

Spatiotemporal distribution of humpback whales off north-west Australia quantifying the Exmouth Gulf nursery area
Kate R. Sprogis A # , Alicia L. Sutton B # , Micheline-Nicole M. Jenner C and K. Curt S. Jenner C *
A # , Alicia L. Sutton B # , Micheline-Nicole M. Jenner C and K. Curt S. Jenner C *
A
B
C
# These authors contributed equally to this paper
Handling Editor: Paul Cooper
Abstract
Humpback whales (Megaptera novaeangliae) migrate from their Antarctic feeding grounds to their north-west Australian breeding grounds. North-west Australia has experienced rapid development of the mining and petroleum industry, and remains under increasing human pressure. The aim of this study was to examine the spatiotemporal distribution of humpback whales across the north-west: North West Cape, Pilbara and Exmouth Gulf. Aerial surveys (n = 184) were conducted from 2000 to 2010. After accounting for survey effort, kernel density estimation was used to assess the monthly distribution of whales. The largest density hotspot of whale groups was the centre to western edge of Exmouth Gulf. The highest encounter rate of mother–calf pairs was in October in Exmouth Gulf (0.0447 ± 0.0082 groups/km), which was significantly higher than in the North West Cape and Pilbara (0.0005 ± 0.0002 and 0.0011 ± 0.0005 groups/km, respectively). The comparison among the three regions confirms and quantifies Exmouth Gulf as an important nursery area for humpback whale mother–calf pairs. Nursery areas are important for lactating humpback whale mothers as they carry the highest energetic demands in the population. It is crucial that nursery areas remain relatively undisturbed from human pressures so mothers can rest to conserve energy and nurse their young.
Keywords: aerial survey, breeding grounds, coastal development, kernel density estimation, marine megafauna, Megaptera novaeangliae, North West Cape, species distribution.
Introduction
Humpback whales (Megaptera novaeangliae) off Western Australia (breeding stock D, group IV) generally migrate from their Antarctic feeding grounds to their tropical breeding grounds off north-west Australia (Chittleborough 1953; Bestley et al. 2019). Whales in this population migrate to warmer calving grounds, with some migrating over 8000 km, extending to Camden Sound (15°S) and north of 10°S (Chittleborough 1965; Dawbin and Gill 1991). On their northern and southern migration, humpback whales pass the North West Cape (21°S) between June and November (Jenner et al. 2001). On the southern migration, whales enter Exmouth Gulf (22°S), which is adjacent to the North West Cape, to breed, rest and nurse their young (Chittleborough 1953; Jenner et al. 2001). It was 70 years ago that the Gulf was described as a nursery area: ‘in September there were large numbers of adult humpback whales with calves in this Gulf, indicating that it was a nursery area for this species’ (Chittleborough 1953). While on the breeding ground, capital breeding humpback whales generally fast, thus, such nursery areas are of importance for whales to rest, especially for lactating mothers and their nursing calves (Christiansen et al. 2016; Bejder et al. 2019).
Historically, humpback whales were hunted off the North West Cape at Norwegian Bay (22.5°S) and Point Cloates (22.7°S) (Dakin 1934; Ruud 1952; Chittleborough 1965). Whaling of Stock D also occurred off Carnarvon, Albany, Antarctica and in the Southern Ocean (Chittleborough 1965). Due to low numbers of humpback whales, a resolution was passed for a ban on the killing of humpback whales by the International Whaling Commission (IWC) in 1963 (Bannister 1964). By then, Stock D was nearly decimated, and by 1968 was estimated as <270 individuals (Bannister and Hedley 2001). Illegal Soviet whaling continued until 1972, contributing to low population numbers (Yablokov 1994; Ivashchenko and Clapham 2014). Sighting rates remained low along the Western Australian coastline for almost 10 years (Bannister et al. 1991; Dawbin and Gill 1991). Since then, the population has been increasing at 9–12.7% per annum and has made a substantial recovery, with over 25,000 individuals estimated in 2008 (Hedley et al. 2011; Salgado Kent et al. 2012). Based on this rate, the population is estimated to have recovered (Bejder et al. 2016), and as a result has recently been removed from the list of threatened species for Australia’s Environment Protection and Biodiversity Conservation Act (DCCEEW 2022).
Today, humpback whales face different pressures in north-west Australia (Groom and Coughran 2012). This region has had near continuous mineral and petroleum industrial activity since the 1960s (Hanf et al. 2016). More recently, Exmouth Gulf has been the focus of proposals for the construction of evaporative salt ponds for mining, service wharfs, as well as a deep-water port and shipping channels for oil and gas facilities (EPA 2021; Sutton and Shaw 2021). Anthropogenic stressors may place pressures on the continued population success of humpback whales (Erbe et al. 2019; Smith et al. 2020; Sprogis et al. 2020b). When assessing future industrialised uses off north-west Australia, the spatiotemporal distribution and encounter rates of humpback whales should be considered.
The importance of north-west Australia to humpback whales became the focus of vessel and aerial based research studies decades ago (e.g. Chittleborough 1953; Jenner et al. 2001) and continues today. The aim of this study is to quantify the spatiotemporal distribution and encounter rates of humpback whales during the breeding season across north-west Australia (including North West Cape, Pilbara and Exmouth Gulf). The study utilises long-term aerial survey datasets collected from 2000 to 2010. Furthermore, opportunistic, but systematic data collection for other marine megafauna are also presented in the supplementary material to augment existing but scant records of species of management interest over this large, remote region. Opportunistic data collection included all cetaceans (mysticetes and odontocetes), dugongs (Dugong dugon), whale sharks (Rhincodon typus), manta rays (Mobula spp.) and sea turtles (Chelonioidea) (Supplementary material 1).
Materials and methods
Study area and aerial survey design
Aerial surveys were conducted off north-west Australia (~20.5–22.5°S) from 2000 to 2010. The study area encompassed state waters; in three regions (1) North West Cape (13,400 km2), (2) Pilbara (16,700 km2) and (3) Exmouth Gulf (3600 km2). The three regions vary in benthic habitat, turbidity and water depth; North West Cape (0–2500 m depth), Pilbara (0–960 m depth) and Exmouth Gulf (0–22 m depth) (Fig. 1).
The study area off the North West Cape, Pilbara and Exmouth Gulf, Western Australia, where aerial surveys were flown searching for humpback whales between 2000 and 2010. The three regions surveyed for marine megafauna are indicated in the legend by lines representing the aerial flight paths. The inset on the bottom right shows the location of the study area in the context of Australia. The UNESCO Ningaloo Reef World Heritage Area which was implemented in 2011 is shaded in grey. The bathymetry underlay and grey contour lines represent the water depth (in metres), showing the shallow shelf waters in Exmouth Gulf and the Pilbara region, and the deeper waters off the North West Cape. The location of the historic Norwegian Bay Whaling Station and Point Cloates whaling station are shown.
