

Where there’s smoke, there’s cats: long-unburnt habitat is crucial to mitigating the impacts of cats on the Ngarlgumirdi, greater bilby (Macrotis lagotis)
Harry A. Moore A B * , A G , Ben Corey A H , Karen Bettink A , Fiona M. Carpenter A , Ruth McPhail A , Tracy Sonneman A and Bruce Greatwich A
A B * , A G , Ben Corey A H , Karen Bettink A , Fiona M. Carpenter A , Ruth McPhail A , Tracy Sonneman A and Bruce Greatwich A A
B
C
D
E
F
G
H
Abstract
The decline of the greater bilby (Macrotis lagotis), or Ngarlgumirdi (Yawuru), like other critical-weight range Australian mammals, is believed to be primarily due to the synergetic impacts of predation by feral cats and foxes, habitat disturbance caused by large introduced herbivores, and increases in the frequency and intensity of wildfires. Although it has been demonstrated that low-intensity prescribed burning mosaics in some habitats have the potential to benefit mammals, including Ngarlgumirdi, by creating habitat with sufficient vegetation cover, the contributions of specific fire-mosaic attributes to Ngarlgumirdi persistence remain unclear.
To elucidate the impacts of fire-mosaic attributes on the occupancy of Ngarlgumirdi on the Dampier Peninsula.
We used 2-ha sign-plot data collected by four Indigenous Ranger groups, in combination with 20 years of satellite-derived fire-history information to investigate the multiscale impacts of fire attributes on Ngarlgumirdi and feral cats (Felis catus) on the Dampier Peninsula in the West Kimberley region, a large, unfenced landscape in the most fire-prone section of the Ngarlgumirdi’s current range.
We found that Ngarlgumirdi was more common in areas that had a higher proportion of habitat that had not burnt for at least 3 years, whereas feral cats were less prevalent in these areas. Similarly, Ngarlgumirdi was less likely to occur in landscapes affected by frequent fires, whereas cats were more common there.
Our findings have highlighted the importance of decreasing fire frequency and increasing the extent of long-unburnt habitats (>3 years) for preserving Ngarlgumirdi on the Dampier Peninsula and mitigating ecological damage inflicted by feral cats. Findings were consistent across spatial scales (1-, 3-, 5- and 10-km radius from each monitoring site).
These results have demonstrated the potential of fire management to increase native species resilience in the absence of direct feral cat control methods. Further, they support a recent cross-tenure initiative led by Traditional Owners to implement fire management that aims to reduce large, frequent high-severity wildfires and increase areas of long-unburnt vegetation on the Dampier Peninsula.
Keywords: applied ecology, biodiversity, conservation ecology, conservation management, ecological function, ecological systems, ecology, ecosystem management, natural resource management.
Introduction
Since European colonisation, Australia’s diverse mammal fauna has been affected by species loss and high rates of population decline (Woinarski et al. 2015), particularly those species in the small to medium size range (Johnson and Isaac 2009). Although most known extinctions took place in southern and arid regions of Australia during the early years of colonisation (Woinarski et al. 2015), recent mammal declines in northern Australia’s tropical savannas have occurred in areas that were largely unaffected until much later (McKenzie 1981; Burbidge and McKenzie 1989; Woinarski et al. 2011; Ziembicki et al. 2013), and similarly appear most prevalent among species in the small to medium size range (35–5500 g) (Murphy and Davies 2014).
A broad suite of factors has been attributed to mammal declines in southern Australia, including predation by introduced foxes and feral cats (Abbott et al. 2014), changing fire regimes, and the spread of large-scale agriculture and introduced herbivores (Woinarski et al. 2015). However, declines in northern Australia remain less clearly understood (Stobo-Wilson et al. 2020). Although evidence suggests that feral cats play a role, the decline of northern Australian mammals in the late 20th century is likely to be attributable to a combination of factors (Woinarski et al. 2011; Davies et al. 2020; Radford et al. 2021). One such factor is the frequent, high-intensity fires that have become increasingly common in northern Australia since the cessation of traditional Indigenous burning practices (Santos et al. 2022). Although the effects of these fires on mortality may be modest (Jolly et al. 2022), they have been shown to reduce the availability of resources, particularly for species that rely on vegetation cover. Moreover, the interaction between predators and fire further complicates the survival of these species, because altered habitats from fire events can make it easier for predators such as feral cats to hunt (McGregor et al. 2015), thereby exacerbating the impact on native fauna (Geary et al. 2020; Doherty et al. 2022).
Although there is evidence that supports the concept of predation as a primary driver of northern Australian mammal declines, with changed disturbance regimes exacerbating impacts (Griffiths et al. 2015; Lawes et al. 2015; Leahy et al. 2016; Radford et al. 2021), there is still much to be understood from a management perspective (Griffiths and Brook 2014). For example, although it has been demonstrated that low-intensity prescribed burning mosaics in some habitats have the potential to benefit mammals by reducing wildfires and creating habitat with sufficient vegetation cover (Davies et al. 2018; Radford et al. 2020), the contributions of specific fire-mosaic attributes to species persistence remain unclear. Further, whereas the scale at which these attributes are most important has been examined for some species (e.g. Legge et al. 2019), for the majority of species this detail remains poorly understood. Improving our understanding of these factors is crucial from a conservation management perspective to design effective interventions for threatened species across Australia’s north.
The greater bilby (Macrotis lagotis) is a medium-sized burrowing marsupial that was once widespread in Australia’s arid and semi-arid regions. The species is now restricted to a northern subset of its former range (DCCEEW 2023) and is listed as vulnerable by the IUCN (Burbidge and Woinarski 2016). The decline of the bilby, like other critical weight-range Australian mammals (Woinarski et al. 2015), is believed to be primarily due to impacts of predation (Southgate 1990), which are thought to be exacerbated by grazing from introduced herbivores, and frequency and intensity of wildfire (Abbott 2001; Doherty et al. 2015a; DCCEEW 2023), both of which can cause changes to habitat and resource availability through modification of vegetation structure, composition and cover (Bowman et al. 2020). Further understanding of the interactive effects of fire and predation is a research priority for the bilby (Cramer et al. 2016), particularly in the most fire-prone areas of its range where the synergetic impacts from these threats are likely to be greatest. Previous research investigating these threats is mostly limited to arid areas of the species range (Southgate 1990; Southgate et al. 2007), although see Dawson (2017). Results suggest that whereas bilby occurrence tends to be negatively correlated with predators such as the fox, the effect of fire on its distribution appears relatively weak. However, there does appear to be a preference for habitat located in close proximity to burnt areas in arid systems (Southgate et al. 2007), which is likely to be due to the high availability of fire-promoted food plants (Southgate and Carthew 2006). Whether bilbies respond similarly in habitats that burn more frequently, or how fire influences their vulnerability to predators in these environments, remain unclear.
The Dampier Peninsula in the Kimberley region of north-western Western Australia represents the most tropical extent of the current range of the bilbies, receiving an average of 650–850 mm of rain per year (BOM 2022), and is extremely prone to fire (NAFI 2022). Although the response of bilbies to fire has previously been investigated on part of the Dampier Peninsula (Dawson 2017), a more broad-scale approach is required to assist in developing fire management guidelines for bilbies in this area, which has been identified as a priority action in the most recent greater bilby recovery plan (DCCEEW 2023). Evidence from nearby caves suggests that Aboriginal peoples have been living in this region for at least 46,000 years (Balme et al. 2019). Like in other parts of Australia prior to European colonisation (Enright and Thomas 2008), fire-management practices implemented by Aboriginal peoples are likely to have played a significant role in shaping the ecosystems of the Kimberley (Vigilante 2001). This includes creating fine-scale habitat mosaics and limiting the extent of large-scale wildfires, both of which can be favourable for native mammals (Bowman 1998). Although contemporary burning patterns in the Kimberley are still influenced by people (Vigilante et al. 2004), the large-scale migration of Aboriginal pastoral workers and their families into towns and communities following the introduction of equal pay in 1968 (Jebb 2002) has largely removed traditional burning practices from the landscape. Concurrently, the demand for labour as part of the Dampier Peninsula’s pearling industry contributed to social disruption within Aboriginal communities, and thus also the pre-existing fire regime, because people were often forcibly removed from their lands to work in pearling or were displaced by the industry infrastructure and activities (Nilsson 2017). These developments, alongside other social factors, have contributed to shaping current fire regimes in large parts of the Kimberley, including the Dampier Peninsula, which became largely characterised by extensive mid-to-late season wildfires, often reoccurring on a 1–3-year cycle (Russell-Smith et al. 2003a).
