

Co-design and Traditional Owner participation in an assessment of abundance and size of Donax deltoides (garlaany, pipi) in Ngambaa Country, New South Wales, Australia
Jade Fredericks A * , Hannah B. Smith B , Kirsten Benkendorff A , Anna Scott A and Karina C. Hall A C
A * , Hannah B. Smith B , Kirsten Benkendorff A , Anna Scott A and Karina C. Hall A C
A
B
C
Abstract
Integration of Indigenous ecological knowledge (IEK) and Traditional Owner participation in the co-design of field-sampling techniques can optimise data collection on fisheries resources, that may be under-represented when using conventional scientific knowledge (CSK) alone.
To co-design an IEK-informed sampling methodology to assess counts and size–frequency distribution of an aggregated sandy shore mollusc, Donax deltoides (garlaany, pipi).
We worked with Traditional Owners to co-design a sampling regime synonymous with traditional harvesting techniques. This method was then applied to an assessment of D. deltoides on the Mid North Coast, New South Wales, Australia, at Stuarts Point Beach, over 13 months from November 2021 to November 2022.
This co-designed methodology differed from CSK by using non-random site selection and sampling techniques informed by cultural practice. The standardised methodology was repeatable, quantitative and successfully detected significant spatial and temporal variation in D. deltoides counts and size–frequency distribution, as well as correlations with environmental variables.
A co-designed framework that integrates IEK can provide quantitative assessments of counts and size–frequency distributions in an exploited aggregated sandy shore mollusc.
IEK and Traditional Owner participation can facilitate scientifically rigorous data collection on culturally important species, while fostering stewardship and co-management.
Keywords: bivalve, cultural harvesting techniques, environmental indicators, fisheries assessment, IEK, Indigenous ecological knowledge, infauna, population biology, sandy shores, soft sediment communities.
Introduction
First Nations people are being increasingly recognised as the first scientists, with knowledge systems that reflect a deep understanding of the complexity and inter-connectedness between people and the environment (Nursey-Bray 2011; Prober et al. 2011; Berkes 2012). Collaboration with Traditional Owners in co-designing and executing research projects is therefore of great value (Schemmel et al. 2016). This is particularly true for biological and cultural resources that are becoming harder to detect and accurately assess because of population declines resulting from increasing anthropogenic pressure and environmental change (Pauly and Zeller 2016; Carvalho et al. 2017; Food and Agriculture Organization of the United Nations 2020). Traditional Owners, many of whom still live in their clan-based territories, have intimate knowledge of the landscape and are intrinsically linked with these environments (Teixeira et al. 2013). This amalgamation of knowledge, referred to as Indigenous ecological knowledge (IEK), forms the basis for more complex traditional management and cultural harvesting strategies (Golden et al. 2014; Chakraborty and Gasparatos 2019).
Cultural harvesting techniques that have been utilised and evolved over generations include information on how and when to catch species by using detailed biocultural knowledge. Other IEK includes information on reproductive strategies and niche utilisation that aim to minimise harvesting effort while conserving resources (Deepananda et al. 2016; Tavares et al. 2020). When traditional harvesting techniques can be quantified using conventional scientific knowledge (CSK) into a co-designed framework, optimisation of scientific research strategies and sampling regimes can occur over broad and localised spatial and temporal scales (Gaspare et al. 2015; Schemmel et al. 2016; Lima et al. 2017). This may be particularly useful for species that have distributions that vary in space and time, with population dynamics that can be difficult to predict and may therefore be under-represented when using standard ecological sampling regimes.
Indigenous ecological knowledge accumulates over time, evolving over generations through interactions with the environment. In many instances, these interactions are entwined with cultural practices, leading to spiritual connections to specific natural resources (Berkes 2012; Donda and Manyungwa-Pasani 2018; Turner 2021). However, the evolution of IEK, formation of cultural harvesting strategies and continuation of cultural practices, require target species to be ‘in situ’, to be extensively used and persist through time. In some cases, this supports the transitions of a targeted harvested species to a cultural keystone species (Berkes 2012). Cultural keystone species are of significance not only as subsistence resources, but are also pivotal to stories, songs, ethic or moral tales and support the continuation and evolution of cultural practises (Garibaldi and Turner 2004; Noble et al. 2018; McKemey et al. 2019). Cultural harvesting strategies for cultural keystone species utilise IEK recorded on, but not limited to, biological, ecosystem and seasonal parameters (Schemmel et al. 2016; Ban et al. 2017; Eckert et al. 2018). Further to this, harvesting strategies use techniques specific to target catch, which can include size and sex of individuals on the basis of this knowledge. Fish traps and eel farms located up and down the eastern coast of Australia are evidence of the selective use of targeted cultural knowledge that has evolved over thousands of years (Cruse et al. 2005).
Donax deltoides (garlaany, pipi) is a cultural keystone species for Aboriginal communities of the eastern and southern coast of Australia, being important as a subsistence resource and for the transference of IEK (Sullivan 1978; Schnierer and Egan 2016). Historical evidence of the use of D. deltoides exists in midden sites along the eastern coast of Australia dating back to at least 9000 BCE (Sullivan 1985; McNiven 1989; Knuckey 1999). Contemporary knowledge has recorded D. deltoides along the eastern coast of Australia, stretching from Fraser Island in Queensland to Young Husband Peninsula in South Australia (Murray-Jones and Ayre 1997).
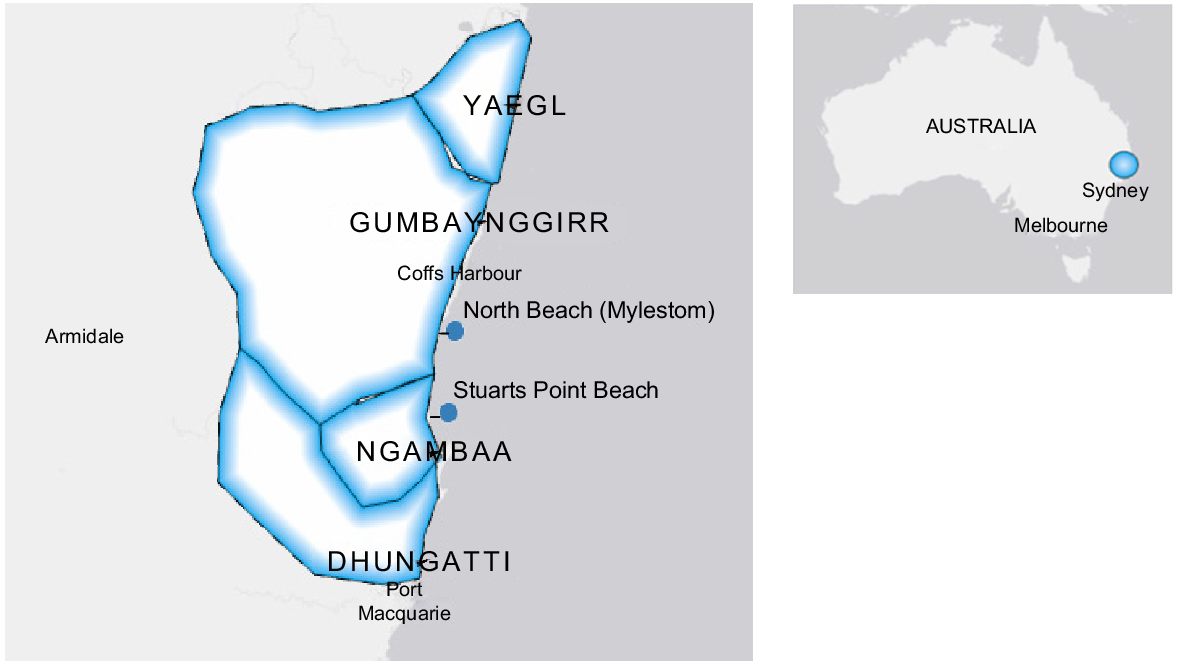
Donax deltoides is a short-lived, fast-growing ‘r-selected’ species (Murray-Jones and Ayre 1997; Versace et al. 2011), with pelagic larvae being produced by dribble spawning through-out the year and recruitment peaks occurring in the austral spring (King 1976; Murray-Jones 1999). As a benthic species that spends most of its time buried under the sand on wave-exposed beaches, D. deltoides populations can be challenging to assess using standard scientific methods. Furthermore, D. deltoides forms aggregated distributions (Murray-Jones 1999; McKenzie and Montgomery 2002) or ‘nests’ (H. Smith, Traditional Owner Ngambaa, pers. obs.) and is driven along beaches by prevailing winds, swell and seas and actively ‘swash ride’ to move up and down the beach slope with the tides (Ellers 1995; McLachlan et al. 1995). Several studies have rapidly assessed presence or absence and relative abundance of D. deltoides on the eastern coast of Australia; however, these have been limited to south of Dhungatti (Murray-Jones 1999; McKenzie and Montgomery 2002; Johnson 2022) and north of Yaegl (Schnierer and Egan 2016) on the eastern coast of New South Wales (NSW) (Fig. 1). Except for the documentation of presence or absence in macrofauna assessments of sandy shores (Hacking 1998, 2007), there has been no comprehensive assessment of D. deltoides within Yaegl, Gumbaynggirr, Ngambaa or Dhungatti, which are prominent Nations on the Mid North Coast of NSW (Fig. 1). Donax deltoides is called ‘garlaany’ in the local Ngambaa language and is considered a cultural keystone species in this region (Morelli 2015).
Beach locations used in the consultation strategy (North Beach, Mylestom) for co-designed method development for sampling Donax deltoides, and the application to the spatial and temporal assessment of populations in Stuarts Point Beach, Ngambaa, NSW (note: Nation borders are approximate).
