

Infection with an asymptomatic virus in rice results in a delayed drought response
Jaymee R. Encabo A B C , Reena Jesusa A. Macalalad-Cabral A , Jerlie Mhay K. Matres A , Sapphire Charlene Thea P. Coronejo A , Gilda B. Jonson A , Yuji Kishima B , Amelia Henry D E and Il-Ryong Choi A EA Rice Breeding Platform, International Rice Research Institute, Pili Drive, Los Baños, Laguna, 4031 Philippines.
B Laboratory of Plant Breeding, Research Faculty of Agriculture, Hokkaido University, Sapporo, 060-8589, Japan.
C Microbiology Division, Institute of Biological Sciences, University of the Philippines Los Baños, Los Baños, Laguna, 4031, Philippines.
D Strategic Innovation Platform, International Rice Research Institute, Pili Drive, Los Baños, Laguna, 4031, Philippines.
E Corresponding authors. Email: a.henry@irri.org; ichoi51@hotmail.com
Functional Plant Biology 47(3) 239-249 https://doi.org/10.1071/FP19241
Submitted: 22 August 2019 Accepted: 1 November 2019 Published: 12 February 2020
Journal Compilation © CSIRO 2020 Open Access CC BY
Abstract
Infection of viruses in plants often modifies plant responses to biotic and abiotic stresses. In the present study we examined the effects of Rice tungro spherical virus (RTSV) infection on drought response in rice. RTSV infection delayed the onset of leaf rolling by 1–2 days. During the delay in drought response, plants infected with RTSV showed higher stomatal conductance and less negative leaf water potential under drought than those of uninfected plants, indicating that RTSV-infected leaves were more hydrated. Other growth and physiological traits of plants under drought were not altered by infection with RTSV. An expression analysis of genes for drought response-related transcription factors showed that the expression of OsNAC6 and OsDREB2a was less activated by drought in RTSV-infected plants than in uninfected plants, further suggesting improved water status of the plants due to RTSV infection. RTSV accumulated more in plants under drought than in well-watered plants, indicating the increased susceptibility of rice plants to RTSV infection by drought. Collectively, these results indicated that infection with RTSV can transiently mitigate the influence of drought stress on rice plants by increasing leaf hydration, while drought increased the susceptibility of rice plants to RTSV.
Additional keywords: DREB2, Rice tungro spherical virus.
Introduction
Plants are often exposed simultaneously to multiple biotic and abiotic stresses. Among the interactions of plants with various biotic and abiotic stress combinations, the responses of plants under simultaneous stresses from pathogens and drought have been most well studied (Mayek-Pérez et al. 2002; McElrone et al. 2003; Xu et al. 2008; Ramegowda et al. 2013; Davis et al. 2015; Gupta et al. 2016). The outcome of such interactions varies depending on drought severity and pathogens, the order of occurrence of each stressor, and the intensity and duration of exposure (Gupta et al. 2016). One of the typical outcomes from combined drought and pathogen stresses is increased susceptibility of plants to pathogens. Exposure of common bean and sorghum to drought stress resulted in increased susceptibility to the charcoal rot fungus, Macrophomina phaseolina (Diourte et al. 1995; Mayek-Pérez et al. 2002). Similarly, simultaneous exposure of Virginia creeper (Parthenocissus quinquefolia (L.) Planch. plants to drought stress and Xylella fastidiosa infection resulted in more severe leaf scorch symptoms, and greater reductions in leaf area and shoot length (McElrone et al. 2001, 2003; McElrone and Forseth 2004). Exposure to drought stress resulted in higher infection levels of an avirulent strain of Pseudomonas syringae in Arabidopsis thaliana (L.) Heynh. (Mohr and Cahill 2003). Another typical outcome from combined drought and pathogen stresses is improved tolerance of plants to drought. A. thaliana plants infected with P. syringae showed increased stomatal closure under drought, which resulted in reduced water loss from the infected plant (Goel et al. 2008). In another study, infection with Verticillium longisporum in A. thaliana induced de novo xylem formation resulting in enhanced water storage capacity (Reusche et al. 2014). Xu et al. (2008) demonstrated that infection of various plant species with Brome mosaic virus (BMV), Cucumber mosaic virus (CMV) or Tobacco mosaic virus (TMV) induced accumulation of osmoprotectants and antioxidants, and delayed the appearance of drought symptoms in the plants.
Rice tungro spherical virus (RTSV) and Rice tungro bacilliform virus (RTBV) cause rice tungro disease (RTD) (Hibino et al. 1978). RTSV is the type member of the genus Waikavirus in the family Secoviridae (Sanfaçon et al. 2011). RTSV has a positive-sense, single-stranded RNA genome of ~12-kb in a polyhedral particle (Shen et al. 1993; Hull 1996). In RTD, RTSV plays the role of a helper virus for insect transmission of RTBV, and also enhances the yellow-orange leaf discoloration and stunting symptoms caused by RTBV (Hibino 1983; Hibino et al. 1987). RTSV itself can be considered as a latent (asymptomatic) virus since infection with RTSV alone does not cause noticeable symptoms in most Asian rice (Oryza sativa L.) plants (Hibino et al. 1978), although RTSV causes significant stunting in some rice cultivars such as Reiho (Shinkai 1977), and in some African rice (Oryza glaberrima Steud.) plants (Cabauatan 1993; Budot et al. 2014). Transcriptome analyses of rice plants infected with RTSV (Encabo et al. 2009; Satoh et al. 2013) revealed the significant changes in expression levels of genes for transcription factors belonging to DREB, SNAC, WRKY, bZIP and AP2-EREBP families that had been previously reported to respond to drought stress (Ohnishi et al. 2005; Hu et al. 2006; Nakashima et al. 2007; Cui et al. 2011; Rachmat et al. 2014). Based on these observations, we hypothesised that RTSV infection changes the response of rice plants to drought stress.
In the present study, we examined the physiological, growth, and gene expression responses in rice plants infected with RTSV under varying soil moisture conditions in order to determine the effect of infection with RTSV on plant response to drought. Our results showed that infection with RTSV can transiently mitigate the influence of drought stress on rice plants by increasing leaf hydration, while drought increased the susceptibility of rice plants to RTSV.
