

Seasonal flowering and evolution: the heritage from Charles Darwin
R. W. King A C and O. M. Heide BA CSIRO Plant Industry, P.O. Box 1600, Canberra, ACT 2601, Australia.
B Department of Ecology and Natural Resource Management, Norwegian University of Life Sciences, P.O. Box 5003, NO-1432 Ås, Norway.
C Corresponding author. Email: rod.king@csiro.au
This paper is part of an ongoing series: ‘The Evolution of Plant Functions’.
Functional Plant Biology 36(12) 1027-1036 https://doi.org/10.1071/FP09170
Submitted: 8 July 2009 Accepted: 2 September 2009 Published: 3 December 2009
Abstract
To survive, plants optimise their seasonal flowering time and set seed to avoid extremes of the environment including frost, heat and drought. Additionally, pollination may need to be tightly regulated in time so that it coincides with flowering of other individuals and/or with the presence of bird or insect pollinators. It is now clear that plants use seasonal changes in natural light intensity, daylight duration and temperature to achieve reproducible timing of flowering year-in-year-out. In more recent studies, genetic and molecular approaches are beginning to provide a basis for understanding heritability, an essential component of Darwin’s concept of evolution.
Additional keywords: Arabidopsis, Bromus, daylength, FLC, Fragaria, FRI, FT, light intensity, Pimelea, Poa, speciation, temperature, Themeda.
Introduction
Charles Darwin wrote in ‘The Origin of Species’:
‘the organs of vegetation on which their (the plants) whole life depends are of little signification compared with organs of reproduction with their product the seed of paramount importance’ (Darwin 1859, p. 414).
Control of flowering is placed centrally in the evolution of plants by Darwin’s recognition that species survive best if they produce more seed. Environmental regulation of seasonality of flowering and set seed is critical for this survival as it allows seeds to develop in the most favourable conditions. However, it was not until the early part of the last century that some understanding of seasonal regulation of flowering was reported. Both daylength (photoperiod; see Garner and Allard 1920) and winter cold (vernalisation; see Gassner 1918) were shown to regulate flowering time and Bünning (1936) proposed that endogenous circadian rhythms provided the clock mechanism that was essential for accurate measurement of the length of day. Armed with the knowledge of how the extent and timing of flowering is regulated seasonally and with current evidence of its molecular and genetic regulation, Darwin would certainly have speculated on the evolutionary importance of the regulation of flowering.
A further aspect of Darwin’s view of evolution related to plant migration leading to geographical isolation:
‘All the grand leading facts of geographical distribution are explicable in the theory of migration together with subsequent modification and the multiplication of new forms’ (Darwin 1859, p. 408).
Thus, adaptation to different latitudinal environments should be evident in the types of seasonal inputs used by plants to control their flowering.
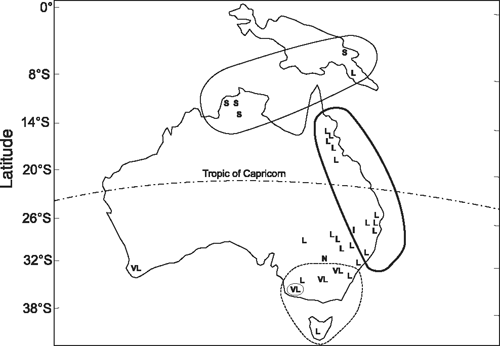
An early study of flowering response with 31 lines of an Australasian perennial grass, Themeda australis (R. Br.) Stapf, collected over a wide geographical range of origin illustrates the match between the type of response to environment and expectation for each line based on site daylength and temperature. In controlled environment studies, strains of Themeda from latitudes 6°S to 15°S flowered in short days (SD or day neutral plants; Fig. 1). Those from mid latitudes where daylength is longer, responded to longer daylengths (LD), while accessions from more southerly, colder sites, required prolonged exposure to winter cold (vernalisation) followed by LD. A daylength-insensitive selection was found in a desert environment where independence from normal seasonal cues would be required for rapid flowering (Evans and Knox 1969).
|
Darwin allowed for flexibility in the path to speciation. He accepted the possibility of species retaining similarities as in those with bipolar distribution. In contrast, he also accepted differences associated with geographical isolation and subsequent random population drift. As Darwin observed during his voyage on the Beagle, Australian species provide an excellent illustration of the effects of divergence following isolation. In some regions of Australia, 75% of the plant species are endemic and in their evolution they developed unique shapes and forms especially of their flowers. Interestingly, despite this clear divergence in plant shape and form, Australian species and those across the globe utilise common mechanisms for regulating seasonal flowering (see above for Themeda) although, as discussed below, across species, and even between individual plants within a species, there is great flexibility in the choice of environmental signals used to regulate seasonality of flowering.
The seasonal environment and flowering
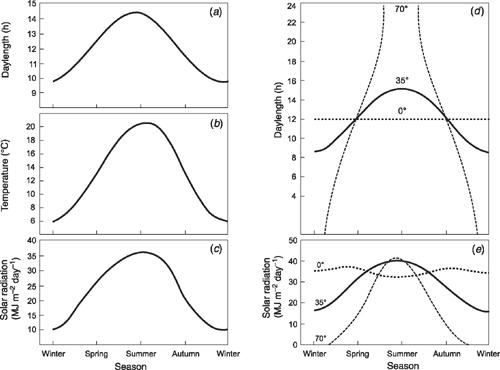
The seasonal pattern of change in daylength (Fig. 2a, d) is identical from year to year and many plants regulate their flowering by responding to daylengths shorter than a critical length, (short day responsive types, SDP), or to daylengths exceeding a critical length (long day plants, LDP) (see summary in Thomas and Vince Prue 1997). Summer flowering at high latitudes typically will involve a LD response (cf. Fig. 2d) but there are notable exceptions. For example, Koenigia islandica L., a tiny arctic–alpine annual plant with geographic distribution extending beyond 80°N in Spitzbergen and Greenland is day neutral across a temperature range of 6−21°C (Heide and Gauslaa 1999). Conversely, insensitivity might be expected in the tropics where there is little or no change in daylength. Nevertheless, some tropical species flower in response to SD and their time measurement is so precise that they can detect seasonal daylength changes of 1–3% (i.e. 7 to 21-min difference in the length of the day). Such sensitivity is seen in rice (Oryza sativa L.), a tropical SDP, where some lines respond to a SD of 11 h 50 min, while others respond to 12 h; the 10 min longer daylength delaying flowering by 30 to 50 days (Dore 1959).