The current stock status of D. deltoides in NSW is viewed as sustainable with a stabilised annual catch of ~116 Mg (Johnson 2022). However, there are limited data on cultural take and ecosystem services (Murray-Jones and Steffe 2000; Owner and Rohweder 2003; Schnierer 2011). The current total allowable catch for commercial fishers is 116 Mg year−1, with a 40-kg daily catch limit. A minimum legal length (MLL) of 45-mm shell length (SL) is also regulated for commercial fishers. Recreational fishers are restricted to 50 individuals per day bag limit to be used as bait only (Department of Primary Industries 2015, 2023). However, recreational catches for the Mid North Coast are much higher than in other fishing zones and account for an estimated 60% of the total NSW recreational catch (87,760 counts ± 31,272 s.e.) (West et al. 2016; Johnson 2022). Anecdotal evidence from Aboriginal, recreational and commercial fishing sectors have indicated localised declines in D. deltoides populations. Specifically, observations from many Traditional Owners and recreational fishers in northern NSW suggest that many intermediate dissipative beaches that historically contained a large biomass of D. deltoides now have depleted populations (J. Fredericks, pers. comm.). This decline is mainly attributed to anthropogenic disturbances such as overharvesting, vehicle access and sand mining (Schlacher et al. 2008; Gray 2016a). Further studies are required to assess the spatial and temporal trends in counts (as a measure of relative abundance) and size–frequency distributions of D. deltoides in relation to environmental parameters and anthropogenic disturbance. Because D. deltoides is a cultural keystone species and an aggregated sandy shore mollusc that is difficult to assess on beaches with low abundance, it is a suitable target species for the application of IEK for population assessment on the Mid North Coast of NSW. Here, we use an integrated fisheries research project that has been co-designed by Traditional Owners, to include IEK, along with participation of Traditional Owners in the data collection to assess spatial and temporal patterns in D. deltoides counts and size–frequency distributions on Stuarts Point Beach. This study provides validation of a rigorous co-designed methodological approach for broader application along the NSW coast.
Materials and methods
Indigenous ethics approval and consultation strategy
This project was undertaken using codes of conduct and ethical guidelines for research with First Nations communities as outlined in Australian Institute of Aboriginal and Torres Strait Islander Studies (2020) and National Health and Medical Research Council (2018). These guidelines were used in developing the consultation strategy, which included Traditional Owner participation in most stages of the research project (Box 1). Pre-consultation was undertaken with Traditional Owners from Nambucca Heads Local Aboriginal Land Council Board and community members. This included 60 face-to-face interviews on the use of D. deltoides in southern Gumbaynggirr and Ngambaa (J. Fredericks, unpubl. data, 2018). Results from the surveys indicated that populations of D. deltoides in southern Gumbaynggirr are smaller, have lower abundance and are harder to find. Anecdotal evidence also indicated that clan-based local beaches, which had been historically fished, are now being depleted of D. deltoides.
| Box 1.Stages of research and guiding principles strategies for Traditional Owner engagement. |
Following the pre-consultation surveys, further consultation with Board and Community members of Galambila, Ngambaa and Yaegl Nations facilitated the development of a funding proposal to the Aboriginal Fishing Trust Committee. This project was supported and, subsequently, a sampling design for assessing D. deltoides populations was developed in consultation with Traditional Owners of Gumbaynggirr, Ngambaa and Dhungatti, to combine the use of IEK and CSK. The co-designed methodology was then applied to a spatial and temporal assessment of D. deltoides on a nominated beach.
Integration of Indigenous knowledge
The sampling design and method development for the spatial and temporal assessment of D. deltoides was developed via a series of consultations undertaken by the investigators with the Coffs Harbour Local Aboriginal Land Council (CHLALC), Nambucca Heads Local Aboriginal Land Council (NHLALC), Yaegl Native Title Holders, Ngambaa Traditional Custodians, Gumma Indigenous Protected Area (IPA) Sea Country rangers, the Gumma IPA Operations manager, Gumbaynggirr Elders, Board members and the wider community.
Under request from Gumbaynggirr Traditional Owners, cultural protocols were confidential and used discretely by rangers during the sampling procedure. To support the cultural integrity of the project, a Sea Country ranger or steward accompanied the principal investigator while undertaking the assessments. This would ensure that culturally sensitive sites were not disturbed, as well as supporting the incorporation of confidential IEK into the ongoing sampling strategy.
Consultation for the sampling-method development occurred at one of the more extensive beaches in Gumbaynggirr, North Beach (Mylestom), a historical, cultural harvesting site for D. deltoides (Fig. 1). North Beach is located in Gumbaynggirr Waajar (Country) at 30.464769°S, 153.047250°E. The beach measures 8.75 km in length, borders Bundageree Creek to the north and Bellinger River to the south and is characterised as an intermediate dissipative beach (Short 2006). Three beach-access points were deemed suitable for starting points for testing the co-designed sampling methods: one north at Tuckers Rock, one mid-beach north of the Mylestom Surf Life Saving Club (SLSC), and one south of the Mylestom SLSC. The southern access point was used for the purpose of the consultation and co-development of the sampling design.
IEK-informed sample-unit development
The IEK-informed sample-unit was developed using IEK and traditional harvesting methods and was quantified using CSK. Once a part of the beach was assessed as having potential for harbouring D. deltoides, on the basis of IEK of topography, including slope, sand-grain size, and proximity to swash area, a test hole was dug in the area. Test holes were made by placing bare feet ~5 cm apart and moving both feet left to right while applying pressure to the balls of the feet. The sand was then excavated using hands and finger dredging to search for individual D. deltoides according to traditional harvesting methods. If an individual was found, a series of other test holes were excavated near the first test hole, moving perpendicular and parallel along the beach. If one or more D. deltoides individuals were found in two or more test holes, further sample holes were then randomly dug by participants until adequate sampling effort had resulted in a sizeable number for the area.
After further discussion with the Traditional Owners, it was decided that a 10- × 10-m grid would be used for the ongoing assessment. Therefore, the final IEK- and CSK-designed sampling approach used a 10- × 10-m sample grid, with 25 holes excavated. The dimensions of the test holes and total area of beach sampled at each site during the consultation phase were measured with a tape measure for comparison with recommended CSK sampling methods and to help validate the co-designed sampling methods.
To further adhere to CSK methods, the volume of excavated sand from each hole was quantified using the formula for a cylinder (Eqn 1) to ensure that an adequate percentage of each sample area was covered by the IEK-informed sample unit.
Study location
After further consultation with the Traditional Owners, Stuarts Point Beach, located within Ngambaa Country (Fig. 1), was selected to test the IEK-informed methodology for population assessment of D. deltoides, by monitoring the spatial and temporal variation in D. deltoides counts and size–frequency distributions. Stuarts Point Beach has a history of commercial, recreational and cultural fishing, along with recent anecdotal declines in population as observed by the Traditional Owners and commercial and recreational fishers.
Stuarts Point Beach, located towards the southern end of Ngambaa, is ~8.7 km in length and extends from Grassy Head Headland (30.79927°S, 152.9694486°E) to the Macleay River Headwaters (30.869834°S, 153.019992°E). Ngambaa is a gatekeeper Nation, extending the area in between Gumbaynggirr and Dhungatti, from the Macleay River to Nambucca River (Tindale 1974; Kijas 2005) and, therefore, is home to descendants of both Gumbaynggirr and Dhungatti. Gumbaynggirr and Dhungatti are two prominent Aboriginal Nations on the Mid North Coast of NSW (Fig. 1). Gumbaynggirr extends an estimated 6000 km2, bordering the Clarence River to the north, Nambucca River to the south and the Great Dividing Range to the west. Dhungatti extends ~5100 km2 from the Macleay River in the north, Hasting River in the south and the Great Dividing Range to the west. Clybucca midden is located within Ngambaa and holds evidence of past meeting grounds among Nations. It is the largest midden in the Southern Hemisphere, ~14 km in length, and dates back at least 4000 years (Sullivan 1985; Knuckey 1999).
Data collection
For the spatial and temporal assessment at Stuarts Point Beach, surveys were undertaken twice monthly, on two random days over 13 months from November 2021 to November 2022. The beach was divided into three locations (north, middle and south) to enable comparisons along the beach, because significant variation in D. deltoides populations is known to occur according to the position on the beach, on the basis of previous CSK studies (James and Fairweather 1995; Hacking 1998; Gorman et al. 2010) and IEK.
On each sampling occasion, two sites were sampled within each of the three locations. Once a site was chosen using IEK indicators, a 10- × 10-m grid was set up using markers on the sand in the swash zone and a hole was randomly excavated in every 2- × 2-m grid section, as per the results of the sample-unit development. Holes were sampled using bare feet, hands and finger dredging as per the traditional harvesting methods. For each excavated hole, sand was also sieved with a 5-mm mesh hand-held net. After completing each hole, all D. deltoides individuals were counted and measured for maximum shell length (SL) by using vernier callipers to the nearest 1 mm. All individuals were then returned to the sample site in the swash zone and observed until they had re-buried into the substrate. If stranded, they were assisted into deeper water.
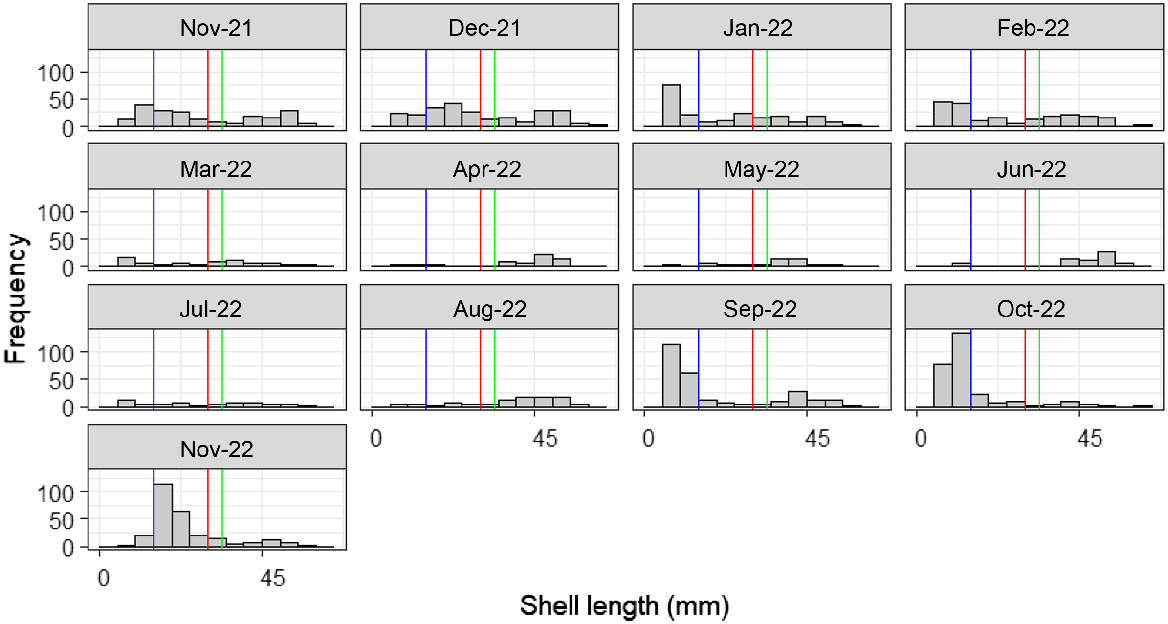
The counts and length–frequency data from replicate holes were collated for each site and then aggregated into three size classes. The adult size class (SK ≥ 30 mm) was defined on the basis of published information for the first size recorded for gametes (SL 27 mm) and size at sexual maturity for 50% of the population (SAM50, SL ≥ 37 mm), as estimated by Murray-Jones (1999). Visual analysis of length–frequency data from the current study indicated a bimodal distribution at the 30-mm junction. The juvenile size class (SL 15–30 mm) was defined by the adult and recruit cut-off points. The recruit size-class cut-off (SL ≤ 15 mm) was based on size–frequency information gathered within this study (refer to Results section and Fig. 2).