In addition to human life and property, large-scale wildfire on the Dampier Peninsula threatens species and communities that are of high cultural and ecological value (Wysong et al. 2021). For example, an ecological community endemic to the Dampier Peninsula, the monsoon vine thicket, is now listed as Endangered nationally, in part owing to increased fire frequency in areas where it occurs (DCCEEW 2013). For thousands of years, the monsoon vine thickets have provided Aboriginal peoples with food, tools, medicines, shelters, and meeting places, while also providing important habitat for native fauna (Lindsay et al. 2022). Like the monsoon vine thickets, bilby, hereafter referred to as Ngarlgumirdi, the name used by Yawuru people from the Broome region, is an ecologically important species that has likely been adversely affected by changing fire regimes on the Dampier Peninsula. Ngarlgumirdi holds significant cultural importance to the Traditional Owners of the Dampier Peninsula, as well as other sections of its current and former range, and frequently features in the ancestral narratives often referred to as The Dreaming (Skroblin et al. 2022; DCCEEW 2023). Protecting Ngarlgumirdi, along with other important species and communities from the damaging impacts of wildfire is a priority for Traditional Owners on the Dampier Peninsula. Achieving this aim requires a coordinated approach, such as that implemented by the Dampier Peninsula Fire Working Group, a collaboration led by Indigenous Ranger groups, pastoralists, fire managers and other stakeholders, that aims to improve fire regimes on the Dampier Peninsula (Wysong et al. 2021). To help focus these efforts so as to improve Ngarlgumirdi persistence, we investigated the multiscale impacts of different fire attributes on Ngarlgumirdi across the Dampier Peninsula, and also examined how these fire attributes influence the occurrence of feral cats, an important predator of the species (Moseby et al. 2011). The implications of the patterns observed are discussed in relation to enhancing Ngarlgumirdi conservation through improved fire management within the study area.
Methods
Study area
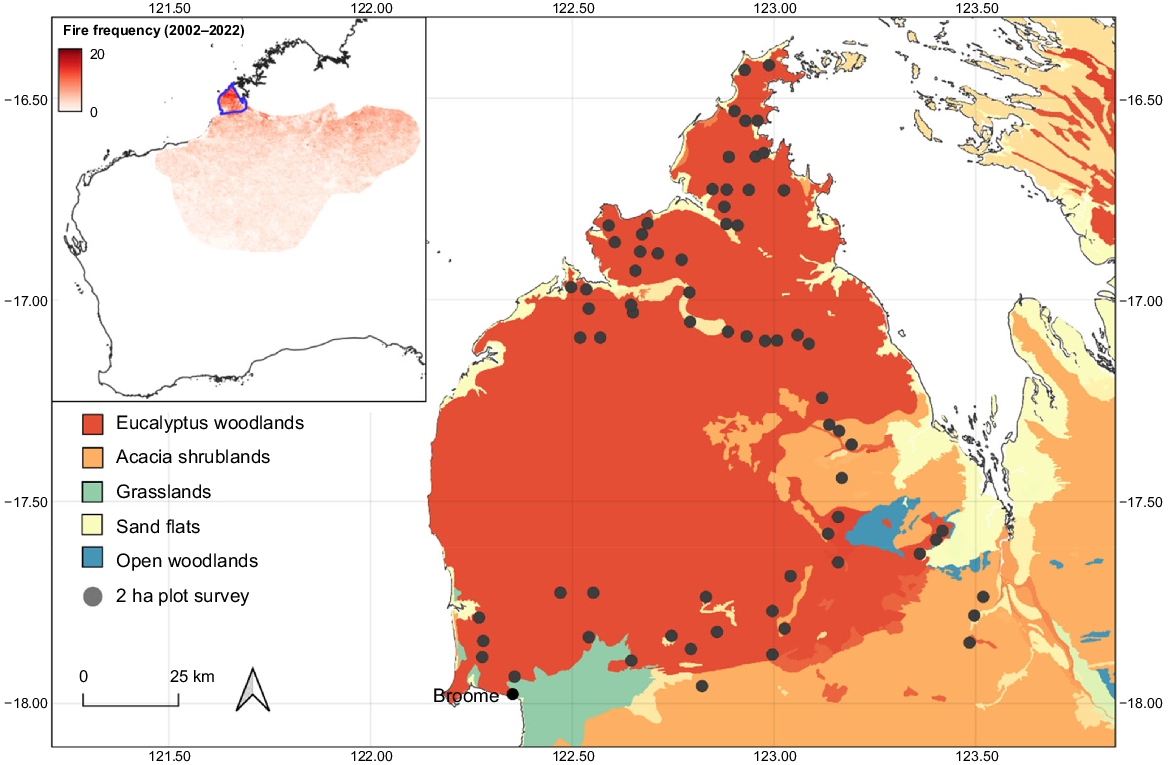
The study area was located on the Dampier Peninsula within the western Kimberley region of north-western Australia (Fig. 1). Covering ~18,000 km2, the area comprises Indigenous Protected Areas, pastoral leases, unallocated Crown Land as well as a small nature reserve (Coulomb Point). This area represents the most north-western section of the Ngarlgumirdi’s contemporary and historic range (Southgate 1990), and also the section of their contemporary range that receives the most annual rainfall (BOM 2022) and burns the most frequently (NAFI 2022). The majority of land within the study region is subject to a determined or registered Native Title Claims (National Native Title Tribunal 2023). The area experiences a tropical monsoonal climate, with the majority of rainfall falling between December and March. Annual rainfall totals across the peninsula typically range from ~650 mm in the south to ~850 mm in the north (BOM 2022).
Location of 2-ha sign plots located on the Dampier Peninsula in north-western Australia. Inset shows fire frequency across the Ngarlgumirdi’s (Macrotis lagotis) contemporary range (excluding the isolated Queensland population), with the study area outlined in blue.
Vegetation on the Dampier Peninsula is largely dominated by eucalypt woodlands (Eucalyptus tectifica, Corymbia polycarpa, Corymbia grandifolia) and acacia shrublands (Acacia tumida, A. eriopoda) over a sparse understorey comprising herbs, and mixed annual and perennial grasses (Sorghum stipoideum, Chrysopogon pallidus, Senna notabilis, Ptilotus corymbosus) (Kenneally et al. 1996; Dziminski and Bettink 2017) (Fig. 2). Within these communities, Ngarlgumirdi tends to be associated with species that harbour root-dwelling larvae (A. colei, A. eriopoda, A. tumida, Grevillea refracta, Dodonaea hispidula, S. notabilis), which are an important component of its diet (Dziminski and Bettink 2017; Southgate et al. 2019). These habitat types provide a mix of more open areas for mobility and sufficient vegetation cover for protection from predators.
Dominant vegetation types on the Dampier Peninsula in north-western Australia. (a) Eucalyptus woodland, comprising Dampier bloodwood (Corymbia sp.) over sparse to dominant blade grass (Chrysopogon pallidus) and Jacquemontia sp. (b) Acacia shrubland, comprising mixed acacia (Acacia tumida, A.colei) over tussock grass.

Fire management on the Dampier Peninsula
Fire management on the Dampier Peninsula is primarily the responsibility of landowners, with state-owned land managed by designated authorities such as the Department of Biodiversity, Conservation and Attractions (DBCA; DPLH 2022). Natural events such as lightning, as well as human factors, including campfires, discarded cigarettes, and deliberate arson, are the main contributors to wildfire ignition on the Dampier Peninsula. The intensity and spread of these fires are influenced by available fuel and moisture content; however, under certain conditions, fires can burn over 6500 km2 in the space of a single month (NAFI 2022). As the region navigates the challenges of managing fires of this scale, there is an ongoing effort to reintegrate traditional burning practices into broad-scale fire management.
The Dampier Peninsula Fire Working Group (DPFWG) was established in 2016 as a collaborative initiative that brings together Traditional Owners, Indigenous Ranger groups, government agencies, conservation and non-profit organisations, as well as the pastoral and natural resources industries (Wysong et al. 2021). Emphasising Aboriginal-led management as a core principle, the group is dedicated to enhancing fire management on the Peninsula through coordinated training, skills-sharing, communication, and prescribed burning efforts. A focus on Aboriginal leadership ensures that traditional knowledge and practices play a central role in shaping fire-management strategies, reinforcing the importance of culturally informed conservation efforts.