Materials and methods
Plant materials and experimental conditions
Taichung Native 1 (TN1), an indica rice cultivar susceptible to RTSV, was used in this study. PVC cylinders of 21 cm in height and 10 cm in diameter were filled with 2.0 kg of dry and sieved soil. One 7-day-old seedling was planted in each cylinder when the soil moisture level was at 100% of field capacity. The plants were allowed to grow in flooded conditions for two weeks before starting a drought stress treatment. Three separate experiments comprised of eight separate trials were conducted: (1) physiological evaluation of response to RTSV and drought under non-shaded conditions, (2) physiological evaluation of response to RTSV and drought under shaded conditions, and (3) gene expression analysis of response to RTSV and drought under non-shaded conditions. In the experiment in which shading was used, drought-treated plants were placed under a shade structure of a metal frame (156 × 61 × 89 cm) covered with black fabric. All experiments were conducted under greenhouse conditions at the International Rice Research Institute, with daytime temperature ranging from 24.7 to 29.0°C, and RH ranging from 70.0 to 85.8%.
Virus inoculation and drought stress application
In all experiments, plants were inoculated with RTSV before initiating the well-watered and drought stress treatments (Fig. 1). At 14 days after sowing (DAS), the plants were inoculated with RTSV by the tube method (Cabauatan et al. 1995). RTSV strain A that had been maintained in variety TN1 was used as the inoculum. Green leafhoppers (GLH, Nephotettix virescens) that are known to transmit RTSV were fed on RTSV-infected TN1 plants for 3 days to acquire RTSV. Five RTSV-viruliferous GLH were transferred to, and allowed to feed on, each healthy plant for 24 h to transmit RTSV. Mock-inoculated plants were prepared likewise by feeding five virus-free GLH on each plant for 24 h. After RTSV was inoculated, the soil in each cylinder was kept flooded for 7 days. Infection of plants with RTSV was confirmed at 28 DAS (corresponding to 14 days post inoculation, DPI) by an enzyme-linked immunosorbent assay using an antibody for RTSV (Bajet et al. 1985; Encabo et al. 2009) (Fig. 1).

|
At 21 DAS (7 DPI), cylinders with single plants were covered using plastic sheets around the base of the plants to minimise non-transpirational water loss, and were randomly assigned to either the well-watered or the drought stress treatments (Fig. 1). The soil moisture in the drought stress treatment was gradually brought down to targeted soil moisture levels by weighing and re-watering the cylinders at 09:00 to 10:00 hours every 2–3 days for 3 weeks. The soil moisture of cylinders for the drought stress treatment was decreased by 9% at each weighing date, while plants of the well-watered treatment were kept flooded throughout the experiments. For all experiments, five replicates of the four treatments (RTSV-infected and well-watered, RTSV-infected and drought stressed, uninfected and well-watered, and uninfected and drought stressed) were included, and each experiment was repeated three times as separate trials.
Measurements of physiological and growth traits
Leaf traits
Measurements of leaf traits were conducted between 33 and 38 DAS (corresponding to 12 to 17 days after drought initiation) (Fig. 1). Leaf rolling due to drought stress was evaluated at 09:00 to 10:00 hours using the 0–9 scale according to the standard evaluation system for rice (IRRI 2002). Apparent leaf area was measured by the digital imaging method described by Kijoji et al. (2013). Digital images were analysed using ImageJ software (ImageJ 1.43; Java 1.6.0_10; Wayne Rasband, Abramoff et al. 2004). Leaf water potential (LWP) was determined as the pressure necessary to force sap from the cut end of the leaf (Plant Water Status Console, Soilmoisture Equipment Corp.) using compressed N2 gas. Leaf osmotic potential (LOP) was measured by collecting the youngest fully-expanded leaf, freezing tissue inside a 5-mL syringe, pressing the sap from the thawed sample and measuring 10 µL with a vapour pressure osmometer (Vapro model 5520, Wescor). Osmolality (c; mmol kg–1) was converted to leaf osmotic potential (MPa) using the formula: MPa = –c × 2.58 × 10–3.
Photosynthetic parameters
Net photosynthetic rate (Pn, µmol m–2 s–1) and stomatal conductance (gs, mmol m–2 s–1) were measured using a LI-COR 6400 portable photosynthesis system (LI-COR Biosciences).
Water uptake
Water uptake rates were determined following the method described by Kijoji et al. (2013). Cylinders were weighed three times per week following drought stress initiation. Water uptake rates were calculated as ((the amount of water loss from the cylinder/the number of days between two successive measurements) – any water added to maintain the targeted gradual drydown). Water uptake (kg day–1) between two weighing dates was divided by the estimated leaf area (cm2) to calculate water uptake rates normalised for plant size. Water use efficiency was estimated as the ratio of the total grams of shoot biomass accumulated over the drought stress period to the amount of water transpired by the plant in kilograms during the entire period of the drought stress.
Plant growth
Plant height was measured before harvesting the shoots of individual plants at 40 DAS (Fig. 1). The dry weights of shoots were determined after 72 h of oven drying at 70°C.
Data presented are the average with the standard error of at least three replicates. The results were analysed separately by trial because a significant effect of trial was observed. Means were compared using Student’s t-test (α = 0.05), except for leaf rolling score in which the treatments were compared by Friedman’s test for ordinal data.