Length–frequency histograms of Donax deltoides sampled over a 13-month period from November 2021 to November 2022 at Stuarts Point Beach (n = 2072). Shell length cut-off points for size classes are indicated: recruits (0–15 mm; blue line), adults (>30 mm; red line), size at 50% sexual maturity (SAM50; green line), and minimum legal length (MLL) for commercial harvesting (grey line at 45).
At the start of each sampling occasion, information on a range of environmental variables was also recorded, including sand-grain size, slope, swell, wave frequency, cloud cover and wind direction (Table 1). At each site, a GPS reading of longitude and latitude was recorded, along with information on the IEK indicators and beach geomorphology. To assess sand-grain size, three samples of sand were taken from each site with a 15-cm-long 5-cm diameter polystyrene pipe. Samples were oven dried and sieved to obtain sand-grain sizes on the basis of Wentworth (1922, cited in Cheetnam et al. 2008). Slope gradient (%) was calculated using a theodolite, staff and transect tape at each site. The height of theodolite, length of transect tape and height of staff were used to calculate the angle of beach slope, then grouped into categories (Table 1). A marked protractor was used as a visual cue to confirm categories at each site. Sand penetration was determined with a steel rod (2-m length, 0.5-cm diameter), which was dropped from 50-cm height, at highest point in the swash zone, and penetration depth was measured to the nearest millimetre. Three measurements were taken per site, two steps apart, parallel to the shore, then averaged for each site. Wave frequency was assessed from counts of the number of crest waves to pass a fixed point over 10 s, with an average of three counts per site. Tidal information including maximum tide height, minimum tide height, tide difference and moon illumination (Table 1) were later accessed from online meteorological sources (NSW tide tables 2020–2021, 2021–2022, 2022–2023).
| Variable | Variable description | Variable type | |||
|---|---|---|---|---|---|
| Morphological | |||||
| Sand-grain size | Category | Size range (μm) | Wentworth size class | Categorical | |
| 1. Silt | <63 | Silt | |||
| 2. Very fine | 63–125 | Very fine sand | |||
| 3. Fine | 126–250 | Fine sand | |||
| 4. Medium | 250–500 | Medium sand | |||
| 5. Coarse | 501–1000 | Coarse sand | |||
| 6. Very coarse | 1001–2000 | Very coarse sand | |||
| Slope | Category | Gradient (%) | Degree equivalent (°) | ||
| 1. Flat | 0–3 | <1 | Categorical | ||
| 2. Very slight incline | 3–8 | 1–5 | |||
| 3. Slight incline | 9–15 | 5–8.5 | |||
| 4. Slightly steep | 16–25 | 8.5–16.5 | |||
| 5. Steep | 26–45 | 16.5–24 | |||
| 6. Very steep | >45 | >24 | |||
| Sand penetration | Depth into sand (cm) | Continuous | |||
| Tidal | |||||
| Tide phase | Category | Time from low tide (min) | Categorical | ||
| 1. Very low | 0–90 | ||||
| 2. Low | 91–180 | ||||
| 3. Low mid | 180–270 | ||||
| 4. High mid | 271–360 | ||||
| 5. High | 360–450 | ||||
| 6. Very high | 451–540 | ||||
| Maximum tide height | Maximum tide height (cm) | Continuous | |||
| Minimum tide height | Minimum tide height (cm) | Continuous | |||
| Meteorological | |||||
| Wave frequency | Number of waves | Continuous | |||
Data analysis
Statistical analysis was performed using the R (ver. 4.2.3, R Foundation for Statistical Computing, Vienna, Austria, see https://www.r-project.org/) programming environment and the lme4 package (ver. 1.1-35.5, see https://CRAN.R-project.org/package=lme4; Bates et al. 2015). Statistical analyses were undertaken to compare spatial and temporal variation in D. deltoides counts and size–frequency distributions and relationships with environmental variables by using the following four response variables: (1) counts of adults (SL ≥ 30 mm); (2) counts of juveniles (SL 15–30 mm); (3) counts of recruits (SL < 15 mm); and (4) mean size (SL, mm). Counts (relative abundance estimates), which were recorded per hole excavated, were pooled to enable aggregation into size classes for analysis and are presented as means ± s.e. counts per site throughout the paper.
For the temporal analysis of D. deltoides counts and size at Stuarts Point Beach, the base model included day (two levels, random), nested in month (13 levels, fixed) and site (two levels, random) nested in location (three levels, fixed) and their interactions to form a generalised linear mixed-effects model (GLMM). However, owing to total count data being aggregated to counts per site, variation explained by the random effects of days and site in the models was reduced to insignificant amounts and was removed, such that the final models were generalised linear models (GLMs). Count data were modelled using a Poisson distribution and mean size (length) data were modelled using a Gaussian distribution. Pairwise comparisons were made using Tukey’s honest significant difference (HSD) test with the Kenward–Roger degrees of freedom method (Kenward and Roger 1997).
First, collinearity between variables was assessed using generalised variation inflation factor (GVIF) and variables were removed starting from the highest GVIF score, until all variables were <5 (Supplementary Tables S1–S4). Then, a step-up approach was used to determine which environmental variables significantly improved the base models for count and mean length data, analysed using GLM. Any continuous variables were first fitted with GLM by using the lowest Akaike information criterion (AIC) and Chi-Square values, with P < 0.05.
Results
Comparison of IEK-informed and CSK methods
The IEK-informed method differed from the traditional conventional scientific approach through the use of bare feet and hands to excavate holes and the use of IEK indicators to target sites. CSK methods typically use a shovel and quadrat or corer. The IEK indicators applied to the selection of sample sites in our co-designed study included active observation of, but were not limited to, visible lumps in the sand, D. deltoides swash riding (loose individuals in shore waves) or feeling D. deltoides underfoot. On occasions where these indicators were not present, sites were chosen on the basis of beach topography, such as beach slope and sand appearance (Table 1). Test holes were used to confirm the presence of D. deltoides before sites were selected for full sampling. In comparison, CSK approaches typically apply random or fixed sites along beaches and all sites are sampled regardless of evidence for the presence of D. deltoides. Once a suitable site for sampling was selected, the IEK-informed design was standardised in a way that is similar to CSK methods. Quantification of the IEK-informed sample unit provided hole diameters of 35–45 cm and depths of 10–20 cm, which were similar in dimension to the recommended quadrat size (0.33 × 0.33 m) and depth (0.1 m) for efficient sampling of D. deltoides tested by CSK (James and Fairweather 1995). At the end of the consultation, ~25 holes between 35 and 45 cm in diameter were excavated using feet and hands to ~10–20 cm. The total area of beach that was sampled measured between 10 and 15 m in length and width. The final sample-hole allocation design resembled CSK methods, with a hole excavated randomly within every 2 × 2 m of the 10- × 10-m grid. This is equivalent to a replicated block design with a total of 25 holes per 100-m2 sampled at each site. To reduce sampling bias, the holes were randomly placed within each 2-m2 quadrat. CSK methods more commonly sample quadrats every 2 m along a random or haphazard 50-m transect, but both methods result in 25 replicates per 100 m2.
IEK-informed sampling at Stuart’s Point Beach
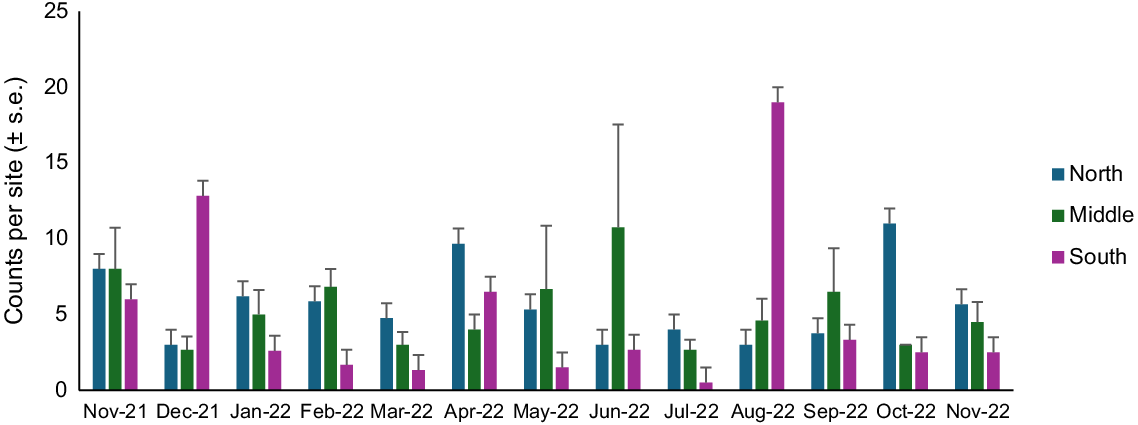
The IEK-informed method was successful in detecting spatial and temporal differences in counts and size–frequency distributions and correlations with environmental variables. During the 13 monthly surveys at Stuarts Point Beach, 2072 D. deltoides individuals were counted, from 3750 individual holes across 156 sites, providing an overall average of 13.75 counts per site. Higher overall counts were recorded from November 2021 to February 2022 and September–November 2022 compared with March–August 2022. The highest average counts across the months sampled were recorded in October and November (21.75 ± 2.25 counts per site) and lowest in July 2022 (4.5 ± 0.75 counts per site) (Fig. 2).
Size–frequency distributions
Application of the co-designed sampling method was successful for recovering sufficient counts of D. deltoides, across a range of size classes, for assessment of temporal changes in size–frequency distributions (Fig. 2). Total counts across size classes for the entire sampling period were higher for recruits (755) and juveniles (561) combined than for adults (756). Counts for those larger than SAM50 (625) accounted for ~30% of the population; counts of MLL (335) was low, only ~15% overall (Fig. 2). A visual analysis of size–frequency information for Stuarts Point Beach from November 2021 to 2022 showed a bimodal distribution for November 2021 through to February 2022 (Fig. 2). Overall, counts of adults larger than SAM50 and the MLL declined over the sampling period; however, recruitment (5–15 mm) was high from September through November 2022. A recruitment cohort emerged in January 2022 and again in September 2022, showing highest counts from 6 to 10 mm and a cohort moving through November 2022 that were up to 15 mm SL (Fig. 2).