|
For mid latitudes, despite its wide daily fluctuations, temperatures averaged over many years show consistent seasonal patterns (Fig. 2b for average monthly temperatures at Canberra, Australia, 35.3°S). At higher latitudes and at altitude, there is comparable seasonality but with a greater range while, over the year, temperature varies little at the equator. For many species, flower number and, hence, seed set declines almost linearly with increase in temperature above 20 to 25°C as in an Australian perennial shrub Crowea exalata F. Muell (King et al. 2008a and references therein for other species). Thus, flowering early in spring is beneficial for avoiding later higher temperatures but not if too early as flowers can be damaged by winter/early spring frosts. To meet these seasonal requirements, some species only flower after up to 2–3 months winter vernalisation involving exposure to cold temperatures below 10 to 12°C (see Lang 1965 and references therein). Other species may only flower when temperatures reach between 15 and 22°C although they may grow at temperatures outside this range (see references in Heide 1994; King 1998). When flowering is induced by these mild temperatures, there may be little or no effect of daylength (see King et al. 1996) or both mild temperature and daylength may be important (see later).
Light intensity, although highly variable from day to day, is rather constant near the equator but shows marked seasonality further away (Fig. 2c, e). As well as responding to daylength, several plant species show dramatic increases in flowering with increasing light intensity. Independence between a true photosynthetic and a true daylength response is clear when the daily light integral is equalised (see King et al. 2008a and references therein). Further, as discussed later, total radiation in combination with a thermal sum strongly correlates with reproductive output (see Rawson 1988; for wheat (Triticum aestivum L.) grown at different latitudes). Clearly, along with daylength, increasing light intensity in spring and summer is likely to impact strongly on flowering.
Rainfall shows seasonal consistency in tropical areas where there is a marked monsoon. However, it may be the drop in temperature associated rainfall that leads to flowering of tropical species such as mango (Mangifera indica L.; Wilkie et al. 2008). Although its significance is unclear, Stinchcombe et al. (2004) reported an interesting finding in a common garden experiment at one site with lines of the annual herb, Arabidopsis thaliana (L.), differing in the FRIGIDA (FRI) gene for vernalisation response. Flowering was associated with January rainfall at their sites of origin. Perhaps there has been selection pressure for other factors including photosynthesis and plant form, which could allow a flowering response to variable inputs including winter rainfall.
The regulation of flowering involves multiple adaptations to local environments
Response to daylength and vernalisation
SD and LD photoperiodic responses often predominate in annual plants, which complete their entire life cycle in a single growing season but perennials may also show simple one component responses. For example, as indicated in Fig. 1, different lines of the perennial grass Themeda flower in LD, in SD, in all daylengths (day neutral plants), or only in intermediate daylengths of ~12 h. Two component responses involving exposure first to winter vernalisation and then to LD may also be important in Themeda (Fig. 1) as also in some herbaceous winter annuals including Arabidopsis and in cereals including wheat (see references in Thomas and Vince Prue 1997). However, at least for Arabidopsis, it can also behave as a summer annual, other of its multiple floral pathways allowing it to bypass its vernalisation requirement.
Molecular and genetic studies with Arabidopsis have highlighted critical roles in vernalisation for two genes, FRI and FLOWERING LOCUS C (FLC). Cold conditions promote flowering by causing FLC degradation while FRI delays flowering by promoting FLC expression. The most important genes for its LD response are CONSTANS (CO) and FLOWERING LOCUS T (FT) (reviewed in Turck et al. 2008).
As for some annuals, perennials often flower after exposure to winter chilling (vernalisation) followed by LD in late spring and summer (Fig. 1). Other perennials may show a dual daylength induction response and this is particularly important for perennial plants. A dual daylength requirement for sequential SD and LD, or LD and SD enables a plant to distinguish between spring and autumn when photoperiodic conditions are identical. Because the sequence of inductive conditions is crucial, dual induction photoperiodic plants will not flower in either constant SD or constant LD conditions and, hence, they are distinct from daylength neutral species (Heide 2004). Such a dual daylength response is also not to be confused with intermediate daylength responses where LD and SD inputs may be satisfied simultaneously (Runkle et al. 2001).
Dual daylength, short-long-day plants are found in cold temperate regions where both temperature and daylength vary greatly between seasons, and where early flowering is important for seed maturation. Some tropical and sub-tropical plants also show a dual daylength adaptation but often require the opposite sequence of daylength (long-short-day plants see references in Lang 1965; Thomas and Vince Prue 1997).
Interestingly, in some species, winter SD can substitute for the winter cold response and for many perennial plants, notably temperate perennial grasses and sedges, the two types of winter responses exist side by side and may act independently and interchangeably (Heide 1994, 1997). The two pathways are distinct, as demonstrated by differences in critical exposure times and other kinetic characteristics but, as an evolutionary mechanism, the versatility of the alternative SD/vernalisation primary induction system offers a beautiful safety mechanism with SD acting as a fall-back alternative in case of inadequate winter chill.
Response to mild temperatures
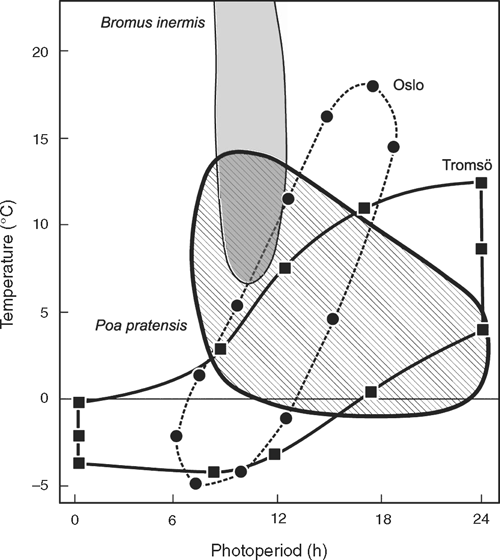
Another two-component, dual-induction response is seen in species that flower in response to moderate/mild temperatures (>12°C) and sometimes along with simultaneous exposure to an appropriate daylength. In such species, the physical displacement between average temperature and daylength (e.g. Fig. 2a, c) can be represented by the physical photothermal envelopes shown for two locations in Norway in Fig. 3 (lines joining the symbols).
|
The impact of such seasonal phototherms on flowering is illustrated in Fig. 3 for the grass Bromus inermis Leyss., an American species introduced to Scandinavia over 100 years ago. It has an obligatory SD requirement for primary floral induction but the experimentally determined response window for B. inermis only overlaps the physical phototherm for the lower latitude (55°55′N) where it is currently found. Thus, precision in the timing of seasonal flowering of B. inermis places clear geographical constraints on its survival. In contrast, Poa pratensis L., a native arctic alpine grass species, would be successful in both environments (Fig. 3). Precision similar to that in B. inermis is evident in several perennial grasses where their seasonal flowering may require a combination of vernalisation and/or SD during autumn and winter, followed by increasing daylength (LD) and warmer temperatures in spring (see Heide 1994).
