogleScholarGoogle Scholar |
Leblanc, M., Tweed, S., Van Dijk, A., and Timbal, B. (2012). A review of the historic and future hydrological changes in the Murray–Darling Basin. Global and Planetary Change 80–81, 226–246.
| A review of the historic and future hydrological changes in the Murray–Darling Basin.Crossref | GoogleScholarGoogle Scholar |
Lieschke, J. A., Lyon, J. P., Moloney, P. D., and Nicol, S. J. (2016). Spatial partitioning in the use of structural woody habitat supports the cohabitation of two cod species in a large lowland river. Marine and Freshwater Research 67, 1835–1843.
| Spatial partitioning in the use of structural woody habitat supports the cohabitation of two cod species in a large lowland river.Crossref | GoogleScholarGoogle Scholar |
Lyon, J. P., and O’Connor, J. P. (2008). Smoke on the water: can riverine fish populations recover following a catastrophic fire-related sediment slug? Austral Ecology 33, 794–806.
| Smoke on the water: can riverine fish populations recover following a catastrophic fire-related sediment slug?Crossref | GoogleScholarGoogle Scholar |
Lyon, J. P., Bird, T., Tonkin, Z., Raymond, S., Sharley, J., and Hale, R. (2021). Does life history mediate the effects of discharge as a driver of multi-decadal changes in population abundance and size structure of freshwater fish? Ecological Applications , e02430.
| Does life history mediate the effects of discharge as a driver of multi-decadal changes in population abundance and size structure of freshwater fish?Crossref | GoogleScholarGoogle Scholar | 34309984PubMed |
Mallen-Cooper, M., and Zampatti, B. P. (2020). Restoring the ecological integrity of a dryland river: why low flows in the Barwon-Darling River must flow. Ecological Management & Restoration 21, 218–228.
| Restoring the ecological integrity of a dryland river: why low flows in the Barwon-Darling River must flow.Crossref | GoogleScholarGoogle Scholar |
Moyle, P. B. (2014). Novel aquatic ecosystems: the new reality for streams in California and other Mediterranean climate regions. River Research and Applications 30, 1335–1344.
| Novel aquatic ecosystems: the new reality for streams in California and other Mediterranean climate regions.Crossref | GoogleScholarGoogle Scholar |
New South Wales Department of Planning, Industry and Environment (2020). Murray–Lower Darling long-term water plan part A: Murray–Lower Darling catchment.
New South Wales Department of Primary Industries Fisheries (2020). Fish Stocking Database. Available at https://www.dpi.nsw.gov.au/fishing/recreational/resources/stocking [Verified 18 December 2020].
Nilsson, C., Reidy, C. A., Dynesius, M., and Revenga, C. (2005). Fragmentation and flow regulation of the world’s large river systems. Science 308, 405–408.
| Fragmentation and flow regulation of the world’s large river systems.Crossref | GoogleScholarGoogle Scholar | 15831757PubMed |
Petriki, O., Patsia, A., Vafeiadou, A., Tekidis, I., Michalopoulou, P., Samartzi, S., Mallinis, D., and Bobori, D. C. (2021). Tracking the causes of a mass fish kill at a Mediterranean River within a protected area. Water 13, 989.
| Tracking the causes of a mass fish kill at a Mediterranean River within a protected area.Crossref | GoogleScholarGoogle Scholar |
Poff, N. L., Allan, J. D., Bain, M. B., Karr, J. R., Prestegaard, K. L., Richter, B. D., Sparks, R. E., and Stromberg, J. C. (1997). The natural flow regime. Bioscience 47, 769–784.
| The natural flow regime.Crossref | GoogleScholarGoogle Scholar |
Propst, D. L., Gido, K. B., and Stefferud, J. A. (2008). Natural flow regimes, nonnative fishes, and native fish persistence in arid-land river systems. Ecological Applications 18, 1236–1252.
| Natural flow regimes, nonnative fishes, and native fish persistence in arid-land river systems.Crossref | GoogleScholarGoogle Scholar | 18686584PubMed |
Richter, B. D., and Thomas, G. A. (2007). Restoring environmental flows by modifying dam operations. Ecology and Society 12, art12.