Dampier Peninsula bilby-offset project
In 2016, Main Roads Western Australia provided offset funding (associated with the Cape Leveque Road upgrade) to the Western Australian Department of Biodiversity Conservation and Attractions, to undertake a 3-year project to survey, monitor and commence adaptive management of threats to Ngarlgumirdi populations across the Dampier Peninsula, including fire (Dziminski and van Leeuwen 2019). Partners on this project included Nyul Nyul, Bardi Jawi Oorany, Nyikina Mangala Indigenous Rangers, and Yawuru Country Managers.
Data used in this study were collected during the first component of this project, which, in addition to measuring the influence of fire-mosaic attributes on Ngarlgumirdi occurrence, aimed to map Ngarlgumirdi occurrence across the Dampier Peninsula. Other project objectives included implementing fine-scale fire management at a select number of sites where Ngarlgumirdi was confirmed to be present, in conjunction with an annual abundance-monitoring program using a scat-genotyping approach (Dziminski and van Leeuwen 2019).
Presence data
Ngarlgumirdi and feral cat-presence data were collected at 69 sites across the study area, between April and October 2019. The average distance between a site and its nearest neighbouring sites was 5.7 km (minimum = 2.5 km; Fig. 1). Although Ngarlgumirdi is known to move distances of up to 5 km between burrows (DCCEEW 2023), the majority of sites were deemed to be suitably independent from one another. Collaborative planning sessions for sign-plot locations were conducted with Nyul Nyul, Bardi Jawi, Nyikina Mangala Rangers and Yawuru Country Managers on the basis of their local knowledge of suitable habitats. The location of sites was stratified according to fire frequency by using data collected from the North Australia Fire Information Service (NAFI 2022).
Species presence at sites was verified using the 2-ha sign-plot approach, which involved trained sign observers searching a 2-ha area (100 m × 200 m) for secondary signs, including tracks, diggings and scats, for 25 min (Moseby et al. 2009) per visit. The 2-ha sign-plot approach is currently one of the most widely used methods for detecting Ngarlgumirdi (Bradley et al. 2015; Southgate et al. 2019; Dziminski et al. 2021a, 2021b), especially by Indigenous Ranger groups (Paltridge 2016). In addition to Ngarlgumirdi, this survey method is also effective at detecting a range of other vertebrates, including introduced predators such as feral cats (Moseby et al. 2009).
Sites were surveyed at least twice by Nyul Nyul, Bardi Jawi, Nyikina Mangala Rangers and Yawuru Country Managers between March and October 2019, with most sites being surveyed four times (n total visits = 259; Fig. 3). Repeat visits were used to track changes in site occupancy at different post-fire successional stages. The average interval between surveys was 65 days. Only fresh sign was used to record presence.

Environmental data
Fire data used in this study comprised Moderate Resolution Imaging Spectroradiometer (MODIS; 250-m resolution) vector data available from the North Australia Fire Information service (http://firenorth.org.au). The status of data pixels (burnt or unburnt) was measured on a monthly basis from the Year 2000 up until the date a 2-ha plot was completed. Four fire-history attributes were recorded for each site, and each visit, by using MODIS data. Fire attributes were pyrodiversity, fire frequency, as well as proportion of site recently burnt (<1 year post fire) and long unburnt (>3 years post fire) (Table 1). Long unburnt habitat was defined as an area that was last burnt more than 3 years prior, in accordance with the study by Wysong et al. (2021) conducted in the same study area.
| Attribute | Method | Justification | |
|---|---|---|---|
| Pyrodiversity | Sum of post-successional vegetation ages within a landscape. | Pyrodiversity refers to the diversity of fire regimes within a landscape. Previous research has suggested that pyrodiversity can be linked to aspects of biodiversity (Martin and Sapsis 1992; Tingley et al. 2016), as well as individual species habitat suitability (Senior et al. 2021), although not always (Jones and Tingley 2022). | |
| Fire frequency | The number of years in which a landscape burnt between the Year 2000 and the time of a 2-ha plot survey. | Fire frequency is an important predictor of fauna biodiversity in northern Australia (Griffiths et al. 2015; Ondei et al. 2020; Andersen 2021), including mammal species (Davies et al. 2018; Einoder et al. 2018). | |
| Proportion recently burnt (<1 year post-fire) | The proportion of a landscape that was burnt up to 1 year before a 2-ha plot survey. | The proportion of recently burnt habitat within a landscape has previously been demonstrated to have a strong influence on mammal presence in northern Australia (Radford et al. 2015). In central Australia, proximity to recently burnt habitat is associated with Ngarlgumirdi occurrence (Southgate et al. 2007) | |
| Proportion longer unburnt (>3 years post-fire) | The proportion of a landscape that was last burnt more than 3 years before a 2-ha plot survey, following Wysong et al. (2021) | Long-unburnt habitat has repeatedly been demonstrated to be important to the persistence of northern Australian mammal species (Davies et al. 2018; Legge et al. 2019; Shaw et al. 2021; Shaw et al. 2021; von Takach et al. 2022) |
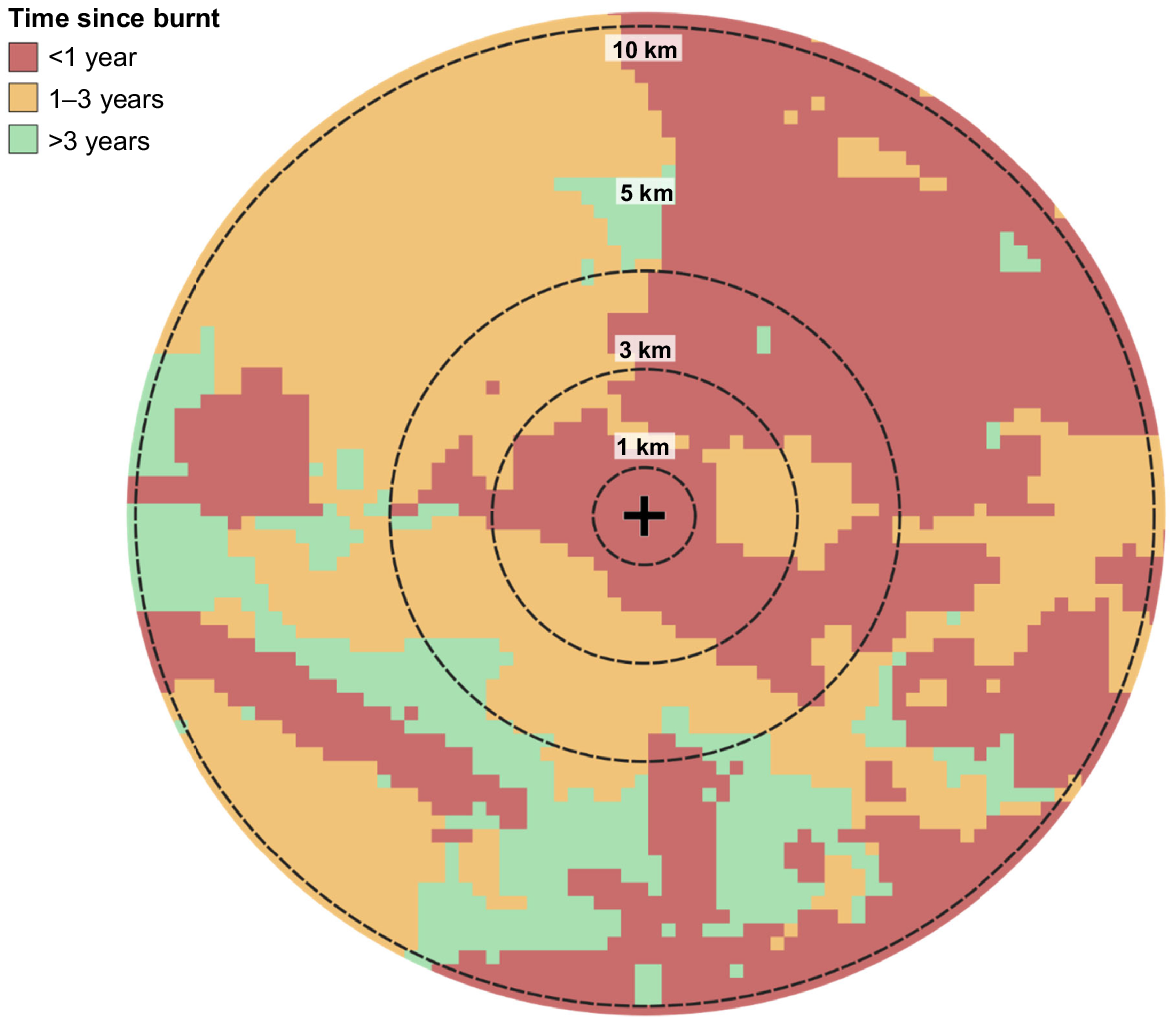
Research has shown that fire attributes can have varying impacts on species depending on the scale at which they are measured (Nimmo et al. 2019; Wan et al. 2020). To account for this, we measured fire attributes at multiple scales for each plot location (Fig. 4). We did this by creating buffers of different sizes at each plot location, and then clipping fire data to those buffers, following Radford et al. (2021). Buffer sizes were 1 km (A = 3.14 km2), 3 km (28.26 km2), 5 km (78.5 km2) and 10 km (314 km2).
