

Stomatal conductance responses to evaporative demand conferred by rice drought-yield quantitative trait locus qDTY12.1
Amelia Henry A C , Hilary Stuart-Williams B , Shalabh Dixit A , Arvind Kumar A and Graham Farquhar B
A C , Hilary Stuart-Williams B , Shalabh Dixit A , Arvind Kumar A and Graham Farquhar B
A International Rice Research Institute, DAPO Box 7777 Metro Manila, Philippines.
B Research School of Biology, Robertson Building (46), Sullivan’s Creek Road, Australian National University, Canberra, ACT 2601, Australia.
C Corresponding author. Email: a.henry@irri.org
Functional Plant Biology 46(7) 660-669 https://doi.org/10.1071/FP18126
Submitted: 15 May 2018 Accepted: 2 March 2019 Published: 8 May 2019
Journal Compilation © CSIRO 2019 Open Access CC BY
Abstract
Rice quantitative trait locus (QTL) qDTY12.1 is a major-effect drought yield QTL that was identified from a cross of Vandana (recipient parent) and Way Rarem (donor parent) through breeding efforts to improve rice yield under upland drought stress conditions. The two main physiological effects previously observed to be related to the presence of qDTY12.1 were (i) increased lateral root growth, and (ii) increased transpiration efficiency. Since relatively more progress has thus far been made on characterising the lateral root growth response related to qDTY12.1, the present study focussed on characterising how qDTY12.1 confers higher transpiration efficiency under upland drought stress in the Vandana background. In a series of field experiments in which stomatal conductance was measured across different times of day in four qDTY12.1 near isogenic lines (NILs), the NILs and Way Rarem showed consistently higher stomatal conductance than Vandana under conditions of low vapour pressure deficit (VPD) and low photosynthetically active radiation (PAR), and consistently lower stomatal conductance than Vandana under high VPD and high PAR. Leaf δ18O was higher in the qDTY12.1 NIL than in Vandana, and although this trend was previously observed for leaf δ13C it appeared to be more consistent across measurement dates and treatments for leaf δ18O. The qDTY12.1 NILs and Way Rarem tended to show greater large vein to small vein interveinal distance and mesophyll area than Vandana, also consistent across treatments. In terms of aquaporin-related plant hydraulics, variation among NILs in terms of aquaporin inhibition of root hydraulic conductivity (Lpr) was observed, with the highest-yielding NIL showing a lack of Lpr inhibition similar to Way Rarem. The results reported here suggest that the effects of qDTY12.1 are in response not only to soil moisture, but also to atmospheric conditions. An interaction among multiple mechanisms including leaf anatomy and aquaporin function appear to confer the transpiration efficiency effect of qDTY12.1.
Additional keywords: aquaporin, isotope, leaf anatomy, photosynthetically active radiation, vapour pressure deficit.
Introduction
Of all rice-growing agro-ecosystems, upland systems are the most drought-prone and produce among the lowest yields, averaging 0.6–1.5 t ha–1 (Singh and Singh 2000). One strategy to improve rice yield in drought-prone upland systems has been to conduct direct selection for yield under drought and identify genetic loci related to increased yield, with identification of the associated physiological mechanisms to be conducted at a later stage (Bernier et al. 2008). The rice quantitative trait locus (QTL) qDTY12.1 is a major-effect drought yield QTL detected from a cross of drought-tolerant upland japonica/aus variety Vandana and drought susceptible indica variety Way Rarem (Bernier et al. 2007). qDTY12.1 is unique in that it was contributed by the drought-susceptible parent in the cross (Way Rarem), and its effect was detected based on increased yield under drought of a variety that was already considered to be drought-tolerant (Vandana). Near isogenic lines (NILs) of qDTY12.1 in the background of Vandana were characterised to show a significant yield advantage across upland environments in the 0.5–2.0 t ha–1 yield range, in comparison with Vandana – an effect which increased with increasing drought severity. Among four NILs characterised, genotype IR 84984-83-15-481-B has consistently shown the highest yield (an increase of up to 1.0 t ha–1 over Vandana) and the most significant physiological differences from Vandana (Henry et al. 2014; Dixit et al. 2015), and is a candidate for varietal release in drought-prone upland areas of east and north-east India.
Across field trials of the NILs, the physiological traits related to qDTY12.1 appeared to be more stable than the yield advantage (Henry et al. 2014). Two main physiological effects have been observed to be related to the presence of qDTY12.1: increased lateral root growth and increased transpiration efficiency (Dixit et al. 2015). Increased lateral root growth/branching was observed in field, soil-filled cylinders, and in PEG-treated gel studies, and increased transpiration efficiency was observed in the field by gas exchange and leaf Δ13C, and gravimetrically in the greenhouse. These two traits were characterised to interact by affecting plant water uptake in that qDTY12.1 NILs showed more conservative water uptake than Vandana early in the drought stress treatment and subsequently greater water uptake than Vandana at later growth stages (see supplementary figure S6D in Dixit et al. (2015)).