Temporal and spatial patterns of Donax deltoides at Stuarts Point Beach
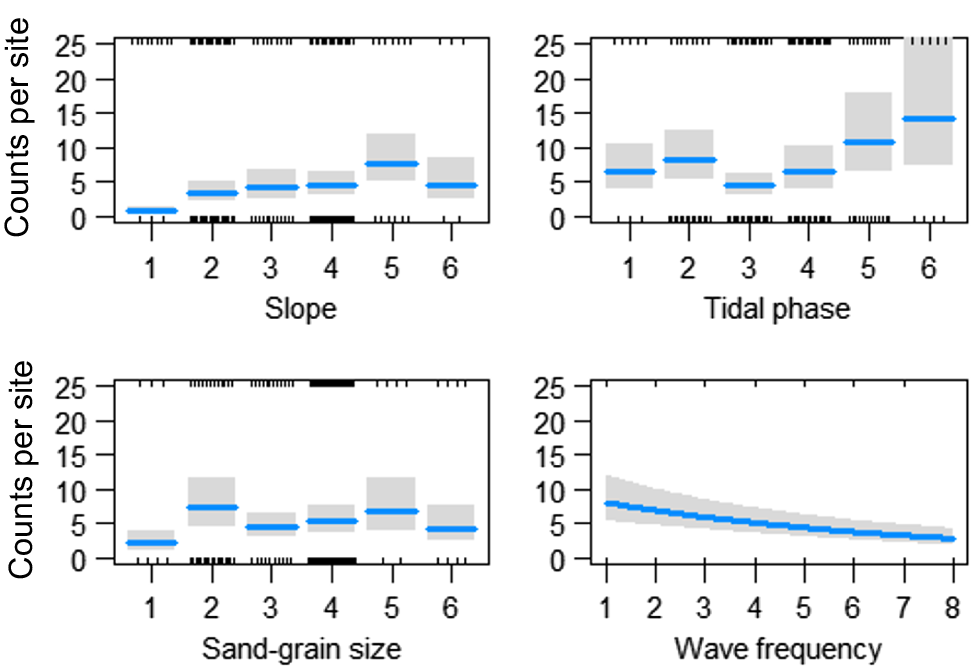
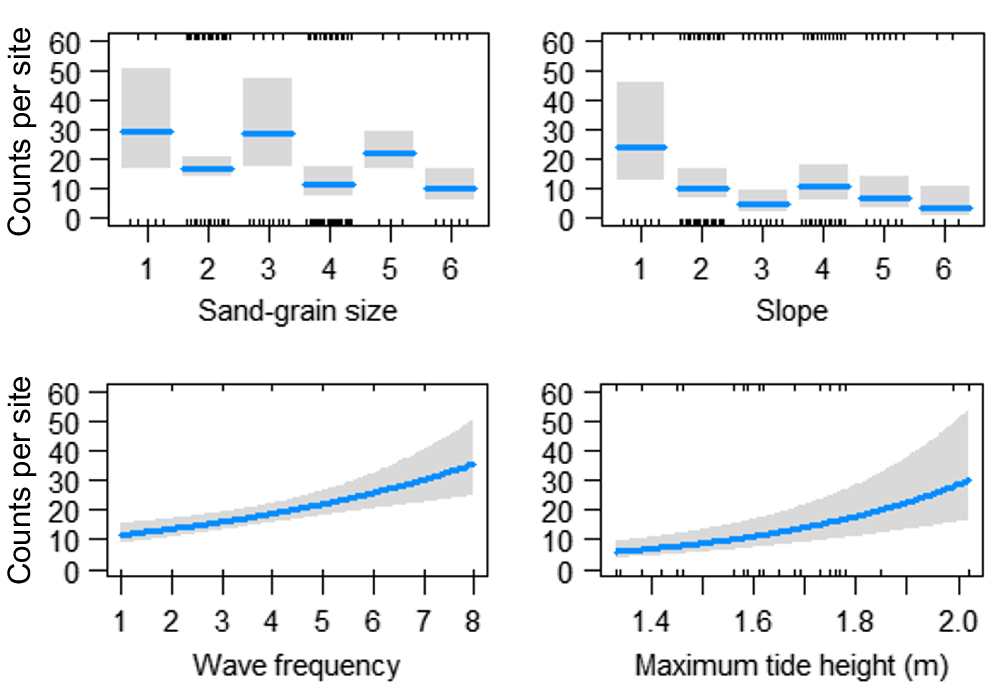
For counts of adult D. deltoides (SL > 30 mm), there was a significant interaction between month and location (χ2 = 215, P < 0.001, d.f. = 24). The highest counts of adults were recorded in August 2022 and December 2021 in the northern location, in June 2022 in the middle of the beach and in August 2022 in the southern beach location (Table 2, Fig. 3). The addition of environmental variables to the model indicated that a significant amount of the variation in the counts of adult D. deltoides at Stuarts Point Beach could be explained by morphological (sand-grain size, beach slope), tidal (tide phase, minimum tide height) and meteorological (wave frequency) variables. However, the most parsimonious model (R2 = 0.67, d.f. = 74) included the effects of spatial and temporal variables plus beach-state values, including slope, sand-grain size, tide phase and wave frequency (Table 2). A visual analysis of environmental variables shows an increase in counts of adult D. deltoides in response to increases in slope, at a higher tide phase, medium sand-grain size and low wave frequency (Fig. 4).
| Variable | d.f. | Deviance | Residual d.f. | Residual deviance | P(>χ 2) | AIC | |
|---|---|---|---|---|---|---|---|
| Null | 667.7 | 128 | |||||
| Month | 12 | 71.1 | 116 | 596.6 | <0.05* | 1036.2 | |
| Location | 2 | 0.9 | 114 | 595.6 | 0.618 | 1039.3 | |
| Month × location | 24 | 215.0 | 90 | 380.6 | <0.001*** | 868.2 | |
| Minimum tide | 1 | 6.6 | 89 | 373.9 | <0.010** | 863.5 | |
| Wave frequency | 1 | 22.4 | 89 | 358.1 | <0.001*** | 870.1 | |
| Sand-grain size | 5 | 31.8 | 85 | 348.7 | <0.001*** | 846.3 | |
| Tidal phase | 5 | 36.7 | 85 | 343.8 | <0.001*** | 841.5 | |
| Slope | 5 | 100.5 | 85 | 280.1 | <0.001*** | 777.7 | |
| +Tidal phase | 5 | 19.2 | 80 | 260.8 | 0.002** | 768.4 | |
| +Sand-grain size | 5 | 16.3 | 75 | 244.5 | 0.006** | 762.1 | |
| +Wave frequency | 1 | 25.1 | 74 | 219.3 | <0.001*** | 739.0 |
d.f., degrees of freedom; P, probability of χ2 value significant at: *, P < 0.05; **, P < 0.01; ***, P < 0.001. AIC, Akaike information criterion.
Counts (mean ± s.e. counts per site) of adult (SL > 30 mm) Donax deltoides individuals sampled over 13 months (November 2021–November 2022) and three locations (north, middle, south) at Stuarts Point Beach, Ngambaa, NSW.
Fitted values (with 95% confidence intervals indicated by shading) for significant relationships identified in the GLM analyses between counts of adult (SL > 30 mm) Donax deltoides and environmental variables: slope (1, flat; 2, very slight incline; 3, slight incline; 4, slightly steep; 5, steep; 6, very steep); tide phase (1, very low; 2, low; 3, low middle; 4, high middle; 5, high; 6, very high); sand-grain size (1, silt; 2, very fine; 3, fine; 4, medium; 5, coarse; 6, very course); sampled across 13 months (November 2021–November 2022) and three locations (north, middle, south) at Stuarts Point Beach, Ngambaa, NSW.
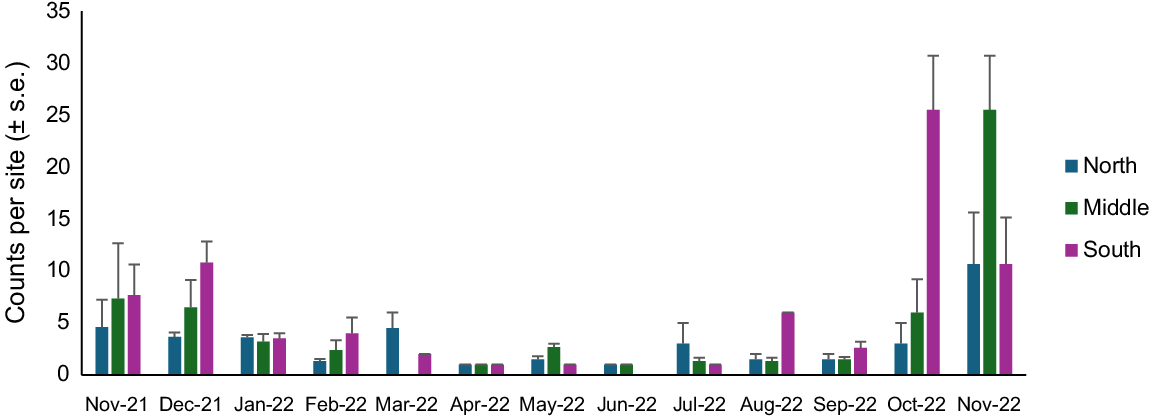
Counts of juveniles varied at different locations across the beach in different months and were highest in October and November 2022. There was also a peak in counts in November and December 2021, indicating cohorts moving through during the austral summer (Fig. 5). There was a significant interaction between month and location on the beach for juvenile counts (Table 3). Low counts occurred from January 2022 through to September 2022, with the northern location having the lowest counts of D. deltoides across all months apart from March 2022 and July 2022. Highest counts were in the southern end of beach in October 2022 and the middle of the beach in November 2022.