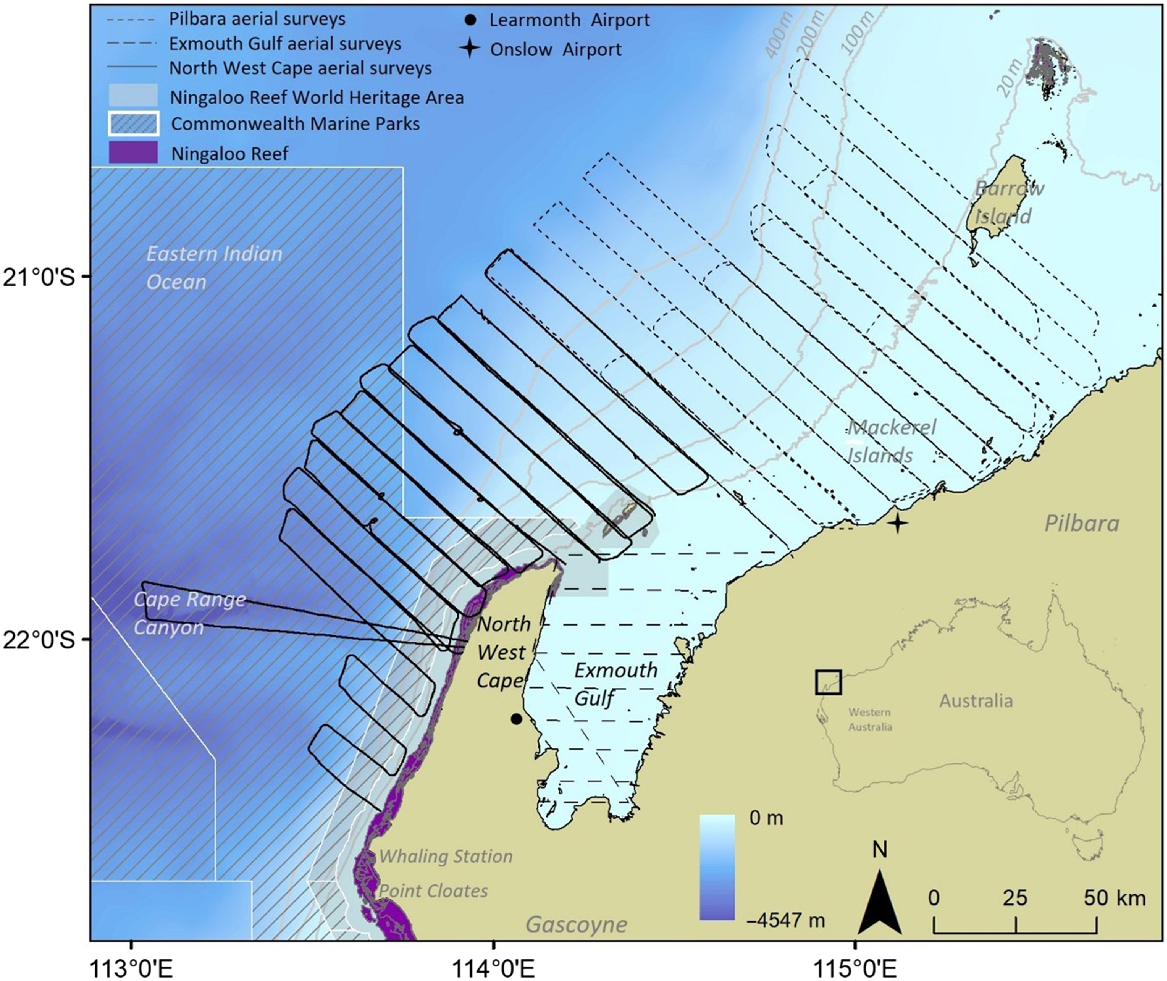
Aerial surveys were designed for detecting humpback whales and other large cetaceans, and were originally conducted for the purpose of population modelling and impact assessments for industry proponents (Jenner and Jenner 2005). These data have contributed to various research studies in the region (Sleeman et al. 2007; Braithwaite et al. 2012; Salgado Kent et al. 2012). Aerial survey design was consistent and followed protocols defined in the Distance v. 5.1 software program (Buckland et al. 2001). This program specifically allowed users to design line transect surveys and analyse data resulting from these surveys for the purpose of estimating density and abundance. Using the principles of this system, transects were drawn over the study area in order to maximise coverage probability during a single flight. Although parallel line transect designs are disadvantaged because the time spent in between transects is ‘off survey’, this technique results in an even probability of coverage for non-rectangular survey areas such as the current study area (Buckland et al. 2001). Surveys followed a set flight path along a parallel strip-transect design, where transects were perpendicular to bathymetric contours and to the humpback whale migration route. Transects were spaced ~10 km (±0.5 km) apart, and megafauna were scanned to ~5 km out (Fig. 1). North West Cape and Exmouth Gulf surveys were flown from Learmonth Airport, and Pilbara surveys from Onslow Airport (Fig. 1), using a twin-engine over-head wing airplane (Cessna 337 or Partenavia P68B fitted with bubble or standard windows). The plane maintained an altitude of 1000 feet (~305 m) and a speed of 120 knots (~222 km/h). Flights were conducted in wind speeds <18 knots, with no precipitation, beginning at ~0700 hours.
Humpback whale sightings were recorded in passing mode (i.e. the plane did not deviate from the flight path). For surveys flown off North West Cape and Pilbara, flights were flown north to south during the northern migration (June–August) and south to north during the expected southern migration period (August–November), to minimise the possibility of double counting groups of whales on successive transects. Flights in Exmouth Gulf were flown from north to south on all occasions, and it is acknowledged that some double counting of whales may have occurred in the Gulf as whales move both in and out of the Gulf.
Staff on each flight consisted of two pilots, two observers and, on some flights, a third researcher who was the data recorder with a time-coded Mini-Disk recorder (Sony Mini Disk Recorder NH900) which was synchronised to the Global Positioning System (GPS). The observer team was consistent for all flights each year and typically between years, and involved trained personnel who had prior experience spotting marine wildlife and/or humpback whale experience. Pilots were separated acoustically from the two observers and, data-wise, were responsible for recording the planes’ angle of drift on each transect, so that angles reported from the compass boards could be corrected relative to the flight path. Using clinometers (Suunto PM-5/360PC) and compass boards, the observers recorded the relative vertical and horizontal position of each humpback whale sighting, with the associated GPS location. A group of whales consisted of one or more individuals within 100 m of each other, predominantly involved in the same behavioural state and moving in the same general direction (Whitehead 1983). Observers recorded the group size and group composition, including the estimated number of adults and mother–calf pairs. A calf was defined as a whale small in size (less than 1/3 length of the adult it was travelling with) that may have light pigmentation (Chittleborough 1953; Clapham et al. 1999). Since the end of data collection for this study in 2010, the definition of a calf in the region has been updated (see Discussion). Observers recorded the environmental conditions during each transect line, including Beaufort Sea State and glare (scale: 0 = no glare, 1 = slight, 2 = significant, 3 = severe).
To obtain the correct location of the whales, the true angle from the aircraft to each sighting was later calculated with the following formula:
where AW is the angle to the whale, AC is the aircraft course, MHA is the measured horizontal angle and DA is the drift angle of the aircraft, which was either subtracted or added depending upon the side of the aircraft the animal was sighted on (as defined and described in Lerczak and Hobbs 1998). The GPS location of the whale sighting was then updated for density plotting purposes.
Aerial survey perception and availability bias
There are inherent biases with aerial surveys. Perception bias and availability bias are two forms of detection bias that could influence the number of animals sighted (Marsh and Sinclair 1989; Fuentes et al. 2015). Perception bias is when animals are missed by observers although they are potentially visible (available) (e.g. due to sun glare, worsening weather conditions). Perception bias was limited by flying in good weather conditions (e.g. <low glare, <18 knot winds, no precipitation). To further meet this bias, surveys were filtered to only include transect lines where the Beaufort Sea State was ≤3 prior to analyses. In addition, the average glare on the up-sun side of the plane was no more than 1.83 ± 0.71 (s.d.), reducing perception bias.
Availability bias is when animals are in the search area but are not visible to the observers, such as when they are underwater. Availability bias is influenced by environmental conditions (e.g. depth of water, turbidity) and/or animal behaviour (e.g. diving patterns, whether the animals are air breathing animals or not). Availability bias differs among the three different survey regions in several ways: (1) One example is in regard to dive duration, and if the whale is on or near the surface to be detected (Heide-Jørgensen and Laidre 2015). Dive durations across the three regions likely differ; for example, the North West Cape is a migratory corridor where whales are travelling and may dive for extended durations (~150–250 s: Sprogis et al. 2020a), whereas Exmouth Gulf is a resting area where whales commonly rest on or near the surface (Ejrnæs and Sprogis 2021). Furthermore, off the North West Cape groups with calves have a shorter dive duration compared to groups without calves (Sprogis et al. 2020a), therefore availability bias also differs within the area based on group composition. (2) Another example of availability bias is in regard to water turbidity (Marsh and Sinclair 1989). For example, Exmouth Gulf waters are highly turbid compared to the Pilbara and North West Cape waters (Cartwright et al. 2021), where Secchi disk measurements can be 0.5 m depth from the surface in Exmouth Gulf (Sprogis and Parra 2022). Therefore, sighting numbers in Exmouth Gulf are likely negatively biased due to the highly turbid waters. (3) A third example is in regard to water depth, where there are different water depths across the three areas. Humpback whales may use the water column differently and travel at depth (e.g. the mean maximum depth on migration is 83 m: Bestley et al. 2019) or rest near the surface (e.g. <15 m depth: Videsen et al. 2017). Lastly, the assumption is made that marine megafauna did not have a behavioural response to the sound of the plane prior to being within the field of vision of the observers (Patenaude et al. 2002). This assumption is made given the high altitude (~305 m) and fast speed flown (~222 km/h) to exit the area quickly without conducting closing mode. For the purpose of describing the distribution of whales, a sighting correction factor was not applied to the results as per analyses focussing on abundance estimates (e.g. Salgado Kent et al. 2012), and therefore the sighting records should be considered conservative estimates.
Identifying the spatiotemporal distribution of humpback whale groups and mother–calf pairs
The spatiotemporal density distribution of humpback whales was assessed for (1) all humpback whales, regardless of age class and/or sex, and (2) mother–calf pairs, as they are the most vulnerable to anthropogenic disturbance (Stamation et al. 2010). To identify ‘hotspot’ densities of humpback whales, kernel density estimation was used, which is a tool commonly used to visually assess the spatial distribution of humpback whales (Harris et al. 2012; Martins et al. 2013; MacKay et al. 2016; Dulau et al. 2017; Gonçalves et al. 2018; Derville et al. 2019b). Here, the kernel density tool was used from the Geostatistical toolbox in Esri’s ArcGIS© 10.8.1 (Esri, Redlands, California). Densities of whale groups and mother–calf pairs were assessed by month across the years of data collection (2000–2010). The geographical coordinate system used was GDA 2020. Prior to using the kernel density estimator (KDE) tool, survey effort across years were accounted for by dividing the sighting point (1) by the total kilometres of distance surveyed within a grid cell. Within ArcGIS, grid cells of 10 × 10 km spanned across the three regions of the study area. The size of the grid cells was selected due to the strip transect design with 10 km spacing and also to account for the large distances travelled by whales.
The kernel density estimator tool is based on the kernel function outlined in Silverman (1986). The output cell size (resolution) of the output raster was set to 1 × 1 km, which was rounded up from the automatically generated 0.7 × 0.7 km by the KDE tool for better interpretation and visualisation. A user defined option in the tool is the search radius, which is the bandwidth smoothing value (Worton 1989; Gitzen et al. 2006). The search radius was set to 20 km to include data points across two transect lines, which were spaced 10 km apart. The search radius value was the same across the three regions, and was constant across the plane to represent fixed kernels (Powell 2000). The tool calculated the density of point features around each output raster cell, thereby representing more tightly clustered group sightings as a higher density. A heat ramp colour scheme was used to highlight any hotspots of humpback whales.