Increased lateral root growth and transpiration efficiency as conferred by qDTY12.1 have been observed under drought, but not under well-watered conditions; these two traits appear to act in combination to improve yield under drought (Henry et al. 2014; Dixit et al. 2015). NIL IR 84984-83-15-481-B showed a range of metabolic and proteomic changes under drought that differed from that of Vandana (Raorane et al. 2015a) and Way Rarem (Raorane et al. 2015b). Within a 1.8 Mb qDTY12.1 region, a cluster of 13 genes have been reported to be involved in the molecular mechanisms behind the yield effect of qDTY12.1. Consistent with the physiological characterisation, the expression and interaction of several of these genes appears to be drought-induced and their characterisation has thus far focussed on the lateral root growth response related to qDTY12.1 (Dixit et al. 2015).
To contribute to a better understanding of rice response to upland drought stress, in the present study we focussed on understanding the transpiration efficiency effect of qDTY12.1. Our objective was to characterise how qDTY12.1 confers higher transpiration efficiency under upland drought stress in the Vandana background. Transpiration efficiency can be defined as the amount of water transpired per amount of carbon assimilated. Specifically, transpiration efficiency can be measured as biomass/water transpired (measured gravimetrically), assimilation rate/stomatal conductance (measured by gas-exchange rates), or estimated by isotope ratios, although carbon isotope discrimination has not always consistently reflected transpiration efficiency as compared with the other methods (Vadez et al. 2014). Genetic variation in transpiration efficiency has been observed in several crops in response to varying levels of vapour pressure deficit (VPD, Vadez et al. 2014; Sinclair et al. 2017). As a component trait of transpiration efficiency, aquaporin function has been reported to affect guard cell osmotic pressure regulating stomatal aperturture and whole plant hydraulics, and can be regulated by ambient humidity levels (Maurel et al. 2016). These functions suggest a role of aquaporins in transpiration efficiency. Another component trait reported to affect transpiration via leaf hydraulics is the leaf anatomy, specifically that of the vascular tissue and mesophyll, in Oryza genotypes (Xiong et al. 2017). Therefore we hypothesised that either differences in stomatal conductance at different times of day (at which VPD would vary), aquaporin-related hydraulics, leaf oxygen isotope discrimination (as an alternative to carbon isotope discrimination), or leaf anatomy might explain the overall higher transpiration efficiency of qDTY12.1 lines compared with Vandana.
Materials and methods
Field experiments
Four NILs of qDTY12.1, the recipient parent Vandana, and the donor parent Way Rarem, were grown in five field experiments in the dry season (DS) and wet season (WS) at the International Rice Research Institute, Los Baños, Laguna, Philippines (14°10′N, 121°15′E) from 2013 to 2016 (Table 1). All experiments were conducted under upland conditions in which the soil was dry-ploughed and harrowed, and seeds were sown directly into the dry soil.
|
Seeds (2–3) were sown at 5-cm intervals into rows 3 m in length. Each plot consisted of 3–4 planted rows and experiments were arranged in a randomised complete block design with 3–4 replicates per genotype. Two treatments (drought-stress and well-watered) were planted in separate areas of the fields in each season. The longer duration genotype, Way Rarem, was sown 19 and 30 days before Vandana and the qDTY12.1 NILs in order to impose drought at reproductive stage only in Field Experiment (Exp.) 3 and Field Exp. 4 respectively. After sowing, all experiments were sprinkler irrigated 2–3 times per week or as needed to supplement rainfall. Fertiliser was applied in the furrows at the time of sowing at 50-50-50 (N-P-K) kg ha–1, and 50 kg ha–1 N was applied as a topdressing around 4 weeks after sowing. Sprinkler irrigation was continued throughout the season in the well-watered treatments, whereas irrigation was stopped at 38–56 days after sowing Vandana and the qDTY12.1 NILs in the drought stress treatments. The drought stress treatments were interrupted by rainfall or rewatering, and soil water potential at a depth of 30 cm was recorded from 2–3 tensiometers per experiment. VPD and photosynthetically active radiation (PAR) levels were monitored hourly by the IRRI Experiment Station.
All field physiology measurements were conducted during reproductive stage of Vandana and the qDTY12.1 NILs, i.e. from panicle initiation onwards. Stomatal conductance was measured at different times of day from 42–56 days after sowing (DAS) in Field Exp. 1, 53–69 DAS in Field Exp. 4, and 64–85 DAS in Field Exp. 5 using a diffusion conductance porometer (AP4, Delta-T). Each measurement set was completed within 40 min, and two porometer units were used in Field Exp. 4 and Field Exp. 5 when both well-watered and drought-stress treatments were measured. To compare each genotype with Vandana, the percent increase in stomatal conductance (gs) was calculated as (gs genotype – gs Vandana)/gs Vandana × 100. The percent increase in stomatal conductance was then analysed in respect to the VPD and light intensity values recorded at the time of stomatal conductance measurement. Ten of the youngest, fully-formed leaves per plot were sampled for 18O analysis at 61 and 91 DAS in 2013WS, and at 52 and 67 DAS in Field Exp. 4. In Field Exp. 4, the same leaf samples were also partitioned for 13C analysis. The whole leaves were dried and ground with a Wiley mill (#20 mesh).