Counts (mean ± s.e. counts per site) of juvenile (SL 15–30 mm) Donax deltoides individuals sampled over 13 months (November 2021–November 2022) and three locations (north, middle, south) at Stuarts Point Beach, Ngambaa, NSW.
| Variable | d.f. | Deviance | Residual d.f. | Residual deviance | P(>χ 2) | AIC | |
|---|---|---|---|---|---|---|---|
| Null | 653.1 | 103 | |||||
| Month | 12 | 354.3 | 91 | 298.7 | <0.05* | 635.6 | |
| Location | 2 | 25.4 | 89 | 273.3 | <0.001*** | 614.2 | |
| Month × location | 24 | 76.5 | 65 | 196.7 | <0.001*** | 577.6 | |
| Sand penetration | 1 | 180.3 | 64 | 180.3 | <0.001*** | 563.2 | |
| Slope | 5 | 23.4 | 60 | 173.2 | <0.001*** | 564.1 | |
| Sand-grain size | 5 | 24.0 | 60 | 172.6 | <0.001*** | 563.5 | |
| +Slope | 1 | 18.1 | 59 | 154.4 | <0.003** | 555.3 | |
| +Sand penetration | 1 | 18.3 | 58 | 136.1 | <0.001*** | 539.0 |
d.f., degrees of freedom; P, probability of χ2 value significant at: *, P < 0.05; **, P < 0.01; ***, P < 0.001. AIC, Akaike information criterion.
Variation in counts of juvenile D. deltoides at Stuarts Point Beach was influenced by sand-grain size, slope, temperature, moon illumination, swell, lunar day, wind direction and cloud cover. However, when combined with the base model, sand-grain size and tide phase were the only variables included in the final model and accounted for 79% of the variation (R2 = 0.79, d.f. = 58) (Table 3). Lowest counts of juveniles were found in coarse sand (sand grain size = 6) and low tides (tide phase 1–2), whereas higher counts were found in the fine to medium sand (sand-grain size 1–3) and the middle of the tides (tide phase 3–4) (Fig. 6).
Fitted values (with 95% confidence intervals indicated by shading) for significant relationships identified by the GLM analyses between counts of juvenile (SL 15–30 mm) Donax deltoides and environmental variables: sand-grain size (1, silt; 2, very fine; 3, fine; 4, medium; 5, coarse; 6, very course); and slope (1, flat; 2, very slight incline; 3, slight incline; 4, slightly steep; 5, steep; 6, very steep); sampled across 13 months (November 2021–November 2022) and three locations (north, middle, south) at Stuarts Point Beach, Ngambaa, NSW.
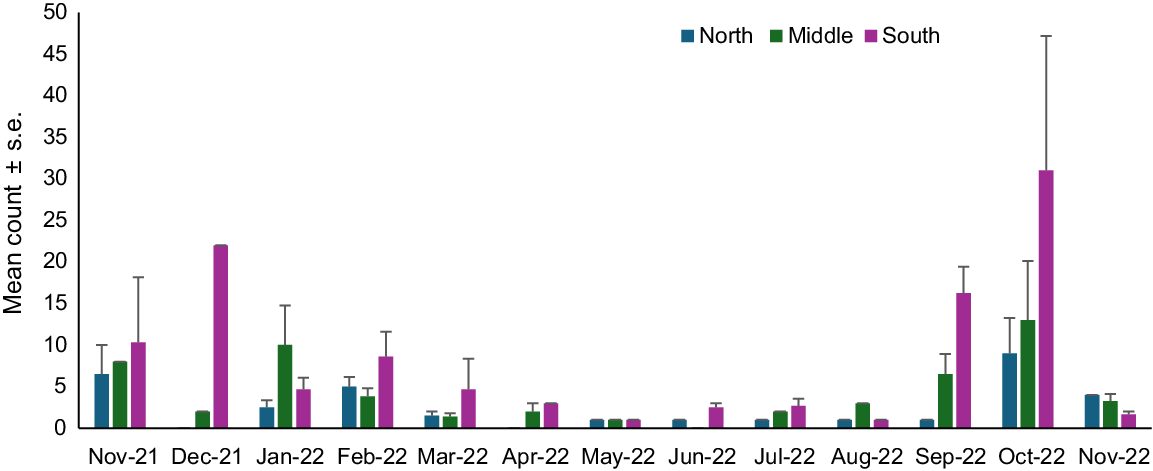
A visual assessment of spatial and temporal variation shows a peak in recruitment in austral summer, November and December 2021, and austral spring, September and October 2022, with low overall counts, including and extending either side of the austral winter (April–August 2022) (Fig. 7). A spatial and temporal assessment of recruit Donax deltoides at Stuarts Point Beach showed a significant interaction between month and location (Table 4). Counts of recruits were highest in the southern beach location across all months apart from January, when recruit numbers were higher in the middle of the beach. The northern beach zone had lowest counts of recruits across all months (Fig. 7).
Counts (mean ± s.e. counts per site) of recruit (SL < 15 mm) Donax deltoides sampled over 13 months (November 2021–November 2022) and three locations (north, middle, south) at Stuarts Point Beach, Ngambaa, NSW.
| Variable | d.f. | Deviance | Residual d.f. | Residual deviance | P(>χ2) | AIC | |
|---|---|---|---|---|---|---|---|
| Null | 802.3 | 100 | 802.3 | ||||
| Month | 12 | 292.5 | 88 | 509.7 | <0.050* | 845.6 | |
| Location | 2 | 66.5 | 86 | 443.1 | <0.001*** | 783.0 | |
| Month × location | 24 | 149.1 | 62 | 294.0 | <0.001*** | 673.8 | |
| Maximum tide | 1 | 12.3 | 61 | 281.6 | <0.001*** | 663.5 | |
| Wave frequency | 1 | 15.7 | 61 | 278.3 | <0.001*** | 660.1 | |
| Minimum tide | 1 | 25.5 | 61 | 268.4 | <0.001*** | 650.2 | |
| Slope | 5 | 33.3 | 57 | 260.6 | <0.001*** | 650.5 | |
| Sand-grain size | 5 | 33.8 | 57 | 260.1 | <0.001*** | 650.0 | |
| +Slope | 5 | 22.6 | 52 | 237.5 | <0.001*** | 637.3 | |
| +Maximum tide | 1 | 23.5 | 51 | 213.9 | <0.001*** | 615.8 | |
| +Wave frequency | 1 | 15.6 | 50 | 168.3 | <0.001*** | 578.1 |
d.f., degrees of freedom; P, probability of χ2 value significant at: *, P < 0.05; **, P < 0.01; ***, P < 0.001. AIC, Akaike information criterion.
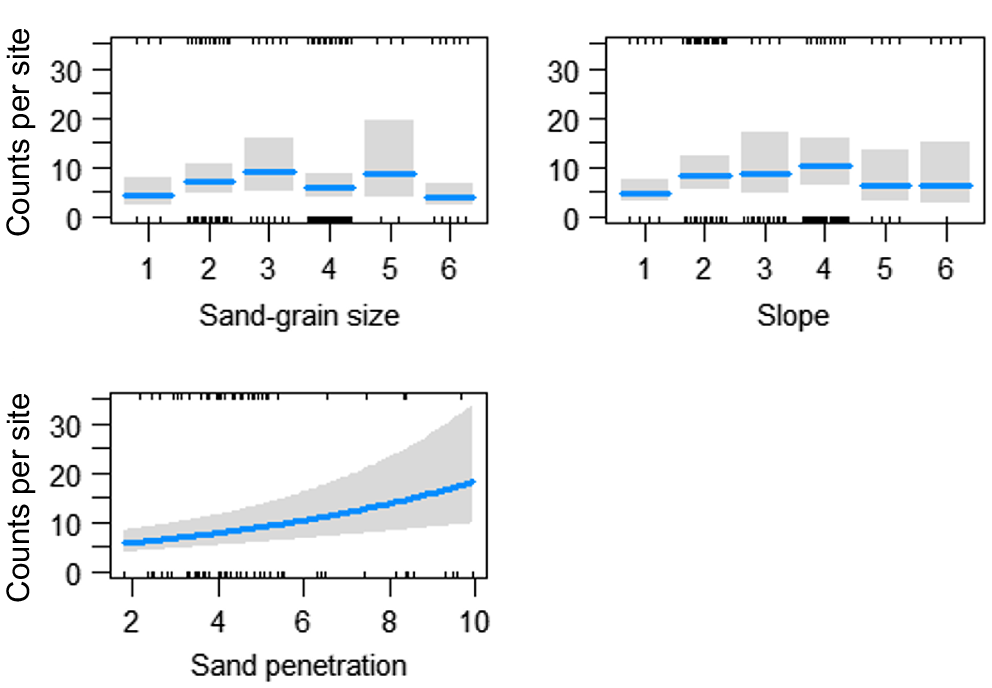
Analysis of environmental variables showed that the variation in counts was influenced significantly by sand-grain size, slope, wave frequency and maximum tide height. The final model, which explained 79% of the variation in the counts of recruits, was attributed to spatial and temporal variation plus slope, sand-grain size, maximum tide height and wave frequency (R2 = 0.79, d.f. = 50) (Table 4). Higher counts of recruit D. deltoides were found on flatter slopes (slope 1–3), at finer sand-grain sizes (1–3) and increased with an increasing maximum tide height (Fig. 8).
Fitted values (with 95% confidence intervals indicated by shading) for the significant relationships identified in the GLM analyses between counts of recruit (SL < 15 mm) Donax deltoides and environmental variables: sand-grain size (1, silt; 2, very fine; 3, fine; 4, medium; 5, coarse; 6, very course); slope (1, flat; 2, very slight incline; 3, slight incline; 4, slightly steep; 5, steep; 6, very steep); wave frequency and maximum tide height (cm); sampled across 13 months (November 2021–November 2022) and three locations (north, middle, south) at Stuarts Point Beach, Ngambaa, NSW.