Recent studies with strawberry (Fragaria spp.) further demonstrate how combinations of daylength and temperature enhance seasonal flowering and, conversely, inappropriate experimental or natural conditions cause a reduction in flowering associated with increased runnering and vegetative growth. For example, a range of Norwegian populations of the diploid wood strawberry (Fragaria vesca L.) all initiated flowers in both SD and LD at 9°C, they only flowered in SD at 15 and 18°C while they did not flower at all at 21°C (Heide and Sønsteby 2007; and references therein). Conversely, leaf growth and stolon formation (runnering) were stimulated by LD and high temperature. Similar, but less extreme temperature modifications of the daylength responses are found in F. × ananass Duch, the common June-bearing (octoploid) cultivated strawberries (Heide 1977; Sønsteby and Heide 2006), and are inherited from the American octoploid parental species Fragaria chiloense L. and Fragaria virginiana Duch (Sønsteby and Heide 2008, 2009 and references therein). In contrast, the ever-bearing (recurrent-flowering) strawberry cultivars are LDPs, which flower freely in LD at temperatures as high as 27°C (Sønsteby and Heide 2007). Clearly, within a single genus there is great potential for creating new and environmentally adapted types under both natural and artificial selection pressures.
The negative effects of high temperature on flower number of strawberry (Sønsteby and Heide 2007) highlight a further complexity of environmentally regulated flowering, which is dramatically illustrated by the interplay between temperature and daylength for the leguminous Australian perennial Hardenbergia violaceae (Schneeve) Stem. This latter species requires SD of 12 h or less for flowering but it only sets seed at an average daily temperature of 15°C, but not at 12°C or 18°C (King 1998). At 21°C, flowers form but quickly abort and, presumably, there was also abortion at 18°C. Thus, in nature, the very narrow photothermal window of Hardenbergia allows flowering in late autumn and early spring so that seeds develop in spring. Then with rising summer temperatures, its seasonal window for reproduction closes and the plant returns to vigorous vegetative growth and any immature seed rapidly abort (King 1998).
Response to light intensity
Although photoperiod may be a dominant factor of the light environment, radiation can also have large effects and especially in LD species. In the annual grass Lolium temulentum L., irradiance in SD for the 2 weeks before exposure to a single florally-inductive LD was highly correlated with the rate of flower development. Without the LD exposure, the same full sunlight exposure did not induce flowering (King and Evans 1991). Furthermore, when light intensity was varied during 16 h of the single LD, flowering was strongly enhanced in association with a photosynthetically-driven increase in shoot apex sugar content (King and Evans 1991).
In Arabidopsis, the link between photosynthesis and flowering is both compelling and even more immediate. A single photosynthetic LD upregulates expression of the gene FT whose protein is a transmitted florigen (reviewed in Turck et al. 2008; for Arabidopsis and other LD and SD plants). For Arabidopsis, within 8 h of starting a single high irradiance LD, there is a photosynthetically-driven increase in leaf and shoot apex sucrose content. At the same time, FT expression in the leaf blade increases and by 10 days, flower buds are evident (King et al. 2008b). Independently, a separate photoresponse via phytochrome causes FT upregulation, but as part of a low light photoperiodic LD response involving no photosynthetic input (King et al. 2008b). Consequently, in nature, the LD regulation of flowering via FT will involve both a response to the photosynthetic light integral and to a light durational component.
As an aside, it is unclear why high light intensity upregulates FT expression in LD but not in SD. In SD, increasing light intensity to give matched daily light integrals only slightly increased FT expression (King et al. 2008b). Perhaps, in addition to phytochrome, there is a second photoresponse in the LD photoperiod via the CRYPTOCHROME (CRY) photoreceptor proteins, which respond to blue-rich light.
Like L. temulentum and Arabidopsis, several other LD plants show independent and additive flowering responses to light intensity (daily light integral) and daylength (King and Ben-Tal 2001 and references therein). In SD species, the daily light integral is unlikely to be as important while some species, such as the herbaceous perennial C. exalata, may be unresponsive to daylength but flower when their daily light receipt exceeds a third or more of full sunlight levels (King et al. 2008a and see references therein). Seasonally, such a light intensity regulation of flowering of Crowea is ‘opportunistic’ and somewhat atypical, but clearly, some qualification is required of Darwin’s claim that there is ‘little signification of organs of vegetation’ (Darwin 1859, p. 414).
Response to rainfall and drought
Opportunistic, drought-avoidance flowering of the ephemeral desert annual, Pectis papposa Harvey and A. Gray, illustrates an extreme in adaptation involving insensitivity to both daylength and temperature. After rainfall, it germinates and forms as few as two or three leaves before flowering with either the terminal shoot apex or the axillary buds becoming floral. In this way, should rainfall persist, some vegetative meristems are available for further growth and reproduction (Hayashi et al. 1994). Adaptively, there is some similarity between the rapid flowering of P. papposa and that of the arctic annual K. islandica, which is also constrained by a brief growing season (Heide and Gauslaa 1999). Such adaptation for rapid flowering could be characterised by little or no dependence on flowering time genes.
The persistence of vegetative meristems when P. papossa flowers is consistent with the way perennation is achieved in Arabis alpina L. where the PERPETUAL FLOWERING 1 (PEP1) gene blocks flowering of some branch meristems. Interestingly, like its Arabidopsis homologue FLC, expression of PEP1 is under epigenetic control in response to temperature and seasonal perennation is reversible through loss and gain of chromatin modifications (Wang et al. 2009). Additional meristem modifications in other species are likely to relate to genes such as TERMINAL FLOWER (TFL) and the MORE AXILLARY GROWTH/RAMOSUS (MAX/RMS) branching genes (Beveridge 2006). For example, in citrus before flowering and fruiting, the expression of a homologue of TFL is elevated in juvenile trees and then declines in association with the development of a sufficient vegetative structure (Pillitteri et al. 2004). Similarly, for the perennial grass, Lolium perenne (L.), seasonally reduced expression of a TFL homologue may also be essential for its flowering as suggested by Jensen et al. (2001).