Analysis of δ18O was conducted at the Farquhar Laboratory Stable Isotope Facility at the Research School of Biology, Australian National University, Canberra, ACT. Pyrolysis was performed in a nickel-lined ceramic tube on nickelised carbon (Farquhar et al. 1997). The carbon monoxide produced by the reduction of the samples was analysed on an Isoprime IRMS (Micromass). δ13C analysis was conducted by mass spectrometry at the IRRI Analytical Service Laboratory using a wheat flour standard calibrated to Vienna Pee Dee Belemnite (Elemental Microanalysis).
To pinpoint an anatomical phenotype that might be related to the transpiration efficiency effect of qDTY12.1, a range of parameters were measured that are used in standard evaluation of leaf anatomical traits potentially related to photosynthetic efficiency, according to work by Chatterjee et al. (2016). Several first, fully-formed leaves per plot were collected for leaf anatomy analysis at 67 DAS in Field Exp. 3 and 65 DAS in Field Exp. 5. Leaf segments from the central length of each leaf were stored in 75% ethanol until they were hand-sectioned and imaged using a Zeiss Axioplan 2 compound microscope. Three images were taken from each side of the mid-vein from the leaf samples of each plot. A set of 18 anatomical attributes were measured using ImageJ ver. 1.46r (Schneider et al. 2012) and Gimp ver. 2.8 (GNU Image Manipulation Program).
Statistical analysis was conducted by ANOVA and l.s.d. mean comparison in R ver. 3.3.1 (R Core Team 2016).
Greenhouse hydraulic conductance and aquaporin inhibition experiment
The effects of aquaporin inhibition on plant hydraulic conductance in qDTY12.1 NILs was assessed by measuring root hydraulic conductivity (Lpr) with and without application of sodium azide, which has been previously documented as an inhibitor of PIP aquaporin function (Tournaire-Roux et al. 2003). The Lpr experiment was conducted according to the method described by Grondin et al. (2016). Four genotypes (IR 90020:22–283-B-1, IR 84984-83-15-481-B, Vandana and Way Rarem) were sown into soil-filled (920 g soil to a height of 40 cm) 4-cm diameter tubes in five replicates per treatment. Since only 12 plants per day could be accommodated for the Lpr measurement, sowing was staggered over 7 days so that all plants would be the same age at the time of measurement. Plants were grown in a greenhouse at IRRI in January 2014, with an average temperature of 28°C and relative humidity of 58%. Two water treatments (drought-stress and well-watered) and two aquaporin inhibitor treatments within each water treatment (azide and water) were included. The well-watered treatment was maintained by re-watering to field capacity by weight three times per week, and the drought-stress treatment was initiated at 75% of field capacity and allowed to dry down throughout the plant growth period to an average soil moisture level of 50% of field capacity.
Lpr measurements were conducted at 21 DAS by adding up to 150 mL of water or 4 mM NaN3 (azide) for 30 min to the plants in the greenhouse before placing the planted tube into a pressure chamber (300HGBL Plant Water Status Console, Soilmoisture Equipment Corp.) and removing all but the main tiller, which was inserted through the opening of the pressure chamber lid. Two pressure chambers were used simultaneously in order to accommodate Lpr measurements of 12 plants per day, all conducted between 07:00 and 13:00 hours. The chamber was pressurised for 10 min at 0.2 MPa, and then sap was collected from the cut end of the main tiller for 10 min each at 0.2, 0.35, and 0.5 MPa. Lpr in m–3sap m–2root s–1 MPa–1 was determined by calculating the slope of the amount of sap collected across the three pressure levels, normalised for the root surface area and duration of the measurement. Any samples not resulting in linear increases in sap amounts were excluded from further analysis.