Spatial scales of buffers used to assess fire attributes on the Dampier Peninsula in northern Australia. In this example, time since burnt is used. The black cross represents the centre of a 2-ha plot location.
Other environmental factors likely to influence the occurrence of Ngarlgumirdi and feral cats, including dominant vegetation type and recent rainfall (previous 6-month total) were also recorded for each site, at each scale. Vegetation data were sourced from the National Vegetation Information System (NDVI 2020) at 100 m × 100 m resolution. The majority of sites (n = 65) were dominated by either Eucalyptus woodland or Acacia shrublands. The four sites that did not fall into either of these two categories were removed because of inadequate sampling replication. Although substrate is known to be an important predictor of Ngarlgumirdi occurrence, it was not included in models here, given that almost all sites were located on calcareous and siliceous sands (Rudosols), which are deemed suitable substrate for Ngarlgumirdi occurrence.
Data analysis
To measure the effect of fire attributes on species’ occurrences, we fit binomial generalised linear mixed-effects models by using the lme4 package (Bates et al. 2015) in R ver. 4.1.2 (R Core Team 2021). A global model was fit for each fire attribute (pyrodiversity, fire frequency, proportion recently burnt, and proportion longer unburnt), at each scale (1 km, 3 km, 5 km, 10 km), for each species (Ngarlgumirdi, feral cat). To avoid collinearity issues, separate models were fitted for each fire attribute. This resulted in a total of 16 models per species (four fire attributes × four scales). The presence or absence of species at each site on each visit was set as the response variable for all models.
In addition to fire attributes, each global model included vegetation type and recent rainfall (previous 6 months) as fixed effects. The site ID was included in the models as a random factor to account for sampling replication within sites. Fixed effects were deemed to have a strong effect on species occurrence when estimate confidence intervals did not overlap zero (Nakagawa and Cuthill 2007).
Results
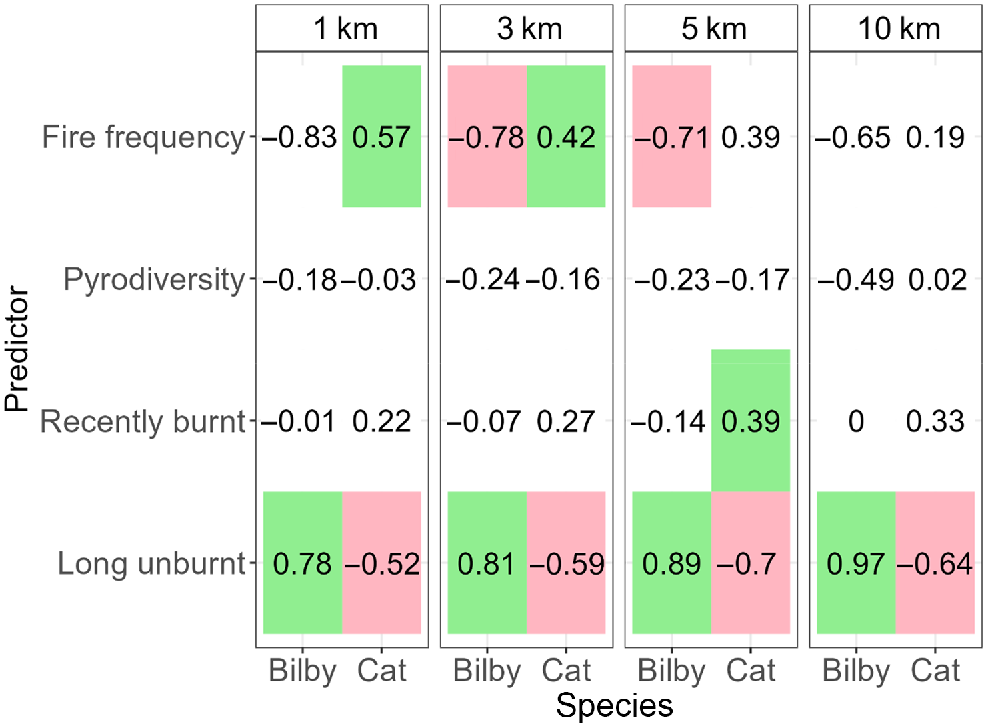
Ngarlgumirdi was recorded at 18.8% of the sites. Ngarlgumirdi presence was influenced by the proportion of long-unburnt habitat, with the largest effect being observed at the largest scale (10 km) (Fig. 5). Here, the likelihood of Ngarlgumirdi presence increased from absent (0%) when there was no >3 years unburnt habitat within a 10-km radius of the site, to 24.2% when the entire area within a 10-km radius site was >3 years unburnt (Figs 5, 6). Fire frequency also influenced Ngarlgumirdi presence at the 3-km and 5-km scales. At the 3-km scale, presence decreased from 34.8% at sites that had been burnt seven times in the previous 20 years to 0% at sites that had been burnt 20 times in the same period (Fig. 6). Predictions of Ngarlgumirdi occurrence were associated with wide confidence intervals, which were likely to be due to the limited sample size.
Model estimates from generalised linear mixed-effects models testing the influence of fire attributes on the presence of Ngarlgumirdi and feral cats on the Dampier Peninsula in north-western Australia. Coloured cells show estimates where the effect was significant (P < 0.05). Red cells indicate a negative effect, and green cells indicate a positive effect.
Model predictions from generalised linear mixed-effects models testing the influence of fire attributes on the presence of Ngarlgumirdi and feral cats on the Dampier Peninsula in north-western Australia. Grey shading indicates 95% confidence intervals. Each row shows predictions from models using data collected at varying distances from 2-ha plots (1 km, 3 km, 5 km, 10 km).
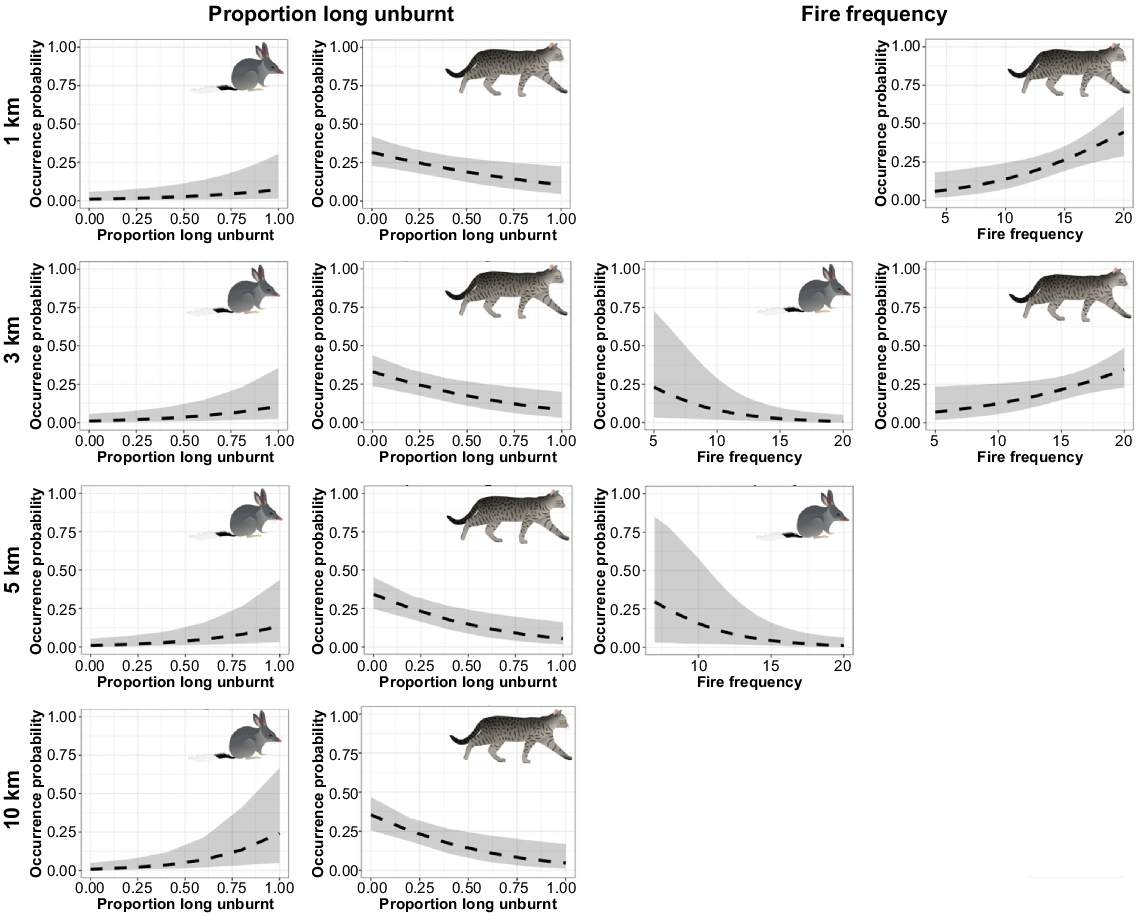
Feral cats were recorded at 59.4% of the sites. Feral cat presence was also related to the proportion of long-unburnt habitat, but the effect was inversed when compared with Ngarlgumirdi (Fig. 5). The likelihood of feral cat presence decreased by 30% from sites with no surrounding long-unburnt habitat to sites surrounded by 100% long-unburnt habitat. This effect was consistent across spatial scales (Fig. 6). The likelihood of feral cat presence increased with an increasing fire frequency at the 3-km and 5-km scales, as well as the proportion of recently burnt habitat at the 5-km scale. Pyrodiversity, annual rainfall and dominant vegetation type had no significant influence on the presence of Ngarlgumirdi or feral cats at any scale (Supplementary Tables S1, S2, S3, S4).