Species in their natural environment: processes regulating flowering show geographical adaptation
Several studies have provided broad support for adaptation of a species to its local environment. For example, in the study cited above with Themeda, when tested in controlled conditions, it showed latitudinal differences in daylength and vernalisation response that matched the conditions at their sites of origin (Fig. 1). Other species including both dicots and grasses with wide latitudinal distribution also show clinal variation in their critical daylength for flowering (Cumming 1969; Heide 1994; Aamlid et al. 2000).
Similar geographical differences in the vernalisation requirement for flowering are illustrated from studies with ecotypes of European thistle (Cirsium vulgare Suvi) grown in a common nursery garden. Compared with lines originating from the Mediterranean, there was a greater requirement for vernalisation in lines from colder more northerly sites as in Scandinavia or from high altitudes (Wesselingh et al. 1994).
Latitude also affects the balance between perennial and annual forms. At high latitudes perennial plants predominate (Körner 1995; and references in Heide and Gauslaa 1999) and they have a clear advantage because their overwintering photosynthetic tissues can contribute to early, rapid new spring growth and, so, to early flowering (Miller and Tieszen 1972). In such cases, as discussed above, perenniality of flowering is often achieved, by the sequential action over two seasons of two components of the environment.
A further adaptation for obtaining ‘pseudo’ perenniality involves change to flower development. Some high arctic species form floral primordia in one year, they overwinter and then, in the following spring, they rapidly flower (Heide et al. 1990; Heide 1992). A similar perenniality with its requirement for renewed floral induction occurs in some species from mid latitudes when they pass through a seasonal phase of excessive summer vegetative growth as in Fragaria (see Heide and Sønsteby 2007) or of winter bud dormancy as in chrysanthemum (T. Hisamatsu, pers. comm.).
Adaptation does not require identical mechanisms for regulation of flowering time at the same location across species or even for local variants within a species. For example, for two herbaceous perennial species of Lechenaultia originating from similar latitudes in Western Australia, one flowers in response to LD, the other to SD (King et al. 2008a). Similarly, within accessions of T. australis from northerly latitudes, one flowered in LD but other lines responded to SD (Fig. 1).
A further complexity is the presence of mechanisms that control flowering but lack adaptive value. As an example, Cerastium regelii Ostenf., an arctic-alpine species has a strong SD primary induction requirement for flowering in subsequent LD even though it will never experience SD conditions in a non-frozen condition in its natural environment (Heide et al. 1990). Like many other high-latitude dual induction species, it has an alternative, low temperature (vernalisation) pathway of primary induction, which is operative also under LD conditions (see above).
Species in their natural environment; some lessons from domesticated plants
‘Why if man can by patience select variations most useful to himself, should not nature …?’ (Darwin 1859, p. 469).
Changes in the flowering responses of crop species associated with their domestication raise interesting parallels with adaptation of flowering response to the natural environment. Consistently, introductions of foreign species into a new climate have been associated with selection for earliness of flowering and links have been suggested with photoperiod and vernalisation responses with all major crops including wheat, maize (Zea mays L.), rice and soybean (Glycine max L.) (see review in Evans 1993). Particularly for temperate cereals, there is extensive information on the genetic changes associated with differences in vernalisation response to winter cold (summarised in Trevaskis et al. 2007). However, caution is required in ascribing all changes in earliness to photoperiod and vernalisation responses. For rice and soybean, juvenility rather than photoperiodic response may have been the focus of this selection by humans (Evans 1993).
Early flowering with domestication of crop species is also evident as change in the effects of field temperatures on their development rate (phenology). The importance of temperature is clear from computations involving a heat sum approach in combination with daylength information. Such computations provide reasonable assessments of flowering times of annual crops (see references in Craufurd and Wheeler 2009) but little is known of associated genetic mechanisms. Studies of flowering in natural populations could benefit from this computational approach especially for the growing literature on effects of global warming on phenology.
Another predictor of the physical environment involves the photothermal quotient, which assesses the combined response to light intensity and temperature (i.e. radiation per unit area per degree day). For wheat, this quotient approximates well to the effect of latitude on wheat yield. For example, Rawson (1988) reported a very strong positive linear relationship between wheat yield at different maritime latitudes and the photothermal quotient for the period of 4 weeks pre-anthesis (r2 = 0.92). Obviously, greater seed production in nature should confer a selective advantage so response to the photothermal quotient could be usefully incorporated into studies of adaptation, at least in annual plants.
In other cases, domestication appears to have focussed not just on the initiation of flowering but, as well, on subsequent stem elongation. For example, temperate winter cereals avoid winter cold damage by delaying flowering and stem elongation until vernalised over winter. However in assessing the response of winter wheat lines developed in the 1980s in the UK, Evans (1987) found one line, Maris Templar, which was insensitive to vernalisation. It initiated flowers in winter but was able to withstand cold because its stem did not elongate until exposed to LD of spring and early summer.
Overall, change in flowering time has been critical in domestication of crop species and may involve several environmental adaptations in addition to those for daylength and vernalisation response. However, it is uncertain whether this evidence from studies of domestication provides a template for understanding evolution of wild species.
Has adaptation of flowering time to the environment been significant for evolution of a species?
Evidence for the adaptive nature of seasonal flowering does not address the question of evolutionary advantage and species fitness. More informative are observations of the adaptation of introduced species and of the environmental tolerance of a species when its historical range of adaptation is examined in reciprocal transplant studies.
Adaptation of an introduced species by the establishment of latitudinal flowering ecotypes can take place in surprisingly short periods of time. Thus, for the short day annual cocklebur (Xanthium strumarium L.), McMillan (1973) reported the establishment of latitudinally determined ecotypes in Australia in less than 150 years after the original introduction. In the short-long-day grass B. inermis 100 years after its introduction to Norway, Heide (1984) found changes in the critical photoperiods for both primary and secondary induction of flowering.
The value of two way transfers to uncover historic adaptation is illustrated in studies with a native Australian perennial shrub, Pimelea ferruginea Labill, a maritime species that grows over a 600 km latitudinal range in Western Australia on sand dunes within 100 m of the Indian Ocean. Large batches of vegetatively propagated lines of P. ferruginea were generated from cuttings from single plants collected over the full range of the natural distribution of the species.