| Restoring environmental flows by modifying dam operations.Crossref | GoogleScholarGoogle Scholar |
Richter, B. D., Andrews, S., Dahlinghaus, R., Freckmann, G., Ganis, S., Green, J., Hardman, I., Palmer, M., and Shalvey, J. (2020). Buy me a river: purchasing water rights to restore river flows in the Western USA. Journal of the American Water Resources Association 56, 1–15.
| Buy me a river: purchasing water rights to restore river flows in the Western USA.Crossref | GoogleScholarGoogle Scholar |
Rowland, S. (1988). Hormone-induced spawning of the Australian freshwater fish Murray Cod, Maccullochella peeli (Mitchell) (Percichthyidae). Aquaculture 70, 371–389.
| Hormone-induced spawning of the Australian freshwater fish Murray Cod, Maccullochella peeli (Mitchell) (Percichthyidae).Crossref | GoogleScholarGoogle Scholar |
Rowland, S. (1998). Aspects of the reproductive biology of Murray cod, Maccullochella peelii peelii. Proceedings of the Linnean Society of New South Wales 120, 147–162.
Rowland, S. (2004). Overview of the history, fishery, biology and aquaculture of Murray cod (Maccullochella peelii peelii). In ‘Management of Murray Cod in the Murray–Darling Basin’. Statement, Recommendations and Supporting Papers Proceedings of a Workshop held in Canberra, ACT, Australia. (Eds M. Lintermans, and B. Phillips.) pp 38–61. (Murray–Darling Basin Commission: Canberra, ACT, Australia.)
Rowland, S. J. (2020). ‘The Codfather. A Life Dedicated to the Study and Conservation of Australian Freshwater Fish.’ (Optima Press: Perth, WA, Australia.)
Sargent, J. C., and Galat, D. L. (2002). Fish mortality and physicochemistry in a managed floodplain wetland. Wetlands Ecology and Management 10, 113–119.
| Fish mortality and physicochemistry in a managed floodplain wetland.Crossref | GoogleScholarGoogle Scholar |
Serafini, L., and Humphries, P. (2004). Preliminary guide to the identification of larvae of fish, with a bibliography of their studies, from the Murray–Darling Basin. Presented at the taxonomy workshop, Lake Hume resort, Cooperative Research Centre for Freshwater Ecology.
Sharpe, C. P. (2011). Spawning and recruitment ecology of golden perch (Macquaria ambigua Richardson, 1845) in the Murray and Darling Rivers. Ph.D. Thesis, Griffith University, Nathan, Qld, Australia.
Sisto, N. P. (2009). Environmental flows for rivers and economic compensation for irrigators. Journal of Environmental Management 90, 1236–1240.
| Environmental flows for rivers and economic compensation for irrigators.Crossref | GoogleScholarGoogle Scholar | 18678441PubMed |
Stoffels, R. J., Weatherman, K. E., Bond, N. R., Morrongiello, J. R., Thiem, J. D., Butler, G., Koster, W., Kopf, R. K., McCasker, N., Ye, Q., and Zampatti, B. (2020). Stage-dependent effects of river flow and temperature regimes on the growth dynamics of an apex predator. Global Change Biology 26, 6880–6894.
| Stage-dependent effects of river flow and temperature regimes on the growth dynamics of an apex predator.Crossref | GoogleScholarGoogle Scholar | 32970901PubMed |
Stuart, I., and Sharpe, C. (2020). Riverine spawning, long distance larval drift and floodplain recruitment of a pelagophilic fish, a case study of golden perch (Macquaria ambigua) in the arid Darling River, Australia. Aquatic Conservation 30, 675–690.
| Riverine spawning, long distance larval drift and floodplain recruitment of a pelagophilic fish, a case study of golden perch (Macquaria ambigua) in the arid Darling River, Australia.Crossref | GoogleScholarGoogle Scholar |
Stuart, I., Sharpe, C., Stanislawski, K., Parker, A., and Mallen-Cooper, M. (2019). From an irrigation system to an ecological asset: adding environmental flows establishes recovery of a threatened fish species. Marine and Freshwater Research 70, 1295–1306.
| From an irrigation system to an ecological asset: adding environmental flows establishes recovery of a threatened fish species.Crossref | GoogleScholarGoogle Scholar |
Thiem, J. D., Wooden, I. J., Baumgartner, L. J., Butler, G. L., Forbes, J. P., and Conallin, J. (2017). Recovery from a fish kill in a semi-arid Australian river: can stocking augment natural recruitment processes? Austral Ecology 42, 218–226.
| Recovery from a fish kill in a semi-arid Australian river: can stocking augment natural recruitment processes?Crossref | GoogleScholarGoogle Scholar |
Thiem, J. D., Baumgartner, L. J., Fanson, B., Sadekov, A., Tonkin, Z., and Zampatti, B. P. (2021). Contrasting natal origin and movement history informs recovery pathways for three lowland river species following a mass fish kill. Marine and Freshwater Research , .
| Contrasting natal origin and movement history informs recovery pathways for three lowland river species following a mass fish kill.Crossref | GoogleScholarGoogle Scholar |
Thoms, M. C., and Sheldon, F. (2000). Water resource development and hydrological change in a large dryland river: the Barwon–Darling River, Australia. Journal of Hydrology 228, 10–21.
| Water resource development and hydrological change in a large dryland river: the Barwon–Darling River, Australia.Crossref | GoogleScholarGoogle Scholar |
Tockner, K., and Stanford, J. A. (2002). Riverine flood plains: present state and future trends. Environmental Conservation 29, 308–330.
| Riverine flood plains: present state and future trends.Crossref | GoogleScholarGoogle Scholar |
Tonkin, Z., Kitchingman, A., Lyon, J., Kearns, J., Hackett, G., O’Mahony, J., and Bird, T. (2017). Flow magnitude and variability influence growth of two freshwater fish species in a large regulated floodplain river. Hydrobiologia 797, 289–301.
| Flow magnitude and variability influence growth of two freshwater fish species in a large regulated floodplain river.Crossref | GoogleScholarGoogle Scholar |
Tonkin, Z., Stuart, I., Kitchingman, A., Thiem, J. D., Zampatti, B., Hackett, G., Koster, W., Koehn, J., Morrongiello, J., Mallen-Cooper, M., and Lyon, J. (2019). Hydrology and water temperature influence recruitment dynamics of the threatened silver perch Bidyanus bidyanus in a regulated lowland river. Marine and Freshwater Research 70, 1333–1344.
| Hydrology and water temperature influence recruitment dynamics of the threatened silver perch Bidyanus bidyanus in a regulated lowland river.Crossref | GoogleScholarGoogle Scholar |
Tonkin, Z., Yen, J., Lyon, J., Kitchingman, A., Koehn, J. D., Koster, W. M., Lieschke, J., Raymond, S., Sharley, J., Stuart, I., and Todd, C. (2021). Linking flow attributes to recruitment to inform water management for an Australian freshwater fish with an equilibrium life-history strategy. The Science of the Total Environment 752, 141863.
| Linking flow attributes to recruitment to inform water management for an Australian freshwater fish with an equilibrium life-history strategy.Crossref | GoogleScholarGoogle Scholar | 32889283PubMed |
Vertessy, R., Barma, D., Baumgartner, L., Mitrovic, S., Sheldon, F., and Bond, N. (2019). Independent assessment of the 2018–19 fish deaths in the lower Darling River. (Australian Government: Canberra, ACT, Australia.) Available at https://www.mdba.gov.au/publications/mdba-reports/response-fish-deaths-lower-darling
Vilizzi, L., Meredith, S. N., Sharpe, C. P., and Rehwinkel, R. (2008). Evaluating light trap efficiency by application of mesh to prevent inter-and intra-specific in situ predation on fish larvae and juveniles. Fisheries Research 93, 146–153.