Results
Environmental conditions
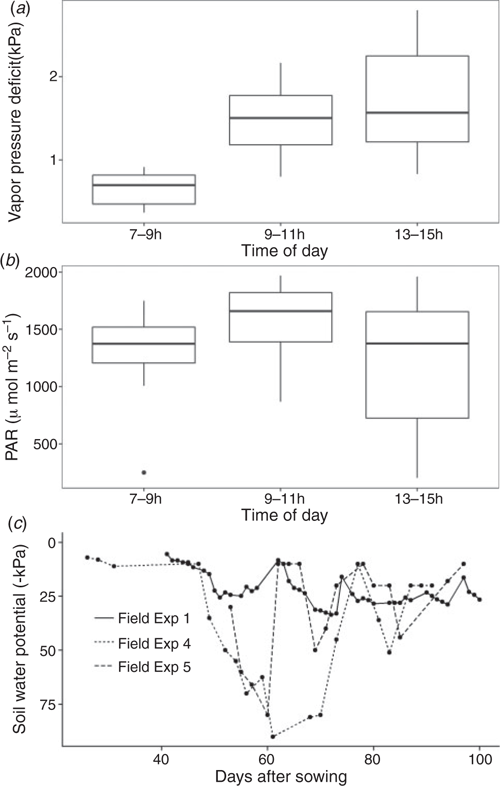
To dissect the mechanisms behind our previous observations of higher transpiration efficiency of qDTY12.1 NILs under drought compared with Vandana, we measured stomatal conductance at multiple times of day in both drought-stress and well-watered field experiments. Across three field experiments, stomatal conductance measurements were conducted during three main times of day (07:00–09:00, 09:00–11:00, and 13:00–15:00 hours) for a total of 16–18 and 7–11 measurements per time period under drought-stress and well-watered treatments, respectively, for each genotype. Of these measurement times, the VPD was typically greatest in the early afternoon reaching >2 kPa (Fig. 1a), and photosynthetically active radiation was typically highest in the late morning reaching >1500 μmol m–2 s–1 (Fig. 1b). Of all field experiments conducted for this study the soil water potential progressed to the most negative values (< –70 kPa) in Field Exp. 4 and Field Exp. 5 (Fig. 1; Fig. S1, available as Supplementary Material to this paper).
|
Diurnal stomatal conductance measurements
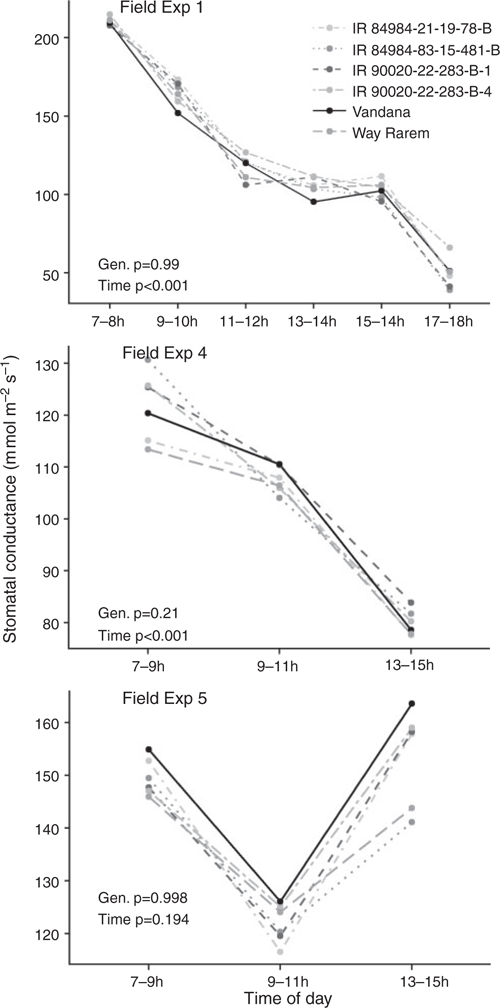
The stomatal conductance of all genotypes declined over the course of the day under drought-stress in Field Exp. 1 (4–5 measurement dates averaged) and under both drought-stress and well-watered treatments in Field Exp. 4 (seven measurement dates averaged), whereas the average stomatal conductance increased in the afternoon measurement (one measurement date only) in both treatments compared with the other measurement times (4–5 dates) in Field Exp. 5 (Figs 2, S2). No genotypic differences were apparent when stomatal conductance was analysed by time of day considering all qDTY12.1 NILs (Figs 2, S2).
|
Stomatal conductance tended to decrease over time as the drought stress progressed in each experiment (Fig. 3). When the best-performing NIL IR 84984-83-15-481-B was compared with the recurrent parent Vandana by time of day and measurement date, some genotypic effects were observed at early measurement dates in the drought treatments (for example, at 09:00–11:00 hours in Field Exp. 1, and at 07:00–09:00 and 09:00–11:00 hours in Field Exp. 4; Fig. 3). Although these trends were more apparent in the drought-stress than in the well-watered treatment (Fig. S3), the times of day at which these differences were observed were not consistent across seasons.