In controlled environment studies, all plants flowered 4–6 weeks after transfer from a warm growth temperature to mild temperatures matching those of early winter (i.e. 15 to 18°C). Over many years of records, temperatures at the most northerly site (28°S) consistently track 4°C higher than those at the cooler most southerly site (31°S). Most interestingly, the ecotypes collected from the latitudinal extremes (28 v. 31°S) showed a matching 3−4°C difference in their temperature optimum for flowering (King et al. 1996).
Simultaneously, batches of the same plants were transferred back to their natural sites of origin. They flowered normally in response to cool winter conditions at their sites of origin. However, when reciprocally transplanted, a more southerly line with an 18°C optimum for flowering in the controlled environment studies, failed to flower in a nursery site at the 4°C warmer extreme of distribution of this species. Only the line with a 21−22°C optimum, which originated from this warmer site, flowered at this site. All lines flowered in a nursery at the cooler, more southerly site of origin. Thus, not only has this species of Pimelea developed a very sensitive thermoregulation of flowering, but, to survive at the warmer extreme of its range of natural distribution, its ‘thermoregulator’ tolerates higher temperatures characteristic of that site.
A further and reasonably demanding test of the adaptive advantage of mechanisms regulating seasonal flowering comes from studies with trans-global species. Darwin raised this question in relation to migration and isolation, which he suggested occurred following a period of glaciation. One such example involves bipolar species of the genus Carex. Seasonal regulation of flowering in such species is the same in the matching alpine environments of northerly and southerly alpine sites (Heide 2002). Furthermore, based on DNA fingerprinting studies (AFLP) there is no apparent genetic difference between northern and southern hemisphere representatives (Vollan et al. 2006). Thus, although geographically isolated, these Carex species have retained common mechanism(s) of flowering under matching environmental conditions in the two hemispheres.
In contrast, the cosmopolitan grass Poa annua L. illustrates the opposite situation, a micro-locational distinction in the choice of mechanisms for controlling flower initiation, Populations from two suburban parks in Canberra (35.3°S), separated by a distance of 7 km, exhibited very different flowering responses; one being a quantitative SD plant with no vernalisation response, the other flowering as a quantitative LD plant promoted by vernalisation (Heide 2001). Furthermore, while flowering responses were uniform within the latter population, the former segregated into early- and late-flowering types. This variation in earliness of flowering was shown to be genetically determined, and tests of selfed progenies indicated that individual plants collected from an area of 1 m2 were an aggregate of several largely homozygous lines with divergent flowering responses. Likewise, two lowland P. annua populations from southern Norway were both shown to be quantitative LD plants with no response to vernalisation, while two alpine snow bed populations from southern Norway and two high-latitude, subarctic populations were perennial, quantitative SD plants with an obligatory SD or vernalisation requirement for flowering. Thus, virtually any kind of photoperiodic and vernalisation response could be found among various populations of P. annua.
The versatility in choice of mechanisms regulating flowering of P. annua reflects the contrasting flowering responses of its putative ancestors, Poa infirma Kunth and Poa supine Schrader, the former being a summer annual, quantitative SD plant with no response to vernalisation, whereas the latter is a perennial quantitative LD plant with an obligatory SD or low temperature vernalisation requirement (Heide 2001). Such findings demonstrate the large environmental adaptation potential of allotetraploid species such as P. annua in which two different genomes have been brought together (Thompson and Lumarat 1992). Clearly, such a system is of great evolutionary significance and may explain why P. annua has spread around the world as a successful cosmopolitan weed with a variety of new ecotypes, each apparently well adapted to its new environment. On the other hand, the putative ancestors have remained restricted to limited and well separated areas of Europe.
Overall, at any one site, plants may flower at the same time but may achieve this by quite different responses to the environment. Furthermore, as discussed above, for survival the critical ‘floral’ regulatory processes may include juvenility, perenniality and seasonal dormancy in addition to processes directly controlling flower initiation. Thus, it is unwise to focus on a limited number of processes and often on single genes as the basis of selection; what is essentially a selfish gene concept is unlikely to capture the variety of responses and gene complexes involved in adapting flowering to season and site. We return to this issue below as part of an examination of genetic aspects of evolutionary fitness.
Genetic plasticity, selective advantage and strong heritability are central to environmental adaptation in the control of flowering
To argue evolutionary importance of a character requires evidence not only of broad latitudinal/geographical variation in flowering time but also of heritable responses. The foundation for much of our current knowledge of heritability of flowering time was provided by Laibach (1951) in his studies of flowering of the annual species A. thaliana. He established heritability of its flowering response to the seasonal environment. Subsequently, the extensive natural variation in its flowering response to daylength and vernalisation has been documented by Karlsson et al. (1993) and screening of induced mutations and natural populations of Arabidopsis for vernalisation- and/or daylength-induced flowering has indicated roles for several candidate genes (see review in Koornneef et al. 2003).
Which genes are important is now becoming clear from molecular/genetic studies. For example, the CO and FT proteins play a central role in LD flowering of Arabidopsis (reviewed in Turck et al. 2008) and whether in SD or LD response types, proteins similar to CO are found in a wide range of higher plants (Griffiths et al. 2003) and in a moss (Shimizu et al. 2004). Furthermore, a CO homologue in the unicellular green alga Chlamydomonous reinhardtii, could be reproductively significant as, when expressed in Arabidopsis, it complements a co mutant (Serrano et al. 2009). In parallel, response to seasonal environment is seen not only across higher plants but also in some mosses and liverworts (Lang 1965; Thomas and Vince Prue 1997).
Quantitative trait loci (QTL) screens with hybrid populations have confirmed the importance for flowering of vernalisation response of the two genes FLC and FRI (Johanson et al. 2000) and of other genes including the photoreceptor CRYPTOCHROME 2 (CRY2) (El-Assal et al. 2001). Subsequent analysis of allelic variants in 45 arbitrarily selected lines from natural populations of Arabidopsis showed that FRI accounted for up to a third of the heritable variation in vernalisation-induced flowering (Stinchcombe et al. 2004). In a larger grouping of 192 natural accessions where there was a wide range of expression of both FRI and FLC, Shindo et al. (2007) suggested a heritability of up to 70%. However, this may be a high estimate. While FRI could be identified as a gene of some importance in the variation of flowering time in European populations of Arabidopsis, there was no relationship between FRI and vernalisation in Asian lines (Shindo et al. 2007). Not only must other genes be important, but when Scarcelli et al. (2007) and Scarcelli and Kover (2009) used outbred populations developed by intermating 19 Arabidopsis accessions, they found that only 12% of the variation in flowering time was explained by FRI.