Temporal patterns in Donax deltoides shell length at Stuart Point Beach
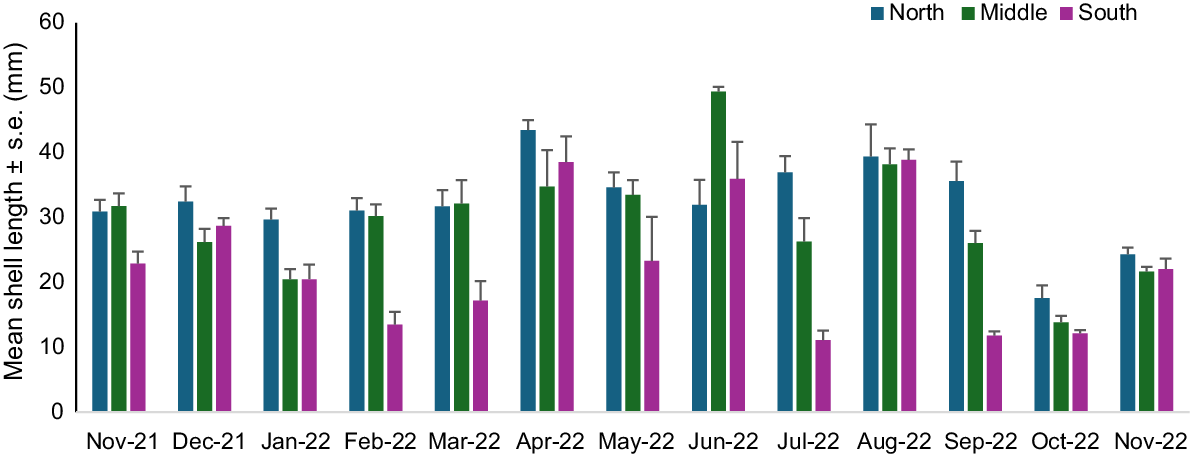
Overall, the mean shell length for D. deltoides at Stuarts Point Beach across the entire sampling period was 25.14 mm (±s.e. 0.35), with a minimum length measured being 6 mm (±1 mm) and a maximum being 61 mm (±1 mm). Higher overall mean lengths were found from April to August 2022 and the lowest in October and November 2022 (Fig. 9). Higher mean lengths were found in the northern end of the beach over most months, apart from June 2022, where the highest overall mean length was recorded in the middle of the beach. Low overall mean lengths were recorded for the southern end of Stuarts Point Beach, with October having low counts across all locations (Fig. 9). Statistical analysis of mean shell length identified significant differences in months and locations and there was an interaction between these factors (Table 5). The decrease in mean shell length coincided with the influx of recruits and juveniles during the austral spring and summer months.
Mean shell length (mm, ±s.e.) of Donax deltoides sampled over 13 months (November 2021–November 2022) and three locations (north, middle, south) at Stuarts Point Beach, Ngambaa, NSW.
| Variable | d.f. | Deviance | Residual d.f. | Residual deviance | F | P(>F) | |
|---|---|---|---|---|---|---|---|
| Null | 156 | 990.0 | |||||
| Month | 12 | 256.3 | 144 | 733.6 | 21.3 | <0.001*** | |
| Location | 2 | 98.1 | 142 | 635.5 | 49.0 | <0.001*** | |
| Slope | 5 | 23.9 | 137 | 611.5 | 4.7 | <0.001*** | |
| Sand-grain size | 5 | 20.0 | 132 | 591.5 | 4.0 | <0.001** | |
| Sand penetration | 1 | 29.7 | 131 | 561.7 | 29.7 | <0.001*** | |
| Wave frequency | 1 | 4.1 | 130 | 557.6 | 4.1 | <0.050* | |
| Month × location | 24 | 130.2 | 106 | 427.4 | 5.4 | <0.001*** |
d.f., degrees of freedom; P, probability of F-value significant at: *, P < 0.05; **, P < 0.01; ***, P < 0.001.
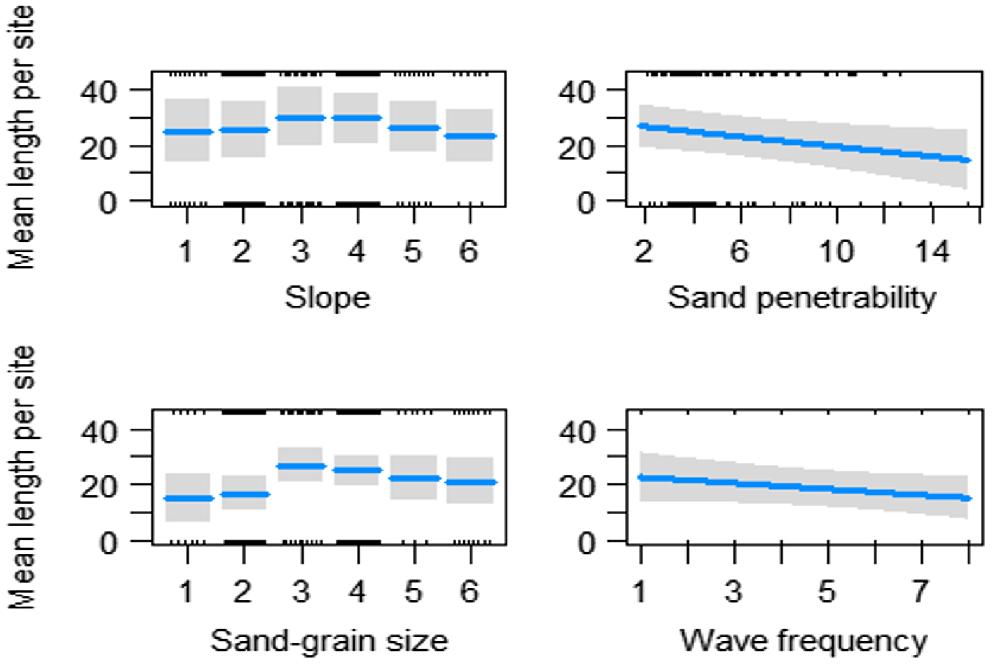
Analysis of environmental variables and mean shell length showed that significant variation was explained by spatial and temporal variation in the base model and slope, sand-grain size, sand penetrability and wave frequency. Smaller individuals tended to be found on flatter slopes and smaller sand-grain sizes (Fig. 10). A greater number of larger individuals were found on medium slopes and medium to course sand-grain sizes (Table 1, Fig. 10).
Fitted values (with 95% confidence intervals indicated by shading) for significant relationships identified from the GLM analyses between mean shell length (mm) of Donax deltoides and environmental variables: slope (1, flat; 2, very slight incline; 3, slight incline; 4, slightly steep; 5, steep; 6, very steep); sand-grain size (1, silt; 2, very fine; 3, fine; 4, medium; 5, coarse; 6, very course); sand penetrability (cm); and wave frequency; sampled across 13 months (November 2021–November 2022) and three locations (north, mid, south) at Stuarts Point Beach, Ngambaa, NSW.
Discussion
Collaboration with Traditional Owners enabled the design, optimisation and implementation of a sampling strategy that differed from traditional CSK methods and was successful in assessing an important cultural and fisheries resource on sandy beaches. Incorporating Traditional Owner co-design, site selection and participation in the execution of field sampling, we successfully detected significant differences in the counts and mean size of D. deltoides (garlaany, pipi) among months and locations within Stuarts Point Beach in Gumbaynggirr Country. We also identified a range of environmental variables that significantly explained size-specific variation in D. deltoides abundance. Further to these results, a co-designed framework provided a platform for two-way information and skill sharing, increased knowledge of the target species and habitat and fostered relationship development for ongoing co-management, monitoring and stewardship.
The IEK-informed method differed from the CSK-informed method through the use of hands, feet and IEK indicators synonymous with traditional harvesting methods. In comparison, methods using CSK sampling on sandy shores often utilise shovels, transects and quadrats at fixed or random sites along the beach. Being aligned to cultural practice, the IEK-informed approach is more likely to engage Traditional Owners in participation in co-designed research and empower communities with a tool suitable for adaptive management and longer-term monitoring of cultural resources on Sea Country. By comparison, the need for more equipment, and requirements for formal randomisation methods or measuring fixed locations by using the CSK approach, could deter ongoing participation. Apart from engaging with the Traditional Owners in the data-collection process, additional benefits arise from the use of IEK indicators to inform site selection. Although not specifically tested in this study, IEK could reduce sample time and potentially increase precision and accuracy, especially on beaches of low abundance where populations may be missed altogether when using conventional random site selection.
The identification of sites by using IEK indicators differs substantially from the CSK sampling, which uses random or fixed sites along beaches (e.g. fixed for broad-scale and random for fine-scale variability; Ferguson et al. 2021). The main difference in the co-designed sampling strategy is that sites are first selected using IEK indicators and then test holes are sampled until D. deltoides individuals are found. This differs from the ‘random’ approach often used by CSK, but is beneficial for studies on cryptic, mobile infaunal species, because it helps ensure that sampling is conducted in suitable habitat, which can be influenced by environmental drivers that are variable at small spatial and temporal scales. The spatio-temporal distribution of macroinfauna on physically dynamic beaches appears to be influenced by a combination of factors, including beach geomorphology, grain size, nutrient availability, picoplankton and other physical, chemical and biological properties (e.g. Ortega Cisneros et al. 2011).
For highly aggregated and mobile species such as D. delatoides (McLachlan et al. 1995), random sampling on long beaches could entirely miss populations and would require much more sampling effort, particularly on beaches were population levels are now very low. By comparison, the IEK approach acknowledges the temporal and spatial variability of D. deltoides within beaches and uses visual indicators to target sites that are most likely to support D. deltoides populations at the time of sampling. The co-designed methodology uses a hierarchical approach, whereby direct or indirect evidence of D. deltoides presence is used first, but if none of these biological indicators is present, geomorphological signs of habitat suitability are used, such as beach slope, penetrability and sand-grain size. This approach improves the efficiency of locating suitable sites for sampling and reduces the likelihood of zero inflated data by maximising the chances of sampling an actual D. deltoides population.
Our co-designed methods were successful in recording spatial and temporal variation in the size–frequency distribution and counts of D. deltoides within three different size classes. A significant interaction was evident between month and location across all size classes. This spatial and temporal variation may be attributed to alongshore theories of beach communities being driven by hydrodynamic and topographic conditions (Defeo and Rueda 2002; Gray 2016b). Sandy-shore invertebrates can be pushed north in the austral autumn and winter, with the southerly winds, and south in the austral spring and summer, with the northerly winds. This study supports this theory. Counts of adults were highest in the northern zone during the austral winter (April–August), juveniles in the middle and southern locations during the austral summer (December–March) and recruits the highest in the southern zone in the austral spring (September–November). Northward movement by adults in winter, followed by a peak spawning in winter–spring, may facilitate greater localised retention of larvae. Along the NSW coast, larvae are transported south with the (spring–summer) northerly winds and the East Australian Current (EAC) (Murray-Jones 1999; Miller et al. 2013).
Temporal variation was evident in adult D. deltoides counts in austral autumn and spring and juveniles and recruits in austral spring and summer. Adult D. deltoides had slightly higher counts in the austral spring (August–October) and the lowest in late austral summer into autumn (January–March). This could be attributed to seasonal zonation as suggested by other studies, where adults move towards the subtidal in the summer and the intertidal in the winter, to avoid thermal stress (Totterman 2019). Alternatively, it could be attributed to harvesting pressure on adult D. deltoides, which can be difficult to assess directly (Gray 2016a; Ferguson et al. 2021). Harvesting pressure was not assessed in this study; however, an overall decline in adult and harvestable D. deltoides was evident. A peak in recruitment was also evident in spring, which aligns with previous studies, showing seasonal peaks in recruitment, also in spring (King 1976; Murray-Jones and Ayre 1997; Murray-Jones 1999). However, the overall numbers of recruits were low. This could be due to relatively low numbers of adult D. deltoides also found on this beach, when in comparison to other studies (Gray 2016b; Ferguson et al. 2021). The low recruitment is consistent with theories of local recruitment via larval retention (Murray-Jones 1999; Bani et al. 2019), and less influence of EAC, which would facilitate re-seeding from external northern beaches, as suggested by other studies in different regions (Versace et al. 2011; Miller et al. 2013).
The environmental parameters, slope, sand-grain size, wave frequency and sand penetrability were the main factors contributing to variation in counts across all size classes. The majority of smaller D. deltoides individuals was found in finer sands on flatter slopes and juveniles and adults were more prevalent in medium to course sand on steeper slopes. This is consistent with other studies on sandy shore molluscs, where recruits were found in the middle of the swash zone and juveniles and adults across the tidal zones (Cardoso and Veloso 2003; Hacking 2007). These trends may be related to the burrowing ability, passive movement and settlement of larvae, settlement cues, and morpho- and aqua-dynamics (McLachlan et al. 1995; Hull et al. 1998; Duong and Fairweather 2011). These results also support the ‘swash riding’ hypothesis, whereby individuals utilise the tides to move up and down the beach and therefore are more likely to be distributed under the influence of tidal and meteorological factors (Ellers 1995; Takeuchi et al. 2015). The ability to burrow and move within the sandy shore environment is more controlled for larger individuals with bigger morphological features to anchor within the sand, whereas smaller individuals can be swept up from the sand easier and are more likely to be moved passively along with the waves (Ellers 1995; Hull et al. 1998; Takeuchi et al. 2015). For the recruits at Stuarts Point Beach, this passive movement south in spring with the tides and winds locates larva in the southern zone near the main river system, which has predominantly finer sand and flatter slopes. This aligns with other studies, indicating recruitment towards freshwater inflow of larger beach systems (Gorman et al. 2010; Versace et al. 2011).
Overall counts of mature and harvestable D. deltoides individuals that were larger than SAM50 and the MLL respectively, were low, and a decline over the sampling period was evident. On the basis of the counts recorded for D. deltoides at Stuarts Point Beach, this survey indicated that there are too few to sustain cultural, recreational and commercial fishing. This is particularly problematic for the Traditional Owners who utilise D. deltoides as a subsistence resource and for cultural practices. Low counts were found across all months and locations on Stuarts Point Beach and were significantly less at the end of the assessment in November 2022, than in May 2021, at the start of the survey period. This indicates a potentially depleted resource that is not accessible to Traditional Owners of this Nation. Stuarts Point Beach is one of the main beaches in the region that is accessed by the Ngambaa community and has historically been culturally harvested and adaptively managed for at least the past 9000 years (Knuckey 1999).
Increased vehicle use was observed from June to November, which may affect beach health. Consequent compaction of sand and beach erosion was observed on Stuarts Point Beach during the assessment. Compacted sand affects the ability of D. deltoides to burrow and move and vehicle pressure can cause high mortality (Schlacher et al. 2008; Takeuchi et al. 2015). To support the enhancement of current depleted D. deltoides populations at Stuarts Point Beach, restrictions to vehicle access are suggested. Although recruits may thrive in fine and very fine sand, grow-out-to-mature (SAM50) and harvestable (MLL) sizes requires medium to coarse sand with greater penetrability and areas with steeper slopes and moderate wave action. A seasonal beach closure during peak spawning times (June–September) and spatial restriction to vehicle access are recommended. For Stuarts Point Beach, ideally, access would be restricted to north of the walking-access point (special restricted licensing during the mullet run). This would enable access to D. deltoides by foot for Traditional Owners and recreational fishers, while leaving a proportion (2–3 km) of the southern section of the beach for replenishment. Further testing of potential pesticide and herbicide run-off from intensive agriculture in these catchments is also recommended for Macleay River creeks to investigate impacts on grow-out for these D. deltoides populations. The primary food source for D. deltoides is phytoplankton, which are known to be affected by a range of herbicides (Wang et al. 2016; Yang et al. 2021). Further ongoing testing on the environmental health of marine and inflow waterways is recommended, as well as cessation of Bitou Bush spraying with herbicide.
Managing fisheries species that are targeted across fishing sectors is essential, if social, cultural, economic and environmental values are to be maintained. More importantly, cultural keystone species are vital for Aboriginal communities as a subsistence resource and for maintaining the transference and evolution of cultural practices and knowledge. However, cultural knowledge requires the maintenance of sustainable populations of cultural keystone species ‘in situ’ for cultural knowledge evolution. Access to cultural keystone species depends on adequate management of resources within clan and Nation areas. Although key fisheries species are managed at state levels, with catch reports within ocean zones, Aboriginal communities continue to operate resource management at Nation levels and often within clan areas. Further to this, Aboriginal communities still operate under traditional management regimes. Surveys with Traditional Owners identified traditional management practises and cultural terms of ‘only taking enough for a feed’, ‘throwing the little ones back’ and ‘adaptive harvesting’, which takes time and effort into account when targeting overall catch (J. Fredericks, pers. comm. with Traditional Owners of Ngambaa). Access to D. deltoides is vital for the Ngambaa and wider Gumbaynggirr and Dhungatti communities to continue to undertake cultural activities and therefore it is imperative that D. deltoides is managed adequately at local clan and Nation scales.
Conclusions
Our co-designed survey method using IEK and CSK, along with the execution of sampling involving Traditional Owners in the data collection, has been successful in assessing spatial and temporal differences in counts and correlations with environmental variables in D. deltoides (garlaany, pipi) at Stuarts Point Beach, Ngambaa Country. Importantly, the surveys highlighted the reasonably low numbers of harvestable-size D. deltoides and mature (SAM50) individuals and the correlation between shell length of D. deltoides and sand-grain size, slope and tide phases. This information can support management strategies towards sustainable harvesting, re-seeding and protection of vital habitat for settlement of recruits and adult grow-out. The scope of this model can be broadened for application to aquatic-resource management statewide, enabling more in-depth assessments of cultural food sources. The abundance and availability of cultural species can be directly affected by the commercial and recreational fishing industries, which impedes access to cultural resources and Aboriginal fishing rights. Further to this, co-design and collaboration with Traditional Owners to collect contemporary count data and size–frequency distributions for important cultural resources fosters stewardship and co-management.
Declaration of funding
Funding for this research project was provided by the Aboriginal Fishing Trust Fund (NSW Department of Primary Industries) and a Southern Cross University RTP scholarship to Jade Fredericks.
Acknowledgements
We extend a most grateful thanks to the Traditional Owners of Gumbaynggirr, Ngambaa, Yaegal and Dhungatti for sharing knowledge, time and for the ongoing collaboration, support and forged partnerships, as well as to the invaluable external collaborators, technical and administration staff and fieldwork volunteers. This project was supported by the Aboriginal Fishing Trust Fund, Southern Cross University, Department of Primary Industries, Coffs Harbour Local Aboriginal Land Council, Gumma Indigenous Protected Area Management, Stuarts Point Traditional Owner Group and Nambucca Heads Local Aboriginal Land Council.
References
Australian Institute of Aboriginal and Torres Strait Islander Studies (2020) AIATSIS Code of conduct for Aboriginal and Torres Strait Islander Research. (AIATSIS) Available at https://aiatsis.gov.au/research/ethical-research [Verified 1 January 2021]
Ban NC, Eckert L, McGreer M, Frid A (2017) Indigenous knowledge as data for modern fishery management: a case study of Dungeness crab in Pacific Canada. Ecosystem Health and Sustainability 3(8), 1379887.
| Crossref | Google Scholar |
Bani R, Fortin M-J, Daigle RM, Guichard F (2019) Dispersal traits interact with dynamic connectivity to affect metapopulation growth and stability. Theoretical Ecology 12, 111-127.
| Crossref | Google Scholar |
Bates D, Mächler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4’. Journal of Statistical Software 67(1), 1-48.
| Crossref | Google Scholar |
Cardoso R, Veloso V (2003) Population dynamics and secondary production of the wedge clam Donax hanleyanus (Bivalvia: Donacidae) on a high-energy, subtropical beach of Brazil. Marine Biology 142, 153-162.
| Crossref | Google Scholar |
Carvalho AN, Vasconcelos P, Piló D, Pereira F, Gaspar MB (2017) Socio-economic, operational and technical characterisation of the harvesting of gooseneck barnacle (Pollicipes pollicipes) in SW Portugal: insights towards fishery co-management. Marine Policy 78, 34-44.
| Crossref | Google Scholar |
Chakraborty S, Gasparatos A (2019) Community values and traditional knowledge for coastal ecosystem services management in the ‘satoumi’ seascape of Himeshima Island, Japan. Ecosystem Services 37, 100940.
| Crossref | Google Scholar |
Cheetnam MD, Keene AF, Bush RT, Sullivan LA, Erskine WD (2008) A comparison of grain-size analysis methods for sand-dominated fluvial sediments. Sedimentology 55(6), 1905-1913.
| Crossref | Google Scholar |
Deepananda KHMA, Amarasinghe US, Jayasinghe-Mudalige UK, Berkes F (2016) Stilt fisher knowledge in southern Sri Lanka as an expert system: a strategy towards co-management. Fisheries Research 174, 288-297.
| Crossref | Google Scholar |
Defeo O, Rueda M (2002) Spatial structure, sampling design and abundance estimates in sandy beach macroinfauna: some warnings and new perspectives. Marine Biology 140, 1215-1225.
| Crossref | Google Scholar |
Department of Primary Industries (2015) Marine biotoxin management plan. NSW Shellfish Program. (NSW DPI Food Authority, NSW Government) Available at https://www.foodauthority.nsw.gov.au/sites/default/files/_Documents/industry/marine_biotoxin_management_plan.pdf
Department of Primary Industries (2023) Aboriginal cultural fishing local management plans. (NSW DPI) Available at https://www.dpi.nsw.gov.au/dpi/fishing/aboriginal-fishing/Aboriginal-cultural-fishing-local-management-plans
Donda S, Manyungwa-Pasani CL (2018) Understanding indigenous knowledge: its role and potential in fisheries resources management in Malawi. Aquatic Ecosystem Health & Management 21, 176-184.
| Crossref | Google Scholar |
Duong HLS, Fairweather PG (2011) Effects of sandy beach cusps on wrack accumulation, sediment characteristics and macrofaunal assemblages. Austral Ecology 36, 733-744.
| Crossref | Google Scholar |
Eckert LE, Ban NC, Frid A, McGreer M (2018) Diving back in time: extending historical baselines for yelloweye rockfish with Indigenous knowledge. Aquatic Conservation: Marine and Freshwater Ecosystems 28, 158-166.
| Crossref | Google Scholar |
Ellers O (1995) Behavioral control of swash-riding in the clam Donax variabilis. The Biological Bulletin 189, 120-127.
| Crossref | Google Scholar | PubMed |
Ferguson GJ, Hooper G, Mayfield S (2021) Temporal and spatial variability in the life-history of the surf clam Donax deltoides: influences of density dependent processes. Estuarine, Coastal and Shelf Science 249, 107104.
| Crossref | Google Scholar |
Food and Agriculture Organization of the United Nations (2020) The state of world fisheries and aquaculture 2020. Sustainability in action. Available at www.fao.org/3/ca9229en/ca9229en.pdf [Verified 10 June 2021]
Garibaldi A, Turner N (2004) Cultural keystone species: implications for ecological conservation and restoration. Ecology and Society 9, 1.
| Crossref | Google Scholar |
Gaspare L, Bryceson I, Mgaya YD (2015) Temporal and spatial trends in size, biomass and abundance of groupers (Epinephelinae) in Mafia Island Marine Park: fishers’ perceptions and underwater visual census surveys. Fisheries Management and Ecology 22, 337-348.
| Crossref | Google Scholar |
Golden AS, Naisilsisili W, Ligairi I, Drew JA (2014) Combining natural history collections with fisher knowledge for community-based conservation in Fiji. PLoS ONE 9, e98036.
| Crossref | Google Scholar |
Gorman D, Fergusan G, Matthews C, Ward TM (2010) Inter-tidal infauna of the Younghusband peninsula: assessing the potential impacts of discharge of hypersaline water. SARDI Publication number F2009/000699-1, South Australian Research and Development Institute (Aquatic Sciences), Adelaide, SA, Australia.
Gray CA (2016a) Effects of fishing and fishing closures on beach clams: experimental evaluation across commercially fished and non-fished beaches before and during harvesting. PLoS ONE 11, e0146122.
| Crossref | Google Scholar |
Gray CA (2016b) Tide, time and space: scales of variation and influences on structuring and sampling beach clams. Journal of Experimental Marine Biology and Ecology 474, 1-10.
| Crossref | Google Scholar |
Hacking N (1998) Macrofaunal community structure of beaches in northern New South Wales, Australia. Marine and Freshwater Research 49, 47-53.
| Crossref | Google Scholar |
Hacking N (2007) Effects of physical state and latitude on sandy beach macrofauna of eastern and southern Australia. Journal of Coastal Research 2007, 899-910.
| Crossref | Google Scholar |
Hull PJ, Cole RG, Creese RG, Healy TR (1998) An experimental investigation of the burrowing behaviour of Paphies australis (bivalvia: Mesodesmatidae). Marine and Freshwater Behaviour and Physiology 31(3), 167-183.
| Crossref | Google Scholar |
James RJ, Fairweather PG (1995) Comparison of rapid methods for sampling the Pipi, Donax deltoides (Bivalvia: Donacidae), on sandy ocean beaches. Marine and Freshwater Research 46, 1093-1099.
| Crossref | Google Scholar |
Kenward MG, Roger JH (1997) Small sample inference for fixed effects from restricted maximum likelihood. Biometrics 53(3), 983-997.
| Google Scholar |
Kijas J (2005) Revival, renewal & return: Ray Kelly & the NSW Sites of significance survey. (NSW Department of Environment and Conservation: Sydney, NSW, Australia) Available at https://www.environment.nsw.gov.au/-/media/OEH/Corporate-Site/Documents/Aboriginal-cultural-heritage/revival-renewal-return-ray-kelly-nsw-sites-of-significance-survey-050002.pdf
Knuckey G (1999) A shell midden at Clybucca, near Kempsey, New South Wales. Australian Archaeology 48, 1-11.
| Crossref | Google Scholar |
Lima MSP, Oliveira JEL, de Nóbrega MF, Lopes PFM (2017) The use of Local Ecological Knowledge as a complementary approach to understand the temporal and spatial patterns of fishery resources distribution. Journal of Ethnobiology and Ethnomedicine 13, 30.
| Crossref | Google Scholar |
McKemey MB, Patterson M (Lesley), Rangers B, Ens EJ, Reid NCH, Hunter JT, Costello O, Ridges M, Miller C (2019) Cross-cultural monitoring of a cultural keystone species informs revival of Indigenous burning of country in South-Eastern Australia. Human Ecology 47, 893-904.
| Crossref | Google Scholar |
McLachlan A, Jaramillo E, Defeo O, Dugan J, de Ruyck A, Coetzee P (1995) Adaptations of bivalves to different beach types. Journal of Experimental Marine Biology and Ecology 187, 147-160.
| Crossref | Google Scholar |
McNiven I (1989) Aboriginal shell middens at the mouth of the Maroochy River, southeast Queensland. Queensland Archaeological Research 6, 28-52.
| Crossref | Google Scholar |
Miller AD, Versace VL, Matthews TG, Montgomery S, Bowie KC (2013) Ocean currents influence the genetic structure of an intertidal mollusc in southeastern Australia – implications for predicting the movement of passive dispersers across a marine biogeographic barrier. Ecology and Evolution 3, 1248-1261.
| Crossref | Google Scholar | PubMed |
Murray-Jones SE, Ayre DJ (1997) High levels of gene flow in the surf bivalve Donax deltoides (Bivalvia: Donacidae) on the east coast of Australia. Marine Biology 128, 83-89.
| Crossref | Google Scholar |
Murray-Jones S, Steffe AS (2000) A comparison between the commercial and recreational fisheries of the surf clam, Donax deltoides. Fisheries Research 44, 219-233.
| Crossref | Google Scholar |
National Health and Medical Research Council (2018) Ethical conduct in research with Aboriginal and Torres Strait Islander peoples and communities: guidelines for researchers and stakeholders. (NHMRC) Available at www.nhmrc.gov.au/about-us/resources/ethical-conduct-research-aboriginal-and-torres-strait-islander-peoples-and-communites [Verified 1 January 2021]
Noble MM, Fulton CJ, Pittock J (2018) Looking beyond fishing: conservation of keystone freshwater species to support a diversity of socio-economic values. Aquatic Conservation: Marine and Freshwater Ecosystems 28, 1424-1433.
| Crossref | Google Scholar |
Nursey-Bray M (2011) Social contexts and customary fisheries: marine protected areas and indigenous use, Australia. Environmental Management 47, 671-683.
| Crossref | Google Scholar | PubMed |
Ortega Cisneros K, Smit AJ, Laudien J, Schoeman DS (2011) Complex, dynamic combination of physical, chemical and nutritional variables controls spatio-temporal variation of sandy beach community structure. PLoS ONE 6(8), e23724.
| Crossref | Google Scholar |
Owner D, Rohweder DA (2003) Distribution and habitat of Pied Oystercatchers (Haematopus longirostris) inhabiting ocean beaches in northern New South Wales. Emu – Austral Ornithology 103, 163-169.
| Crossref | Google Scholar |
Pauly D, Zeller D (2016) Catch reconstructions reveal that global marine fisheries catches are higher than reported and declining. Nature Communications 7, 10244.
| Crossref | Google Scholar |
Prober SM, O’Connor MH, Walsh FJ (2011) Australian Aboriginal people’s seasonal knowledge: a potential basis for shared understanding in environmental management. Ecology and Society 16, 12.
| Crossref | Google Scholar |
Schemmel E, Friedlander AM, Andrade P, Keakealani K, Castro LM, Wiggins C, Wilcox BA, Yasutake Y, Kittinger JN (2016) The codevelopment of coastal fisheries monitoring methods to support local management. Ecology and Society 21, 34.
| Crossref | Google Scholar |
Schlacher TA, Thompson LMC, Walker SJ (2008) Mortalities caused by off-road vehicles (ORVs) to a key member of sandy beach assemblages, the surf clam Donax deltoides. Hydrobiologia 610, 345-350.
| Crossref | Google Scholar |
Schnierer S, Egan H (2016) Composition of the Aboriginal harvest of fisheries resources in coastal New South Wales, Australia. Reviews in Fish Biology and Fisheries 26, 693-709.
| Crossref | Google Scholar |
Short A (2006) Australian beach systems: nature and distribution. Journal of Coastal Research 22, 11-27.
| Crossref | Google Scholar |
Takeuchi S, Yamada F, Shirozu H, Ohashi S, Tamaki A (2015) Burrowing ability as a key trait in the establishment of infaunal bivalve populations following competitive release on an extensive intertidal sandflat. Journal of Experimental Marine Biology and Ecology 466, 9-23.
| Crossref | Google Scholar |
Tavares AS, Mayor P, Loureiro LF, Gilmore MP, Perez-Peña P, Bowler M, Lemos LP, Svensson MS, Nekaris KA-I, Nijman V, Valsecchi J, Morcatty TQ (2020) Widespread use of traditional techniques by local people for hunting the yellow-footed tortoise (Chelonoidis denticulatus) across the Amazon. Journal of Ethnobiology 40, 268-280.
| Crossref | Google Scholar |
Teixeira JB, Martins AS, Pinheiro HT, Secchin NA, Leão De Moura R, Bastos AC (2013) Traditional Ecological Knowledge and the mapping of benthic marine habitats. Journal of Environmental Management 115, 241-250.
| Crossref | Google Scholar | PubMed |
Totterman S (2019) Seasonal zonation patterns of the sandy beach bivalve Donax deltoides (Bivalvia: Donacidae) in subtropical eastern Australia. bioRxiv 2019, 610576 [Preprint, posted 18 April 2019].
| Crossref | Google Scholar |
Turner AD (2021) Traditional food systems. In ‘Social–ecological diversity and traditional food systems: opportunities from the biocultural world’. (Eds RK Singh, NJ Turner, V Reyes-Garcia, J Pretty) (CRC Press: London, UK) 10.1201/9781003246220
Wang C, Lin X, Li L, Lin S (2016) Differential growth responses of marine phytoplankton to herbicide Glyphosate. PLoS ONE 11(3), e0151633.
| Crossref | Google Scholar |
Yang L, Mou S, Li H, Zhang Z, Jiao N, Zhang Y (2021) Terrestrial input of herbicides has significant impacts on phytoplankton and bacterioplankton communities in coastal waters. Limnology and Oceanography 66, 4028-4045.
| Crossref | Google Scholar |