

Monitoring methods influence native predator detectability and inferred occupancy responses to introduced carnivore management
Tim S. Jessop A * and Graeme R. Gillespie B
A * and Graeme R. Gillespie B
A Centre For Integrative Ecology, School of Life and Environmental Sciences, Deakin University, Waurn Ponds, Vic. 3216, Australia.
B Department of Environment and Natural Resources, Palmerston, NT 0830, Australia.
Wildlife Research 50(1) 16-27 https://doi.org/10.1071/WR22012
Submitted: 24 January 2022 Accepted: 14 April 2022 Published: 4 July 2022
© 2023 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Context: Management actions that suppress introduced predator densities can benefit the population recovery of native species. Nevertheless, ensuring that predator management produces measurable population-level benefits can be influenced by multiple factors affecting species detection. Monitoring designs using multiple survey methods may perform better than increasing sampling effort with single-method protocols.
Aims: This study aimed to estimate individual and cumulative detection probabilities and site occupancy estimates from the use of five different monitoring methods to survey a native mesopredator, the lace monitor (Varanus varius). Second, we assessed the effect of lethal red fox (Vulpes vulpes) baiting on lace monitor detection probabilities and site occupancy estimates collected from each monitoring method.
Methods: Multi-method sampling for Varanus varius occurred at 76 sites across lethal fox baited and non-baited habitats in East Gippsland, Victoria. Bayesian site occupancy models were used to estimate the effects of detection method and fox-baiting treatments on Varanus varius detection probability and site occupancy.
Key results: Method-specific detection probabilities (P = 0.00–0.12) and site occupancy estimates (Ψ = 0–0.53) varied considerably among methods, but combinations of multi-method monitoring improved lace monitor detection probability (P = 0.11–0.18) and site occupancy (Ψ = 0.87 ± [0.66–0.93]−0.91 ± [0.76–0.97] mean ± [95% credible intervals]) above any single method. However, there was extreme heterogeneity in the size and direction of the introduced predator baiting effect on method-specific lace monitor detection. Three methods (box traps and two different visual search surveys) all indicated lace monitor detection probabilities increased in fox-baited sites. However, sand pads reported a decrease in lace monitor detection at fox-baited sites, whereas pipe traps obtained no detections.
Conclusions: Combining detection data from all methods led to the inference of a positive fox-baiting effect, albeit with a smaller magnitude and better certainty than that estimated using a reduced method monitoring design, which had fewer detection data after excluding biased detection from sand pads.
Implications: Using a multi-method monitoring approach improved lace monitor detection and reduced sampling effort. However, depending on sampling methodology, the management effects on lace monitors can change.
Keywords: biodiversity monitoring, detection method evaluation, detection probability, lace monitor, lethal fox baiting, management inference, site occupancy, Varanus varius.
Introduction
Introduced mammalian predators have caused globally significant ecological impacts, including the decline and extinction of native species (Doherty et al. 2016). For instance, the 19th century introduction of the European red fox (Vulpes vulpes) into Australia has had well documented and continental-scale biodiversity impacts (Glen and Dickman 2008; Saunders et al. 2010; Woinarski et al. 2019). Since the 1980s, landscape-scale poison baiting programmes have been a key management action used to suppress the density of introduced fox populations across Australia (Kinnear et al. 2002; Hayward and Somers 2012; Braysher 2017; Legge et al. 2018). Fox baiting has produced measurable biodiversity benefits for both threatened and non-threatened native species, and helped restore ecological processes (Kinnear et al. 1998; Dexter and Murray 2009; Claridge et al. 2010; de Tores and Marlow 2012). However, it is also sometimes evident that fox baiting may have weak, absent, or unintended (e.g. introduced mesopredator or herbivore release) biodiversity consequences (Walsh et al. 2012; Marlow et al. 2015; Lindenmayer et al. 2018; Jessop et al. 2021). Evaluating the processes underpinning ineffective or unforeseen native biodiversity responses from fox-baiting outcomes is essential for innovating and guiding future management plans, and helps ensure that scarce conservation funds can best achieve positive conservation benefits (Walsh et al. 2012; Lindenmayer et al. 2018). Not discounting that environmental, ecological, or baiting protocol-related processes could affect the magnitude and direction of native biodiversity responses to fox-baiting programmes. It is also important to recognise that the monitoring method(s) and associated experimental designs used to measure native species responses can have significant implications for data quality, survey success, and arising population response estimates used to evaluate wildlife management effectiveness (Thompson and Thompson 2007; Nichols et al. 2008; Lindenmayer et al. 2020). For example, indirect or direct population estimates for a target species obtained from different monitoring methods may be poorly correlated and indicate method-specific detection biases (Long et al. 2007, 2012; Hayward et al. 2015). Similarly, monitoring methods that use baits, lures or traps can affect animal behaviour that may positively or negatively influence detection, and consequently bias population-level estimates (Thompson 2013; Comer et al. 2018; Stewart et al. 2019). Ultimately, any monitoring method that achieves poor, or biased, detection probability could obscure environmental managers from accurately assessing their actions on wildlife populations (Yoccoz et al. 2001; Kéry and Schmidt 2008).