|
When stomatal conductance measurements across seasons, dates, and times of day were grouped based on atmospheric or soil moisture conditions, NIL IR 84984-83-15-481-B showed consistently increased stomatal conductance values compared with Vandana under conditions of relatively lower evapo-transpirational demand (i.e. low VPD (Fig. 4a) and low PAR (Fig. 4b). These genotypic trends were reversed under conditions of high evapo-transpirational demand in that the percent increase in stomatal conductance of IR 84984-83-15-481-B over Vandana was negative when the VPD and PAR levels were relatively high (Fig. 4). These genotypic responses appeared to be specific to changes in atmospheric conditions and did not show them same trends in response to relative changes in soil water potential (Fig. 4c). Similar responses to VPD, PAR, and soil water potential were also observed for the other qDTY12.1 NILs and donor parent Way Rarem (Fig. S4). In the well-watered treatment, most NILs and Way Rarem also showed increased stomatal conductance compared with Vandana under low VPD and low PAR and decreased stomatal conductance compared with Vandana under high VPD and high PAR.

|
Leaf isotopic signatures
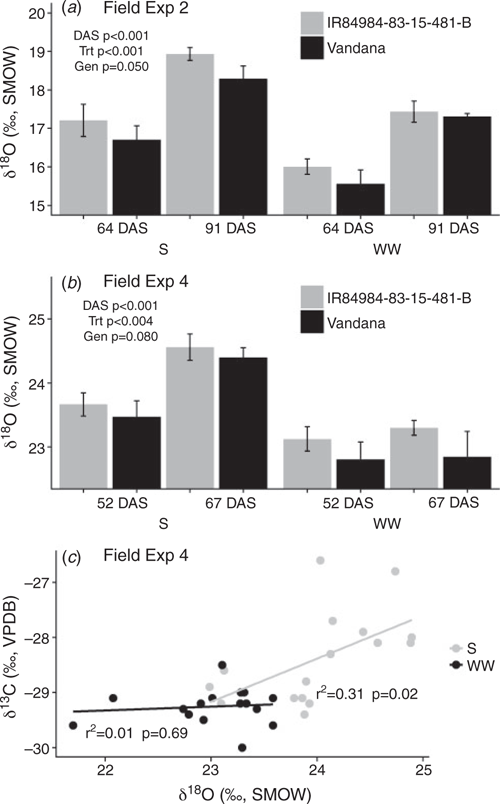
The δ18O values in leaf tissue of IR 84984-83-15-481-B were consistently higher than those of Vandana across field seasons, measurement dates, and treatments (Fig. 5a, b). These genotypic differences in leaf δ18O values were more consistent than the δ13C values measured on the same leaf tissue in Field Exp. 4, which showed only the previously observed trend of less negative δ13C in IR 84984-83-15-481-B compared with Vandana when sampled during moderate drought stress and not under very severe drought stress or well-watered conditions (Fig. S6). Leaf δ18O values generally showed a positive relationship with leaf δ13C values under drought stress, due to the relative increase in δ13C at the sampling date under severe drought stress (Fig. 5c).
|
Leaf anatomy
The effects of experiment, treatment, and genotype on leaf anatomical parameters varied (Table S1, available as Supplementary Material to this paper). The most consistent genotypic effects were observed for the interveinal distance between the large vein and small vein, for which IR 84984-83-15-481-B and Way Rarem were consistently greater than Vandana (Fig. 6a, c, d). The qDTY12.1 NILs and Way Rarem also showed larger mesophyll cell total area (large vein to small vein) than Vandana (Fig. 6b). The interveinal distance between small veins and the large vein height were also affected by genotype, but these effects were mostly driven by Way Rarem rather than any differences between the NILs and Vandana (Fig. S7) and therefore not attributed to qDTY12.1

|
Hydraulic conductance and response to aquaporin inhibition
In the greenhouse study, application of azide to the root zone inhibited the Lpr of all four genotypes by 25–40% in the well-watered treatment (Fig. 7a). In the drought stress treatment, two genotypes, IR 84984-83-15-481-B and Way Rarem, stood out as having little or no inhibition of Lpr by azide application, whereas IR 90020:22-283-B-1 and Vandana showed Lpr reductions of more than 50% by the application of azide (Fig. 7b).

|
Discussion
Results of the present study indicate that under upland drought stress conditions, rice qDTY12.1 confers not only a physiological response to soil moisture levels, but also a response to atmospheric conditions. Multiple component traits appear to be related to the previously reported transpiration efficiency effect of qDTY12.1, some of which have been observed only under drought stress and some of which have been observed under both drought-stress and well-watered conditions.
The stomatal conductance response of qDTY12.1 NILs showed an interesting trend of conserving water under conditions of high evapo-transpirational demand while apparently increasing transpiration under conditions of low evapo-transpirational demand, compared with the recipient parent Vandana. Conservative water uptake under drought and limitation of transpiration at increased VPD levels have been reported as key drought-response strategies in many dryland crops, and these trends have been attributed to partial stomatal closure due to aquaporin-related hydraulics (as reviewed by Sinclair et al. 2017). In the present study the qDTY12.1-related responses to atmospheric conditions were subtle, and detected only when relative genotypic differences in stomatal conductance over multiple measurement dates were considered (Figs 4, S4, S5). However, these stomatal conductance responses to atmospheric conditions (VPD and PAR) were quite consistent among the NILs and the qDTY12.1 donor (Way Rarem) and were observed both in drought-stress and well-watered conditions.