Quantification of RTSV accumulation
Total RNA were isolated from the second-youngest leaves of individual plants using TRIzol (Invitrogen) as per manufacturer’s instructions. The standard curve for RTSV was prepared using an in vitro transcript corresponding to the NTP-Q2 region of the RTSV genome. First-strand cDNA from the RTSV genome was synthesised from 1 µg of RNA samples combined from those of three independent plants of the same treatment using an oligonucleotide primer specific to the RTSV NTP-Q2 region (Primer RTSV RT R in Table S1, available as Supplementary Material to this paper) (Sharma and Dasgupta 2012) and Superscript III synthesis system (Invitrogen) as per the manufacturer’s instructions. A double-stranded cDNA segment of 1.6 kbp corresponding to the NTP-Q2 region of the RTSV genome (nucleotide positions 6469–8110 of RTSV-Phil A, NC.001632) was synthesised using RTSV 5F and RTSV 5R (Table S1) (Sharma and Dasgupta 2012) and cloned in pGEM T-easy vector. The resultant plasmid was linearised with SalI and used to synthesise the NTP-Q2 in vitro transcript using the Riboprobe in vitro transcription system with T7 RNA polymerase (Promega) according to the manufacturer’s protocol. The copy numbers of the in vitro transcript were calculated as ((ng of transcript) × Avogadro constant (6.023 × 1023 molecules mol–1))/(1641 (length of transcript) × 330 g mol–1) × 109 (ng/g)). Serial dilutions of the NTP-Q2 in vitro transcript were prepared to give a range from 106 to 1010 copies per 2 µL, and were subjected to a quantitative reverse transcription polymerase chain reaction (qRT-PCR). The standard curve for the RTSV genomes was constructed by plotting mean threshold cycle (CT) values against the logarithm of the in vitro transcript copy number in a linear regression curve.
The accumulation of RTSV in plants was evaluated by a qRT-PCR for the NTP-Q2 segment of the RTSV genome by a method modified from those described in Sharma and Dasgupta (2012) and Fronhoffs et al. (2002). First-strand cDNA from the RTSV genome was synthesised as described above. qPCR amplifications were carried out using the ABI Step-One Plus Real-Time PCR system (Applied Biosystems). Each reaction mix consisted of 1× Power SYBR PCR master mix (Applied Biosystems), first-strand cDNA synthesised from 1 μg of RNA, and 200 nM of each primer RTSV RT F and RTSV RT R (Table S1). The cycling profile was 95°C for 10 min, followed by 40 cycles of 95°C for 15 s and 60°C for 60 s. The CT values obtained were used to determine the quantity of RTSV genome based on the standard curve. The experiment was repeated three times with independent samples. Differences in the number of RTSV genomes in plants were examined by Student’s t-test (α = 0.05).
Gene expression analysis
Drought-responsive transcription factor genes examined for their expression by qRT-PCR were selected based on their RTSV-induced expression patterns previously revealed by transcriptome analyses for TN1 infected with RTSV (Encabo et al. 2009; Satoh et al. 2013). The locus IDs of selected genes were based on the Rice Pseudomolecules, release 7 available at the Rice Genome Annotation Project database (http://rice.plantbiology.msu.edu/, accessed 28 October 2019). The annotations of selected genes referred to information available at the databases of Rice Genome Annotation Project, National Centre for Biotechnology Information and Rice Database Oryzabase (http://shigen.nig.ac.jp/rice/oryzabase/, accessed 28 October 2019). Total RNA was isolated from plants as described above. First-strand cDNA was synthesised using 1 µg of RNA samples combined from those of three independent plants of the same treatment using an oligo(dT)20 primer. qPCR was performed as described above using gene-specific primers (Table S2). Expression values were normalised to the expression of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) gene (LOC_Os04g40950) as the internal control. Quantification of the relative changes in gene expression was performed using the 2–ΔΔCT method (Schmittgen and Livak 2008). Data from three technical replicates and three biological replicates per treatment and gene were used for the analysis. The differences in the expression changes of a gene under different conditions were examined by an analysis of variance followed by a least significant difference test (α = 0.05).
Results
Effect of RTSV infection on leaf traits under drought
To examine the effects of RTSV infection on the response of plants to drought, uninfected and RTSV-infected plants were grown under the drought condition and the timing of leaf rolling in the plants were compared. Cumulative water uptake of plants was recorded during the three-week observation period to ensure controlled dry-down for the drought stress treatment (Fig. 2). Both uninfected and RTSV-infected plants started showing leaf rolling on the same day between 33 and 38 DAS when the soil moisture level became below 30% of field capacity (Fig. 1), but leaf rolling in uninfected plants started two to three hours earlier than in RTSV-infected plants. At 09:00 hours most uninfected plants showed severe leaf rolling (average leaf rolling score of 8.1), but noticeable leaf rolling was not observed in RTSV-infected plants (Fig. 3a–c). The apparent leaf area of uninfected plants at the time when leaf rolling was observed (09:00 to 10:00 hours) was significantly smaller than that of RTSV-infected plants (Fig. 3d).

|

|
The leaf rolling progressed rapidly during the mornings and the difference in the initiation of leaf rolling between RTSV-infected and uninfected plants were only observed for two to three hours. The appearance of drought-induced symptoms can be affected by many factors such as light intensity (Holmgren et al. 2012). To confirm the observed difference in drought response between uninfected and RTSV-infected plants is indeed associated with RTSV infection, we repeated the experiment under shaded conditions. Shading resulted in larger differences in the timings of leaf rolling appearance between RTSV-infected and uninfected plants (Fig. 4). The appearance and progression of leaf rolling were delayed in RTSV-infected plants compared with those in mock-inoculated plants. When kept shaded, leaf rolling appeared in uninfected plants 1.36 ± 0.29 (average ± s.e.) days earlier than in RTSV-infected plants. Tightly rolled leaves corresponding to a leaf rolling score of 9 were observed in uninfected plants 1.87 ± 0.07 days earlier than in RTSV-infected plants. By the end of the seven-day observation period under the shaded condition, uninfected plants exhibited more wilted tips and tightly rolled leaves compared with RTSV-infected plants.

|
Stomatal conductance (gs) and net photosynthetic rate (Pn) decreased in response to drought stress in both RTSV-infected and uninfected plants (Fig. 5). Under the well-watered condition, gs and Pn in RTSV-infected and uninfected plants were virtually the same (Fig. 5). When measured during the period of greatest visual differences in leaf rolling between RTSV-infected and uninfected plants in the drought treatment, gs and Pn were generally higher in RTSV-infected plants than in uninfected plants (Fig. 5). Moreover, RTSV-infected plants showed less negative LWP than uninfected plants under the drought condition, but not under well-watered conditions (Table S3). Overall, these results indicate that the appearance and progression of leaf rolling due to drought stress were delayed by infection with RTSV, and that the delayed drought symptoms appear to be associated with increased leaf hydration by RTSV infection.