Interestingly, in the study of Scarcelli and Kover (2009), three generations of selection for early flowering in simulated spring conditions led to a reduced frequency of functional FRI alleles in these populations. Conversely, when vernalised in winter conditions the FRI limitation was removed and there was no large shift in the frequency of FRI alleles in the populations. These studies provide clear ‘proof of concept’, that FRI will contribute to fitness and, therefore, in natural populations it could have evolutionary importance. However, because FRI has pleiotropic effects, including changes in branching, the authors cautioned against concluding that selection optimises flowering time per se.
In wild populations, studies with the flowering time gene FT suggest its evolutionary importance for daylength responses of rice, an SD plant (Hagiwara et al. 2009). The genetic evidence for rice is compelling but little is known for other species although in wild poplar (Populus ssp.), latitudinal variation in daylength-regulated autumn bud dormancy is matched by daylength regulation of FT expression (Bohlenius et al. 2006). Perhaps for poplar, FT expression is simply a good marker of dormancy. Alternatively, FT acts in a pleiotropic way on a range of developmental process. This latter possibility appears more than likely as recent findings point to a pleiotropic effect of FLC acting on FT. Vernalising temperatures regulate flowering by suppressing FLC expression but also act in the same way during germination (Chiang et al. 2009). Thus, because FLC regulates FT, there is a broader role for FT in the regulation of plant development. The parallel between FLC regulation of FT (and other genes) both for flowering and germination (Chiang et al. 2009) not only highlights the importance of temperature in regulating FLC but also that the florigenic role for FT in flowering may involve chromatin remodelling as also for FLC.
Of further interest for field studies with Arabidopsis, temperature at germination may be an important determinant of flowering time (Wilczek et al. 2009). Thus, although FLC and the linked gene FRI may be important for flowering responses involving vernalisation, temperature sensing via FLC during germination may have been under most selective pressure in this annual species.
A second and very important issue for evolutionary studies with Arabidopsis is its ability to utilise alternative flowering pathways. Current genetic models list four pathways controlling its flowering; vernalisation, photoperiod, the gibberellin class of plant hormones and an autonomous path. Now, as discussed earlier, there is also evidence for a fifth path involving photosynthetic regulation of FT expression (King et al. 2008b).
Plasticity in the choice of floral pathways in Arabidopsis confers obvious evolutionary advantage but could also reduce selective pressure on any one pathway. Local genetic diversity within a species may in fact be quite extreme as in P. annua where different mechanisms for controlling flower initiation were evident in adjacent plants in a natural population (Heide 2001; see above). Conversely, selection pressure could be stronger where there is less plasticity in a species that flowers in response to one environmental input and by one genetic pathway.
Not only might gene/pathway flexibility complicate estimates of heritability, but it can mean that effective analysis of natural populations will require sampling of many plants in a population. A further qualification is that random genetic drift and not selection pressure may explain differences in the mechanism controlling flowering between adjacent plants or between geographical locations.
Random genetic drift may account for the identification of the CRY2 gene as a QTL for flowering time in an Arabidopsis line from the Cape Verde Islands but not from other lines (El-Assal et al. 2001). To approach this issue, linkage disequilibrium tests for non-random association of polymorphisms in a population allow a formal assessment of this problem of random genetic drift; any divergence, if strong, may indicate selection pressure (Ehrenreich and Purugganan 2006). In the future, in association with intense micro-locational sampling, genome wide mapping (GWA) could provide an even more powerful way to look at the association between genome and phenotype (Norberg and Weigel 2008).
To avoid the challenge of population demography and non-homogeneity, in testing for selective pressure on the FRI gene, Scarcelli et al. (2007) and Scarcelli and Kover (2009) used an intermated, homogeneous Arabidopsis population containing various FRI and FLC alleles (see above). Similarly, genetic homogeneity can be achieved by studying ‘directed gene’ responses in mutants and/or transgenic lines in a common background. For flowering, the first ‘directed gene’ effects have now been reported by Wilczek et al. (2009). Their field sowings of Arabidopsis lines across four sites in Europe over the year confirmed the natural control of flowering by genes considered as regulators of summer LD input (CO) and by winter vernalisation responses (FRI and FLC). However, unexpectedly, FLC repression of summer flowering could be overridden so substitution is possible by other unknown environmental inputs. Competitive advantage was not examined by Wilczek et al. (2009), so it is not possible to draw major conclusions about the evolutionary role of the two genes they used. The ‘directed gene’ approach also introduces a priori, assumptions about which genes determine evolutionary significance but further studies are warranted.
Overview
Darwin (1859) documented numerous characteristics of plants and animals that are beneficial for their survival. Here, we presented, in an evolutionary context, evidence supporting the use by plants of seasonal daylength, temperature and light intensity to time their flowering and seed set to avoid frost, heat or drought. Geographically, their choice of regulatory mechanisms has been selected to match their local environment.
A somewhat overlooked aspect of environmental regulation of flowering involves the dual requirements either for sequential exposure to different daylengths or to a combination of temperature and daylength and/or light intensity. Such dual responses greatly enhance the precision of seasonal timing of flowering and are evident in several species. While more difficult, simultaneous inheritance of two responses is not impossible especially if flowering is crucial for survival.
For the future, tests for selective advantage of any particular environmental input to flowering could involve reciprocal transplant experiments over a natural latitudinal cline (Mitchell-Olds et al. 2007). As an example, although the intensity of population sampling was limited, reciprocal transfers of Pimelea did establish differences in fitness that were related to their temperature responses for flowering and the temperature at their sites of origin (see discussion above and King et al. 1996). However, for other species there may be greater complexity both in environmental control of their flowering and in their natural environment. Thus, it may be difficult if not impossible to gain useful information from reciprocal transplantation studies.
Lacking our current genetic understanding, Darwin could not link inheritance and survival. Now modern molecular and genetic approaches allow tests of selective advantage of genes. One pair of floral regulatory genes, FRI/FLC, show a selective advantage for flowering in vernalising conditions but their low overall heritability (~12%) makes it likely that other flowering genes make substantial contributions to survival. A further issue is that studies of ‘floral’ genes have focussed on their effects on flowering time but various authors are beginning to consider their pleiotropic roles. Potentially how plants synchronise flowering to seasonal change could also involve regulation of germination, juvenility, dormancy or branching patterns.
Acknowledgements
We thank Drs L. T. Evans and E. S. Dennis for their helpful comments on this review.