Encounter rates of humpback whale groups, individuals and mother–calf pairs
Encounter rates were calculated for humpback whale groups, individuals and mother–calf pairs. Rates were derived by dividing the total number of group sightings (group encounter rate) and the total number of individuals (individual encounter rate) by the total kilometres flown within each 10 × 10 km grid cell. Encounter rates were derived monthly from June to November, and as an average across these months. Grid cells containing a distance less than the diagonal of the cell (<15 km) were removed in order to avoid artificial inflation of encounter rates due to small sample biases (e.g. Dinis et al. 2016; Awbery et al. 2022). Average encounter rates of humpback whales across region and months were compared using non-parametric Dunn-Bonferroni pair-wise testing (e.g. Exmouth Gulf versus Pilbara) in the statistical software JASP 0.11.1.0 (Love et al. 2019).
Results
Survey effort and humpback whale sightings
From 2000 to 2010, across 54 months, a total of 184 aerial surveys were flown across the study region (Table 1, Table S2). Of these, 101 surveys were flown off the North West Cape, 66 off Pilbara and 17 over Exmouth Gulf. In total, there was 169,821 linear km surveyed, consisting of 93,645 km off the North West Cape, 67,835 km off Pilbara, and 8341 km over Exmouth Gulf. Surveys began at ~07:00 and were, on average, 4.72 ± 1.35 s.d. h in duration. Flight durations ranged from 2.5 h in Exmouth Gulf to 7 h off Pilbara (inclusive of a mid-survey landing break for observers and refuelling). Humpback whale groups were sighted on 7413 occasions (Table S2). A subset of 6227 sightings was used in further analyses after filtering for Beaufort Sea State ≤3. The average number of whales in a group was 1.52 ± 0.010 s.e. (min = 1, max = 16).
| Year/month | Jan | Feb | Mar | Apr | May | Jun | Jul | Aug | Sep | Oct | Nov | Dec | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2000 | 4 | 4 | 3 | 2 | 3 | 2 | ||||||||||||
| 2001 | 4 | 4 | 2 | 2 | 3 | 1 | ||||||||||||
| 2002 | ||||||||||||||||||
| 2003 | ||||||||||||||||||
| 2004 | 3 | 1 | ||||||||||||||||
| 2005 | 1 | 1 | 2 | 1 | 1 | 1 | 2 | 3 | 1 | |||||||||
| 2006 | 3 | 3 | 3 | 3 | 3 | |||||||||||||
| 2007 | 1 | 4 | 3 | 2 | 3 | |||||||||||||
| 2008 | 2 | 5 | 6 | |||||||||||||||
| 2009 | 2 | 2 | 3 | 2 | 3 | 2 | 3 | 2 | 3 | 4 | 3 | 5 | 4 | |||||
| 2010 | 5 | 4 | 4 | 5 | 2 | 2 | 6 | 4 | 5 | 5 | 4 | 4 | 4 | |||||
Spatiotemporal distribution and encounter rates of humpback whale groups
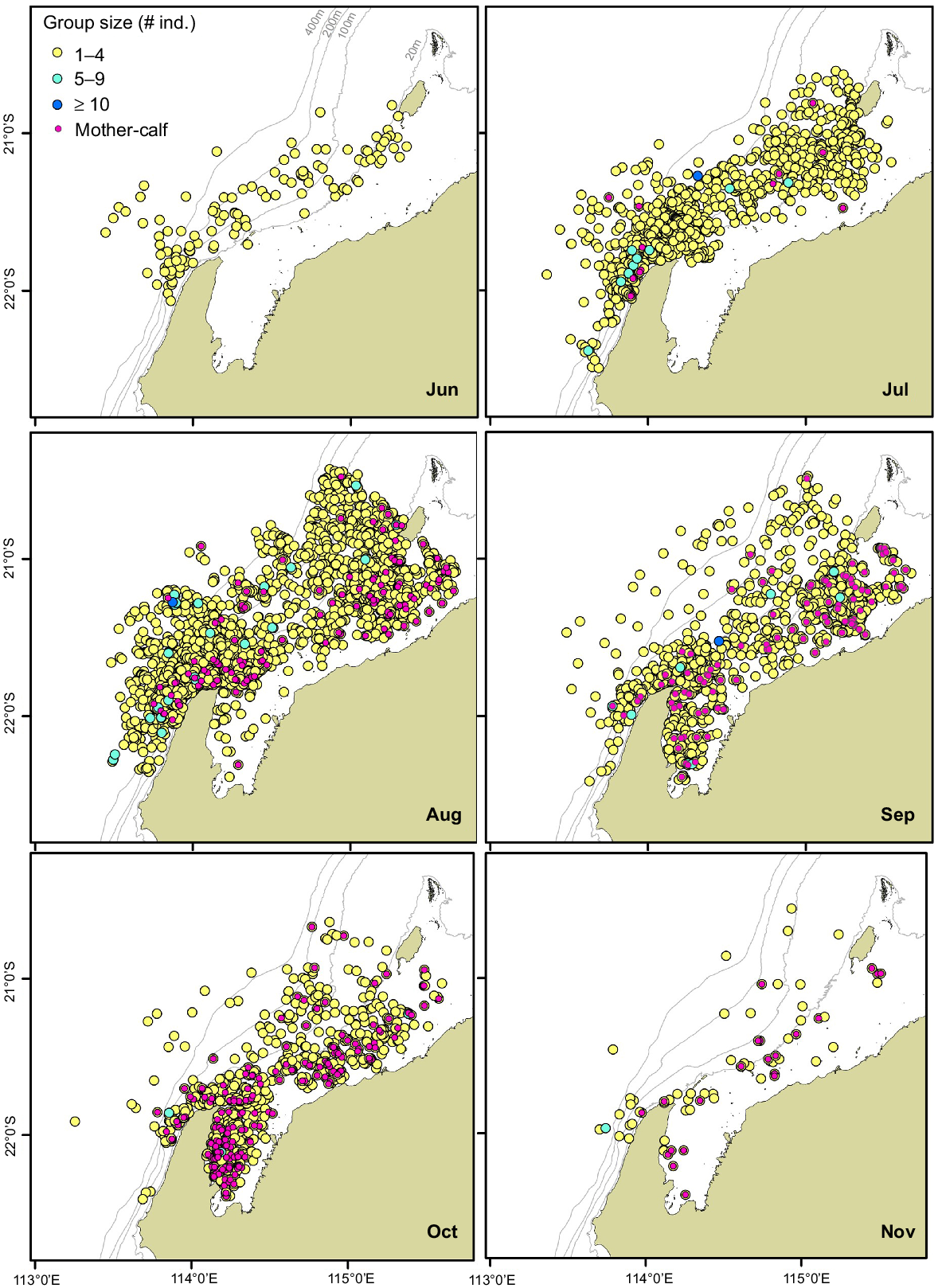
On the northern migration, from early in the austral winter (June and July), humpback whales were concentrated around the North West Cape and south-west of Barrow Island (Fig. 2). During August, a cross-over point between the northern and southern migration timing, there were higher densities of humpback whales observed off the Pilbara. On the southern migration, whales were in larger densities in Exmouth Gulf beginning in August and peaking in October. Within Exmouth Gulf, whales were primarily concentrated along the central to western coast, in shallow waters as far south as 22°25′S. The density of whales reduced across the north-west study region in November as the whales migrated south.
Kernel density plots of humpback whale group sightings (n = 6227; grey circles) off North West Cape, Pilbara and Exmouth Gulf off Western Australia. Sightings were weighted/standardised by survey effort (km) representing relative density. The darker clustered areas (red) of the kernel density heat map represent a higher density of whale group sightings, and the lighter coloured areas (white and light blue) represent a lower density of whale group sightings. Note: Group sightings of whales were recorded in both May (two whales) and December (mother–calf) off the Pilbara; however, data for these months were not included here. The months June to November are presented here (Austral winter and spring). These data represent aerial surveys across 2000 to 2010 (see Table 1 for details). Water depth contours are presented as light grey lines (in June panel). Note: each group sighting represents a different number of individuals: on average, 1.52 ± 0.010 s.e. whales (minimum = 1, maximum = 16 whales).