|
Effect of RTSV infection on water uptake and plant growth under drought
Water uptake rate was not affected by RTSV infection under both well-watered and drought conditions (Table S3). Likewise, no consistent differences in cumulative water uptake (Fig. 2) and water use efficiency (Table S3) were observed between uninfected and RTSV-infected plants under both well-watered and drought conditions. The height and shoot mass of uninfected plants and those of RTSV-infected plants were not significantly different under both well-watered and drought conditions (Table S3). These results indicate that infection with RTSV was asymptomatic in plants, and had no evident effect on water uptake of plants regardless of the water conditions.
Effect of drought on RTSV replication
The accumulation of RTSV in plants under the well-watered and the drought conditions was examined to determine the effect of water conditions on RTSV accumulation. At 14 DPI (28 DAS), the RTSV level in well-watered plants was not significantly different from that in plants under the drought condition (soil moisture level of ~70% of field capacity) (Fig. 6). However, at 21 DPI, RTSV accumulated significantly less in well-watered plants than in plants under severe drought stress (soil moisture level below 30% of field capacity) (Fig. 6). At 21 DPI (35 DAS), drought-stressed plants were found to contain ~65% more RTSV than the well-watered plants, indicating an increased susceptibility of drought-stressed plants to RTSV infection.
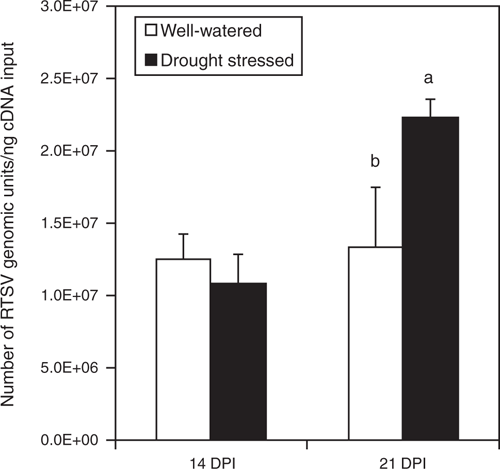
|
Changes in the expression of drought-responsive transcription factor genes by drought and RTSV
From the study of physiological parameters, we observed that leaf water status (gs, Pn, and LWP) was significantly influenced by RTSV infection under drought (Fig. 5, Table S3). It has been well documented that regulation of various transcription factor genes is closely associated with drought stress (Ohnishi et al. 2005; Hu et al. 2006; Nakashima et al. 2007; Cui et al. 2011; Rachmat et al. 2014). Therefore, we selected five transcription factor genes that had been reported to be involved in drought stress response, in order to examine the effects of drought-RTSV interaction on their expression. The expression patterns of genes for dehydration-responsive element-binding protein 2A (OsDREB2A, LOC_Os01g07120) (Chen et al. 2008; Cui et al. 2011), b-ZIP transcription factor 23 (OsbZIP23, LOC_Os02g52780) (Xiang et al. 2008), stress-induced transcription factor NAC1 (OsSNAC1, LOC_Os03g60080.1) (Hu et al. 2006), NAC domain containing protein 6 (OsNAC6, LOC_Os01g66120) (Ohnishi et al. 2005; Nakashima et al. 2007; Rachmat et al. 2014), and ethylene response factor 20 (OsERF20, LOC_Os02g45420) (Fukao et al. 2011) were examined in rice plants in four different treatments: (1) uninfected and well-watered (as the control condition), (2) uninfected and drought stressed, (3) RTSV-infected and well-watered and (4) RTSV-infected and drought stressed. Leaf samples were collected at two time points: the first time point was 14 DPI with RTSV (28 DAS and 7 days after initiation of the drought treatment when soil moisture was ~70% of field capacity). The second time point was 21 DPI with RTSV (35 DAS and 14 days after initiation of the drought treatment when soil moisture was lower than 30% of field capacity).
At 14 DPI, the levels of activation for OsNAC6 (Fig. 7b), OsDREB2a (Fig. 7d), and OsbZIP23 (Fig. 7e) were significantly higher in uninfected, drought-stressed plants than in RTSV-infected, well-watered plants. The level of activation for OsNAC6 in RTSV-infected, drought-stressed plants was significantly lower than that in uninfected, drought-stressed plants (Fig. 7b), indicating that the activation of OsNAC6 by the mild drought stress (soil moisture ~70% of field capacity) was attenuated by RTSV infection.

|
At 21 DPI, all five genes were more activated by the drought stress than by RTSV infection (Fig. 7). The levels of activation for the genes other than OsDREB2a by the drought stress were not affected by RTSV infection (Fig. 7a–c, e). The level of activation for OsDREB2a by the drought stress was affected by RTSV infection, showing less activation of OsDREB2a in RTSV-infected, drought-stressed plants than in uninfected, drought stressed plants (Fig. 7d), indicating that the activation of OsDREB2a by the severe drought stress (soil moisture lower than 30% of field capacity) was attenuated by RTSV infection. These results suggest drought-responsive activation of transcription factor genes such as OsNAC6 and OsDREB2a can be suppressed by RTSV infection.
Discussion
In the present study we observed that RTSV transiently mitigated the influence of drought stress in rice, as evidenced by delayed leaf rolling, higher stomatal conductance, and less negative LWP. As an asymptomatic virus, RTSV may therefore act as a conditional mutualist to the rice plant in the case of drought stress. Infection with RTSV delayed the onset and progression of leaf rolling in rice (Figs 3, 4). Similar observations were previously reported in Nicotiana benthamiana plants infected with BMV, CMV, or TMV (Xu et al. 2008) and in wheat plants infected with Barley yellow dwarf virus (Davis et al. 2015). The results in this study and those from the previous studies clearly indicate that virus infection can alter plant–water relations under drought.