Population monitoring designs that use multiple concurrent methods have been advocated to reduce uncertainty produced by low detection probability from single method monitoring designs (Bailey et al. 2004; Nichols et al. 2007; Otto and Roloff 2011). Multi-method monitoring protocols can be particularly advantageous if they increase sampling opportunities to reduce imperfect detection (MacKenzie et al. 2017; Einoder et al. 2018). Similarly, through improved detection, such designs might reduce sampling effort compared with single method sampling designs. Additionally, one monitoring method may have relatively high detectability, but may be biased if it only detects part of the population. Then integration with other methods can allow concurrent methods to better obtain unbiased estimates (Descalzo et al. 2021). Using multiple methods is not without problems, including that different methods can achieve dissimilar detection of target species, or if one method is far better, it can produce inefficient monitoring (Nichols et al. 2008; Mattfeldt et al. 2009). Similarly, if multiple methods within a sampling site lack spatial independence, they may produce overlapping and correlated detections that produce inefficient monitoring (Clare et al. 2017). Also, method-specific biases can increase or decrease detection probability relative to the population’s actual occurrence (Bailey et al. 2004). In such cases, method-specific biases may propagate uncertainty in estimates of environmental management effects on population responses (Nichols and Williams 2006; MacKenzie et al. 2017).
This study’s objective was to use a multi-method monitoring design to address two aims: (1) compare how survey methods vary in their individual and combined ability to influence detection and site occupancy of a varanid lizard, the lace monitor (Varanus varius); and (2) evaluate how red fox removal (via poison baiting) influenced lace monitor detectability and site occupancy. The lace monitor is a sizeable semiarboreal monitor lizard that can weigh up to 14 kg and attain a maximum body length of approximately 2 m (Guarino 2002; Jessop et al. 2010; Anson et al. 2014; Fig. 1a). It is distributed throughout the non-arid regions of eastern Australia (Smissen et al. 2013). As a generalist predator, lace monitors consume a broad array of prey, including insects, small to medium-sized mammals, and carrion (Guarino 2002; Jessop et al. 2012). These monitor lizards are expected to provide an important trophic influence in Australia’s terrestrial ecosystems (Doody et al. 2015; Feit et al. 2020). However, the way different monitoring methods influence lace monitor detection and site occupancy (and thus their potential to benefit from lethal fox-baiting programmes) remains poorly understood.

|
The first goal was to assess how the use of multi-method monitoring design would allow us to measure the extent to which different survey methods varied in their ability to detect lace monitors and influence method-specific site occupancy estimates. It was presumed that methods could vary substantially in their detection of lace monitors, but overall, a multi-method monitoring design would better reduce the total survey effort needed to conduct population monitoring and achieve good estimates of site occupancy. A reduced survey effort was considered especially important given our study area experiences a highly seasonal climate that restricts reptile activity (and hence detection) to a relatively short summer period (Jessop et al. 2013). Similarly, demonstrating method-specific differences in detection is essential to inform how to best allocate survey effort in future monitoring of lace monitor populations.
The second goal was to determine if different monitoring methods consistently produced results that inform whether fox removal is an effective tool for promoting native wildlife. Here we compared the effect of a large-scale introduced fox lethal baiting programme on method-specific lace monitor detection probabilities and site occupancy. Prior studies have demonstrated that monitor lizard populations can sometimes benefit from fox baiting, possibly because it releases these native predators from predation or competition (Anson et al. 2013; Anson et al. 2014; Hu et al. 2019; Jessop et al. 2021). However, it remains unclear if different monitoring methods also allow reporting consistency in how fox baiting influences monitor lizard detection and site occupancy. If all methods are relatively effective and unbiased, they should produce consistent detection responses to fox baiting (Read and Scoleri 2015). Conversely, the effect of fox baiting could lead to method-specific lace monitor sampling biases. For example, different fox densities between baited and non-baited landscapes could influence how lace monitors (i.e. a native mesopredator) competitively interact with this apex predator (Ritchie and Johnson 2009; Anson et al. 2013). One consequence could be that lace monitors alter behaviour, such as risk-sensitive foraging or daily activity patterns, in unbaited habitats with higher fox densities (Moll et al. 2017). Differences in lace monitor foraging activity could allow sampling biases in monitoring methods that use food-based baits to detect individuals (Anson et al. 2013; Moll et al. 2017). If so, method-related detection biases could affect site occupancy estimates and thus weaken evidence to deduce if fox baiting benefits this lace monitor population.