In contrast to the consistent genotypic differences observed in stomatal conductance responses to atmospheric conditions, only the highest-yielding NIL (IR 84984-83-15-481-B) and Way Rarem showed a lack of aquaporin inhibition effects on Lpr under drought (Fig. 7) and the other NIL tested showed similarly strong aquaporin inhibition effects on Lpr under drought to that of Vandana. However, it is unclear whether a lack of aquaporin inhibition effect on Lpr is correlated with improved performance under drought. Grondin et al. (2016) also reported varied levels of aquaporin inhibition effects on plant hydraulics among drought-tolerant genotypes. Furthermore, the trends in Lpr observed here may differ from trends in leaf hydraulics, and may also represent some side effects due to toxicity of the azide.
Another measurement in which the highest-yielding NIL was consistently different from Vandana across treatments was leaf δ18O. Although genotypic differences in leaf δ13C were also observed, those differences were only under moderate drought conditions and not under severe drought or well-watered conditions in this study and as previously reported (Henry et al. 2014). One reason why IR 84984-83-15-481-B and Vandana showed more consistent differences in leaf δ18O than in leaf δ13C could be the processes integrated by each measurement; leaf δ13C is likely affected by both stomatal conductance and photosynthetic capacity whereas leaf δ18O appears to be mainly sensitive to changes in stomatal conductance (Barbour et al. 2000). In the present study, the consistently higher δ18O values of IR 84984-83-15-481-B reflect a net limitation of transpiration compared with Vandana. The relative trends in leaf δ13C were more similar than leaf δ18O to the trends in grain yield, in which IR 84984-83-15-481-B typically shows higher yield than Vandana under moderate levels of drought stress, but not under very severe drought or well-watered conditions (Henry et al. 2014). Similarly, a closer relationship between grain yield and grain δ13C than that of grain δ18O was reported for Mediterranean drought-prone environments in wheat (Araus et al. 2013).
Greater large vein-to-small vein leaf interveinal distances and resulting large vein-to-small vein mesophyll areas in the NILs and Way Rarem were also observed in both drought and well-watered treatments (Fig. 6), but the relationship between this trait and transpiration efficiency is unclear. It may be that larger interveinal distance presents a diffusional constraint to water, resulting in reduced mesophyll conductance of water and limited stomatal conductance in the NILs and Way Rarem. However, this effect would be in contrast to that reported by Xiong et al. (2015) in which larger interveinal distances in rice were correlated with higher leaf hydraulic conductance. Furthermore, rice leaves have shown decreased mesophyll conductance under drought stress that was completely reversible by rewatering (Centritto et al. 2009), suggesting that the effects of anatomy on mesophyll conductance under drought may not be significant.
It is notable that the stomatal conductance response to ambient conditions, and the differences in leaf δ18O and anatomy between the qDTY12.1 NILs and Vandana all showed consistent trends across drought and well-watered treatments, but that the leaf δ13C and previously reported transpiration efficiency (through gravimetric, gas exchange and δ13C methods) and grain yield differences were observed only under moderate to severe drought stress. This may be due to an interaction of constitutively reduced stomatal conductance with the increase in lateral root growth under drought that affected water uptake (Henry et al. 2014; Dixit et al. 2015).
Although apparently common in dryland crops, the qDTY12.1-related transpiration efficiency response observed under upland drought conditions has not been frequently reported in rice. For example, we have observed the reverse trend in transpiration efficiency under lowland drought conditions as conferred by qDTY2.2 + qDTY4.1 in the IR64 background, in which the NILs showed higher Δ13C values related to deep root growth and lower canopy temperature under drought (Henry et al. 2015). In fact, drought avoidance (increased water uptake through improved water acquisition by roots) has been cited as one of the most promising strategies to increasing rice productivity under drought (Serraj et al. 2009). The discrepancies among rice agro-ecosystems and cultivars reflect the wide range of types of drought, and the wide range of physiological responses to those types of drought, that occur in rice crops. Compared with drought stress in rainfed lowland rice-growing environments, the drought stress that typically occurs in upland rice environments develops much more quickly and reaches lower soil moisture levels (O’Toole and Chang 1978). qDTY12.1 has already been reported to increase yield under drought in other genetic backgrounds under lowland drought stress (Mishra et al. 2013). Considering the potentially different plant responses to lowland drought stress, it could be useful to quantify the relative effects of qDTY12.1 on parameters related to transpiration efficiency in those conditions.
Conflicts of interest
The authors declare no conflicts of interest.