Discussion
Our findings indicated that fire frequency and the extent of long-unburnt habitat (>3 years) are important factors in predicting the presence of both Ngarlgumirdi and feral cats across multiple spatial scales in the highly fire-prone landscape of the Dampier Peninsula. Ngarlgumirdi was more frequently detected in areas with a high proportion of habitat that had not been burnt for at least 3 years, whereas feral cat detections were lower here. Similarly, frequent fires tended to be a deterrent for Ngarlgumirdi, but not for feral cats. These results support findings from global studies that suggest that trends of increasing fire frequency and spatial extent of fires have contributed to declines observed across multiple taxa, presumably owing to an increase in predation and decrease in resource availability (Engstrom 2010; Kelly et al. 2020). These results provide important context for fire practitioners seeking to implement fire regimes that promote habitat suitable for the conservation of Ngarlgumirdi within fire-prone areas of their range, particularly on a landscape scale outside of invasive-predator-free enclosures.
How does fire influence the distribution of Ngarlgumirdi on the Dampier Peninsula?
Previous studies that examined the interaction between Ngarlgumirdi and fire have mostly focused on arid environments, such as in the Tanami Desert. In these drier locations, there is evidence to suggest that Ngarlgumirdi occurs more frequently in close proximity to recently burnt habitat (Southgate et al. 2007), likely owing to the increased availability of post-fire ephemeral grasses (Southgate and Carthew 2007), which can make up a substantial proportion of their diet in arid areas (Southgate and Carthew 2006). Whereas one previous study has investigated the response of Ngarlgumirdi to fire on the Dampier Peninsula, finding limited evidence of a response (Dawson 2017), our research expands on this methodology by using a landscape approach across multiple scales and incorporating a larger study area.
In contrast to arid environments where Ngarlgumirdi is known to prefer recently burnt habitat, we found that Ngarlgumirdi on the Dampier Peninsula was more likely to occur in areas that had not been burnt for at least 3 years (Southgate and Carthew 2007). The observed disparity in habitat preference between arid ecosystems and the current study area may be attributed to differences in the post-fire composition and structure of the vegetation in the higher-rainfall Dampier woodlands from those in arid grasslands. For example, in arid landscapes, post-fire spinifex grasslands are usually characterised by low vegetation cover that lasts for 1–3 years, thereby creating an open environment where Ngarlgumirdi can forage for fire-promoted plants such as Yakirra. In contrast, in more tropical climates such as the Dampier Peninsula, burnt landscapes are often rapidly colonised by fast-growing annual grasses such as Sorghum stipoideum (Radford et al. 2015; Radford and Fairman 2015). This is likely to be especially true for fires that occur in the late-dry season, given the wettest part of the year generally follows soon after, promoting rapid vegetation growth. By dominating the understorey, these annual grasses are likely to reduce the availability of important perennials such as Yakirra spp. and impede the movement of Ngarlgumirdi while foraging. This result highlights the value of undertaking research across the full range of a wide-ranging habitat generalist such as Ngarlgumirdi to better understand ecological associations that may differ across their range.
We found that Ngarlgumirdi favoured habitat that had been less frequently burnt. This result is supported by previous research in northern Australia, where mammal declines have often been linked to high fire frequencies (Woinarski et al. 2010; von Takach et al. 2020; Einoder et al. 2023). High fire frequencies alter vegetation communities by promoting species with high fire tolerance, and displacing species that are fire sensitive (Russell-Smith et al. 2003b; Rossiter et al. 2003; Miller et al. 2010). In addition to killing invertebrate prey and outcompeting native grasses and herbs, which may be important food sources for Ngarlgumirdi (Gibson 2001), these changes can substantially increase the flammability of landscapes, further increasing fire frequency, intensity and scale (Russell-Smith et al. 2003b). Similar effects have been observed within the study area (Wysong et al. 2021), with major increases in highly flammable annual Sorghum, and this is thought to be at least partly responsible for the disappearance of a long-term local Ngarlgumirdi population. In this example, an intense late dry-season fire replaced stands of Acacia tumida, a plant species important for harbouring cossid larvae which are preyed on by Ngarlgumirdi, with annual sorghum grasslands. These changes in the vegetation led to a repeat late dry-season fire event 2 years following the fire, after which Ngarlgumirdi was no longer present at the site (B. Greatwich, unpubl. data).
How does fire influence the distribution of feral cats on the Dampier Peninsula?
In contrast to Ngarlgumirdi, we found that the likelihood of feral-cat presence increased as the extent of long-unburnt habitat decreased, and fire frequency increased. Although the literature suggests that the response of feral cats to fire is variable and dependent on habitat type (Doherty et al. 2015b, 2022), there is evidence that cats do target areas frequently affected by disturbance in tropical savannas (McGregor et al. 2014; Davies et al. 2020). The most widely accepted explanation for this behaviour is related to prey access, because feral cats can locate and pursue prey more easily in structurally simple habitat (Geary et al. 2020). For example, previous studies conducted in the savannas of the Kimberley and Cape York Peninsula regions by McGregor et al. (2014, 2015) demonstrated that feral cats exhibit improved hunting efficiency in grassland and recently burnt habitats. Similarly, Trewella et al. (2023) found that habitat use by feral cats was most frequent in areas with high fire frequencies and low tree basal area. It is plausible, therefore, that mature Acacia shrubland and eucalypt woodland (>3 years post fire) limit the hunting efficiency of feral cats by providing complex vegetation cover for Ngarlgumirdi. In addition to the immediate reduction in vegetation cover caused by fire, long-term reductions in shrubland/woodland structural complexity caused by repeated fires at short intervals are also likely to benefit feral cats, likely to the detriment of Ngarlgumirdi (Davies et al. 2020; Stobo-Wilson et al. 2020). It is important to note that the behaviour of feral cats in response to fire is likely to vary in more arid sections of the Ngarlgumirdi’s range. For instance, in contrast to our findings, a recent study in arid Australia observed that feral cats not only sought out habitats indicating the presence of native prey, including Ngarlgumirdi, through signs like foraging digs, burrows, and scats, but they also frequently occupied areas with dense vegetation cover (Moseby and McGregor 2022). These insights underscore the need for adaptive management strategies that consider the variable responses of feral cats across different habitats.