Methods
Study area
The study area comprised 42 000 ha of coastal forest located within the Cape Conran Coastal Park (37°490S, 148°440E) and Murrungowar State Forest (37°568S, 148°753E) in East Gippsland, Victoria, Australia. Mean maximum and minimum temperatures ranged from 27.0°C (January) to 4.7°C (July), and annual rainfall for the region averages ∼846 mm (http://www.bom.gov.au/climate/averages/tables/cw_084030.shtml, accessed 12 February 2021). Within the area, study sites were primarily located in two widespread vegetation types: (1) coastal woodland dominated by Banksia integrifolia, and Banksia serrata; and (2) lowland forest dominated by Eucalyptus globoidea and Eucalyptus sieberi (see Anson et al. 2013). A minority of sites consisted of heathland or vegetation ecotones. Generally, the habitat transitioned from Banksia woodland near the coast into eucalypt forest further inland.
Introduced predator management
The study area was overlaid onto the Southern Ark programme’s experimental fox population management treatments. This programme is a large-scale and ongoing fox suppression programme managed by the Department of Environment, Land, Water and Planning, Victoria, Australia (Murray et al. 2006; Dexter and Murray 2009). It was initially established to examine fox suppression effects on native threatened mammals. The programme area comprised two management treatments, each with two replicate blocks (∼6000 ha each). Two blocks were assigned as areas of lethal fox baiting to cause fox population density suppression, and two replicates were assigned as control block areas (i.e. deployment of poison-free baits). To avoid any potential ‘halo effect’ where lethal fox baiting can decrease fox or increase native animal densities in non-baited habitat immediately adjacent to the baited area (Glen et al. 2013), a minimum boundary area of 2 km was used to promote spatial independence between and within each management treatment. Before the commencement of this study, poison and control baits had been deployed consistently within their respective blocks since May 1999 (Murray et al. 2006; Dexter and Murray 2009).
Fox suppression was achieved using Foxoff Econobaits (Animal Control Technologies, Melbourne), a commercially available manufactured bait, pre-poisoned with 3 mg of sodium fluoroacetate (‘1080’ poison). This sodium fluoroacetate dose is lethal to canids but not toxic to varanid lizards (Mcilroy et al. 1985; Twigg and King 1991).
Study design and monitoring methods
In the summer of 2008/2009, we established 76 multi-method monitoring sites over the study area (Fig. 1b). We allocated 38 monitoring sites each to the lethal fox-baited treatment and the non-lethal baited control treatment. Each sampling site (15 ha, 500 m long × 300 m wide) was accessed using unsealed management or logging tracks. All sites were at least 2 km apart to promote spatial independence in lace monitor detections among sites (Jessop et al. 2013). Monitoring at each site was conducted for six consecutive days and only during periods with consistently warm days (>25°C) and clear sky (<25% cloud cover), to limit false absences due to unfavourable climatic conditions reducing lizard activity (Jessop et al. 2013). At each site, five concurrent monitoring methods (Fig. 1c) were used to detect lace monitors:
-
Visual drive survey: Two observers in a vehicle travelling at 10 km/h searched for lace monitors along 500 m of management track, which comprised the midline of each site (Fig. 2a). The search area consisted of searching forward from the vehicle’s mid-line out to 90° on either side of the observer’s position. The search area was constrained to a maximum distance of 30 m on either side of the vehicle. This was considered the maximum distance one could reliably see a monitor lizard in forested habitats. Visual drive surveys were the first method used to detect lace monitors in each daily survey. Furthermore, we always surveyed from the first to the last site before we returned to assess for additional lace monitor detections from the remaining four monitoring methods within each site. This method resulted in a standard area (i.e. 500 m × 60 m) being sampled on each census;
-
Mixed visual search: This type of search comprised driving through each site at 40 km/h and then stopping the vehicle at regular intervals to walk into the forest to check each site’s box trap, sand pads and pipe traps. Monitor lizards observed either from the vehicle or on foot within a site during these searches (15 min/site) were recorded. To reduce the potential for repeated detections of any lace monitors encountered at a site during the aforementioned visual drive survey, we waited for 30 min before conducting the mixed visual search method;
-