RTSV infection significantly increased leaf photosynthetic parameters (gs and Pn) under drought (Fig. 5). The higher Pn in RTSV-infected plants under the drought condition might have been a result of the increased gs (Pantaleo et al. 2016). Moreover, plants infected with RTSV consistently showed less negative LWP under the drought condition (Table S3). The less negative LWP, which is indicative of better leaf hydration, might have resulted in the observed phenomena of delayed leaf rolling and prolonged leaf turgidity in RTSV-infected plants. Consistent with our results, previous studies have shown that virus-infected plants exhibited higher LWP than control plants under drought (Hall and Loomis 1972; Lindsey and Gudauskas 1975; Grimmer et al. 2012; Davis et al. 2015). From these observations, the predominant mode of drought response in RTSV-infected plants is likely to be water potential-dependent stomatal control as previously reported (Brodribb and McAdam 2013; El Aou‐ouad et al. 2017). Thus, RTSV infection can improve the leaf water status of rice plants under drought, albeit transiently.
RTSV accumulated more in drought-stressed plants than in well-watered plants (Fig. 6), indicating that the drought stress might have increased the susceptibility of rice plants to RTSV. Combined biotic and abiotic stresses in plants can weaken plant defence mechanisms, resulting in enhanced susceptibility to biotic stress (Goel et al. 2008; Mittler and Blumwald 2010). For example, when plants are exposed to a combination of abiotic and virus stresses, they may deactivate defence responses to virus infection by prioritising the defence systems to abiotic stresses (Prasch and Sonnewald 2013). Similarly, the increase in RTSV accumulation under the severe drought condition observed in this study might have been due to prioritised defence systems to the more damaging effects of drought when they were exposed simultaneously to RTSV infection and drought stress.
Many transcription factors associated with drought stress have been identified in rice. These include the families of DREB (Chen et al. 2008; Cui et al. 2011; Mallikarjuna et al. 2011), NAC (Hu et al. 2006; Nakashima et al. 2007), bZIP (Uno et al. 2000; Xiang et al. 2008), WRKY (Wu et al. 2009) and ERF (Fukao et al. 2011). In the present study we examined the expression of five transcription factor genes (SNAC1, OsNAC6, OsERF20, OsbZIP23 and OsDREB2A) known to be induced by drought stress to examine the effect of RTSV infection on the drought-inducible expression of the transcription factor genes. Consistent with previous reports, the expression of all five genes was induced by drought stress (Fig. 7). However, the levels of activation of OsNAC6 and OsDREB2A during the drought stress treatment were significantly decreased in the presence RTSV (Fig. 7b, d). Therefore, the decreased activation of the two genes may have been associated with the drought-induced symptoms mitigated by RTSV infection.
Along with drought-responsive transcription factor genes, plant microRNAs (miRNAs), a class of non-coding regulatory single strand RNAs of 20–27 nucleotides in length, play an important role in the regulation of plant response to abiotic and biotic stresses. Many miRNA target transcription factors that regulate tolerance to abiotic stresses such as drought in plants (Jatan and Lata 2019). Several miRNAs have been identified to target drought-responsive transcription factor genes such as that for bZIP in rice (Zhou et al. 2010) and those for NAC proteins in grapevines (Pantaleo et al. 2016), which further regulate gene expression involved in drought response. In our experiments, OsNAC6 was induced by drought stress but OsNAC6 activation was less responsive to drought stress in the presence of RTSV. Taken together, it is highly likely that some interactions between miRNAs and drought-responsive transcription factor genes might have resulted in the observed physiological changes induced by RTSV infection under drought.
We observed that the combination of RTSV infection and drought stress led to contrasting impacts on plants. Virus-infected plants exhibited a delayed response to drought as a result of increased leaf hydration whereas drought stress increased the susceptibility of rice plants to RTSV infection, perhaps as a consequence of weakened basal defence to RTSV by drought stress. Our findings provide useful information in understanding virus-induced alterations of plant–water relations that may provide conditionally beneficial outcomes under combined stress conditions. However, our study investigated only short-term physiological and gene expression responses of virus-infected and drought-stressed rice plants. Therefore, further field studies and evaluation of plant growth performance such as yield under drought may be able to assess the long-term effects of infection with an asymptomatic virus under combinatorial stress.
Conflicts of interest
The authors declare no conflicts of interest.
Acknowledgements
This work was supported by grants from the Global Rice Science Partnership (GRISP), Temperate Rice Research Consortium of IRRI and Rural Development Administration of Korea and the Bill and Melinda Gates Foundation STRASA project (Stress Tolerant Rice for Africa and South Asia). We thank E Baguioso and C Lantican for assistance in preparing the virus source and insects for inoculation; and also D Morales, N Sadiasa, A Reyes and M Quintana for assistance with the gas exchange and leaf measurements.
References
Abramoff MD, Magalhaes PJ, Ram SJ (2004) Image Processing with ImageJ. Biophotonics International 11, 36–42.Bajet NB, Daquioag RD, Hibino H (1985) Enzyme-linked immunosorbent assay to diagnose rice tungro. Journal of Plant Protection in the Tropics 2, 125–129.
Brodribb TJ, McAdam SA (2013) Abscisic acid mediates a divergence in the drought response of two conifers. Plant Physiology 162, 1370–1377.
| Abscisic acid mediates a divergence in the drought response of two conifers.Crossref | GoogleScholarGoogle Scholar | 23709665PubMed |
Budot BO, Encabo JR, Ambita IDV, Atienza-Grande GA, Satoh K, Kondoh H, Ulat VJ, Mauleon R, Kikuchi S, Choi I-R (2014) Suppression of cell wall-related genes associated with stunting of Oryza glaberrima infected with Rice tungro spherical virus. Frontiers in Microbiology 5, 26
| Suppression of cell wall-related genes associated with stunting of Oryza glaberrima infected with Rice tungro spherical virus.Crossref | GoogleScholarGoogle Scholar | 24550897PubMed |
Cabauatan PQ, Kobayashi N, Ikeda R, Koganezawa H (1993) Oryza glaberrima: an indicator plant for Rice tungro spherical virus. International Journal of Pest Management 39, 273–276.
| Oryza glaberrima: an indicator plant for Rice tungro spherical virus.Crossref | GoogleScholarGoogle Scholar |
Cabauatan PQ, Cabunagan RC, Koganezawa H (1995) Biological variants of rice tungro viruses in the Philippines. Phytopathology 85, 77–81.
| Biological variants of rice tungro viruses in the Philippines.Crossref | GoogleScholarGoogle Scholar |
Chen JQ, Meng XP, Zhang Y, Xia M, Wang XP (2008) Over-expression of OsDREB genes lead to enhanced drought tolerance in rice. Biotechnology Letters 30, 2191–2198.
| Over-expression of OsDREB genes lead to enhanced drought tolerance in rice.Crossref | GoogleScholarGoogle Scholar | 18779926PubMed |
Cui M, Zhang W, Zhang Q, Xu Z, Zhu Z, Duan F, Wu R (2011) Induced over-expression of the transcription factor OsDREB2A improves drought tolerance in rice. Plant Physiology and Biochemistry 49, 1384–1391.
| Induced over-expression of the transcription factor OsDREB2A improves drought tolerance in rice.Crossref | GoogleScholarGoogle Scholar | 22078375PubMed |
Davis TS, Bosque-Perez N, Foote NE, Magney T, Eigenbrode SD (2015) Environmentally dependent host-pathogen and vector-pathogen interactions in the Barley yellow dwarf virus pathosystem. Journal of Applied Ecology 52, 1392–1401.
| Environmentally dependent host-pathogen and vector-pathogen interactions in the Barley yellow dwarf virus pathosystem.Crossref | GoogleScholarGoogle Scholar |
Diourte M, Starr JL, Jeger MJ, Stack JP, Rosenow DT (1995) Charcoal rot (Macrophomina phaseolina) resistance and the effects of water stress on disease development in sorghum. Plant Pathology 44, 196–202.
| Charcoal rot (Macrophomina phaseolina) resistance and the effects of water stress on disease development in sorghum.Crossref | GoogleScholarGoogle Scholar |
El Aou‐ouad H, Pou A, Tomás M, Montero R, Ribas‐Carbo M, Medrano H, Bota J (2017) Combined effect of virus infection and water stress on water flow and water economy in grapevines. Physiologia Plantarum 160, 171–184.
| Combined effect of virus infection and water stress on water flow and water economy in grapevines.Crossref | GoogleScholarGoogle Scholar | 28044321PubMed |
Encabo JR, Cabauatan PQ, Cabunagan RC, Satoh K, Lee JH, Kwak DY, De Leon TB, Macalalad RJA, Kondoh H, Kikuchi S, Choi I-R (2009) Suppression of two tungro viruses in rice by separable traits originating from cultivar Utri Merah. Molecular Plant-Microbe Interactions 22, 1268–1281.
| Suppression of two tungro viruses in rice by separable traits originating from cultivar Utri Merah.Crossref | GoogleScholarGoogle Scholar | 19737100PubMed |
Fronhoffs S, Totzke G, Stier S, Wernert N, Rothe M, Brüning T, Koch B, Sachinidis A, Vetter H, Ko Y (2002) A method for the rapid construction of cRNA standard curves in quantitative real-time reverse transcription polymerase chain reaction. Molecular and Cellular Probes 16, 99–110.
| A method for the rapid construction of cRNA standard curves in quantitative real-time reverse transcription polymerase chain reaction.Crossref | GoogleScholarGoogle Scholar | 12030760PubMed |
Fukao T, Yeung E, Bailey-Serres J (2011) The submergence tolerance regulator SUB1A mediates crosstalk between submergence and drought tolerance in rice. The Plant Cell 23, 412–427.
| The submergence tolerance regulator SUB1A mediates crosstalk between submergence and drought tolerance in rice.Crossref | GoogleScholarGoogle Scholar | 21239643PubMed |
Goel AK, Lundberg D, Torres MA, Matthews R, Akimoto-Tomiyama C, Farmer L, Dangl JL, Grant SR (2008) The Pseudomonas syringae type III effector HopAM1 enhances virulence on water-stressed plants. Molecular Plant-Microbe Interactions 21, 361–370.
| The Pseudomonas syringae type III effector HopAM1 enhances virulence on water-stressed plants.Crossref | GoogleScholarGoogle Scholar | 18257685PubMed |
Grimmer MK, Foulkes MJ, Paveley ND (2012) Foliar pathogenesis and plant water relations: a review. Journal of Experimental Botany 63, 4321–4331.
| Foliar pathogenesis and plant water relations: a review.Crossref | GoogleScholarGoogle Scholar | 22664583PubMed |
Gupta A, Dixit SK, Senthil-Kumar M (2016) Drought stress predominantly endures Arabidopsis thaliana to Pseudomonas syringae infection. Frontiers in Plant Science 7, 808
| Drought stress predominantly endures Arabidopsis thaliana to Pseudomonas syringae infection.Crossref | GoogleScholarGoogle Scholar | 27375661PubMed |
Hall AE, Loomis RS (1972) An explanation for the difference in photosynthetic capabilities of healthy and beet yellows virus-infected sugar beets (Beta vulgaris L.). Plant Physiology 50, 576–580.
| An explanation for the difference in photosynthetic capabilities of healthy and beet yellows virus-infected sugar beets (Beta vulgaris L.).Crossref | GoogleScholarGoogle Scholar | 16658220PubMed |
Hibino H (1983) Relations of rice tungro bacilliform and rice tungro spherical viruses with their vector Nephotettix virescens. Annals of the Phytopathological Society of Japan 49, 545–553.
| Relations of rice tungro bacilliform and rice tungro spherical viruses with their vector Nephotettix virescens.Crossref | GoogleScholarGoogle Scholar |
Hibino H, Roechan M, Sudarisman S (1978) Association of two types of virus particles with penyakit habang (tungro disease) of rice in Indonesia. Phytopathology 68, 1412–1416.
| Association of two types of virus particles with penyakit habang (tungro disease) of rice in Indonesia.Crossref | GoogleScholarGoogle Scholar |
Hibino H, Tiongco ER, Cabunagan RC, Flores ZM (1987) Resistance to rice tungro-associated viruses in rice under experimental and natural conditions. Phytopathology 77, 871–875.
| Resistance to rice tungro-associated viruses in rice under experimental and natural conditions.Crossref | GoogleScholarGoogle Scholar |
Holmgren M, Gómez-Aparicio L, Quero JL, Valladares F (2012) Non-linear effects of drought under shade: reconciling physiological and ecological models in plant communities. Oecologia 169, 293–305.
| Non-linear effects of drought under shade: reconciling physiological and ecological models in plant communities.Crossref | GoogleScholarGoogle Scholar | 22083284PubMed |
Hu H, Dai M, Yao J, Xiao B, Li X, Zhang Q, Xiong L (2006) Overexpressing a NAM, ATAF, and CUC (NAC) transcription factor enhances drought resistance and salt tolerance in rice. Proceedings of the National Academy of Sciences of the United States of America 103, 12987–12992.
| Overexpressing a NAM, ATAF, and CUC (NAC) transcription factor enhances drought resistance and salt tolerance in rice.Crossref | GoogleScholarGoogle Scholar | 16924117PubMed |
Hull R (1996) Molecular biology of rice tungro viruses. Annual Review of Phytopathology 34, 275–297.
| Molecular biology of rice tungro viruses.Crossref | GoogleScholarGoogle Scholar | 15012544PubMed |
IRRI (2002) ‘Standard evaluation system.’ (International Rice Research Institute: Manila, Philippines)
Jatan R, Lata C (2019) Role of microRNAs in abiotic and biotic stress resistance in plants. Proceedings of the Indian National Science Academy 85, 553–567.
Kijoji AA, Nchimbi-Msolla S, Kanyeka ZL, Klassen SP, Serraj R, Henry A (2013) Water extraction and root traits in Oryza sativa × Oryza glaberrima introgression lines under different soil moisture regimes. Functional Plant Biology 40, 54–66.
| Water extraction and root traits in Oryza sativa × Oryza glaberrima introgression lines under different soil moisture regimes.Crossref | GoogleScholarGoogle Scholar |
Lindsey DW, Gudauskas RT (1975) Effects of maize dwarf mosaic virus on water relations of corn. Phytopathology 65, 434–440.
| Effects of maize dwarf mosaic virus on water relations of corn.Crossref | GoogleScholarGoogle Scholar |
Mallikarjuna G, Mallikarjuna, K, Reddy MK, Kaul T (2011) Expression of OsDREB2A transcription factor confers enhanced dehydration and salt stress tolerance in rice. Biotechnology Letters 33, 1689–1697.
| Expression of OsDREB2A transcription factor confers enhanced dehydration and salt stress tolerance in rice.Crossref | GoogleScholarGoogle Scholar | 21528404PubMed |
Mayek-Pérez N, Garcia-Espínosa R, López-Castañeda C, Acosta-Gallegos JA, Simpson J (2002) Water relations, histopathology and growth of common bean (Phaseolus vulgaris L.) during pathogenesis of Macrophomina phaseolina under drought stress. Physiological and Molecular Plant Pathology 60, 185–195.
| Water relations, histopathology and growth of common bean (Phaseolus vulgaris L.) during pathogenesis of Macrophomina phaseolina under drought stress.Crossref | GoogleScholarGoogle Scholar |
McElrone AJ, Forseth IN (2004) Photosynthetic responses of a temperate liana to Xylella fastidiosa infection and water stress. Journal of Phytopathology 152, 9–20.
| Photosynthetic responses of a temperate liana to Xylella fastidiosa infection and water stress.Crossref | GoogleScholarGoogle Scholar |
McElrone AJ, Sherald JL, Forseth IN (2001) Effects of water stress on symptomatology and growth of Parthenocissus quinquefolia infected by Xylella fastidiosa. Plant Disease 85, 1160–1164.
| Effects of water stress on symptomatology and growth of Parthenocissus quinquefolia infected by Xylella fastidiosa.Crossref | GoogleScholarGoogle Scholar | 30823160PubMed |
McElrone AJ, Sherald JL, Forseth IN (2003) Interactive effects of water stress and xylem limited bacterial infection on the water relations of a host vine. Journal of Experimental Botany 54, 419–430.
| Interactive effects of water stress and xylem limited bacterial infection on the water relations of a host vine.Crossref | GoogleScholarGoogle Scholar | 12493870PubMed |
Mittler R, Blumwald E (2010) Genetic engineering for modern agriculture: challenges and perspectives. Annual Review of Plant Biology 61, 443–462.
| Genetic engineering for modern agriculture: challenges and perspectives.Crossref | GoogleScholarGoogle Scholar | 20192746PubMed |
Mohr PG, Cahill DM (2003) Abscisic acid influences the susceptibility of Arabidopsis thaliana to Pseudomonas syringae pv. tomato and Peronospora parasitica. Functional Plant Biology 30, 461–469.
| Abscisic acid influences the susceptibility of Arabidopsis thaliana to Pseudomonas syringae pv. tomato and Peronospora parasitica.Crossref | GoogleScholarGoogle Scholar |
Nakashima K, Tran LSP, Van Nguyen D, Fujita M, Maruyama K, Todaka D, Ito Y, Hayashi N, Shinozaki K, Yamaguchi‐Shinozaki K (2007) Functional analysis of a NAC‐type transcription factor OsNAC6 involved in abiotic and biotic stress responsive gene expression in rice. The Plant Journal 51, 617–630.
| Functional analysis of a NAC‐type transcription factor OsNAC6 involved in abiotic and biotic stress responsive gene expression in rice.Crossref | GoogleScholarGoogle Scholar | 17587305PubMed |
Ohnishi T, Sugahara S, Yamada T, Kikuchi K, Yoshiba Y, Hirano HY, Tsutsumi N (2005) OsNAC6, a member of the NAC gene family, is induced by various stresses in rice. Genes & Genetic Systems 80, 135–139.
| OsNAC6, a member of the NAC gene family, is induced by various stresses in rice.Crossref | GoogleScholarGoogle Scholar |
Pantaleo V, Vitali M, Boccacci P, Miozzi L, Cuozzo D, Chitarra W, Mannini F, Lovisolo C, Gambino G (2016) Novel functional microRNAs from virus-free and infected Vitis vinifera plants under water stress. Scientific Reports 6, 20167
| Novel functional microRNAs from virus-free and infected Vitis vinifera plants under water stress.Crossref | GoogleScholarGoogle Scholar | 26833264PubMed |
Prasch CM, Sonnewald U (2013) Simultaneous application of heat, drought, and virus to Arabidopsis plants reveals significant shifts in signaling networks. Plant Physiology 162, 1849–1866.
| Simultaneous application of heat, drought, and virus to Arabidopsis plants reveals significant shifts in signaling networks.Crossref | GoogleScholarGoogle Scholar | 23753177PubMed |
Rachmat A, Nugroho S, Sukma D, Aswidinnoor H (2014) Overexpression of OsNAC6 transcription factor from Indonesia rice cultivar enhances drought and salt tolerance. Emirates Journal of Food and Agriculture 26, 519–527.
| Overexpression of OsNAC6 transcription factor from Indonesia rice cultivar enhances drought and salt tolerance.Crossref | GoogleScholarGoogle Scholar |
Ramegowda V, Senthil-Kumar M, Ishiga Y, Kaundal A, Udayakumar M, Mysore KS (2013) Drought stress acclimation imparts tolerance to Sclerotinia sclerotiorum and Pseudomonas syringae in Nicotiana benthamiana. International Journal of Molecular Sciences 14, 9497–9513.
| Drought stress acclimation imparts tolerance to Sclerotinia sclerotiorum and Pseudomonas syringae in Nicotiana benthamiana.Crossref | GoogleScholarGoogle Scholar | 23644883PubMed |
Reusche M, Truskina J, Thole K, Nagel L, Rindfleisch S, Tran VT, Braus-Stromeyer SA, Braus GH, Teichmann T, Lipka V (2014) Infections with the vascular pathogens Verticillium longisporum and Verticillium dahliae induce distinct disease symptoms and differentially affect drought stress tolerance of Arabidopsis thaliana. Environmental and Experimental Botany 108, 23–37.
| Infections with the vascular pathogens Verticillium longisporum and Verticillium dahliae induce distinct disease symptoms and differentially affect drought stress tolerance of Arabidopsis thaliana.Crossref | GoogleScholarGoogle Scholar |
Sanfaçon H, Iwanami T, Karasev AV, van der Vlugt R, Wellink J, Wetzel T (2011) Secoviridae. In ‘Virus taxonomy: classification and nomenclature. Ninth report of the International Committee on Taxonomy of Viruses’. (Eds AMQ King, MJ Adams, EB Carstens, EJ Lefkowitz) pp. 881–899. (Academic Press: San Diego, CA, USA)
Satoh K, Kondoh H, De Leon TB, Macalalad RJA, Cabunagan RC, Cabauatan PQ, Mauleon R, Kikuchi S, Choi I-R (2013) Gene expression responses to Rice tungro spherical virus in susceptible and resistant near-isogenic rice plants. Virus Research 171, 111–120.
| Gene expression responses to Rice tungro spherical virus in susceptible and resistant near-isogenic rice plants.Crossref | GoogleScholarGoogle Scholar | 23183448PubMed |
Schmittgen TD, Livak KJ (2008) Analyzing real-time PCR data by the comparative CT method. Nature Protocols 3, 1101–1108.
| Analyzing real-time PCR data by the comparative CT method.Crossref | GoogleScholarGoogle Scholar | 18546601PubMed |
Sharma S, Dasgupta I (2012) Development of SYBR Green I based real-time PCR assays for quantitative detection of Rice tungro bacilliform virus and Rice tungro spherical virus. Journal of Virological Methods 181, 86–92.
| Development of SYBR Green I based real-time PCR assays for quantitative detection of Rice tungro bacilliform virus and Rice tungro spherical virus.Crossref | GoogleScholarGoogle Scholar | 22326276PubMed |
Shen P, Kaniewska M, Smith C, Beachy RN (1993) Nucleotide sequence and genomic organization of rice tungro spherical virus. Virology 193, 621–630.
| Nucleotide sequence and genomic organization of rice tungro spherical virus.Crossref | GoogleScholarGoogle Scholar | 8460478PubMed |
Shinkai A (1977) Rice waika, a new virus disease, and problems related to its occurrence and control. Japan Agricultural Research Quarterly 11, 151–155.
Uno Y, Furihata T, Abe H, Yoshida R, Shinozaki K, Yamaguchi-Shinozaki K (2000) Arabidopsis basic leucine zipper transcription factors involved in an abscisic acid-dependent signal transduction pathway under drought and high-salinity conditions. Proceedings of the National Academy of Sciences of the United States of America 97, 11632–11637.
| Arabidopsis basic leucine zipper transcription factors involved in an abscisic acid-dependent signal transduction pathway under drought and high-salinity conditions.Crossref | GoogleScholarGoogle Scholar | 11005831PubMed |
Wu X, Shiroto Y, Kishitani S, Ito Y, Toriyama K (2009) Enhanced heat and drought tolerance in transgenic rice seedlings overexpressing OsWRKY11 under the control of HSP101 promoter. Plant Cell Reports 28, 21–30.
| Enhanced heat and drought tolerance in transgenic rice seedlings overexpressing OsWRKY11 under the control of HSP101 promoter.Crossref | GoogleScholarGoogle Scholar | 18818929PubMed |
Xiang Y, Tang N, Du H, Ye H, Xiong L (2008) Characterization of OsbZIP23 as a key player of the basic leucine zipper transcription factor family for conferring abscisic acid sensitivity and salinity and drought tolerance in rice. Plant Physiology 148, 1938–1952.
| Characterization of OsbZIP23 as a key player of the basic leucine zipper transcription factor family for conferring abscisic acid sensitivity and salinity and drought tolerance in rice.Crossref | GoogleScholarGoogle Scholar | 18931143PubMed |
Xu P, Chen F, Mannas JP, Feldman T, Sumner LW, Roossinck MJ (2008) Virus infection improves drought tolerance. New Phytologist 180, 911–921.
| Virus infection improves drought tolerance.Crossref | GoogleScholarGoogle Scholar | 18823313PubMed |
Zhou L, Liu Y, Liu Z, Kong D, Duan M, Luo L (2010) Genome-wide identification and analysis of drought-responsive microRNAs in Oryza sativa. Journal of Experimental Botany 61, 4157–4168.
| Genome-wide identification and analysis of drought-responsive microRNAs in Oryza sativa.Crossref | GoogleScholarGoogle Scholar | 20729483PubMed |