Management implications
Our findings have highlighted the importance of decreasing the frequency of large-scale wildfire and increasing the amount of long-unburnt habitat (>3 years) for the preservation of Ngarlgumirdi and the mitigation of ecological damage inflicted by feral cats on the Dampier Peninsula. For example, on the basis of model predictions at the 3-km scale (a logistically feasible management unit), increasing the proportion of long-unburnt habitat in the landscape from 20% to 60% could almost triple the likelihood of Ngarlgumirdi occurrence, and almost halving the likelihood of feral-cat occurrence. Similarly, reducing fire frequency from once every 2 years to once every ~4 years more than triples the likelihood of Ngarlgumirdi occurrence, and halves the likelihood of feral-cat occurrence. These targets align directly with objectives established by the Dampier Peninsula Fire Working Group (DPFWG) (Wysong et al. 2021). A recent review of the project performance in relation to these objectives demonstrated that fire-management efforts in the region increased the extent of vegetation long unburnt from 18% in 2015 to 65% in 2020 (Wysong et al. 2021). In addition, the proportion of habitat burnt three or more times was halved over the same period. Our study supports the ongoing burning planning and practices being implemented by DPFWG that aim to increase the extent of long-unburnt vegetation on the Dampier Peninsula and reduce fire frequency.
Future research
Although our study has provided important insights into the roles of fire frequency and the extent of long-unburnt habitat on the distribution of Ngarlgumirdi and feral cats in the Dampier Peninsula, several avenues for future research are evident. One is investigating how changes in understorey and overstorey vegetation complexity influence the likelihood of Ngarlgumirdi occurrence. Although it is likely that these metrics are correlated with fire metrics (Fisher et al. 2003; Lynch et al. 2018; Rainsford et al. 2020), a more detailed understanding of how Ngarlgumirdi responds to vegetation structure per se could provide important insights into how habitat suitability can be better managed, particularly for populations located within pastoral leases where grazing pressure is a threat (Dziminski et al. 2021b). Incorporating burn timing and severity as additional predictors is also likely to offer a more nuanced understanding of how Ngarlgumirdi responds to fire in this region (Southgate and Carthew 2007). Both factors are likely to be strong determinants in the response of vegetation in terms of structure and composition (Russell-Smith and Edwards 2006). Moreover, since both fire severity and timing are key metrics used to gauge the success of current fire management in the region (Wysong et al. 2021), quantifying the response of Ngarlgumirdi to these variables could enhance our ability to tailor burn plans to improve Ngarlgumirdi conservation.
It is important to note that the 2-ha sign-plot method used in this study was initially developed for arid environments, where animal sign, particularly tracks, are more easily detected because of the predominance of open sandy areas. Although Ngarlgumirdi diggings and burrows are typically visible regardless of substrate, high vegetation cover within the study area may have reduced Ngarlgumirdi and feral-cat detectability at some sites. To overcome this issue, we suggest future studies couple plot surveys with camera-traps, so that species presence can be better verified.
Finally, improving our understanding of how Ngarlgumirdi diets vary with fire history, similar to what has already been undertaken in other parts of their range (Gibson 2001; Southgate and Carthew 2006), should also be a focus of future research. Integrating this knowledge into burn regimes could enable fire practitioners to actively manage Ngarlgumirdi food resources and expand the extent of suitable habitat available.
Declaration of funding
This project was supported by the Western Australian Department of Biodiversity, Conservation and Attractions as well as environmental offset funding provided by Main Roads Western Australia.
Acknowledgements
We acknowledge and pay respect to the Traditional Custodians of the lands where our research was conducted: the Bardi Jawi, Bindunbur, Jabirr Jabirr, Nyikina Mangala, and Yawuru peoples. We are grateful to the Kimberley Land Council, Rangelands NRM, WWF, Environs Kimberley for their support, advice and assistance on this project.
References
Abbott I (2001) The bilby Macrotis lagotis (Marsupialia: Peramelidae) in south-western Australia: original range limits, subsequent decline, and presumed regional extinction. Records-Western Australian Museum 20, 271-306.
| Google Scholar |
Andersen AN (2021) Faunal responses to fire in Australian tropical savannas: insights from field experiments and their lessons for conservation management. Diversity and Distributions 27, 828-843.
| Crossref | Google Scholar |
Balme J, O’connor S, Maloney T, Vannieuwenhuyse D, Aplin K, Dilkes-Hall IE (2019) Long-term occupation on the edge of the desert: Riwi Cave in the southern Kimberley, Western Australia. Archaeology in Oceania 54, 35-52.
| Crossref | Google Scholar |
Bates D, Mächler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. Journal of Statistical Software 67, 1-48.
| Crossref | Google Scholar |
BOM (2022) Average annual, seasonal and monthly rainfall. Available at http://www.bom.gov.au/jsp/ncc/climate_averages/rainfall/index.jsp
Bowman DMJS (1998) The impact of Aboriginal landscape burning on the Australian biota. New Phytologist 140, 385-410.
| Crossref | Google Scholar | PubMed |
Bowman DMJS, Kolden CA, Abatzoglou JT, Johnston FH, van der Werf GR, Flannigan M (2020) Vegetation fires in the Anthropocene. Nature Reviews Earth & Environment 1, 500-515.
| Crossref | Google Scholar |
Burbidge AA, McKenzie NL (1989) Patterns in the modern decline of western Australia’s vertebrate fauna: causes and conservation implications. Biological Conservation 50, 143-198.
| Crossref | Google Scholar |
Burbidge A, Woinarski JCZ (2016) Macrotis lagotis. The IUCN Red List of Threatened Species Available at.
| Crossref | Google Scholar |
Cramer VA, Dziminski MA, Southgate R, Carpenter FM, Ellis RJ, van Leeuwen S (2016) A conceptual framework for habitat use and research priorities for the greater bilby (Macrotis lagotis) in the north of Western Australia. Australian Mammalogy 39, 137-151.
| Crossref | Google Scholar |
Davies HF, McCarthy MA, Rioli W, Puruntatameri J, Roberts W, Kerinaiua C, Kerinauia V, Womatakimi KB, Andersen AN, Murphy BP (2018) An experimental test of whether pyrodiversity promotes mammal diversity in a northern Australian savanna. Journal of Applied Ecology 55, 2124-2134.
| Crossref | Google Scholar |
Davies HF, Maier SW, Murphy BP (2020) Feral cats are more abundant under severe disturbance regimes in an Australian tropical savanna. Wildlife Research 47, 624-632.
| Crossref | Google Scholar |
DCCEEW (2013) Approved conservation advice for the Monsoon vine thickets on the coastal sand dunes of Dampier Peninsula. Available at https://www.environment.gov.au/biodiversity/threatened/communities/pubs/105-conservation-advice.pdf
Doherty TS, Dickman CR, Nimmo DG, Ritchie EG (2015a) Multiple threats, or multiplying the threats? Interactions between invasive predators and other ecological disturbances. Biological Conservation 190, 60-68.
| Crossref | Google Scholar |
Doherty TS, Bengsen AJ, Davis RA (2015b) A critical review of habitat use by feral cats and key directions for future research and management. Wildlife Research 41, 435-446.
| Crossref | Google Scholar |
Doherty TS, Geary WL, Jolly CJ, Macdonald KJ, Miritis V, Watchorn DJ, Cherry MJ, Conner LM, González TM, Legge SM, Ritchie EG, Stawski C, Dickman CR (2022) Fire as a driver and mediator of predator–prey interactions. Biological Reviews 97, 1539-1558.
| Crossref | Google Scholar | PubMed |
DPLH (2022) BUshfire management plan guidance for the Dampier Peninsula. Available at chrome-extension://efaidnbmnnnibpcajpcglclefindmkaj/https://www.wa.gov.au/system/files/2022-02/Bushfire-Management-Plan-Guidance-for-the-Dampier-Peninsula.pdf
Dziminski M, Greatwich B, McPhail R, Carpenter F, Alai J, Lindsay M, Cherel V, Dick D, Cherel C, Chestnut R, Carter C, Green J, Charles R, Watson W, Watson A, Wallacy L, Green N, Millindee S, Raina K, Blythman M, Sercombe J, Giles D, Middleton P, Middleton J, Shaw S, Uhl J, Chunga K, Middleton M, Cooksey I, Godfrey N, Jessel S, Curtin D, Jnr BD, Ramsey C, Andrews T, Murray G, Lenmardi R, Surprise S, McGinty P, Thirkall R, Kogolo D, Thirkall R, Smiler E, Brown L, Murray K, Smiler E, Skinner K, Andrews J, Smith D, Murray C, Kylon P, Nuggett A, van Leeuwen S, Gibson L (2021b) Monitoring, mapping and safeguarding Kimberley bilbies. DBCA.