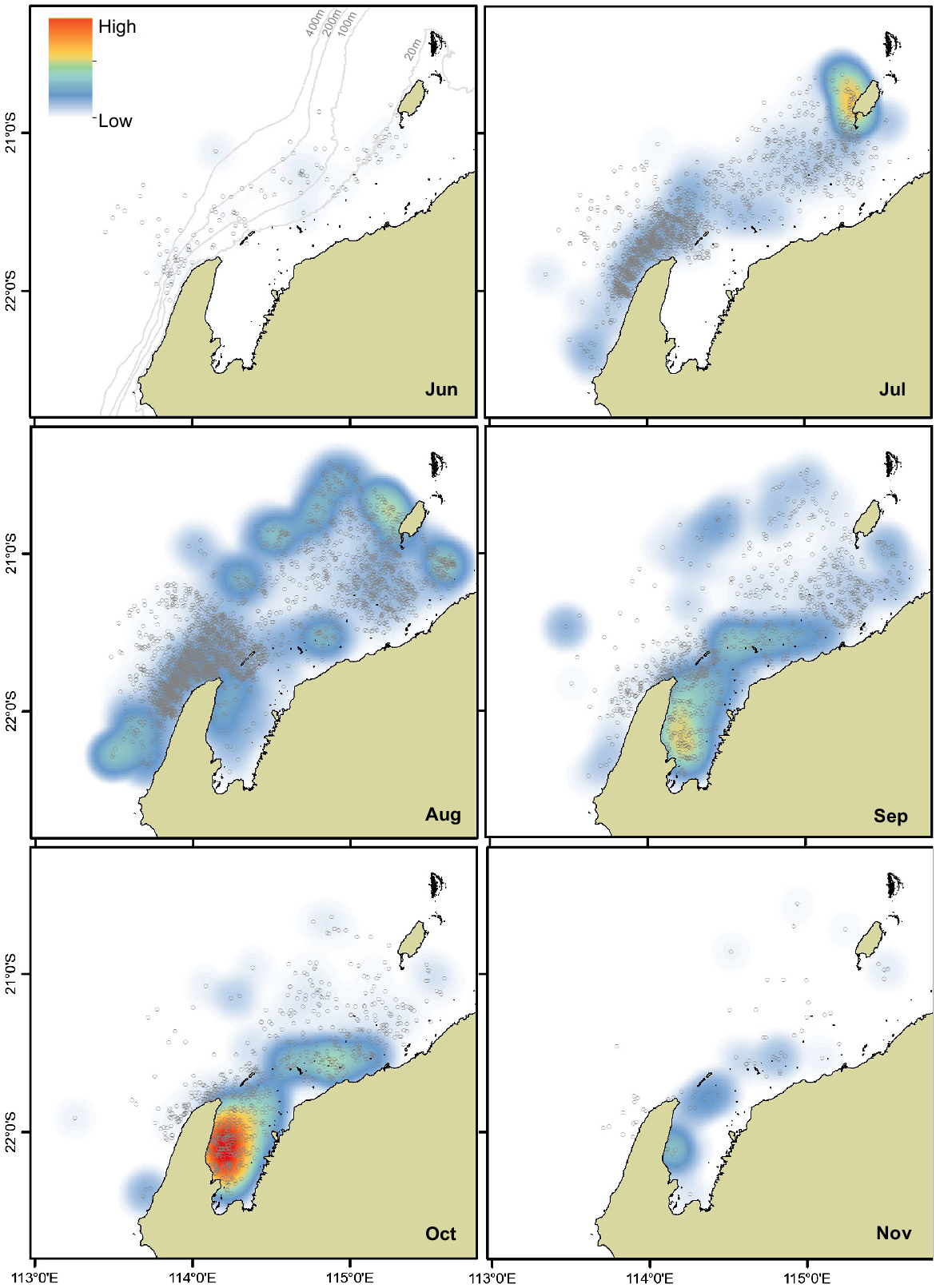
All three regions showed a bell-shaped trend in humpback whale sightings, where the arrival, peak and departure reflected the migratory patterns and timing of the whales. There was one sighting off the Pilbara in May, signifying the beginning of the northern migration. There may have also been sightings in May off the North West Cape; however, aerial flights were not flown in this location during this month due to perceived low numbers.
Group encounter rates across June to November were, on average, higher in Exmouth Gulf compared to North West Cape or Pilbara (P < 0.001) (Table 2). Averaged group encounter rates were not significantly different between North West Cape or Pilbara (P = 0.413). The group encounter rate was highest off the North West Cape in August (0.115 ± 0.007 (s.e.) groups/km), off Pilbara in August (0.151 ± 0.011 groups/km) and in Exmouth Gulf in September and October (Fig. 3a). In Exmouth Gulf, encounter rates of humpback whale groups were not significantly different between September and October (0.126 ± 0.018 and 0.126 ± 0.023 groups/km; P = 0.155) (Fig. 3a), or compared to Pilbara in August (P = 0.0180). Group encounter rates decreased in November (Fig. 3a), with only a single group sighting recorded in December off the Pilbara.
| Encounter rate/Region | North West Cape | Pilbara | Exmouth Gulf | |
|---|---|---|---|---|
| Groups/km | 0.0532 ± 0.0027 | 0.0511 ± 0.0027 | 0.0973 ± 0.0110 | |
| Individuals/km | 0.0794 ± 0.0043 | 0.0775 ± 0.0042 | 0.1574 ± 0.0182 | |
| Mother–calf groups/km | 0.0018 ± 0.0003 | 0.0054 ± 0.0005 | 0.0236 ± 0.0037 |
Encounter rates of humpback whales across the main months (x-axis) of migration in north-west Australia. (a) All whale groups and (b) humpback mother–calf groups were averaged by month across each study area. Group sightings of whales were recorded in both May (two whales) and December (mother–calf) off the Pilbara; however, these months were not included here. Upper standard error bars are provided. Note different y-axis values for (a) and (b).
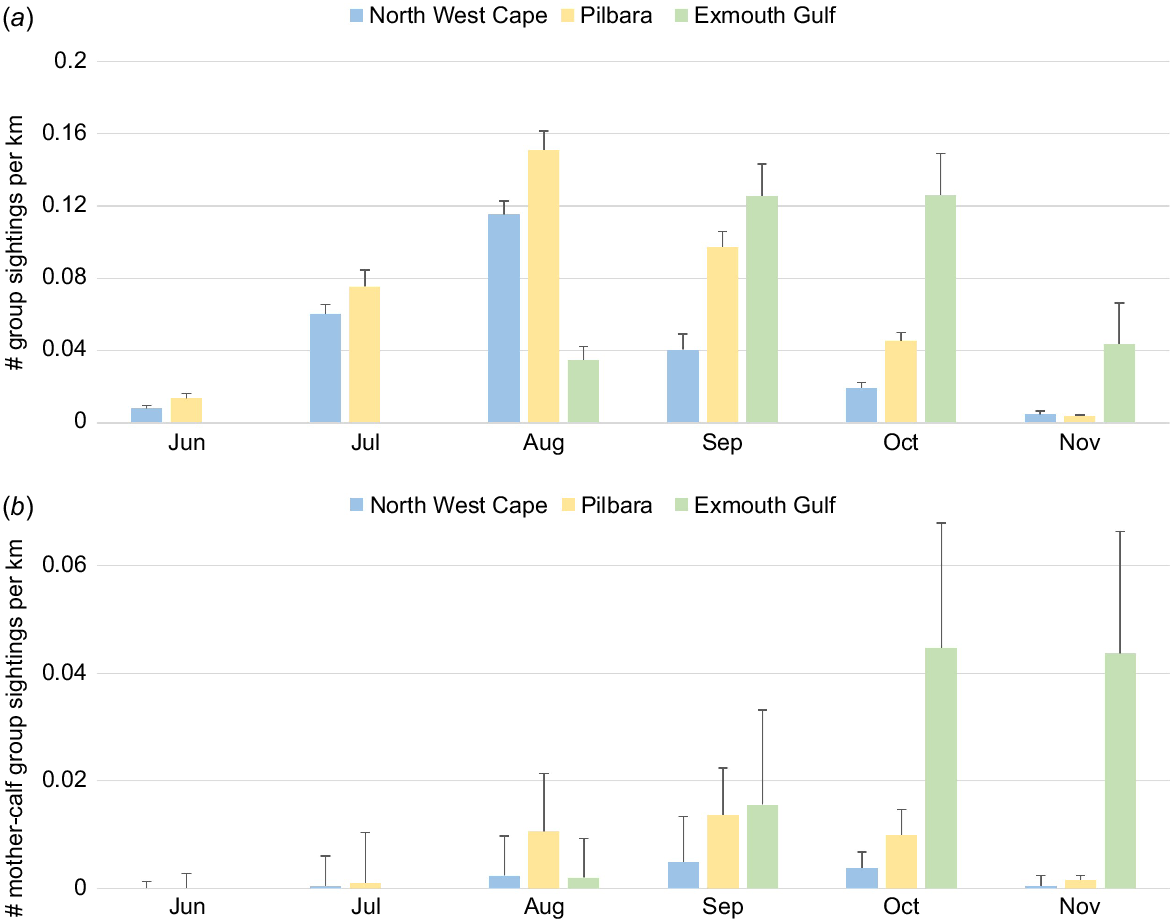
Individual encounter rates across June to November were, on average, higher in Exmouth Gulf compared to North West Cape or Pilbara (P < 0.001) (Table 2). Across the three regions, the largest individual encounter rate was 0.2235 ± 0.0164 individuals/km off the Pilbara in August. The smallest individual encounter rate was 0.0061 ± 0.0014 individuals/km off the Pilbara in November.