Aamlid T,
Heide OM, Boelt B
(2000) Primary and secondary induction requirements for flowering of contrasting European varieties of Lolium perenne. Annals of Botany 86, 1087–1095.
| Crossref | GoogleScholarGoogle Scholar |

Beveridge CA
(2006) Axillary bud outgrowth: sending a message. Current Opinion in Plant Biology 9, 35–40.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Bohlenius H,
Huang T,
Charbonnel-Campaa L,
Brunner AM,
Jansson S,
Strauss SH, Nilsson O
(2006) CO/FT Regulatory module controls timing of flowering and seasonal growth cessation in trees. Science 312, 1040–1043.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Bünning E
(1936) Die endogene Tagesrhythmik als Grundlage der photoperiodischen Reaktion. Berichte der Deutschen Botanischen Gesellschaft 54, 590–607.
Chiang GCK,
Barua D,
Kramer EM,
Amasino RM, Donohue K
(2009) Major flowering time gene FLOWERING LOCUS C, regulates seed germination in Arabidopsis thaliana. Proceedings of the National Academy of Sciences of the United States of America 106, 11 661–11 666.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Craufurd PQ, Wheeler TR
(2009) Climate change and the flowering time of crops. Journal of Experimental Botany 60, 2529–2539.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Dore J
(1959) Response of rice to small differences in length of day. Nature 183, 413–414.
| Crossref | GoogleScholarGoogle Scholar |
Ehrenreich IM, Purugganan MD
(2006) The molecular genetic basis of plant adaptation. American Journal of Botany 93, 953–962.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
El-Assal S,
Alonso-Blanco C,
Peeters AJM,
Raz V, Koornneef MA
(2001) QTL for flowering time in Arabidopsis reveals a novel allele of CRY2. Nature Genetics 29, 435–440.
|
CAS |
Crossref |
PubMed |
Evans LT
(1987) Short day induction of inflorescence initiation in some winter wheat varieties. Australian Journal of Plant Physiology 14, 277–286.
| Crossref | GoogleScholarGoogle Scholar |
Evans LT, Knox RB
(1969) Environmental control of reproduction in Themeda australis. Australian Journal of Botany 17, 375–389.
| Crossref | GoogleScholarGoogle Scholar |
Garner WW, Allard HA
(1920) Effect of the relative length of day and night and other factors of the environment on growth and reproduction in plants. Journal of Agricultural Research 18, 553–603.
Gassner G
(1918) Beiträge zur physiologischen Characteristic sommer- und winterannueller Gewächse, insbesondere der Getreidepflanzen. Zeitschrift fur Botanik 10, 417–480.
Griffiths S,
Dunford RP,
Coupland G, Laurie DA
(2003) The evolution of CONSTANS-like gene families in barley, rice, and Arabidopsis. Plant Physiology 131, 1855–1867.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Hagiwara WE,
Uwatoko N,
Sasaki A,
Matsubara K,
Nagano H,
Onishi K, Sano Y
(2009) Diversification in flowering time due to tandem FT-like gene duplication, generating novel Mendelian factors in wild and cultivated rice. Molecular Ecology 18, 1537–1549.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Hayashi M,
Kawano S, Kuroiwa T
(1994) Rapid flower initiation in a desert annual, Pectis papposa Gray. Cytologia 59, 369–376.
Heide OM
(1977) Photoperiod and temperature interactions in growth and flowering of strawberry. Physiologia Plantarum 40, 21–26.
| Crossref | GoogleScholarGoogle Scholar |
Heide OM
(1984) Flowering requirements in Bromus inermis, a short-long-day plant. Physiologia Plantarum 62, 59–64.
| Crossref | GoogleScholarGoogle Scholar |
Heide OM
(1992) Flowering strategies of the high-arctic and high-alpine snow bed grass species Phippsia algida. Physiologia Plantarum 85, 606–610.
| Crossref | GoogleScholarGoogle Scholar |
Heide OM
(1994) Control of flowering and reproduction in temperate grasses. New Phytologist 128, 347–362.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
Heide OM
(1997) Environmental control of flowering in some Northern Carex species. Annals of Botany 79, 319–327.
| Crossref | GoogleScholarGoogle Scholar |
Heide OM
(2001) Flowering responses of contrasting ecotypes of Poa annua and their putative ancestors Poa infirma and Poa supina. Annals of Botany 87, 795–804.
| Crossref | GoogleScholarGoogle Scholar |
Heide OM
(2002) Climatic flowering requirements of bipolar sedges Carex spp. and the feasibility of their trans-equatorial migration by mountain-hopping. Oikos 99, 352–362.
| Crossref | GoogleScholarGoogle Scholar |
Heide OM
(2004) Dual induction rather than intermediate daylength response of flowering in Echinacea purpurea. Physiologia Plantarum 120, 298–302.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Heide OM, Gauslaa Y
(1999) Developmental strategies of Koenigia islandica, a high-arctic annual plant. Ecography 22, 637–642.
| Crossref | GoogleScholarGoogle Scholar |
Heide OM, Sønsteby A
(2007) Interactions of temperature and photoperiod in the control of flowering of latitudinal and altitudinal populations of wild strawberry (Fragaria vesca). Physiologia Plantarum 130, 280–289.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
Heide OM,
Pedersen K, Dahl E
(1990) Environmental control of flowering and morphology in the high-arctic Cerastium regelii, and the taxonomic status of C. jenisejense. Nordic Journal of Botany 10, 141–147.
| Crossref | GoogleScholarGoogle Scholar |
Jensen CS,
Salchert K, Nielsen KK
(2001) A Terminal Flower1-like gene from perennial ryegrass involved in floral transition and axillary meristem identity. Plant Physiology 125, 1517–1528.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Johanson U,
West J,
Lister C,
Michaels S,
Amasino R, Dean C
(2000) Molecular analysis of FRIGIDA, a major determinant of natural variation in Arabidopsis flowering time. Science 290, 344–347.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Karlsson BH,
Sills GR, Nienhuis J
(1993) Effects of photoperiod and vernilization on the number of leaves at flowering in Arabidopsis thaliana (Brassicaceae) ecotypes. American Journal of Botany 80, 646–648.
| Crossref | GoogleScholarGoogle Scholar |
King RW
(1998) Dual control of flower initiation and development by temperature and photoperiod in Hardenbergia violacea. Australian Journal of Botany 46, 65–74.