Acknowledgements
We thank Eugene Aquino for providing the VPD and solar radiation data. Rolando Torres, Marinell Ramirez, Leo Holongbayan, Nancy Sadiasa, Eleanor Mico, Ariston Reyes, Lesly Satiouquia, Allan Los Añes, Mignon Natividad, and Carlo Cabral contributed to the field experiments and physiological measurements. We thank W. Paul Quick for constructive comments on the manuscript. This work was supported by the Bill and Melinda Gates Foundation STRASA project (Stress Tolerant Rice for Africa and South Asia).
References
Araus JL, Cabrera-Bosquet L, Serret MD, Bort J, Nieto-Taladriz MT (2013) Comparative performance of δ13C, δ18O and δ15N for phenotyping durum wheat adaptation to a dryland environment. Functional Plant Biology 40, 595–608.| Comparative performance of δ13C, δ18O and δ15N for phenotyping durum wheat adaptation to a dryland environment.Crossref | GoogleScholarGoogle Scholar |
Barbour MM, Fischer RA, Sayre KD, Farquhar GD (2000) Oxygen isotope ratio of leaf and grain material correlates with stomatal conductance and grain yield in irrigated wheat. Australian Journal of Plant Physiology 27, 625–637.
Bernier J, Kumar A, Venuprasad R, Spaner D, Atlin GN (2007) A large-effect QTL for grain yield under reproductive-stage drought stress in upland rice. Crop Science 47, 507–516.
| A large-effect QTL for grain yield under reproductive-stage drought stress in upland rice.Crossref | GoogleScholarGoogle Scholar |
Bernier J, Atlin GN, Serraj R, Kumar A, Spaner D (2008) Breeding upland rice for drought resistance. Journal of the Science of Food and Agriculture 88, 927–939.
| Breeding upland rice for drought resistance.Crossref | GoogleScholarGoogle Scholar |
Centritto M, Lauteri M, Monteverdi MC, Serraj R (2009) Leaf gas exchange, carbon isotope discrimination, and grain yield in contrasting rice genotypes subjected to water deficits during the reproductive stage. Journal of Experimental Botany 60, 2325–2339.
| Leaf gas exchange, carbon isotope discrimination, and grain yield in contrasting rice genotypes subjected to water deficits during the reproductive stage.Crossref | GoogleScholarGoogle Scholar | 19443613PubMed |
Chatterjee J, Dionora J, Elmido-Mabilangan A, Wanchana S, Thakur V, Bandyopadhyay A, Brar DS, Quick WP (2016) The evolutionary basis of naturally diverse rice leaves anatomy. PLoS One 11, e0164532
| The evolutionary basis of naturally diverse rice leaves anatomy.Crossref | GoogleScholarGoogle Scholar | 27792743PubMed |
Dixit S, Kumar Biswal A, Min A, Henry A, Oane RH, Raorane ML, Longkumer T, Pabuayon IM, Mutte SK, Vardarajan AR, et al (2015) Action of multiple intra-QTL genes concerted around a co-localized transcription factor underpins a large effect QTL. Scientific Reports 5, 15183
| Action of multiple intra-QTL genes concerted around a co-localized transcription factor underpins a large effect QTL.Crossref | GoogleScholarGoogle Scholar | 26507552PubMed |
Farquhar GD, Henry BK, Styles JM (1997) A rapid on-line technique for determination of oxygen isotope composition of nitrogen-containing organic matter and water. Rapid Communications in Mass Spectrometry 11, 1554–1560.
| A rapid on-line technique for determination of oxygen isotope composition of nitrogen-containing organic matter and water.Crossref | GoogleScholarGoogle Scholar |
Grondin A, Mauleon R, Vadez V, Henry A (2016) Root aquaporins contribute to whole plant water fluxes under drought stress in rice (Oryza sativa L.). Plant, Cell & Environment 39, 347–365.
| Root aquaporins contribute to whole plant water fluxes under drought stress in rice (Oryza sativa L.).Crossref | GoogleScholarGoogle Scholar |
Henry A, Dixit S, Mandal NP, Anantha MS, Torres R, Kumar A (2014) Grain yield and physiological traits of rice lines with the drought yield QTL qDTY12.1 showed different responses to drought and soil characteristics in upland environments. Functional Plant Biology 41, 1066–1077.
| Grain yield and physiological traits of rice lines with the drought yield QTL qDTY12.1 showed different responses to drought and soil characteristics in upland environments.Crossref | GoogleScholarGoogle Scholar |
Henry A, Swamy BPM, Dixit S, Torres RD, Batoto TC, Manalili M, Anantha MS, Mandal NP, Kumar A (2015) Physiological mechanisms contributing to the QTL-combination effects on improved performance of IR64 rice NILs under drought. Journal of Experimental Botany 66, 1787–1799.