Spatiotemporal distribution and encounter rates of humpback whale mother–calf pairs
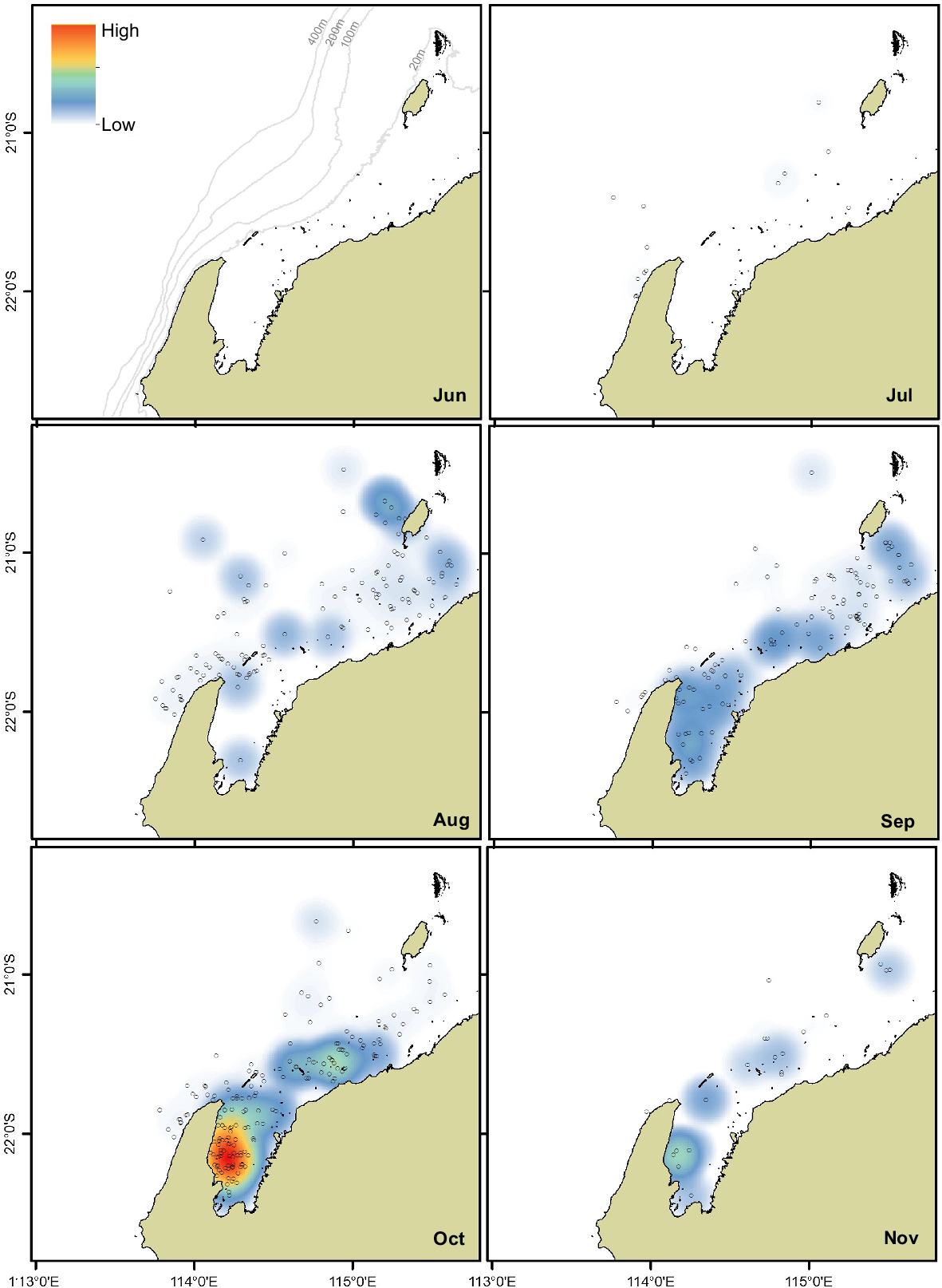
Mother–calf pairs were sighted off North West Cape and Pilbara from July to November (Figs 4, 5). Mother–calf pairs entered Exmouth Gulf from August, peaking in density in October (Fig. 4). Within Exmouth Gulf, mother–calf pairs were primarily concentrated along the central to south-western side. The density of mother–calf pairs decreased across the region in November. Mothers and calves were sighted out to the 400 m depth contour off North West Cape and Pilbara; however, they were generally distributed closer to the coast (Figs 4, 5).
Kernel density plots of humpback whale mother–calf group sightings (n = 428; grey circles) off North West Cape, Pilbara and Exmouth Gulf off Western Australia. Sightings were weighted/standardised by survey effort (km) representing relative density. The darker clustered areas (red) of the kernel density heat map represent a higher density of whale group sightings, whereas the lighter coloured areas (white and light blue) represent a lower density of whale group sightings. The months June to November are presented here (Austral winter and spring). Note: a single mother–calf pair was sighted in December off the Pilbara. These data represent aerial surveys across 2000 to 2010 (see Table 1 for details). Water depth contours are represented in light grey lines (in June panel).
Group size of humpback whales off North West Cape, Pilbara and Exmouth Gulf, Western Australia (n = 6227). Sightings (number of individuals) are from June to November (Austral winter and spring), incorporating aerial survey data from 2000 to 2010 (see Table 1 for details). Sightings show groups with mother–calf pairs present (magenta dots).
Initial sightings of calves occurred in July off the North West Cape and Pilbara (0.0005 ± 0.0002 and 0.0011 ± 0.0005 groups/km, respectively). The highest encounter rate of mother–calf pairs across the three regions was in October in Exmouth Gulf (0.0447 ± 0.0082 groups/km) (Fig. 3b). Fewer humpback whale mother–calf pairs were sighted in November; of these, most were sighted in Exmouth Gulf (0.0436 ± 0.0226 groups/km) (P < 0.001). Overall, the average encounter rate of mother–calf pairs was higher in Exmouth Gulf compared to North West Cape or Pilbara (P < 0.001) (Table 2).
Discussion
Spatiotemporal distribution and encounter rates of humpback whales
Long-term ecological datasets are beneficial to provide data and understanding to support evidence-based policy, decision making and management of an ecosystem (Lindenmayer et al. 2012). However, long-term datasets are difficult to collect partially due to logistical difficulties and financial costs. This study incorporates a long-term dataset spanning from 2000 to 2010, and although these data were collected more than a decade ago, this is the first study to compare humpback whale distribution records across the north-west, which can serve as a baseline that can be used by future studies. The findings from this study show a consistent spatiotemporal distribution of humpback whales across the study region compared to historic records and contemporary records, representing generational and reliable use of this region by what has been an exploited, protected and now recovered population of whales.
Humpback whales were temporally distributed across the North West Cape and Pilbara from May. Mother and calves were generally distributed closer to the coast, and were temporally distributed across the North West Cape and Pilbara from July. Similarly, other studies have shown that newborn calves are present off the North West Cape from July (Chittleborough 1953; Irvine et al. 2018; Ransome et al. 2022). Humpback whales migrate into Exmouth Gulf from August; however, recent studies in the Gulf have shown that whales can arrive during the northern migration from June (Sprogis and Parra 2022). Humpback whales were temporally distributed across the study area until November. Migration departure timing from the region appears to vary slightly each year; for example, whales may remain in Exmouth Gulf until October/November (e.g. in 2018: Irvine and Salgado Kent 2019; Sprogis et al. 2020b), or depart by early–mid October (e.g. in 2019 and 2021: Ejrnæs and Sprogis 2021; Sprogis and Waddell 2022). Drivers influencing migration timing departure from the region likely relate to various factors, including (1) selection pressures on different age, sex and reproductive classes, with mother–calf pairs migrating last (Chittleborough 1965; Craig et al. 2003; Franklin et al. 2011), (2) the decrease in body condition of individuals over the breeding season (Christiansen et al. 2020; Russell et al. 2023), (3) large-scale climatic drivers, such as El Niño Southern Oscillation (Pelayo-González et al. 2022; Denkinger et al. 2023), and/or more locally driven influences, such as (4) variations in sea surface temperature (Fig. S3) (Rasmussen et al. 2007; Derville et al. 2019a), and/or (5) the prevalence of predators and micropredation (Chittleborough 1953; Pitman et al. 2015; Harkness and Sprogis 2020).
Kernel density analysis of whale group sightings across the North West Cape, Pilbara and Exmouth Gulf highlighted that the largest humpback whale hotspot was the centre to western edge of Exmouth Gulf. This is despite the Gulf being the least surveyed area within the study. This spatial distribution in the Gulf is supported by previous (Jenner et al. 2001) and contemporary (Irvine and Salgado Kent 2019; Sprogis and Parra 2022; Sprogis and Waddell 2022) studies. The occurrence of a hotspot in Exmouth Gulf is further supported by encounter rates being higher, on average, in Exmouth Gulf compared to North West Cape or Pilbara. Specifically, group encounter rates were highest in Exmouth Gulf in September and October (0.126 ± 0.018 and 0.126 ± 0.023 groups/km). Similar group encounter rates in Exmouth Gulf were recorded during boat-based surveys in October 2021 (0.102 groups/km: Sprogis and Waddell 2022). The high-density space use within Exmouth Gulf may be driven by social and ecological factors, including (1) the mating system, where aggregations are beneficial for mating purposes (Braithwaite et al. 2012; Clapham and Zerbini 2015), and (2) ideal environmental conditions for resting and nursing, where the Gulf waters may be suitably shallow (<20 m depth) and sheltered from prevailing winds and swell (Franklin et al. 2018). The importance of shallow, sheltered waters for core breeding habitat has been shown for other breeding populations, such as in Oceania (Derville et al. 2019a) and Ecuador (Félix and Botero-Acosta 2011; Denkinger et al. 2023).
The encounter rate of mother–calf pairs in Exmouth Gulf increased from September, peaking in October. This pulse in mother–calf pairs is supported by other studies (Jenner et al. 2001; Braithwaite et al. 2012; Irvine and Salgado Kent 2019). Although the importance of the Gulf for mother–calf pairs was noted 70 years ago (Chittleborough 1953), this study quantifies, for the first time, Exmouth Gulf as an important nursery area by comparing mother–calf densities and encounter rates among the three study regions. The high density and encounter rate for calves in the Gulf are considered to be minimum values, as calves nursing or resting below the surface may not be sighted during aerial surveys due to lack of visibility in the highly turbid waters of the Gulf (i.e. availability bias) (Cartwright et al. 2021). The numbers of calves recorded in this study may also be underestimated due to the definition of a calf used, which was a whale 1/3 the length of its mother (Clapham et al. 1999). This definition represents smaller calves, and recent research in the region has shown that calves grow to >50% of their mother’s length (Christiansen et al. 2016; Irvine et al. 2018), from ~4 to 8 m length, thus representing <2/3 the length of the mother’s length (Sprogis et al. 2020a). Furthermore, it was previously considered that the Gulf was a nursery area for older postneonatal south-bound calves; however, recent research distinguishes between neonates and postneonates, highlighting that the Gulf is also important for neonatal calves (Irvine and Salgado Kent 2019; Ejrnæs and Sprogis 2021). Future research documenting neonates and postneonatal calves is recommended for further describing the importance of the Gulf.
A main difference between the high density area in the western Gulf versus other areas is that the whales in the western Gulf reside for extended periods and are largely resting and nursing their young (Jenner et al. 2001). In the Gulf, whales are able to rest on and below the surface for extended durations (i.e. 35% of their time resting) (Bejder et al. 2019; Sprogis et al. 2020b; Ejrnæs and Sprogis 2021). Nursery areas are crucial for resting and nursing mother–calves, as lactating mothers carry the highest energetic cost in the population through gestation and lactation (Christiansen et al. 2016). Calves need to rest and nurse to grow strong enough to endure the migration, and in Exmouth Gulf calves nurse for around 21% of their time (Videsen et al. 2017; Bejder et al. 2019; Ejrnæs and Sprogis 2021). The shallow, sheltered waters of Exmouth Gulf, which are currently relatively undisturbed by anthropogenic pressures, provide an ideal and unique resting and nursing area for the Western Australian humpback whale population. However, the location of the nursery area close to the coast increases the chances of overlap with anthropogenic stressors, such as boating activities and coastal development.
Management considerations
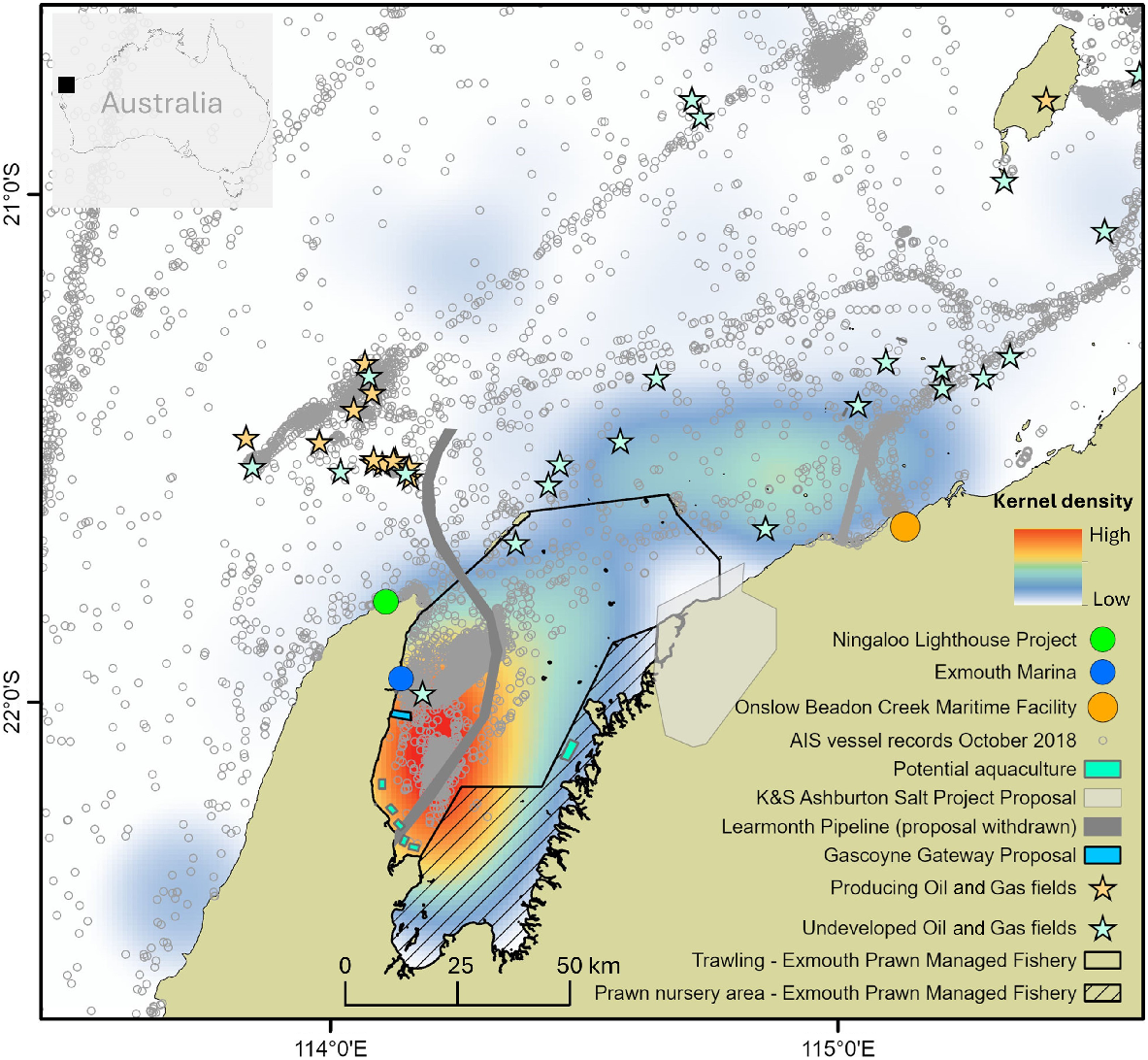
The Western Australian migratory humpback whale population has taken over three decades to recover from commercial and illegal whaling pressures. Today, humpback whales in the region face different pressures, such as from ship strike, underwater vessel noise disturbance, entanglement in fishing gear, habitat modification, disturbance from tourism (e.g. swim-with-humpback-whale tourism) and other human associated stressors (e.g. gull attacks and harassment) (Fig. 6) (Groom and Coughran 2012; Erbe et al. 2019; Harkness and Sprogis 2020; Sprogis et al. 2020a, 2020b; Arranz et al. 2021). Although waters around the North West Cape are included in state and commonwealth marine parks, and are afforded World Heritage Area status, the Pilbara and Exmouth Gulf waters remain largely unprotected from potential cumulative impacts (Fig. 6).
A map to demonstrate the types of overlap that may exist in the region. The map includes data for a kernel density estimation for humpback whale group sightings (October 2004–2005) overlaid with anthropogenic activities, including the current Exmouth Gulf Prawn Managed Fishery and mining and exploration leases, the rejected SubSea 7 Learmonth Pipeline proposal, the proposed Gascoyne Gateway deep-water port and the K+S Ashburton Salt project (including service wharfs) and typical shipping traffic locations (vessels with Automatic Identification System (AIS) presented for the month of October 2018 representing data prior to COVID-19).
Exmouth Gulf is a key resting/nursery area for breeding stock D migrating humpback whales off Western Australia. On the east coast of Australia, there is a similar situation, where Hervey Bay is a crucial resting/nursery area for breeding stock E-1 humpback whales. Hervey Bay is especially important for mother–calves on their southern migration (Franklin et al. 2011; Stack et al. 2021), where the whales predominantly occupy the eastern section of Platypus Bay (Corkeron et al. 1994). Whereas Exmouth Gulf remains unprotected, sections of Platypus Bay are protected within the Great Sandy Marine Park and there are restrictions to limit any disturbance on this important aggregation of humpback whales.
We show that the western side of Exmouth Gulf is a high density area for humpback whales, and overlay the recent rejected and current proposed anthropogenic activities (Fig. 6), such as a deep-water port. The humpback whale high density area on the western side of Exmouth Gulf is of importance as the waters are relatively shallow, and the introduction of future industrialisation or coastal development, such as a deep-water port (Fig. 6), will increase the amount of underwater noise from shipping and increase the chances of ship strike in such shallow waters. Future monitoring and management of the Exmouth Gulf nursery area is recommended to assist in reducing any impact from vessel noise disturbance and vessel strike in these shallow Gulf waters. Furthermore, it is recommended that the humpback whale nursery area in Exmouth Gulf be identified as a priority area and protected for the continued conservation of the species. The recognition of the humpback whale nursery area in Exmouth Gulf is one of the main drivers for the recognition of an Important Marine Mammal Area (IMMA) (IUCN-MMPATF 2022). The IMMA is not currently a protected area, but is identified for use by governments, intergovernmental organisations, conservation groups, and the general public, to prioritise areas for consideration for further conservation measures. Future long-term studies can be used for comparative purposes to this study, and to assess impacts from gull attacks on mother–calf pairs (Harkness and Sprogis 2020) and from a changing climate. With the warming of waters due to climate change, migration timing and duration of stay for humpback whales may be altered, where >28°C water is predicted to become unsuitably warm for humpback whales on the breeding grounds (Fig. S3) (Derville et al. 2019a; von Hammerstein et al. 2022). The generational use of humpback whales across the region is of importance and pertinent for management agencies and industries to assess conflicts with proposed coastal developments, and to make informed decisions about the continued conservation and management of this migratory species.
Declaration of funding
Funding for the aerial flights is from the EPBC Act (Australia) related investigations, in chronological order, from Woodside Energy, BHP Billiton, Straits Salt, Kailis Seafoods, Chevron and Exxon. These funding sources had no involvement in the preparation of the manuscript or the decision to submit for publication.
Acknowledgements
The authors thank the aerial observer team, including, Caroline Williams, Jane Kennedy, Jennifer Thompson, Lyn Irvine, Emily Wilson, Dani Rob, Vanessa Sturrock and pilots coordinated by Eric Rolston. Thank you to the Editor and anonymous reviewers for their input in strengthening the paper.
References
Arranz P, Aguilar de Soto N, Madsen PT, Sprogis KR (2021) Whale-watch vessel noise levels with applications to whale-watching guidelines and conservation. Marine Policy 134, 104776.
| Crossref | Google Scholar |
Awbery T, Akkaya A, Lyne P, Rudd L, Hoogenstrijd G, Nedelcu M, Kniha D, Erdoğan MA, Persad C, Amaha Öztürk A (2022) Spatial distribution and encounter rates of delphinids and deep diving cetaceans in the eastern Mediterranean Sea of Turkey and the extent of overlap with areas of dense marine traffic. Frontiers in Marine Science 9, 860242.
| Crossref | Google Scholar |
Bannister JL, Hedley SL (2001) Southern Hemisphere group IV humpback whales: their status from recent aerial survey. Memoirs-Queensland Museum 47, 587-598.
| Google Scholar |
Bannister JL, Kirkwood GP, Wayte SE (1991) Increase in humpback whales off Western Australia. Reports of the International Whaling Commission 41, 461-465.
| Google Scholar |
Bejder M, Johnston DW, Smith J, Friedlaender A, Bejder L (2016) Embracing conservation success of recovering humpback whale populations: evaluating the case for downlisting their conservation status in Australia. Marine Policy 66, 137-141.
| Crossref | Google Scholar |
Bejder L, Videsen S, Hermannsen L, Simon M, Hanf D, Madsen PT (2019) Low energy expenditure and resting behaviour of humpback whale mother–calf pairs highlights conservation importance of sheltered breeding areas. Scientific Reports 9, 771.
| Crossref | Google Scholar |
Bestley S, Andrews-Goff V, van Wijk E, Rintoul SR, Double MC, How J (2019) New insights into prime Southern Ocean forage grounds for thriving Western Australian humpback whales. Scientific Reports 9, 13988.
| Crossref | Google Scholar | PubMed |
Braithwaite JE, Meeuwig JJ, Jenner KCS (2012) Estimating cetacean carrying capacity based on spacing behaviour. PLoS ONE 7, e51347.
| Crossref | Google Scholar | PubMed |
Cartwright PJ, Fearns PRCS, Branson P, Cuttler MVW, O’Leary M, Browne NK, Lowe RJ (2021) Identifying metocean drivers of turbidity using 18 years of MODIS satellite data: implications for marine ecosystems under climate change. Remote Sensing 13, 3616.
| Crossref | Google Scholar |
Chittleborough RG (1953) Aerial observations on the humpback whale, Megaptera nodosa (Bonnaterre), with notes on other species. Australian Journal of Marine and Freshwater Research 4, 219-226.
| Crossref | Google Scholar |
Chittleborough RG (1965) Dynamics of two populations of the humpback whale, Megaptera novaeangliae (Borowski). Australian Journal of Marine and Freshwater Research 16, 33-128.
| Crossref | Google Scholar |
Christiansen F, Dujon AM, Sprogis KR, Arnould JPY, Bejder L (2016) Noninvasive unmanned aerial vehicle provides estimates of the energetic cost of reproduction in humpback whales. Ecosphere 7, e01468.
| Crossref | Google Scholar |
Christiansen F, Sprogis KR, Gross J, Castrillon J, Warick HA, Leunissen E, Bengtson Nash S (2020) Variation in outer blubber lipid concentration does not reflect morphological body condition in humpback whales. Journal of Experimental Biology 223, jeb213769.
| Crossref | Google Scholar |
Clapham PJ, Zerbini AN (2015) Are social aggregation and temporary immigration driving high rates of increase in some Southern Hemisphere humpback whale populations? Marine Biology 162, 625-634.
| Crossref | Google Scholar |
Clapham PJ, Wetmore SE, Smith TD, Mead JG (1999) Length at birth and at independence in humpback whales. Journal of Cetacean Research and Management 1, 141-146.
| Google Scholar |
Corkeron PJ, Brown M, Slade RW, Bryden MM (1994) Humpback whales, Megaptera novaeangliae (Cetaceaa: Balaenopteridae), in Hervey Bay, Queensland. Wildlife Research 21, 293-205.
| Crossref | Google Scholar |
Craig AS, Herman LM, Gabriele CM, Pack AA (2003) Migratory timing of humpback whales (Megaptera novaeangliae) in the central North Pacific varies with age, sex and reproductive status. Behaviour 140, 981-1001.
| Crossref | Google Scholar |
Dawbin WH, Gill PC (1991) Humpback whale survey along the west coast of Australia: a comparison of visual and acoustic observations. Memoirs of the Queensland Museum 30(2), 255-257.
| Google Scholar |
Denkinger J, Eguiguren A, Rubianes F, Munõz-Abril L, Onã J (2023) Humpback whale (Megaptera novaeangliae) winter distribution and core habitats in relation to El Ninõ Southern Oscillation and depth in coastal and oceanic waters off Ecuador. Marine Mammal Science 39, 876-892.
| Crossref | Google Scholar |
Derville S, Torres LG, Albertson R, Andrews O, Baker CS, Carzon P, Constantine R, Donoghue M, Dutheil C, Gannier A, Oremus M, Poole MM, Robbins J, Garrigue C (2019a) Whales in warming water: assessing breeding habitat diversity and adaptability in Oceania’s changing climate. Global Change Biology 25, 1466-1481.
| Crossref | Google Scholar | PubMed |
Derville S, Torres LG, Dodémont R, Perard V, Garrigue C (2019b) From land and sea, long-term data reveal persistent humpback whale (Megaptera novaeangliae) breeding habitat in New Caledonia. Aquatic Conservation: Marine and Freshwater Ecosystems 29, 1697-1711.
| Crossref | Google Scholar |
Dinis A, Carvalho A, Alves F, Nicolau C, Ribeiro C, Kaufmann M, Cañadas A, Freitas L (2016) Spatial and temporal distribution of bottlenose dolphins, Tursiops truncatus, in the Madeira archipelago, NE Atlantic. Arquipélago-Life and Marine Sciences 33, 45-54.
| Google Scholar |
Dulau V, Pinet P, Geyer Y, Fayan J, Mongin P, Cottarel G, Zerbini A, Cerchio S (2017) Continuous movement behavior of humpback whales during the breeding season in the southwest Indian Ocean: on the road again!. Movement Ecology 5, 11.
| Crossref | Google Scholar | PubMed |
Ejrnæs DD, Sprogis KR (2021) Ontogenetic changes in energy expenditure and resting behaviour of humpback whale mother–calf pairs examined using unmanned aerial vehicles. Wildlife Research 49, 34-45.
| Crossref | Google Scholar |
Erbe C, Marley SA, Schoeman RP, Smith JN, Trigg LE, Embling CB (2019) The effects of ship noise on marine mammals – A review. Frontiers in Marine Science 6, 606.
| Crossref | Google Scholar |
Félix F, Botero-Acosta N (2011) Distribution and behaviour of humpback whale mother–calf pairs during the breeding season off Ecuador. Marine Ecology Progress Series 426, 277-287.
| Crossref | Google Scholar |
Franklin T, Franklin W, Brooks L, Harrison P, Baverstock P, Clapham P (2011) Seasonal changes in pod characteristics of eastern Australian humpback whales (Megaptera novaeangliae), Hervey Bay 1992–2005. Marine Mammal Science 27, E134-E152.
| Crossref | Google Scholar |
Franklin T, Franklin W, Brooks L, Harrison P (2018) Site-specific female-biased sex ratio of humpback whales (Megaptera novaeangliae) during a stopover early in the southern migration. Canadian Journal of Zoology 96, 533-544.
| Crossref | Google Scholar |
Fuentes MMPB, Bell I, Hagihara R, Hamann M, Hazel J, Huth A, Seminoff JA, Sobtzick S, Marsh H (2015) Improving in-water estimates of marine turtle abundance by adjusting aerial survey counts for perception and availability biases. Journal of Experimental Marine Biology and Ecology 471, 77-83.
| Crossref | Google Scholar |
Gitzen RA, Millspaugh JJ, Kernohan BJ (2006) Bandwidth selection for fixed-kernel analysis of animal utilization distributions. Journal of Wildlife Management 70, 1334-1344.
| Crossref | Google Scholar |
Gonçalves MIC, De Sousa-Lima RS, Teixeira NN, Morete ME, De Carvalho GH, Ferreira HM, Baumgarten JE (2018) Low latitude habitat use patterns of a recovering population of humpback whales. Journal of the Marine Biological Association of the United Kingdom 98, 1087-1096.
| Crossref | Google Scholar |
Groom CJ, Coughran DK (2012) Entanglements of baleen whales off the coast of Western Australia between 1982 and 2010: patterns of occurrence, outcomes and management responses. Pacific Conservation Biology 18, 203-214.