| Crossref | GoogleScholarGoogle Scholar |
King RW, Ben-Tal Y
(2001) A florigenic effect of sucrose in Fuchsia hybrida is blocked by gibberellin-induced assimilate competition. Plant Physiology 125, 488–496.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
King RW, Evans LT
(1991) Shoot apex sugars in relation to long-day induction of flowering in Lolium temulentum L. Australian Journal of Plant Physiology 18, 121–135.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
King RW,
Pate JS, Johnston J
(1996) Ecotypic differences in the flowering of Pimelea ferruginea (Thymelaeaceae) in response to cool temperatures. Australian Journal of Botany 44, 47–55.
| Crossref | GoogleScholarGoogle Scholar |
King RW,
Worall R, Dawson IA
(2008a) Diversity in environmental controls of flowering in Australian plants. Scientia Horticulturae 118, 161–167.
| Crossref | GoogleScholarGoogle Scholar |
King RW,
Hisamatsu T,
Goldschmidt EE, Blundell C
(2008b) Photosynthesis and far-red light independently regulate flowering but by increasing expression of FLOWERING LOCUS T (FT). Journal of Experimental Botany 59, 3811–3820.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Koornneef M,
Alonso–Blanco C, Vreugdenhil D
(2003) Naturally occurring genetic variation in Arabidopsis. Annual Review of Plant Biology 56, 141–172.
Laibach F
(1951) Ûber sommer- und winter-annuelle Rassen von Arabidopsis thaliana (L.) Heynh. Ein Beitrag zur Aetiologie der Blütenbildung. Beitrage Biologie Pflanzen 28, 173–210.
McMillan C
(1973) Photoperiodic adaptation in Xanthium populations introduced to Australia. Canadian Journal of Botany 51, 221–229.
| Crossref | GoogleScholarGoogle Scholar |
Miller PC, Tieszen LL
(1972) A preliminary model of processes affecting primary production in the arctic tundra. Arctic and Alpine Research 4, 1–18.
| Crossref | GoogleScholarGoogle Scholar |
Mitchell-Olds T,
Willis JH, Goldstein DB
(2007) Which evolutionary processes influence natural genetic variation for phenotypic traits? Nature Reviews. Genetics 8, 845–856.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Norberg M, Weigel D
(2008) Next generation genetics in plants. Nature 458, 720–723.
Pillitteri LJ,
Lovatt CJ, Walling LL
(2004) Isolation and characterization of a TERMINAL FLOWER homolog and its correlation with juvenility in citrus. Plant Physiology 135, 1540–1551.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Runkle ES,
Heins RD,
Cameron AC, Carlson WH
(2001) Photocontrol of flowering and stem extension of the intermediate-day plant Echinacea purpurum. Physiologia Plantarum 112, 433–441.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Scarcelli N, Kover PX
(2009) Standing genetic variation in FRIGIDA mediates experimental evolution of flowering time in Arabidopsis. Molecular Ecology 18, 2039–2049.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Scarcelli N,
Cheverud JM,
Schaal BA, Kover PX
(2007) Antagonistic pleiotropic effects reduce the potential adaptive value of the FRIGIDA locus. Proceedings of the National Academy of Sciences of the United States of America 104, 16 986–16 991.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Serrano G,
Herrera-Palau R,
Romero JM,
Serrano A,
Coupland G, Valverde F
(2009) Chlamydomonas CONSTANS and the evolution of plant photoperiodic signaling. Current Biology 19, 359–368.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Shimizu M,
Ichikawa K, Aoki S
(2004) Photoperiod-regulated expression of the PpCOL1 gene encoding a homolog of CO/COL proteins in the moss Physcomitrella patens. Biochemical and Biophysical Research Communications 324, 1296–1301.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Shindo C,
Bernasconi G, Hardtke CS
(2007) Natural genetic variation in Arabidopsis: tools, traits and prospects for evolutionary ecology. Annals of Botany 99, 1043–1054.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Sønsteby A, Heide OM
(2006) Dormancy relations and flowering of the strawberry cultivars Korona and Elsanta as influenced by photoperiod and temperature. Scientia Horticulturae 110, 57–67.
| Crossref | GoogleScholarGoogle Scholar |
Sønsteby A, Heide OM
(2007) Long-day control of flowering in everbearing strawberries. The Journal of Horticultural Science & Biotechnology 82, 875–884.
Sønsteby A, Heide OM
(2008) Flowering physiology of populations of Fragaria virginiana. The Journal of Horticultural Science & Biotechnology 83, 641–647.
Sønsteby A, Heide OM
(2009) Environmental control of flowering and runnering in three contrasting populations of Fragaria chiloenis. The Journal of Horticultural Science & Biotechnology 84, 267–272.
Stinchcombe JR,
Weinig C,
Ungerer M,
Olsen KM,
Mays C,
Halldorsdottir SS,
Purugganan MD, Schmitt J
(2004) A latitudinal cline in flowering time in Arabidopsis thaliana modulated by the flowering time gene FRIGIDA. Proceedings of the National Academy of Sciences of the United States of America 101, 4712–4717.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Thompson JD, Lumarat R
(1992) The evolutionary dynamics of polyploid plants: origins, establishment and persistence. Trends in Ecology & Evolution 7, 302–307.
| Crossref | GoogleScholarGoogle Scholar |
Trevaskis B,
Hemming MN,
Dennis ES, Peacock WJ
(2007) The molecular basis of vernalization-induced flowering in cereals. Trends in Plant Science 12, 352–357.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Turck F,
Fornara F, Coupland G
(2008) Regulation and identity of florigen: FLOWERING LOCUS T moves center stage. Annual Review of Plant Biology 59, 573–594.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Vollan K,
Heide OM,
Lye KA, Heun M
(2006) Genetic variation, taxonomy and mountain-hopping of four bipolar Carex species (Cyperaceae) analysed by AFLP fingerprinting. Australian Journal of Botany 54, 305–313.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
Wang R,
Farrona S,
Vincent C,
Joecker A,
Schoof H,
Turck F,
Alonso-Blanco C,
Coupland G, Albani M
(2009) PEP1 regulates perennial flowering in Arabis alpina. Nature 459, 423–427.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Wesselingh RA,
Klinkhamer PGL,
de Jong TJ, Schlatinann EGM
(1994) A latitudinal cline in vernalization requirement in Cirsium vulgare. Ecography 17, 272–277.
| Crossref | GoogleScholarGoogle Scholar |
Wilczek AM,
Roe JL,
Knapp MC,
Cooper MD, Lopez-Gallego C , et al.
(2009) Effects of genetic perturbation on seasonal life history plasticity. Science 323, 930–934.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Wilkie JD,
Sedgley M, Olesen T
(2008) Regulation of floral initiation in horticultural trees. Journal of Experimental Botany 59, 3215–3228.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |