

Activity range and patterns of free-roaming village dogs in a rural Cambodian village
Rachel Ladd A * , Paul Meek A B , Jonathan C. Eames C D and Luke K.-P. Leung A
A * , Paul Meek A B , Jonathan C. Eames C D and Luke K.-P. Leung A
A School of Agriculture and Food Sciences, The University of Queensland, Gatton, Qld 4343, Australia.
B Vertebrate Pest Research Unit, NSW Department of Primary Industries, PO Box 530, Coffs Harbour, NSW 2450, Australia.
C Rising Phoenix Co, Ltd., House 32A, Street 494, Phnom Penh, Cambodia.
D BirdLife International Cambodia Programme, House 32A, Street 494, Phnom Penh, Cambodia.
Abstract
Free-roaming dogs are ubiquitous worldwide and pose a threat to wildlife. An understanding of the roaming behaviour of dogs is useful for developing effective management strategies.
We aimed to assess the activity ranges and patterns of free-roaming dogs in a rural Cambodian village. An adjacent wildlife sanctuary protects populations of threatened species that may be negatively impacted by dogs.
We used rudimentary hand-made GPS collars to track village dogs and quantify their movements. Activity ranges were calculated for male and female dogs, and the mean distances travelled at night and during the day were determined. Additionally, forays outside of the activity range were characterised.
We estimated a mean activity range of 178 ± 190 ha and found that dogs typically travelled longer distances at night. Females had significantly smaller activity ranges, as well as covering shorter distances per day. Foray behaviour was variable, with some dogs entering the wildlife sanctuary regularly and others not at all.
Free-roaming Cambodian dog activity overlapped with native mammals in the study site, posing a risk of predation, resource exclusion, harassment and competition of Cambodian species.
The data suggest that although dogs generally spend the majority of their time within the village, there is significant risk to wildlife. Actions are required to mitigate threats to native wildlife, in particular threatened species such as Eld’s Deer.
Keywords: activity range, Cambodia, forays, free-roaming dogs, GPS tracking, roaming behaviour, village dogs, wildlife.
Introduction
Domestic dogs (Canis familiaris) are globally abundant, with estimates ranging from 700 million to one billion (Hughes and Macdonald 2013; Gompper 2014). Village dogs, those that associate with humans in and around rural dwellings and are typically free-roaming, are the most abundant category of dogs living around the world (Ortolani et al. 2009). Dogs are a recognised threat to biodiversity conservation globally (Seebeck 1979; Kruuk and Snell 1981; Taborsky 1988). However, owned free-roaming dogs have received less attention at an ecological scale than other introduced predators (although see Meek 1999), such as the domestic cat (Felis catus) (Hughes and Macdonald 2013), and have more often been considered in relation to issues of human health (Jenkins and Andrews 1993; Butler and Bingham 2000; Durr and Ward 2014; Morters et al. 2014; Chevalier et al. 2021; Warembourg et al. 2021).
Direct predation is the obvious detrimental effect of dogs on wildlife (Kruuk and Snell 1981; Wierzbowska et al. 2016; Gatti et al. 2018), but they can also have other negative impacts, including fear-mediated behavioural changes (Banks and Bryant 2007; Zapata-Rios and Branch 2016), competition (Vanak et al. 2015), harassment (Weston and Stankowich 2015), hybridisation (Bergman et al. 2009) and disease transmission (Jenkins and Andrew 1993; Alexander et al. 1996; Bergman et al. 2009). The reproductive behaviour of ungulates has been shown to be affected by the presence of domestic dogs (Manor and Saltz 2004; Gingold et al. 2009), and more than 60 zoonoses have been associated with dogs (Matter and Daniels 2000). Dogs have been recorded as having a negative impact on many threatened species, with Southeast Asia having the highest number of impacted species, but this is likely a major underestimate (Doherty et al. 2017). The management of domestic dogs is therefore of serious conservation concern.
Due to the complex roles dogs have in human communities, management strategies need to be informed by ecological, social, cultural and economic perspectives to be effective (White and Ward 2010). Understanding how and where dogs affect wildlife is essential for prioritisation of management actions. Free-roaming dogs in rural landscapes may move between the human interface, where they have access to shelter and food, and the surrounding landscape where they may encounter wildlife (Macdonald and Carr 2016). This is of particular concern where human habitation is adjacent to protected areas (Meek 1999). Roaming behaviour varies among dogs, with some more likely to engage in forays that lead to interactions with wildlife (Meek 1999), and investigating the characteristics of these dogs is important for management action. The roaming behaviour, social structure, diet, demographics and reproductive physiology are important data when managing free-roaming dogs, especially where jurisdictional governance is lacking.
Siem Pang Wildlife Sanctuary in northern Cambodia is home to an ecologically important population of Eld’s deer (Rucervus eldii) (Ladd et al. 2022) and other threatened species (Loveridge et al. 2018), and dogs have been implicated as having a negative impact on Eld’s deer (Gray et al. 2015). There are several villages surrounding the sanctuary (and one located inside the boundary) that are home to free-roaming dogs. In India, 80 species have been identified as being negatively impacted by dogs (Home et al. 2018). Several of these identified species, or similar species, are present in Siem Pang Wildlife Sanctuary, including three critically endangered vulture species (Gyps tenuirostris, G. bengalensis and Sarcogyps calcus), sambar (Rusa unicolor) and red muntjac (Muntiacus vaginalis). Dogs may have also played a role in the decline of Dhole (Cuon alpinus) from the wildlife sanctuary, with dogs being a significant competitor and disease vector of this native canine species (Loveridge et al. 2018).
The objective of the present study was to gain insights into the roaming behaviour of village dogs in relation to the wildlife sanctuary. We specifically aimed to: (1) determine the activity ranges of dogs and the degree of overlap into the wildlife sanctuary; (2) determine the activity patterns of dog movement by examining the accumulated distance travelled; (3) investigate any sex differences in the size and pattern of activity; and (4) characterise forays undertaken by dogs outside of their typical area of activity.
Materials and methods
Study site
The study was conducted in Khes Svay village, Stung Treng Province, Cambodia, due to its proximity to the Siem Pang Wildlife Sanctuary (Fig. 1). The mid-point of the village is approximately 2.2 km from the sanctuary boundary, with some dwellings less than 500 m away. The village consists of 233 households (Siem Pang Administration Office, unpubl. data) with small-scale agriculture being the main source of economic activity in the village. The dog ownership was previously found to be 2.41 dogs per household, and dogs sometimes accompany their owners into the wildlife sanctuary (Ladd 2022). No previous dog management activities have been carried out in this area prior to the present study, making it ideal for observing village dogs behaving in their natural state.
Collar construction
GPS tracking collars were custom made using the iGot-U GT600 GPS logger (Mobile Action Technology Inc., Taiwan) and commercial leather dog collars. A plastic case was modified slightly to fit the logger and curved by a heat gun to attach to the collar and provide increased ergonomy, protection and durability. A number of holes were punched into the leather collars to accommodate different sized dogs. The case containing the logger was fastened to the collar using stainless steel nylon lock nut and bolts. A fishing sinker was attached at the collar buckle as a counterweight so that the logger pointed skywards on the back of the dog’s neck for improved satellite reception. At the time of deployment, the logger was activated, placed in the case and sealed using silicon tape for waterproofing, electrical tape and two zip ties to prevent the case from opening. The collar was then fastened around the dog’s neck, allowing an approximate 2-finger spacing for comfortable wear, and the excess collar tongue zip tied.
Collar deployment
Collars were deployed between January and March 2020. Due to low participation rates, dogs were only rejected if they were too small for the GPS collar or visibly in ill-health. This resulted in the inclusion of one female dog that was ‘possibly pregnant’ at the time of first deployment, and one male dog with minor mobility issues due to a historical leg injury, but was otherwise in good health. All dogs were of no defined breed, or a mix of breeds. Multiple dogs from the same household were tracked if they were of suitable size and health, and they were treated independently.
GPS collars were fitted to dogs with the assistance of the owners, who either held the dog in place for the fitting, or were instructed on how to fit the collar under supervision so that it was not too tight or loose, and so the GPS logger was positioned correctly. The GPS loggers were set to record a position every 15 min. However, in practice this interval could vary slightly or significantly due to temporal noise of the GPS unit or issues with the ability of the GPS logger to connect with satellites. The dogs were all photographed and had their sex (and for females, reproductive status) recorded, along with the owner’s age estimation and any distinguishing features. The location of the household was recorded using a handheld GPS unit. Due to the battery life of the GPS loggers, a collar was exchanged for a new, fully charged collar after 2 weeks of deployment, such that each dog was tracked for an approximate 4-week period. However, due to logistical and technical reasons, the total time each dog was tracked varied slightly, and one owner of three dogs changed their mind soon after deployment and opted to discontinue participation.
Data analysis
GPS data from the loggers were downloaded upon retrieval using the @trip PC software (http://www.a-trip.com). Data were exported as a .csv file and cleaned, with any GPS fixes recorded prior to the collar being secured to the dog or after collar removal being excluded from the dataset. Additionally, three GPS fixes from three different dogs were removed due to a recording error, with the fixes being extreme distances away from the other locations, such that the dog would not be able to travel the distance in the time interval recorded, even if the dog was in a car.
The generally accepted definition of an animal’s home range is the ‘area traversed by the individual in its normal activities of food gathering, mating and caring for young’ (Burt 1943). However, this definition has limitations, including a lack of temporal specification (Cooper 1978). The approximate 4-week period of data in this study was too short and exclusive of some normal activities, which would preclude the calculation of a home range, although home ranges have been calculated from data collected over only 1–3 days (e.g. Durr and Ward 2014). These should more accurately be termed an activity range, that is applicable for the particular conditions under which data were collected (e.g. seasonality). We therefore calculated a dry season activity range for the dogs in this study by the crude calculation of a minimum convex polygon (MCP), using a 95% isopleth to exclude the most extreme fixes. The sizes of male and female activity ranges were compared using a Mann–Whitney U-test. The percentage overlap of individual dog’s activity range with the wildlife sanctuary area was calculated.
The number of forays undertaken by each dog, as well as the average distance, maximum distance, duration and timing of forays, was collated, as well as whether the dog entered the wildlife sanctuary. A foray was generally defined as three or more consecutive fixes outside of the activity range that were also more than 500 m from the household, and that lasted at least 30 min (as measured from time of first fix to last fix outside of the activity range). However, in some cases, a series of fixes were excluded from consideration as forays if they clustered close to the edge of the activity range or 500-m buffer and if there was regular movement back and forth into the activity range or the 500-m buffer area. The average accumulated distance travelled by each dog (adding the distance between each consecutive GPS fix) was calculated per 24-h period, and sex differences in this was tested by a Mann–Whitney U-test. Additionally, the average accumulated distance travelled during daylight (0600 to 1800 hours) and at night (1800 to 0600 hours) was calculated and compared using a Mann–Whitney U-test.
The activity range was calculated for each dog tracked; however, dogs B, D and E were excluded from comparisons due to being tracked for only two full days, and their accumulated distance travelled per day, as well as forays, was not determined. Dog F and dog L were excluded from comparisons because the former gave birth during the fourth week of being tracked, resulting in major changes to her activity levels during this time, and the latter due to mobility issues.
Camera trapping
Camera trapping surveys were conducted in Siem Pang Wildlife Sanctuary during the dry seasons of 2018–2019 and 2020 for population assessment of Eld’s deer (Ladd 2022). Camera traps are not target specific, and non-target wildlife, domestic animals and humans are often detected. To supplement the data provided by the GPS collars, we examined these camera trap datasets for all dog detections.
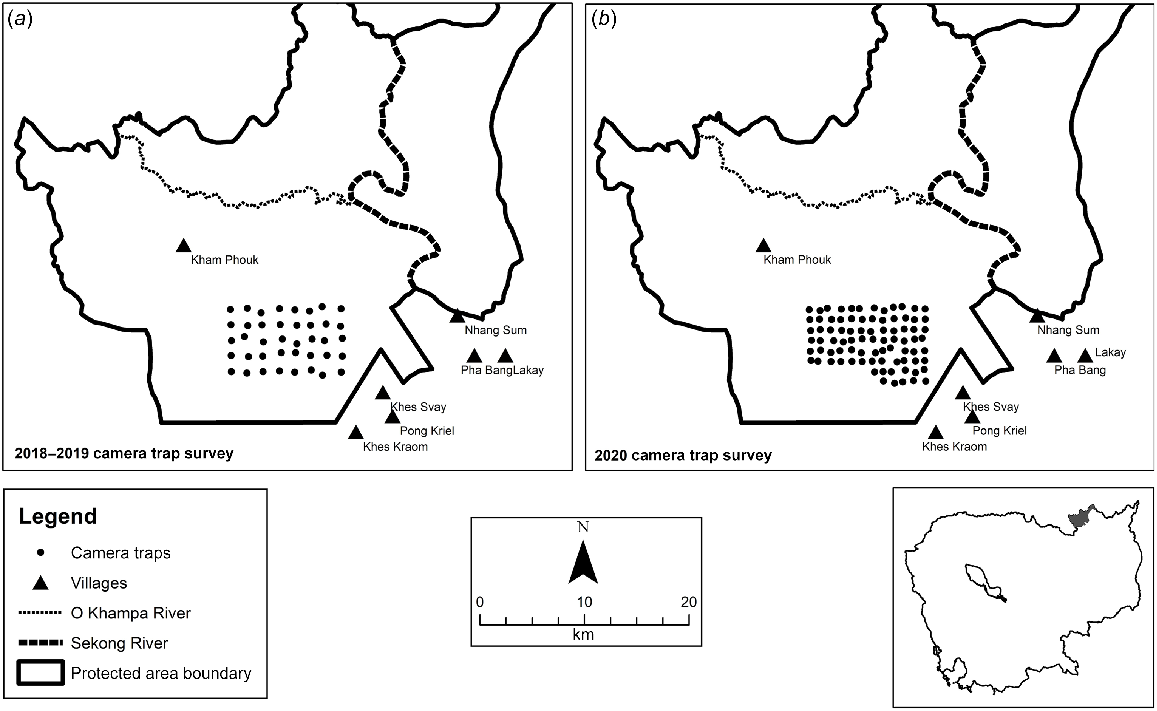
The surveys used two Reconyx camera trap models (HC600 and HP2W), with one camera mounted on a tree at approximately 0.8 m high at each site and vegetation within 2 m of the camera being slashed to avoid wildfire damage and provide an open field of view. The 2018–19 survey consisted of a grid with 40 sites 1.5 km apart, with cameras set to take 10 photos per trigger (Fig. 1). This survey ran from early December to late May. The 2020 survey consisted of a grid of 83 sites 1 km apart, with cameras set to take five photos per trigger (Fig. 1). The camera trap grid was divided into two adjacent blocks, surveyed consecutively from January to mid-March and mid-March to May. In both surveys, the cameras were set with no delay and high sensitivity, and cameras were serviced regularly.
Dog detections were considered independent events if they were separated by 20 min or more, with the location, time of day, group size and whether the dogs were detected in the company of humans also recorded.
Results
GPS tracking
There were 13 dogs tracked from nine households, with seven being female and six male (Table 1). The average age of the dogs tracked was 4 years (min = 1 year, max = 10 years). Of the female dogs, one had had puppies recently but they had been given away prior to collaring, and one dog was said to be possibly pregnant at the time of initial collaring. This was later confirmed, with the dog giving birth during the fourth week of tracking. The number of data points collected per dog varied because the interval between recorded locations often diverged from the 15-min interval set, and could result in a more or less frequent recording of location. The total number of GPS fixes recorded was 57 625, with an average of 5579 fixes per dog, excluding the dogs tracked for only 2 days, which averaged 613 fixes per dog.
| Dog ID | Household | Distance to sanctuary boundary (m) | Sex | Age | Total fixes | Total days (24 h) | |
|---|---|---|---|---|---|---|---|
| A | H1 | 3447 | F | 6 | 5607 | 27 | |
| BA | H2 | 641 | F | 2 | 644 | 2 | |
| C | H3 | 2072 | F | 3 | 6064 | 27 | |
| DA | H2 | F | 5 | 588 | 2 | ||
| EA | H2 | M | 2 | 608 | 2 | ||
| FB | H4 | 1210 | F | 2 | 5397 | 27 | |
| G | H5 | 2078 | M | 6–7 | 5785 | 27 | |
| H | H5 | F | 1 | 6280 | 27 | ||
| I | H6 | 1365 | M | 1 | 4142 | 18 | |
| J | H6 | M | 7 | 4965 | 23 | ||
| K | H7 | 1334 | F | 5 | 5991 | 26 | |
| LC | H8 | 1184 | M | 10 | 5324 | 28 | |
| M | H9 | 1150 | M | 2 | 6234 | 29 |
Distance to sanctuary border is the minimum distance between the household and sanctuary boundary.
AOwners of dogs B, D and E decided against continued participation after 4 days (two 24-h days completed only).
BDog F gave birth to puppies during the fourth week of tracking.
CDog L had mobility issues due to an historic rear leg injury.
The MCP 95% isopleth activity range estimates varied greatly among dogs, with the smallest activity range estimated for the male dog with mobility issues at 0.78 ha, and the largest at 509.52 ha (Table 2). Excluding the dogs tracked for 2 days, as well as dog L and dog F, the mean activity range was 178 ± 190 ha (median = 91.4 ha). The mean distance travelled in 24 h was 8313 ± 4353 m, in daylight hours it was 3431 ± 2444 m and for the night it was 4428 ± 3552 m (Table 2).
| Dog ID | Sex | Activity range size (ha) | Percentage overlap (%) | Mean distance (m/24 h) | Mean distance (m/day) | Mean distance (m/night) | |
|---|---|---|---|---|---|---|---|
| A | F | 2.32 | 0 | 5486 | 3333 | 2176 | |
| B | F | 43.38 | 0 | – | – | – | |
| C | F | 5.69 | 0 | 5648 | 2039 | 3660 | |
| D | F | 21.10 | 0 | – | – | – | |
| E | M | 33.39 | 0 | – | – | – | |
| F | F | 1.60 | 0 | 5157 | 2498 | 2619 | |
| G | M | 97.43 | 0 | 7714 | 2383 | 5492 | |
| H | F | 6.02 | 0 | 5808 | 2665 | 3485 | |
| I | M | 291.69 | 0 | 12 976 | 4304 | 8886 | |
| J | M | 509.52 | 10 | 11 836 | 3623 | 8534 | |
| K | F | 85.36 | 0 | 8045 | 3953 | 4293 | |
| L | M | 0.78 | 0 | 3967 | 2291 | 1672 | |
| M | M | 426.30 | 25 | 10 867 | 5663 | 5341 |
The comparison of activity ranges found a significant difference between males and females (P-value = 0.03038), with the mean female activity ranges being smaller than males (female = 24.8 ± 40.4 ha; male = 331 ± 180 ha). The accumulated distance travelled per 24 h by female and male dogs also differed significantly (P-value = 2.2e-16), with males typically covering more distance (mean = 10 611 ± 4937 m) than females (mean = 6230 ± 2235 m). The accumulated distance travelled at night compared with daylight hours was found to differ significantly (P-value = 0.015), with the mean distance covered at night being greater (day = 3431 ± 2444 m; night = 4428 ± 3552 m).
The number of forays per dog varied from none to 15, with the highest number of forays being recorded for a female dog and the longest distance travelled recorded by a male dog (Table 3). Forays most often commenced between midnight and 0600 hours, followed by 0600–1200 hours, then after 1800 hours. Dog M exhibited particularly consistent foray behaviour, with eight out of nine forays beginning between 0600 and 0700 hours, and typically lasting between 2 and 4 h, although one foray lasted just over 9 h. The other dogs with high numbers of forays, dogs C, I, J and K, were less consistent with when they went on forays, although the majority of forays for these dogs commenced between midnight and 0600 hours. Forays did not usually surpass 4 h in duration, and were often less than 2 h, with only dog M (9 h 9 min) and dog C (8 h 15 min) having a single foray lasting most of the day. Five dogs entered the wildlife sanctuary at least once while on a foray, with four of these dogs entering the wildlife sanctuary on 80–100% of their forays.
| Dog ID | Sex | No. forays | Timing (hours) | Mean duration (h) | Mean total distance (m) | Maximum distance (m) | Forays into sanctuary (%) | |
|---|---|---|---|---|---|---|---|---|
| A | F | 1 | 0600–1200 | 0.49 | 1997 | 888 | 0 | |
| C | F | 13 | 0000−1800 | 1.55 | 1127 | 1496 | 0 | |
| F | F | 2 | 0000−0600 | 2.08 | 5780 | 3039 | 100 | |
| G | M | 2 | 0000−1200 | 3.84 | 2271 | 2074 | 0 | |
| H | F | 2 | 0000−0600 | 0.63 | 590 | 748 | 0 | |
| I | M | 10 | 0000−0600 | 1.83 | 5108 | 5013 | 80 | |
| J | M | 11 | 0000−0600 | 2.06 | 5416 | 5155 | 91 | |
| K | F | 15 | 0000−0600 | 1.32 | 2354 | 2698 | 7 | |
| L | M | 0 | – | – | – | – | – | |
| M | M | 9 | 0600−1200 | 3.34 | 6968 | 6118 | 100 |
The timing refers to the time of day the majority of forays occurred. The mean total distance is calculated accumulatively from the first fix to the last fix outside of the activity range or 500-m buffer within each foray. The maximum total distance is measured from the household location to the furthest fix recorded during forays. The number of forays into the sanctuary is given as a percentage.
Camera trapping
There were 4026 camera trapping days in the first survey during the 2018–2019 dry season, with 34 dog events (Fig. 2). Dogs were detected at 15 of the 40 camera trap sites, and were distributed across the trapping grid, including the site most distant from Khes Svay village. Dogs were detected in the company of humans in only 13 events, and only three events occurred at night.
Maps showing the locations that dogs were detected by camera traps in (a) 2018–2019 and (b) 2020. Black circles indicate dog detections and grey circles where no dog detections occurred.
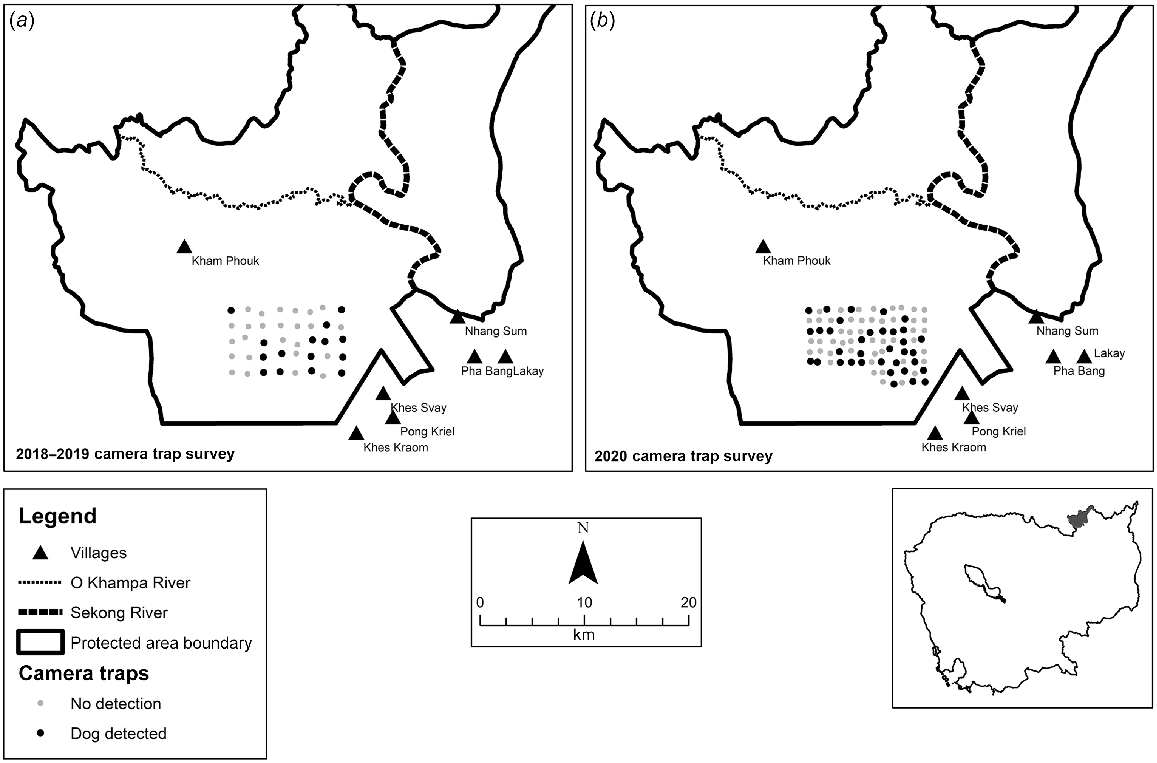
The second camera trap survey totalled 5979 camera trap days and there were 51 dog events, with only 16 events showing dogs in the company of humans (Fig. 2). Dogs were detected at 35 of the 83 camera trap sites and were similarly distributed across the trapping grid, including the site most distant from Khes Svay village, with only 10 events occurring at night.
Discussion
Cambodian village dogs living adjacent to the Siem Pang Wildlife Sanctuary were shown to use the natural habitat at various times, with GPS collars indicating more frequent night and early morning visits, but camera trap detections occurred most often during the day. Male dogs were found to have a larger activity range and travelled longer distances than female dogs on average, with only two male dogs having activity ranges that overlapped with the wildlife sanctuary boundary. The size of activity ranges varied widely, with some dogs being much more sedentary than others. Dogs were more active at night, with a larger accumulated distance recorded between 1800 and 0600 hours for most dogs. The small sample sizes resulted in non-normally distributed data, which was particularly apparent for female dogs. This prevented the use of a two-way ANOVA to determine any interaction between sex and the accumulated distance travelled in night or day time hours, but visualisation of the data suggests there may have been an interaction. Further study with a large sample size is required to critically determine if the interaction is significant. We also chose not to conduct comparisons of foray behaviour due to the small sample size and high degree of variation.
The smaller activity ranges and shorter accumulated distances travelled per day indicate that females are staying closer to their household than males, although female dogs are engaging in similar numbers of forays to male dogs. Dogs living in households more than 2 km from the wildlife sanctuary boundary never entered the sanctuary during the tracking period. The dogs living in the household closest to the sanctuary boundary were only tracked for two full days, and foray behaviour was not examined due the short timeframe. These three dogs had activity ranges that did not overlap with the sanctuary, and although their activity range size was not noticeably different to that of the other dogs, their activity range would potentially be different if they had been tracked for all 4 weeks.
Direct comparisons of the activity range of these Cambodian free-roaming village dogs with the activity or home ranges of other free-roaming dogs around the world is made challenging by the use of different data collection methods and alternate methods for calculating home range, as well as the choice of different isopleth sizes to define home/activity range or core activity. Nevertheless, the estimated median home/activity range of free-roaming dogs from selected studies from various locations are given in Table 4 for comparison. In most cases, these studies assert to be estimating the home range of the dogs, with only Sparkes et al. (2014), Ruiz-Izaguirre et al. (2015) and Kennedy et al. (2018) indicating the estimate is for an activity range. However, this is misleading because, with the exception of Meek (1999), the duration dogs were tracked is very short, often just a couple of days, which is not sufficient to encompass the range of normal activities needed to justify a home range designation. This issue was alluded to in some studies. The majority of the activity ranges were small, less than 10 ha, with only the studies tracking dogs for 7 days or more exceeding this. However, all median activity/home ranges were smaller than the estimated median activity range of 91.4 ha for the free-roaming dogs in this study. However, this is not inconsistent with the individual activity ranges calculated for some individual dogs in the other studies. The duration of tracking likely plays a significant role in the observed difference, although the median activity range of the Cambodian dogs was still much larger than for the free-roaming dogs tracked in Mexico and Chile for somewhat similar time frames. Meek (1999) used VHF radio tracking on foot (GPS tracking was not possible at the time), so fewer fixes were recorded within a session, although the dogs were tracked over a 15-month period. Radio tracking was also used by Ruiz-Izaguirre et al. (2015), but for 45 days. Space use by dogs will likely vary significantly due to temporal factors, and dogs may not move consistently within their home range (Ballard et al. 2018), so it is important to consider what inferences can be made due to the temporal scale of these studies.
| Study | Location | Method | Median home/activity range (ha) | Tracking duration | Tracking method | |
|---|---|---|---|---|---|---|
| Meek (1999) | Australia | MCP isopleth, outliers excluded | 72.50 | 15 months | VHF radio tracking | |
| Van Kesteren et al. (2013) | Kyrgyzstan | Characteristic hull polygon | 2.26 | Mean 20 h | GPS tracking | |
| Sparkes et al. (2014) | Australia | MCP 100% isopleth, forays excluded | 37.47 | 7 days | GPS tracking | |
| Durr and Ward (2014) | Australia | MCP 95% isopleth | 3.60 | Mean 50 h | GPS tracking | |
| Ruiz-Izaguirre et al. (2015) | Mexico | Kernel density | 16.10 | 45 days | VHF radio tracking | |
| Kennedy et al. (2018) | Australia | MCP 100% isopleth, forays excluded | 8.88 | 3–5 days | GPS tracking | |
| Muinde et al. (2021) | Kenya | MCP 95% isopleth | 9.30 | 5 days | GPS tracking | |
| Saavedra-Aracena et al. (2021) | Chile | Kernel density | 19.20 | Mean 20.5 days | GPS tracking | |
| Warembourg et al. (2021) | Chad | Biased random bridge, 95% isopleth | 7.70 | Median 60.3 h | GPS tracking | |
| Guatemala | 5.70 | |||||
| Indonesia | 5.60 | |||||
| Uganda | 5.70 | |||||
| This study | Cambodia | MCP 95% isopleth | 91.40 | Mean 25.5 days | GPS tracking |
MCP, minimum complex polygon.
Foray behaviour was quite variable, although forays commenced most commonly between 0000 and 0600 hours and rarely between 1200 and 1800 hours. This differs from dog forays recorded in Chile, with the majority occurring during the day (Sepulveda et al. 2015), and also differs to the camera trap data, in which the majority of detections, indicative of a foray, occurred during the day. The duration of forays rarely exceeded 4 h, and short forays were common. Dogs did not generally penetrate deeply within the wildlife sanctuary, but covered a fairly wide area, usually within 4 km of the village. Dogs M, J and I spent the most time within the wildlife sanctuary on forays, and only dog M spent time in areas where Eld’s deer are regularly detected by camera traps (Ladd 2022). Tracks within the sanctuary range from well-defined tracks suitable for four-wheel drive vehicles and small tractors to faint and partially reclaimed narrow tracks, but only a few tracks are mapped. It is evident that dog M sometimes travelled along tracks when on forays in the sanctuary, but the extent of off-track travel is unclear. The other dogs that made forays into the sanctuary were in areas with no mapped tracks, so it is unclear how much they may be using them.
It is possible that some forays by dogs were in the company of humans, but we were not able to get information on when owners were travelling with their dogs. Given the typical timings of forays, it is highly unlikely that the majority were in the company of humans. Dog K showed a persistence in using the same route down the main village road to return to the same location within the village on 6 out of 15 forays, which may suggest travelling with a household member to undertake some activity. However, the variation in timing, which included commencing just after midnight, in the early hours of the morning and at nearly 1700 hours, suggests that all of these trips are unlikely to have been accompanied by humans. Still, this dog showed the most consistency in terms of a repeated route used.
The small sample size of dogs in this study was due to a combination of factors, the most difficult being gaining owner permission to collar dogs. Villagers were unfamiliar with GPS technology and generally distrustful of authority. During recruitment, we made efforts to explain how the GPS collars worked, showed pictures of data points on a map to indicate what information was being collected, provided the collar and GPS logger for dog owners to examine, and described exactly what the data would and would not be used for. However, we were repeatedly asked about cameras and audio recorders, and concerns about data being shared with other agencies. This distrust was a difficult hurdle to overcome, with only a small number of willing owners found. Female members of households were generally more willing to give permission and showed less concern about the GPS tracking than male members. Subsequent studies in the village may meet with greater success by using this study to demonstrate the lack of negative impacts and to be able to show results that villagers can more easily relate to. However, achieving higher sample sizes would likely require significant investment in building trust with the villagers. Some sort of incentive, such as a health treatment for the dog (Warembourg et al. 2021), may aid in recruitment for future studies.
Due to limited funding, we were only able to deploy rudimentary GPS collars in this study. Using GPS collars that enabled greater controls over programming and accuracy (e.g. 5-min fixes) would have provided higher fidelity data. However, the bulkier, more sophisticated GPS collars may have caused local Khmer dog owners to be more reluctant to collar their dogs.
The camera trapping data suggest that dogs roam more widely in the sanctuary than indicated by the GPS tracking data, which is expected given the small sample size and relatively short duration of the tracking study. No collared dogs were captured by camera traps, with only one collared dog roaming into the camera trapping grid and only a very limited overlap in time when these cameras were active and the dog being collared. The camera trap information does need to be treated with caution, because the survey was not designed to detect free-roaming dogs, with cameras higher than optimal for an animal of this size and positioned off tracks (Ladd 2022). It is likely that the probability of detecting dogs is reduced, and that their actual distribution and relative abundance in the sanctuary is greater than indicated. It is also difficult to conclusively determine whether these dogs were truly unaccompanied by humans, because dogs tend to wander and are not always in close proximity to their owners (Ladd 2022). Despite these constraints, these data provide valuable insight into roaming and forest use by domestic dogs, and set the benchmark for further GPS radio telemetry studies over longer time frames.
Conclusion
The GPS and camera trap detections in this study have demonstrated that free-roaming village dogs in Cambodia have activity ranges that vary widely in size, with the median size being much larger than for home/activity ranges recorded by free-roaming dogs in other locations (Table 4). Greater distances are typically covered during night than in daylight, with female dogs generally having both smaller activity ranges and travelling less distance per day than male dogs. Activity ranges were centred around each dog’s household and were generally within the village area. Forays into the sanctuary were conducted by five of the 10 dogs for which forays were examined, and although it is unclear if any of these were in the company of humans, it was likely that a high proportion were unaccompanied. This study provides the first observations of the roaming behaviour of village dogs in Cambodia. It provides insight into the potential risk to threatened species in Siem Pang Wildlife Sanctuary by showing the roaming potential of village dogs, and also has implications for epidemiological studies of zoonoses. Further research is required to critically examine the impacts of free-roaming dogs on wildlife, as well as factors causing more frequent and extensive roaming behaviours.
Acknowledgements
We thank BirdLife International Cambodia Programme and Rising Phoenix Co. Ltd. for providing field support for this project. Thank you to the Cambodian villagers who supported our study and allowed us to radio collar their dogs.
References
Alexander KA, Kat PW, Munson LA, Kalake A, Appel MJG (1996) Canine distemper-related mortality among wild dogs (Lycaon pictus) in Chobe National Park, Botswana. Journal of Zoo and Wildlife Medicine 27, 426-427.
| Google Scholar |
Ballard G, Fleming PJS, Meek PD (2018) How long is a dingo’s tale?: impacts of sampling design on our understanding of dingo ecology. Australian Zoologist 39, 581-590.
| Crossref | Google Scholar |
Banks PB, Bryant JV (2007) Four-legged friend or foe? Dog walking displaces native birds from natural areas. Biology Letters 3, 611-613.
| Crossref | Google Scholar |
Burt WH (1943) Territoriality and home range concepts as applied to mammals. Journal of Mammalogy 24, 346-352.
| Crossref | Google Scholar |
Butler JRA, Bingham J (2000) Demography and dog-human relationships of the dog population in Zimbabwean communal lands. Veterinary Record 147, 442.
| Crossref | Google Scholar |
Chevalier V, Davun H, Sorn S, Ly P, Pov V, Ly S (2021) Large scale dog population demography, dog management and bite risk factors analysis: a crucial step towards rabies control in Cambodia. PLoS ONE 16, e0254192.
| Crossref | Google Scholar |
Cooper WE, Jr (1978) Home range criteria based on temporal stability of areal occupation. Journal of Theoretical Biology 73, 687-695.
| Crossref | Google Scholar |
Doherty TS, Dickman CR, Glen AS, Newsome TM, Nimmo DG, Ritchie EG, Vanak AT, Wirsing AJ (2017) The global impacts of domestic dogs on threatened vertebrates. Biological Conservation 210, 56-59.
| Crossref | Google Scholar |
Durr S, Ward MP (2014) Roaming behaviour and home range estimation of domestic dogs in Aboriginal and Torres Strait Islander communities in northern Australia using four different methods. Preventive Veterinary Medicine 117, 340-357.
| Crossref | Google Scholar |
Gatti A, Seibert JB, Moreira DO (2018) A predation event by free-ranging dogs on the lowland tapir in the Brazilian Atlantic Forest. Animal Biodiversity and Conservation 41, 311-314.
| Crossref | Google Scholar |
Gingold G, Yom-Tov Y, Kronfeld-Schor N, Geffen E (2009) Effect of guard dogs on the behavior and reproduction of gazelles in cattle enclosures on the Golan Heights. Animal Conservation 12, 155-162.
| Crossref | Google Scholar |
Gray TNE, Brook SM, McShea WJ, Mahood S, Ranjitsingh MK, Miyunt A, Hussain SA, Timmins R (2015) Rucervus eldii. [WWW document] The IUCN red list of threatened species 2015: e.T4265A22166803. Available at http://dx.doi.org/10.2305/IUCN.UK.2015-2.RLTS.T4265A22166803.en
Home C, Bhatnagar YV, Vanak AT (2018) Canine Conundrum: domestic dogs as an invasive species and their impacts on wildlife in India. Animal Conservation 21, 275-282.
| Crossref | Google Scholar |
Hughes J, Macdonald DW (2013) A review of the interactions between free-roaming domestic dogs and wildlife. Biological Conservation 157, 341-351.
| Crossref | Google Scholar |
Jenkins DJ, Andrew PL (1993) Intestinal parasites in dogs from an Aboriginal community in New South Wales. Australian Veterinary Journal 70, 115-116.
| Crossref | Google Scholar |
Kennedy B, Brown WY, Vernes K, Kurtner G, Butler JRA (2018) Dog and cat interactions in a remote aboriginal community. Animals 8, 65.
| Crossref | Google Scholar |
Kruuk H, Snell H (1981) Prey selection by feral dogs from a population of marine iguanas (Amblyrhynchus cristatus). Journal of Applied Ecology 18, 197-204.
| Crossref | Google Scholar |
Ladd R (2022) Deer, dogs, demographics and detection: conservation management of Eld’s deer (Rucervus eldii) in Cambodia. PhD thesis, School of Agriculture and Food Sciences, The University of Queensland. doi:10.14264/cff018f
Ladd R, Crouthers R, Brook S, Eames JC (2022) Reviewing the status and demise of the Endangered Eld’s deer and identifying priority sites and conservation actions in Cambodia. Mammalia 86, 407-421.
| Crossref | Google Scholar |
Loveridge R, Cusack JJ, Eames JC, Eang S, Willcox D (2018) Mammal records and conservation threats in Siem Pang Wildlife Sanctuary and Siem Pang Khang Lech Wildlife Sanctuary, Cambodia. Cambodian Journal of Natural History 2, 76-89.
| Google Scholar |
Manor R, Saltz D (2004) The impact of free-roaming dogs on gazelle kid/female ratio in a fragmented area. Biological Conservation 119, 231-236.
| Crossref | Google Scholar |
Meek PD (1999) The movement, roaming behaviour and home range of free-roaming domestic dogs, Canis Lupus familiaris, in coastal New South Wales. Wildlife Research 26, 847-855.
| Crossref | Google Scholar |
Morters MK, McKinley TJ, Restif O, Conlan AJK, Cleaveland S, Hampson K, Whay HR, Damriyasa IM, Wood JLN (2014) The demography of free-roaming dog populations and applications to disease and population control. Journal of Applied Ecology 51, 1096-1106.
| Crossref | Google Scholar |
Muinde P, Bettridge JM, Sousa FM, Dürr S, Dohoo IR, Berezowski J, Mutwiri T, Odinga CO, Fèvre EM, Falzon LC (2021) Who let the dogs out? Exploring the spatial ecology of free-roaming domestic dogs in western Kenya. Ecology and Evolution 11, 4218-4231.
| Crossref | Google Scholar |
Ortolani A, Vernooij H, Coppinger R (2009) Ethiopian village dogs: behavioural responses to a stranger’s approach. Applied Animal Behaviour Science 119, 210-218.
| Crossref | Google Scholar |
Ruiz-Izaguirre E, van Woersem A, Eilers KCHAM, van Wieren SE, Bosch G, van der Zijpp AJ, de Boer IJM (2015) Roaming characteristics and feeding practices of village dogs scavenging sea-turtle nests. Animal Conservation 18, 146-156.
| Crossref | Google Scholar |
Saavedra-Aracena L, Grimm-Seyfarth A, Schüttler E (2021) Do dog-human bonds influence movements of free-ranging dogs in wilderness? Applied Animal Behaviour Science 241, 105358.
| Crossref | Google Scholar |
Seebeck JH (1979) Status of the Barred Bandicoot, Perameles Gunnii, in Victoria: with a Note on Husbandry of a Captive Colony. Wildlife Research 6, 255-264.
| Crossref | Google Scholar |
Sepulveda M, Pelican K, Cross P, Eguren A, Singer R (2015) Fine-scale movements of rural free-ranging dogs in conservation areas in the temperate rainforest of the coastal range of southern Chile. Mammalian Biology 80, 290-297.
| Crossref | Google Scholar |
Sparkes J, Kortner G, Ballard G, Fleming PJS, Brown WY (2014) Effects of sex and reproductive state on interactions between free-roaming domestic dogs. PLoS ONE 9, e116053.
| Crossref | Google Scholar |
Taborsky M (1988) Kiwis and dog predation: observations in Waitangi State Forest. Notornis 35, 197-202.
| Google Scholar |
Van Kesteren F, Mastin A, Mytynova B, Ziadinov I, Boufana B, Torgerson PR, Rogan MT, Craig PS (2013) Dog ownership, dog behaviour and transmission of Echinococcus spp. in the Alay Valley, southern Kyrgyzstan. Parasitology 140, 1674-1684.
| Crossref | Google Scholar |
Warembourg C, Wera E, Odoch T, Bulu PM, Berger-González M, Alvarez D, Abakar MF, Maximiano Sousa F, Cunha Silva L, Alobo G, Bal VD, López Hernandez AL, Madaye E, Meo MS, Naminou A, Roquel P, Hartnack S, Dürr S (2021) Comparative study of free-roaming domestic dog management and roaming behavior across four countries: Chad, Guatemala, Indonesia, and Uganda. Frontiers in Veterinary Science 8, 617900.
| Crossref | Google Scholar |
White PCL, Ward AI (2010) Interdisciplinary approaches for the management of existing and emerging human-wildlife conflicts. Wildlife Research 37, 623-629.
| Crossref | Google Scholar |
Wierzbowska IA, Hędrzak M, Popczyk B, Okarma H, Crooks KR (2016) Predation of wildlife by free-ranging domestic dogs in Polish hunting grounds and potential competition with the grey wolf. Biological Conservation 201, 1-9.
| Crossref | Google Scholar |
Zapata-Rios G, Branch LC (2016) Altered activity patterns and reduced abundance of native mammals in sites with feral dogs in the high Andes. Biological Conservation 193, 9-16.
| Crossref | Google Scholar |