Einoder LD, Southwell DM, Lahoz-Monfort JJ, Gillespie GR, Fisher A, Wintle BA (2018) Occupancy and detectability modelling of vertebrates in northern Australia using multiple sampling methods. PLoS ONE 13, e0203304.
| Crossref | Google Scholar | PubMed |
Einoder LD, Fisher A, Hill BM, Buckley K, de Laive AH, Woinarski JCZ, Gillespie GR (2023) Long term monitoring reveals the importance of large, long unburnt areas and smaller fires in moderating mammal declines in fire-prone Savanna of northern Australia. Journal of Applied Ecology 60, 2251-2266.
| Crossref | Google Scholar |
Engstrom RT (2010) First-order fire effects on animals: review and recommendations. Fire Ecology 6, 115-130.
| Crossref | Google Scholar |
Enright NJ, Thomas I (2008) Pre-European fire regimes in Australian ecosystems. Geography Compass 2, 979-1011.
| Crossref | Google Scholar |
Fisher R, Vigilante T, Yates C, Russell-Smith J (2003) Patterns of landscape fire and predicted vegetation response in the North Kimberley region of Western Australia. International Journal of Wildland Fire 12, 369-379.
| Crossref | Google Scholar |
Geary WL, Doherty TS, Nimmo DG, Tulloch AIT, Ritchie EG (2020) Predator responses to fire: a global systematic review and meta-analysis. Journal of Animal Ecology 89, 955-971.
| Crossref | Google Scholar | PubMed |
Gibson LA (2001) Seasonal changes in the diet, food availability and food preference of the greater bilby (Macrotis lagotis) in south-western Queensland. Wildlife Research 28, 121.
| Crossref | Google Scholar |
Griffiths AD, Brook BW (2014) Effect of fire on small mammals: a systematic review. International Journal of Wildland Fire 23, 1034-1043.
| Crossref | Google Scholar |
Griffiths AD, Garnett ST, Brook BW (2015) Fire frequency matters more than fire size: testing the pyrodiversity–biodiversity paradigm for at-risk small mammals in an Australian tropical savanna. Biological Conservation 186, 337-346.
| Crossref | Google Scholar |
Johnson CN, Isaac JL (2009) Body mass and extinction risk in Australian marsupials: the ‘Critical Weight Range’ revisited. Austral Ecology 34, 35-40.
| Crossref | Google Scholar |
Jolly CJ, Dickman CR, Doherty TS, van Eeden LM, Geary WL, Legge SM, Woinarski JCZ, Nimmo DG (2022) Animal mortality during fire. Global Change Biology 28, 2053-2065.
| Crossref | Google Scholar | PubMed |
Jones GM, Tingley MW (2022) Pyrodiversity and biodiversity: a history, synthesis, and outlook. Diversity and Distributions 28, 386-403.
| Crossref | Google Scholar |
Kelly LT, Giljohann KM, Duane A, Aquilué N, Archibald S, Batllori E, Bennett AF, Buckland ST, Canelles Q, Clarke MF, Fortin M-J, Hermoso V, Herrando S, Keane RE, Lake FK, McCarthy MA, Morán-Ordóñez A, Parr CL, Pausas JG, Penman TD, Regos A, Rumpff L, Santos JL, Smith AL, Syphard AD, Tingley MW, Brotons L (2020) Fire and biodiversity in the Anthropocene. Science 370, eabb0355.
| Crossref | Google Scholar |
Lawes MJ, Murphy BP, Fisher A, Woinarski JCZ, Edwards AC, Russell-Smith J (2015) Small mammals decline with increasing fire extent in northern Australia: evidence from long-term monitoring in Kakadu National Park. International Journal of Wildland Fire 24, 712-722.
| Crossref | Google Scholar |
Leahy L, Legge SM, Tuft K, McGregor HW, Barmuta LA, Jones ME, Johnson CN (2016) Amplified predation after fire suppresses rodent populations in Australia’s tropical savannas. Wildlife Research 42, 705-716.
| Crossref | Google Scholar |
Legge S, Smith JG, James A, Tuft KD, Webb T, Woinarski JCZ (2019) Interactions among threats affect conservation management outcomes: livestock grazing removes the benefits of fire management for small mammals in Australian tropical savannas. Conservation Science and Practice 1, e52.
| Crossref | Google Scholar |
Lindsay M, Beames L, Yawuru Country Managers, Nyul Nyul Rangers, Bardi Jawi Rangers (2022) Integrating scientific and Aboriginal knowledge, practice and priorities to conserve an endangered rainforest ecosystem in the Kimberley region, northern Australia. Ecological Management & Restoration 23, 93-104.
| Crossref | Google Scholar |
Lynch D, Russell-Smith J, Edwards AC, Evans J, Yates C (2018) Incentivising fire management in Pindan (Acacia shrubland): a proposed fuel type for Australia’s Savanna burning greenhouse gas emissions abatement methodology. Ecological Management & Restoration 19, 230-238.
| Crossref | Google Scholar |
McGregor HW, Legge S, Jones ME, Johnson CN (2014) Landscape management of fire and grazing regimes alters the fine-scale habitat utilisation by feral cats. PLoS ONE 9, e109097.
| Crossref | Google Scholar | PubMed |
McGregor H, Legge S, Jones ME, Johnson CN (2015) Feral cats are better killers in open habitats, revealed by animal-borne video. PLoS ONE 10, e0133915.
| Crossref | Google Scholar | PubMed |
McKenzie NL (1981) Mammals of the Phanerozoic South-West Kimberley, Western Australia: biogeography and recent changes. Journal of Biogeography 8, 263-280.
| Crossref | Google Scholar |
Miller G, Friedel M, Adam P, Chewings V (2010) Ecological impacts of buffel grass (Cenchrus ciliaris L.) invasion in central Australia – does field evidence support a fire-invasion feedback? The Rangeland Journal 32, 353-365.
| Crossref | Google Scholar |
Moseby KE, McGregor HM (2022) Feral cats use fine scale prey cues and microhabitat patches of dense vegetation when hunting prey in arid Australia. Global Ecology and Conservation 35, e02093.
| Crossref | Google Scholar |
Moseby KE, Read JL, Paton DC, Copley P, Hill BM, Crisp HA (2011) Predation determines the outcome of 10 reintroduction attempts in arid South Australia. Biological Conservation 144, 2863-2872.
| Crossref | Google Scholar |
Murphy BP, Davies HF (2014) There is a critical weight range for Australia’s declining tropical mammals. Global Ecology and Biogeography 23, 1058-1061.
| Crossref | Google Scholar |
NAFI (2022) North Australia and rangelands fire information. Available at https://firenorth.org.au/nafi3/
Nakagawa S, Cuthill IC (2007) Effect size, confidence interval and statistical significance: a practical guide for biologists. Biological Reviews 82, 591-605.
| Crossref | Google Scholar | PubMed |
National Native Title Tribunal (2023) Western Australia native title claimant applications and determination areas. Available at chrome-extension://efaidnbmnnnibpcajpcglclefindmkaj/http://www.nntt.gov.au/Maps/WA_NTDA_Schedule.pdf
NDVI (2020) NVIS 6.0 major vegetation subgroups. Available at https://www.dcceew.gov.au/environment/land/native-vegetation/national-vegetation-information-system/data-products#mvsg60
Nilsson L (2017) The human cost of pearl. Voiceworks 107, 89-94.
| Google Scholar |
Nimmo DG, Avitabile S, Banks SC, Bliege Bird R, Callister K, Clarke MF, Dickman CR, Doherty TS, Driscoll DA, Greenville AC, Haslem A, Kelly LT, Kenny SA, Lahoz-Monfort JJ, Lee C, Leonard S, Moore H, Newsome TM, Parr CL, Ritchie EG, Schneider K, Turner JM, Watson S, Westbrooke M, Wouters M, White M, Bennett AF (2019) Animal movements in fire-prone landscapes. Biological Reviews 94, 981-998.
| Crossref | Google Scholar | PubMed |
Ondei S, Prior LD, McGregor HW, Reid AM, Johnson CN, Vigilante T, Goonack C, Williams D, Bowman DMJS (2020) Small mammal diversity is higher in infrequently compared with frequently burnt rainforest–savanna mosaics in the north Kimberley, Australia. Wildlife Research 48, 218-229.
| Crossref | Google Scholar |
Radford IJ, Fairman R (2015) Fauna and vegetation responses to fire and invasion by toxic cane toads (Rhinella marina) in an obligate seeder-dominated tropical savanna in the Kimberley, northern Australia. Wildlife Research 42, 302-314.
| Crossref | Google Scholar |
Radford IJ, Gibson LA, Corey B, Carnes K, Fairman R (2015) Influence of fire mosaics, habitat characteristics and cattle disturbance on mammals in fire-prone savanna landscapes of the northern Kimberley. PLoS ONE 10, e0130721.