| Physiological mechanisms contributing to the QTL-combination effects on improved performance of IR64 rice NILs under drought.Crossref | GoogleScholarGoogle Scholar | 25680791PubMed |
Maurel C, Verdoucq L, Rodrigues O (2016) Aquaporins and plant transpiration. Plant, Cell & Environment 39, 2580–2587.
| Aquaporins and plant transpiration.Crossref | GoogleScholarGoogle Scholar |
Mishra KK, Vikram P, Yadaw RB, Swamy BP, Dixit S, Cruz MT, Maturan P, Marker S, Kumar A (2013) qDTY12.1: a locus with a consistent effect on grain yield under drought in rice. BMC Genetics 14, 12
| qDTY12.1: a locus with a consistent effect on grain yield under drought in rice.Crossref | GoogleScholarGoogle Scholar | 23442150PubMed |
O’Toole JC, Chang TT (1978) ‘Drought and rice improvement in perspective.’ (International Rice Research Institute: Los Baños, Philippines)
R Core Team (2016) ‘R: A language and environment for statistical computing.’ (R Foundation for Statistical Computing: Vienna, Austria) Available at http://www.R-project.org/ [Verified 4 March 2019]
Raorane ML, Pabuayon IM, Miro B, Kalladan R, Reza-Hajirezai M, Oane RH, Kumar A, Sreenivasulu N, Henry A, Kohli A (2015a) Variation in primary metabolites in parental and near isogenic lines of the QTL qDTY12.1: altered roots and flag leaves but similar spikelets of rice under drought. Molecular Breeding 35, 138
| Variation in primary metabolites in parental and near isogenic lines of the QTL qDTY12.1: altered roots and flag leaves but similar spikelets of rice under drought.Crossref | GoogleScholarGoogle Scholar | 26069451PubMed |
Raorane NL, Pabuayon IM, Varadarajan AR, Mutte SK, Kumar A, Treumann A, Kohli A (2015b) Proteomic insights into the role of the large-effect QTL qDTY12.1 for rice yield under drought. Molecular Breeding 35, 139
| Proteomic insights into the role of the large-effect QTL qDTY12.1 for rice yield under drought.Crossref | GoogleScholarGoogle Scholar |
Schneider CA, Rasband WS, Eliceiri KW (2012) NIH Image to ImageJ: 25 years of image analysis. Nature Methods 9, 671–675.
| NIH Image to ImageJ: 25 years of image analysis.Crossref | GoogleScholarGoogle Scholar | 22930834PubMed |
Serraj R, Kumar A, McNally KL, Slamet-Loedin I, Bruskiewich R, Mauleon R, Cairns J, Hijmans RJ (2009) Improvement of drought resistance in rice. Advances in Agronomy 103, 41–99.
| Improvement of drought resistance in rice.Crossref | GoogleScholarGoogle Scholar |
Sinclair TR, Devi J, Shekoofa A, Choudhary S, Sadok W, Vadez V, Riara M, Rufty T (2017) Limited-transpiration response to high vapor pressure deficit in crop species. Plant Science 260, 109–118.
| Limited-transpiration response to high vapor pressure deficit in crop species.Crossref | GoogleScholarGoogle Scholar | 28554468PubMed |
Singh VP, Singh RK, Eds (2000) ‘Rainfed rice: a sourcebook of best practices and strategies in eastern India.’ (International Rice Research Institute: Los Baños, Philippines)
Tournaire-Roux C, Sutka M, Javot H, Gout E, Gerbeau P, Luu DT, Bligny R, Maurel C (2003) Cytosolic pH regulates root water transport during anoxic stress through gating of aquaporins. Nature 425, 393–397.
| Cytosolic pH regulates root water transport during anoxic stress through gating of aquaporins.Crossref | GoogleScholarGoogle Scholar | 14508488PubMed |
Vadez V, Kholova J, Medina S, Kakkera A, Anderberg H (2014) Transpiration efficiency: new insights into an old story. Journal of Experimental Botany 65, 6141–6153.
| Transpiration efficiency: new insights into an old story.Crossref | GoogleScholarGoogle Scholar | 24600020PubMed |
Xiong D, Yu T, Zhang T, Li Y, Peng S, Huang J (2015) Leaf hydraulic conductance is coordinated with leaf morpho-anatomical traits and nitrogen status in the genus Oryza. Journal of Experimental Botany 66, 741–748.
| Leaf hydraulic conductance is coordinated with leaf morpho-anatomical traits and nitrogen status in the genus Oryza.Crossref | GoogleScholarGoogle Scholar | 25429002PubMed |
Xiong D, Flexas J, Yu T, Peng S, Huang J (2017) Leaf anatomy mediates coordination of leaf hydraulic conductance and mesophyll conductance to CO2 in Oryza. New Phytologist 213, 572–583.
| Leaf anatomy mediates coordination of leaf hydraulic conductance and mesophyll conductance to CO2 in Oryza.Crossref | GoogleScholarGoogle Scholar | 27653809PubMed |