Box traps: One aluminium box trap (2 × 0.3 × 0.3 m) for lace monitors was positioned randomly within each site to allow direct capture of individual lizards (Fig. 2b). Traps were baited with raw beef infused with tuna emulsion oil. Meat baits were replaced if removed or on the morning of the 4th day of the survey if not consumed. Replacement of unused baits on the morning of the 4th day was done to limit variation in bait attractiveness (i.e. mass and odour) and reduce heterogeneity in daily bait detection by lace monitors across the survey period. This step was informed from a similar study that demonstrated that monitoring methods that do not replace meat baits after 3 days had reduced varanid lizard detections (Ariefiandy et al. 2013). All individual lizards were microchipped to provide a means to record the potential for repeat captures (i.e. non-independent detections);
-
Sand pads: Two sand pads (75 cm in diameter and 10 cm high) were constructed from beach sand (Fig. 2c). A small piece of cow heart covered in tuna oil was buried to a 10 cm depth in each pad’s centre. Meat baits in sand pads were replaced if removed or on the morning of the 4th day of the survey if not consumed. Because lace monitors are the only large lizard in the study area, their presence on sand pads was determined from their diagnostic claw and tail drag marks (Jessop et al. 2013);
-
Pipe traps: Two PVC pipes (1.5 m length × 15 cm diameter) closed-off at one end were placed vertically on two large trees and secured with rubber strapping (Fig. 2d). Traps were baited with raw beef. Meat baits were replaced if removed or on the morning of the 4th day of the survey if not consumed. The size of pipe traps and their placement were identical to those used to successfully capture arboreal juveniles of another large varanid lizard, the Komodo dragon (Varanus komodoensis) (Purwandana et al. 2021).

|
The locations of box and pipe traps and sand pads were assigned using a random number table to give x (0–500 m) and y (−150–150 m) coordinates within each site. All monitoring methods were positioned a minimum of 50 m away from the nearest edge of the management track. Each monitoring method was separated by a minimum distance of 100 m from any other method to reduce the likelihood of overlapping detections among methods. The spacing interval among each method was based on a pilot study conducted in 2006/2007, which indicated limited overlap in lace monitor detections between box traps deployed at 100-m intervals. In the current study, we further conducted a two-tailed Pearson correlation test with 1000 bootstraps to statistically assess the extent of correlations among method-specific detection patterns within sites.
Estimating detection and site occupancy estimates
Hierarchical occupancy models (MacKenzie et al. 2006; Nichols et al. 2007) were used to estimate method-specific lace monitor detectability and site occupancy, and the effect of introduced predator baiting on these estimates. An integrated monitoring design assumes that each sampling method is sufficiently separated to produce independent detections (Clare et al. 2017; Descalzo et al. 2021). A consequence of non-independence between methods is expected to inflate the overall detection probability of a species and produce a negative bias in state parameters (Clare et al. 2017). Although our sampling methods were spatially discrete, we did not explicitly test this assumption by using occupancy models that can accommodate observation covariance among each survey method (Clare et al. 2017). However, we did qualitatively assess the capacity of our sampling design to result in non-independence of detections via assessing the recapture rate of lace monitors within and between sites, and by assessing the degree of correlation between detection methods across sampling sites.
To address our aims, we modelled the occurrence of lace monitors at site i as a draw from the Bernoulli distribution with parameter ψ, which is the probability that the lace monitors occupies site i:
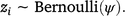
An occurrence is represented by zi, = 1, and an absence is represented by zi, = 0.
It was assumed that a site was either occupied or not across all sampling occasions and that the probability that a site was occupied was the same for all sites.
We modelled the probability of detecting lace monitors given that they were present using method j for n surveys at site i as a draw from the Bernoulli distribution with parameters d,j, which is the probability of detecting lace monitors using method j on a single sampling occasion given they were present. Thus, we calculated:
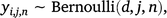
where a detection is represented by yi,j,n = 1, and non-detection is represented by:
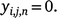
Combining these two processes produced the observation model that modelled the observations as the product of occupancy and detection:
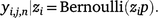
An observation is represented by yi,j,n |zi = 1, and a failure to observe lace monitors is represented by yi,j,n|zi = 0.
The model was run using Winbugs 1.4.2 (Lunn et al. 2000) called from programme Presence 2.12.36 (Hines 2006). A uniform prior distribution for occupancy and detection probabilities was used to represent a lack of prior information and ensure that the parameter estimates were driven by the data. Parameter estimates were based on 1000 samples subsampled from 5000 samples after a 5000 burn in, which was more than sufficient for WinBUGS to stabilise convergence.
Following Descalzo et al. (2021), we first ran occupancy models that combined all five sampling methods as a reference to then compare the mean posterior distribution estimates [±95% credible intervals (CI)] for detection probabilities and site occupancy against models that were evaluated for each individual method and combinations of the four, three and two best lace monitor sampling methods. To compare individual and combined sampling method performance, we used an accuracy index that combines measures of bias and precision through estimating the mean square error (MSE) to assess the performance of each individual, or the combinations, of sampling methods:
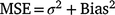
Thus, we estimated for each lace monitor sampling method and combinations therein the probability of detection (mean and s.d.), occupancy (mean, s.d. and coefficient of variation), and the MSE.
Next, we ran occupancy models that tested the effect of fox baiting on the individual method-specific probability of lace monitor detection reported. Similarly, we tested the effect of fox baiting on combinations of methods used to estimate lace monitor site occupancy. The strength of evidence for meaningful differences between posterior mean detection probabilities from the different monitoring methods was evident when their 95% CIs did not overlap. Similarly, evidence for a strong posterior effect of introduced predator management on lace monitor detection and site occupancy was obtained when their 95% CIs did not overlap (McCarthy 2007).
Ethical standards
The protocols used were approved by the Animal Ethics Committee of the University of Melbourne (Permit Number: 0911328). The research was carried out on public land under a Victorian Department of Sustainability Wildlife and National Parks Act (1975) research Permit 10005037.
Results
Method specific detection probability and site occupancy
We surveyed 76 composite monitoring sites for six consecutive days, resulting in 109 lace monitor detections at 52 sites. Box traps, sand pads, mixed visual searches, visual drive surveys, and pipe traps reported 43, 43, 13, 10 and 0 lace monitor detections respectively. At least from box trap-related detections (i.e. direct capture), there was little evidence (two of 43 individuals) that the same individual was detected on multiple occasions (i.e. false positives) within the sampling period. There was no evidence of significant correlations among detections reported from box traps, sand pads, mixed visual search or the visual drive survey within sites (Pearson two-tailed test: all P-values > 0.05; pairwise Pearson R2 = −0.060–0.190) (Supplementary Table S1, available online as Supporting Information).
The method-specific estimates of lace monitor detection probability (P) ranged from 0.000 to 0.121 (Table 1). Box traps (P = 0.120 ± [0.086–0.162] [mean ± 95% credible intervals (CI)]) and sand pads (P = 0.121 ± [0.086–0.160]) performed similarly and substantially better than the visual search (P = 0.030 ± [0.015–0.049]) and mixed visual search surveys (P = 0.038 ± [0.021–0.060]). Because pipe traps failed to detect lace monitors, no estimates of detection were possible. Overall, pending the combination of multiple sampling methods, the detection probability of lace monitors could be increased above estimates obtained from any single best method.

|
The method-specific lace monitor site occupancy (ψ) estimates ranged from 0.00 to 0.53 (Table 1). Box traps (ψ = 0.53 ± [0.03–0.73] [mean ± 95% credible intervals (CI)]) and sand pads (ψ = 0.49 ± [0.03–0.660]) produced higher occupancy estimates that the mixed visual search (ψ = 0.28 ± [0.07–0.68]) and visual search surveys (ψ = 0.24 ± [0.06–0.60]). Because pipe traps failed to detect lace monitors, no occupancy estimates could be obtained. It was evident that different combinations of multi-method monitoring designs substantially increased the lace monitor site occupancy estimate compared with any single method used (Table 1). Indeed, all individual methods reported a strong negative bias and higher MSE scores than sampling designs that incorporated two or more sampling methods (Table 1). Furthermore, it was evident that reduced combinations of different sampling methods could lead to a similar performance in site occupancy estimates compared with the full method sampling design (Table 1).
Lethal fox-baiting effects on method-specific lace monitor detection probability and site occupancy
The magnitude of the fox-baiting effect on detection probabilities varied depending on the monitoring method used to make the assessment (Fig. 3a). Baiting resulted in a positive effect on lace monitor detectability reported from box traps (P baiting effect = 0.045 ± [−0.024–0.120]), visual search (P baiting effect = 0.035 ± [0.001–0.072]) and mixed visual search (P baiting effect = 0.038 ± [−0.002–0.079]) methods (Fig. 3b). However, the lace monitor detection probability on sand pads exhibited a strong negative baiting effect (P baiting effect = −0.150 ± [−0.235 to −0.074]), indicating that lace monitor detections increased on sand pads in unbaited control sites compared with sites sampled in fox-baited sites. There was no effect of fox baiting on pipe trap detection probability (P baiting effect = 0.000 ± [0.010 to −0.010]).

|
The use of combined detection methods indicated that lace monitor site occupancy was higher at fox-baited sites (ψ = 0.867 ± [0.543–0.994]; naive occupancy = 30 of 38 sites occupied) than at unbaited sites (ψ = 0.645 ± [0.323–0.904], naive occupancy = 22 of 38 sites occupied) (Fig. 4a). To further consider the influence of the negative fox-baiting effect on lace monitor detection probability estimated from sand pads, we removed these data and reanalysed our estimates using the remaining four methods. Removal of sand pad data reduced estimates of lace monitor site occupancy by ∼8% across the study area (four methods – sand pad detection data ψ = 0.794 ± [0.665–0.923], naive occupancy = 52 of 76 sites occupied vs five methods ψ = 0.711 ± [0.486–0.870], naive occupancy = 43 of 76 sites). This effect was most evident in unbaited sites where site occupancy was reduced by 15% compared with that estimated using all five methods combined (Fig. 4b). This in turn caused the mean fox-baiting effect on lace monitor site occupancy to increase by ∼50% (B = 0.479 ± [−0.012–0.973] vs 0.241 ± [0.080–0.442]). However, this revised estimate had much larger credible intervals (i.e. spanned zero), meaning evidence of a fox-baiting effect on lace monitor site occupancy was weakened relative to the original estimate (Fig. 4b).

|
Discussion
Introduced mammalian predators, such as the red fox, remain a significant threat to Australia’s biodiversity (Radford et al. 2018; Hradsky et al. 2019). As important native Australian predators, varanid lizards are expected to benefit in landscapes where management actions have caused adequate exclusion or suppression of fox populations (Anson et al. 2014; Read and Scoleri 2015; Hu et al. 2019). This study assessed how five different monitoring methods and lethal fox baiting influenced lace monitor detection and site occupancy estimates.
Our results indicated that different monitoring methods varied considerably in their capacity to detect lace monitors and influence site occupancy estimates. Box traps and sand pads were 3–4-fold more effective in obtaining lace monitor detections than either of the visual search methods, and pipe traps were entirely ineffective. These detection differences similarly translated into a two-fold increase in site occupancy estimates for box traps and sand pads over both visual search methods. Higher detection and site occupancy estimates obtained at baited box traps and sand pads likely occurred because both methods use meat baits to attract lace monitors (Ariefiandy et al. 2013; du Preez et al. 2014; Comer et al. 2018). Visual survey methods had lower detection and site occupancy, possibly due to a reliance on random encounters, less sampling effort (i.e. time duration) and the difficulty of sighting lace monitors in forest (Griffiths and McKay 2007; Ariefiandy et al. 2014; Boback et al. 2020). Unlike other varanid lizard studies, pipe traps failed to capture any juvenile lace monitors, which are presumably a significant fraction of the population (Imansyah et al. 2008; Jessop et al. 2020).
A key advantage of multi-method monitoring protocols was to improve combined estimates of lace monitor detection and site occupancy over any single method (i.e. high bias, low detection) (MacKenzie et al. 2017; Einoder et al. 2018; Descalzo et al. 2021). It is assumed that our multi-, relative to single, method monitoring protocols increased lace monitor detections and site occupancy estimates by multiple processes. First, having additional detectors within a sampling site would increase the sampling effort (i.e. total detection area and sampling duration) relative to using any single method. Increased site-specific sampling effort would thus increase the probability that lace monitors are detected during the survey. Second, each sampling device is expected to be associated with detection biases that favour or preclude different individuals (e.g. life stages, behavioural phenotypes) of the lace monitor population to be detected. Thus, when different methods are combined, they are expected to increase the overall proportion of individuals available for detection (Clare et al. 2017; Descalzo et al. 2021). For example, as discussed below, sand pads could detect neophobic or wary individuals that might otherwise avoid entering box traps. Thus, the combined use of sand pads and box traps within sites is expected to sample a more significant proportion of the lace monitor population than each method alone.