| Crossref | Google Scholar |
Hanf DM, Hunt T, Parra GJ (2016) Humpback dolphins of Western Australia: a review of current knowledge and recommendations for future management. Advances in Marine Biology 73, 193-218.
| Crossref | Google Scholar |
Harkness P, Sprogis KR (2020) Silver gull harassment of humpback whales in Exmouth Gulf, Western Australia. Marine and Freshwater Research 72, 584-592.
| Crossref | Google Scholar |
Harris K, Gende SM, Logsdon MG, Klinger T (2012) Spatial pattern analysis of cruise ship–humpback whale interactions in and near Glacier Bay National Park, Alaska. Environmental Management 49, 44-54.
| Crossref | Google Scholar | PubMed |
Hedley SL, Bannister JL, Dunlop RA (2011) Abundance estimates of breeding stock ‘D’ humpback whales from aerial and land-based surveys off Shark Bay, Western Australia, 2008. Journal of Cetacean Research and Management [Special Issue 3] 209-221.
| Crossref | Google Scholar |
Heide-Jørgensen MP, Laidre KL (2015) Surfacing time, availability bias and abundance of humpback whales in west Greenland. Journal of Cetacean Research and Management 15, 1-8.
| Crossref | Google Scholar |
Irvine L, Salgado Kent C (2019) The distribution and relative abundance of marine mega-fauna, with a focus on humpback whales (Megaptera novaeangliae), in Exmouth Gulf, Western Australia, 2018. Subsea 7, Attachment 2J Humpback Whale Aerial Survey Report. (Marine mega-fauna in Exmouth Gulf: Exmouth, WA, Australia). p. 25.
Irvine LG, Thums M, Hanson CE, McMahon CR, Hindell MA (2018) Evidence for a widely expanded humpback whale calving range along the Western Australian coast. Marine Mammal Science 34, 294-310.
| Crossref | Google Scholar |
IUCN-MMPATF (2022) Ningaloo Reef to Montebello Islands IMMA factsheet. In ‘IUCN Joint SSC/WCPA Marine Mammal Protected Areas Task Force’. Available at https://www.marinemammalhabitat.org/portfolio-item/ningaloo-reef-montebello-islands/
Ivashchenko YV, Clapham PJ (2014) Too much is never enough: the cautionary tale of Soviet illegal whaling. Marine Fisheries Review 76, 1-21.
| Crossref | Google Scholar |
Jenner KCS, Jenner MN, McCabe KA (2001) Geographical and temporal movements of humpback whales in Western Australian waters. APPEA Journal 38, 692-707.
| Google Scholar |
Lerczak JA, Hobbs RC (1998) Calculating sighting distances from angular readings during shipboard, aerial, and shore-based marine mammal surveys. Marine Mammal Science 14, 590-598.
| Crossref | Google Scholar |
Lindenmayer DB, Likens GE, Andersen A, Bowman D, Bull CM, Burns E, Dickman CR, Hoffmann AA, Keith DA, Liddell MJ, Lowe AJ, Metcalfe DJ, Phinn SR, Russell-Smith J, Thurgate N, Wardle GM (2012) Value of long-term ecological studies. Austral Ecology 37, 745-757.
| Crossref | Google Scholar |
Love J, Selker R, Marsman M, Jamil T, Dropmann D, Verhagen J, Ly A, Gronau QF, Šmíra M, Epskamp S, Matzke D, Wild A, Knight P, Rouder JN, Morey RD, Wagenmakers E-J (2019) JASP: graphical statistical software for common statistical designs. Journal of Statistical Software 88(2), 1-17.
| Crossref | Google Scholar |
MacKay MM, Würsig B, Bacon CE, Selwyn JD (2016) North Atlantic humpback whale (Megaptera novaeangliae) hotspots defined by bathymetric features off western Puerto Rico. Canadian Journal of Zoology 94, 517-527.
| Crossref | Google Scholar |
Marsh H, Sinclair DF (1989) Correcting for visibility bias in strip transect aerial surveys of aquatic fauna. The Journal of Wildlife Management 53, 1017-1024.
| Crossref | Google Scholar |
Martins CCA, Andriolo A, Engel MH, Kinas PG, Saito CH (2013) Identifying priority areas for humpback whale conservation at eastern Brazilian Coast. Ocean & Coastal Management 75, 63-71.
| Crossref | Google Scholar |
Patenaude NJ, Richardson WJ, Smultea MA, Koski WR, Miller GW, Würsig B, Greene CR, Jr (2002) Aircraft sound and disturbance to bowhead and beluga whales during spring migration in the Alaskan Beaufort Sea. Marine Mammal Science 18, 309-335.
| Crossref | Google Scholar |
Pelayo-González L, Herra-Miranda D, Pacheco-Polanco JD, Guzmán HM, Goodman S, Oviedo L (2022) Decreases in encounter rate of endangered Northeast Pacific humpback whales in southern Costa Rica: possible changes in migration pattern due to warming events. Frontiers in Marine Science 9, 927276.
| Crossref | Google Scholar |
Pitman RL, Totterdell JA, Fearnbach H, Ballance LT, Durban JW, Kemps H (2015) Whale killers: prevalence and ecological implications of killer whale predation on humpback whale calves off Western Australia. Marine Mammal Science 31, 629-657.
| Crossref | Google Scholar |
Ransome N, Bejder L, Jenner M, Penfold G, Brosig VJ, Kitson C, Skjothaug R, Neilson E, Loneragan NR, Smith JN (2022) Observations of parturition in humpback whales (Megaptera novaeangliae) and occurrence of escorting and competitive behavior around birthing females. Marine Mammal Science 38, 408-432.
| Crossref | Google Scholar |
Rasmussen K, Palacios DM, Calambokidis J, Saborío MT, Dalla Rosa L, Secchi ER, Steiger GH, Allen JM, Stone GS (2007) Southern Hemisphere humpback whales wintering off Central America: insights from water temperature into the longest mammalian migration. Biology Letters 3, 302-305.
| Crossref | Google Scholar | PubMed |
Russell G, Cagnazzi D, Colefax A, Sprogis KR, Christiansen F (2023) Cost of migration and migratory timing in Western Australian humpback whales. Marine Mammal Science 40, e13074.
| Crossref | Google Scholar |
Salgado Kent CP, Jenner C, Jenner M, Bouchet P, Rexstad E (2012) Southern Hemisphere breeding stock D humpback whale population estimates from North West Cape, Western Australia. Journal of Cetacean Research and Management 12, 29-38.
| Crossref | Google Scholar |
Sleeman JC, Meekan MG, Wilson SG, Jenner CKS, Jenner MN, Boggs GS, Steinberg CC, Bradshaw CJA (2007) Biophysical correlates of relative abundances of marine megafauna at Ningaloo Reef, Western Australia. Marine and Freshwater Research 58, 608-623.
| Crossref | Google Scholar |
Smith JN, Kelly N, Childerhouse S, Redfern JV, Moore TJ, Peel D (2020) Quantifying ship strike risk to breeding whales in a multiple-use marine park: the Great Barrier Reef. Frontiers in Marine Science 7, 67.
| Crossref | Google Scholar |
Sprogis KR, Parra GJ (2022) Coastal dolphins and marine megafauna in Exmouth Gulf, Western Australia: informing conservation management actions in an area under increasing human pressure. Wildlife Research 50, 435-450.
| Crossref | Google Scholar |
Sprogis KR, Bejder L, Hanf D, Christiansen F (2020a) Behavioural responses of migrating humpback whales to swim-with-whale activities in the Ningaloo Marine Park, Western Australia. Journal of Experimental Marine Biology and Ecology 522, 151254.
| Crossref | Google Scholar |
Sprogis KR, Videsen S, Madsen PT (2020b) Vessel noise levels drive behavioural responses of humpback whales with implications for whale-watching. eLife 9, e56760.
| Crossref | Google Scholar | PubMed |
Stack SH, Sprogis KR, Olson GL, Sullivan FA, Machernis AF, Currie JJ (2021) The behavioural impacts of commercial swimming with whale tours on humpback whales (Megaptera novaeangliae) in Hervey Bay, Australia. Frontiers in Marine Science 8, 696136.
| Crossref | Google Scholar |
Stamation KA, Croft DB, Shaughnessy PD, Waples KA, Briggs SV (2010) Behavioral responses of humpback whales (Megaptera novaeangliae) to whale-watching vessels on the southeastern coast of Australia. Marine Mammal Science 26, 98-122.
| Crossref | Google Scholar |
Videsen SKA, Bejder L, Johnson M, Madsen PT (2017) High suckling rates and acoustic crypsis of humpback whale neonates maximise potential for mother–calf energy transfer. Functional Ecology 31, 1561-1573.
| Crossref | Google Scholar |
von Hammerstein H, Setter RO, van Aswegen M, Currie JJ, Stack SH (2022) High-resolution projections of global sea surface temperatures reveal critical warming in humpback whale breeding grounds. Frontiers in Marine Science 9, 837772.
| Crossref | Google Scholar |
Whitehead H (1983) Structure and stability of humpback whale groups off Newfoundland. Canadian Journal of Zoology 61, 1391-1397.
| Crossref | Google Scholar |
Worton BJ (1989) Kernel methods for estimating the utilization distribution in home-range studies. Ecology 70, 164-168.
| Crossref | Google Scholar |
Yablokov AV (1994) Validity of whaling data. Nature 367, 108.
| Crossref | Google Scholar |