| Evaluating light trap efficiency by application of mesh to prevent inter-and intra-specific in situ predation on fish larvae and juveniles.Crossref | GoogleScholarGoogle Scholar |
Vörösmarty, C. J., McIntyre, P. B., Gessner, M. O., Dudgeon, D., Prusevich, A., Green, P., Glidden, S., Bunn, S. E., Sullivan, C. A., Liermann, C. R., and Davies, P. M. (2010). Global threats to human water security and river biodiversity. Nature 467, 555–561.
| Global threats to human water security and river biodiversity.Crossref | GoogleScholarGoogle Scholar | 20882010PubMed |
Walker, K. F., Sheldon, F., and Puckridge, J. T. (1995). A perspective on dryland river ecosystems. Regulated Rivers 11, 85–104.
| A perspective on dryland river ecosystems.Crossref | GoogleScholarGoogle Scholar |
Wallace, T., Sharpe, C., Fraser, P., Rehwinkel, R., and Vilizzi, L. (2008). The impact of drought on water quality and fish communities within refuge pools on the Lower Darling River. A technical report prepared for the Lower Murray Darling Catchment Management Authority by the Murray–Darling Freshwater Research Centre.
Ward, J. V., Tockner, K., Uehlinger, U., and Malard, F. (2001). Understanding natural patterns and processes in river corridors as the basis for effective river restoration. Regulated Rivers 17, 311–323.
| Understanding natural patterns and processes in river corridors as the basis for effective river restoration.Crossref | GoogleScholarGoogle Scholar |
Watts, R. J., Keller Kopf, R., McCasker, N., Howitt, J. A., Conallin, J., Wooden, I., and Baumgartner, L. (2018). Adaptive management of environmental flows: using irrigation infrastructure to deliver environmental benefits during a large hypoxic blackwater event in the Southern Murray–Darling Basin, Australia. Environmental Management 61, 469–480.
| Adaptive management of environmental flows: using irrigation infrastructure to deliver environmental benefits during a large hypoxic blackwater event in the Southern Murray–Darling Basin, Australia.Crossref | GoogleScholarGoogle Scholar | 28929206PubMed |
Webb, J. A., Arthington, A. H., and Olden, J. D. (2017). Models of ecological responses to flow regime change to inform environmental flows assessments. In ‘Water for the Environment’. pp. 287–316. (Academic Press.)
Wheeler, S. A., Zuo, A., and Bjornlund, H. (2014). Investigating the delayed on-farm consequences of selling water entitlements in the Murray–Darling Basin. Agricultural Water Management 145, 72–82.
| Investigating the delayed on-farm consequences of selling water entitlements in the Murray–Darling Basin.Crossref | GoogleScholarGoogle Scholar |
Whitworth, K. L., Baldwin, D. S., and Kerr, J. L. (2012). Drought, floods and water quality: drivers of a severe hypoxic blackwater event in a major river system (the southern Murray–Darling Basin, Australia). Journal of Hydrology 450–451, 190–198.
| Drought, floods and water quality: drivers of a severe hypoxic blackwater event in a major river system (the southern Murray–Darling Basin, Australia).Crossref | GoogleScholarGoogle Scholar |
Yarnell, S. M., Petts, G. E., Schmidt, J. C., Whipple, A. A., Beller, E. E., Dahm, C. N., Goodwin, P., and Viers, J. H. (2015). Functional flows in modified riverscapes: hydrographs, habitats and opportunities. Bioscience 65, 963–972.
| Functional flows in modified riverscapes: hydrographs, habitats and opportunities.Crossref | GoogleScholarGoogle Scholar |
Yarnell, S. M., Stein, E. D., Webb, J. A., Grantham, T., Lusardi, R. A., Zimmerman, J., Peek, R. A., Lane, B. A., Howard, J., and Sandoval-Solis, S. (2020). A functional flows approach to selecting ecologically relevant flow metrics for environmental flow applications. River Research and Applications 36, 318–324.
| A functional flows approach to selecting ecologically relevant flow metrics for environmental flow applications.Crossref | GoogleScholarGoogle Scholar |