Additionally, there was no substantial evidence of correlated lace monitor detections within or among methods, suggesting the improved efficacy of different methods to record largely independent site lace monitor detections. Thus, multi-method monitoring considerably reduced sampling effort compared with the single best method (e.g. box traps) for detecting this native predator (Nichols et al. 2008; Martin et al. 2009). Similarly, just using the single best sampling method would have required a greatly expanded survey effort that would have been both costly and potentially unfeasible, given that a highly seasonal climate reduces annual lace monitor activity in the study area (Michael et al. 2012; Jessop et al. 2013; McGrath et al. 2015). Furthermore, it was possible to ascertain the most effective sampling design by comparing how different combinations of methods influenced the accuracy and precision of lace monitor detection and site occupancy estimates. It was evident that using the three best methods (box traps, sand pads and mixed visual search) could produce similar results with similar accuracy (i.e. low bias) and precision (i.e. low CV) to that obtained from all five methods. Consequently, future studies that used this reduced design would benefit from less survey effort and cost without loss of monitoring performance.
This study’s second aim was to assess how different monitoring methods could influence inference due to the effect of fox baiting on lace monitor detection and site occupancy. A multi-method monitoring design demonstrated that fox baiting was associated with method-specific heterogeneity in lace monitor detection probabilities (Boback et al. 2020). Of the four successful detection methods, three (i.e. box traps and two visual survey methods) indicated small positive effects of fox baiting on lace monitor detection. Positive increases in detection with fox baiting are consistent with lace monitors experiencing mesopredator release, resulting in higher population densities in fox-suppressed landscapes (Anson et al. 2014; Read and Scoleri 2015; Jessop et al. 2016; Hu et al. 2019).
In contrast, the lace monitor detection probability estimated from sand pads was higher at unbaited sites, indicating a strong negative fox-baiting effect. This reversed detection probability effect on sand pads is possibly the consequence of the interplay between food rewards (i.e. meat bait within sand pads) and introduced predator-mediated influences on lace monitor foraging behaviour (Anson et al. 2013; Jessop et al. 2015). In particular, risk-sensitive foraging theory predicts that prey depletion by competitors is likely to affect food-based reward–risk ratios influencing the foraging decisions of inferior competitors (Caraco et al. 1980; Barnard and Brown 1985). So if lace monitors exhibit stronger competition-induced reward-motivated foraging behaviour for novel meat baits, it could explain this positive bias in sand pad-based detections in non-fox-baited sites compared with fox-baited sites (Phillips and Winchell 2011; Willson et al. 2011). Behavioural studies of monitor lizards in areas with and without baiting for foxes could help clarify this scenario.
How could method-specific sampling biases affect the capacity to determine if fox baiting benefited the lace monitor population within the study area? For example, because sand pads produced a positive detection bias, this inflated lace monitor site occupancy in non-fox baited sites. This result reduced the overall fox-baiting effect on lace monitor site occupancy estimated across the study area. Re-analysis excluding the sand pad data produced a more significant mean fox-baiting effect on lace monitor site occupancy. However, removal of the sand pad detection data produced an estimate with larger credible intervals, reducing the evidence of a fox-baiting effect on the lace monitor population. Thus, these results highlight how the use of multiple monitoring methods, while producing better overall detection, may nevertheless influence the assessment of target species population management (Nichols et al. 2008).
Since this study was undertaken, we note that camera traps have been demonstrated to be a highly effective monitoring tool for some larger reptile species, including monitor lizards, with high and unbiased detection probabilities reported (Ariefiandy et al. 2013, 2014; Einoder et al. 2018; Moore et al. 2020). Therefore, replacing sand pads with baited camera traps in our multi-method approach could retain high, but less biased, detection that would increase the magnitude and certainty that fox baiting had a positive effect on lace monitor populations (Ariefiandy et al. 2013; Jessop et al. 2013; Purwandana et al. 2014). Noting that camera traps can be placed in the environment for relatively long periods, without checking, extending the time during which detections can be obtained.