| Crossref | Google Scholar | PubMed |
Radford IJ, Woolley L-A, Corey B, Vigilante T, Wunambal Gaambera Aboriginal Corporation, Hatherley E, Fairman R, Carnes K, Start AN (2020) Prescribed burning benefits threatened mammals in northern Australia. Biodiversity and Conservation 29, 2985-3007.
| Crossref | Google Scholar |
Radford IJ, Corey B, Carnes K, Shedley E, McCaw L, Woolley L-A (2021) Landscape-scale effects of fire, cats, and feral livestock on threatened savanna mammals: unburnt habitat matters more than pyrodiversity. Frontiers in Ecology and Evolution 9, 739817.
| Crossref | Google Scholar |
Rainsford FW, Kelly LT, Leonard SWJ, Bennett AF (2020) Post-fire development of faunal habitat depends on plant regeneration traits. Austral Ecology 45, 800-812.
| Crossref | Google Scholar |
R Core Team (2023) R: A Language and Environment for Statistical Computing. R version 4.3.2 – ‘Eye Holes’. R Foundation for Statistical Computing, Vienna, Austria. Available at https://www.R-project.org/. SVN revision 85441. Release date: 2023-10-31
Rossiter NA, Setterfield SA, Douglas MM, Hutley LB (2003) Testing the grass-fire cycle: alien grass invasion in the tropical savannas of northern Australia. Diversity and Distributions 9, 169-176.
| Crossref | Google Scholar |
Russell-Smith J, Edwards AC (2006) Seasonality and fire severity in savanna landscapes of monsoonal northern Australia. International Journal of Wildland Fire 15, 541-550.
| Crossref | Google Scholar |
Russell-Smith J, Yates C, Edwards A, Allan GE, Cook GD, Cooke P, Craig R, Heath B, Smith R (2003a) Contemporary fire regimes of northern Australia, 1997–2001: change since Aboriginal occupancy, challenges for sustainable management. International Journal of Wildland Fire 12, 283-297.
| Crossref | Google Scholar |
Russell-Smith J, Whitehead PJ, Cook GD, Hoare JL (2003b) Response of Eucalyptus-dominated savanna to frequent fires: lessons from Munmarlary, 1973–1996. Ecological Monographs 73, 349-375.
| Crossref | Google Scholar |
Santos JL, Hradsky BA, Keith DA, Rowe KC, Senior KL, Sitters H, Kelly LT (2022) Beyond inappropriate fire regimes: a synthesis of fire-driven declines of threatened mammals in Australia. Conservation Letters 15, e12905.
| Crossref | Google Scholar |
Senior KL, Giljohann KM, McCarthy MA, Rainsford FW, Kelly LT (2021) Predicting mammal responses to pyrodiversity: from microbats to macropods. Biological Conservation 256, 109031.
| Crossref | Google Scholar |
Shaw RE, James AI, Tuft K, Legge S, Cary GJ, Peakall R, Banks SC (2021) Unburnt habitat patches are critical for survival and in situ population recovery in a small mammal after fire. Journal of Applied Ecology 58, 1325-1335.
| Crossref | Google Scholar |
Skroblin A, Carboon T, Bidu G, Taylor M, Bidu N, Taylor W, Taylor K, Miller M, Robinson L, Williams C, Chapman N, Marney M, Marney C, Biljabu J, Biljabu L, Jeffries P, Samson H, Charles P, Game ET, Wintle B (2022) Developing a two-way learning monitoring program for Mankarr (greater bilby) in the Western Desert, Western Australia. Ecological Management & Restoration 23, 129-138.
| Crossref | Google Scholar |
Southgate R, Carthew SM (2006) Diet of the bilby (Macrotis lagotis) in relation to substrate, fire and rainfall characteristics in the Tanami Desert. Wildlife Research 33, 507-519.
| Crossref | Google Scholar |
Southgate R, Carthew S (2007) Post-fire ephemerals and spinifex-fuelled fires: a decision model for bilby habitat management in the Tanami Desert, Australia. International Journal of Wildland Fire 16, 741-754.
| Crossref | Google Scholar |
Southgate R, Paltridge R, Masters P, Carthew S (2007) Bilby distribution and fire: a test of alternative models of habitat suitability in the Tanami Desert, Australia. Ecography 30, 759-776.
| Crossref | Google Scholar |
Southgate R, Dziminski MA, Paltridge R, Schubert A, Gaikhorst G (2019) Verifying bilby presence and the systematic sampling of wild populations using sign-based protocols – with notes on aerial and ground survey techniques and asserting absence. Australian Mammalogy 41, 27.
| Crossref | Google Scholar |
Stobo-Wilson AM, Stokeld D, Einoder LD, Davies HF, Fisher A, Hill BM, Mahney T, Murphy BP, Scroggie MP, Stevens A, Woinarski JCZ, Bawinanga Rangers, Warddeken Rangers, Gillespie GR (2020) Bottom-up and top-down processes influence contemporary patterns of mammal species richness in Australia’s monsoonal tropics. Biological Conservation 247, 108638.
| Crossref | Google Scholar |
Tingley MW, Ruiz-Gutiérrez V, Wilkerson RL, Howell CA, Siegel RB (2016) Pyrodiversity promotes avian diversity over the decade following forest fire. Proceedings of the Royal Society B: Biological Sciences 283, 20161703.
| Crossref | Google Scholar |
Trewella GJ, Cremona T, Nevard H, Murphy BP (2023) Habitat structure facilitates coexistence of native and invasive mesopredators in an Australian tropical savanna. Wildlife Research 50, 1058-1070.
| Crossref | Google Scholar |
Vigilante T (2001) Analysis of explorers’ records of aboriginal landscape burning in the Kimberley Region of Western Australia. Australian Geographical Studies 39, 135-155.
| Crossref | Google Scholar |
Vigilante T, Bowman DMJS, Fisher R, Russell-Smith J, Yates C (2004) Contemporary landscape burning patterns in the far North Kimberley region of north-west Australia: human influences and environmental determinants. Journal of Biogeography 31, 1317-1333.
| Crossref | Google Scholar |
von Takach B, Scheele BC, Moore H, Murphy BP, Banks SC (2020) Patterns of niche contraction identify vital refuge areas for declining mammals. Diversity and Distributions 26, 1467-1482.
| Crossref | Google Scholar |
von Takach B, Jolly CJ, Dixon KM, Penton CE, Doherty TS, Banks SC (2022) Long-unburnt habitat is critical for the conservation of threatened vertebrates across Australia. Landscape Ecology 37, 1469-1482.
| Crossref | Google Scholar |
Wan HY, Cushman SA, Ganey JL (2020) The effect of scale in quantifying fire impacts on species habitats. Fire Ecology 16, 9.
| Crossref | Google Scholar |
Woinarski JCZ, Armstrong M, Brennan K, Fisher A, Griffiths AD, Hill B, Milne DJ, Palmer C, Ward S, Watson M, Winderlich S, Young S (2010) Monitoring indicates rapid and severe decline of native small mammals in Kakadu National Park, northern Australia. Wildlife Research 37, 116.
| Crossref | Google Scholar |
Woinarski JCZ, Legge S, Fitzsimons JA, Traill BJ, Burbidge AA, Fisher A, Firth RSC, Gordon IJ, Griffiths AD, Johnson CN, McKenzie NL, Palmer C, Radford I, Rankmore B, Ritchie EG, Ward S, Ziembicki M (2011) The disappearing mammal fauna of northern Australia: context, cause, and response. Conservation Letters 4, 192-201.
| Crossref | Google Scholar |
Woinarski JCZ, Burbidge AA, Harrison PL (2015) Ongoing unraveling of a continental fauna: decline and extinction of Australian mammals since European settlement. Proceedings of the National Academy of Sciences 112, 4531-4540.
| Crossref | Google Scholar |
Wysong M, Legge S, Clark A, Maier S, Bardi Jawi Rangers, Nyul Nyul Rangers, Yawuru Country Managers, Cowell S, Mackay G (2021) The sum of small parts: changing landscape fire regimes across multiple small landholdings in north-western Australia with collaborative fire management. International Journal of Wildland Fire 31, 97-111.
| Crossref | Google Scholar |
Ziembicki MR, Woinarski JCZ, Mackey B (2013) Evaluating the status of species using Indigenous knowledge: novel evidence for major native mammal declines in northern Australia. Biological Conservation 157, 78-92.
| Crossref | Google Scholar |