Management implications
There are two broader outcomes for management from this study. First, integrating different monitoring methods can improve detection and site occupancy estimates for target wildlife species because it inherently increases the number of detections and reduces the influence of species-specific detection biases or method failures (Stokeld et al. 2015; Boback et al. 2020). This result is significant because it can often be challenging to infer target species responses to management actions if they have poor detection. Second, even when individual methods detect target species well, they might not accurately report management effects on wildlife due to detection biases that affect site occupancy estimates. Thus wildlife ecologists must be aware of the strengths and limitations of different detection methods, and associated sampling biases, to best demonstrate the benefits of wildlife management for biodiversity (Lindenmayer and Likens 2010; Hayward et al. 2015).
Supplementary material
Supplementary material is available online.
Data availability
The data supporting this study will be shared upon reasonable request to the corresponding author.
Conflicts of interest
The authors have no conflicts of interest to declare.
Declaration of funding
Zoos Victoria provided funding for this project.
Author contributions
Study design and fieldwork: TSJ and GRG; data analysis: TSJ; writing: TSJ and GRG.
Acknowledgements
We thank Tim Lockwood for his assistance in the field.
References
Anson, JR, Dickman, CR, Boonstra, R, and Jessop, TS (2013). Stress triangle: do introduced predators exert indirect costs on native predators and prey? PLoS ONE 8, e60916.| Stress triangle: do introduced predators exert indirect costs on native predators and prey?Crossref | GoogleScholarGoogle Scholar | 23585861PubMed |
Anson, JR, Dickman, CR, Handasyde, K, and Jessop, TS (2014). Effects of multiple disturbance processes on arboreal vertebrates in eastern Australia: implications for management. Ecography 37, 357–366.
| Effects of multiple disturbance processes on arboreal vertebrates in eastern Australia: implications for management.Crossref | GoogleScholarGoogle Scholar |
Ariefiandy, A, Purwandana, D, Seno, A, Chrismiawati, M, Ciofi, C, and Jessop, TS (2014). Evaluation of three field monitoring-density estimation protocols and their relevance to Komodo dragon conservation. Biodiversity and Conservation 23, 2473–2490.
| Evaluation of three field monitoring-density estimation protocols and their relevance to Komodo dragon conservation.Crossref | GoogleScholarGoogle Scholar |
Ariefiandy, A, Purwandana, D, Seno, A, Ciofi, C, and Jessop, TS (2013). Can camera traps monitor Komodo dragons a large ectothermic predator? PLoS ONE 8, e58800.
| Can camera traps monitor Komodo dragons a large ectothermic predator?Crossref | GoogleScholarGoogle Scholar | 23527027PubMed |
Bailey, LL, Simons, TR, and Pollock, KH (2004). Estimating site occupancy and species detection probability parameters for terrestrial salamanders. Ecological Applications 14, 692–702.
| Estimating site occupancy and species detection probability parameters for terrestrial salamanders.Crossref | GoogleScholarGoogle Scholar |
Barnard, CJ, and Brown, CAJ (1985). Risk-sensitive foraging in common shrews (Sorex araneus L.). Behavioral Ecology and Sociobiology 16, 161–164.
| Risk-sensitive foraging in common shrews (Sorex araneus L.).Crossref | GoogleScholarGoogle Scholar |
Boback, SM, Nafus, MG, Yackel Adams, AA, and Reed, RN (2020). Use of visual surveys and radiotelemetry reveals sources of detection bias for a cryptic snake at low densities. Ecosphere 11, e03000.
| Use of visual surveys and radiotelemetry reveals sources of detection bias for a cryptic snake at low densities.Crossref | GoogleScholarGoogle Scholar |
Braysher M (2017) ‘Managing Australia’s Pest Animals: a Guide to Strategic Planning and Effective Management.’ (CSIRO Publishing: Melbourne, Vic., Australia)
Caraco, T, Martindale, S, and Whittam, TS (1980). An empirical demonstration of risk-sensitive foraging preferences. Animal Behaviour 28, 820–830.
| An empirical demonstration of risk-sensitive foraging preferences.Crossref | GoogleScholarGoogle Scholar |
Clare, J, McKinney, ST, DePue, JE, and Loftin, CS (2017). Pairing field methods to improve inference in wildlife surveys while accommodating detection covariance. Ecological Applications 27, 2031–2047.
| Pairing field methods to improve inference in wildlife surveys while accommodating detection covariance.Crossref | GoogleScholarGoogle Scholar | 28644579PubMed |
Claridge, AW, Cunningham, RB, Catling, PC, and Reid, AM (2010). Trends in the activity levels of forest-dwelling vertebrate fauna against a background of intensive baiting for foxes. Forest Ecology and Management 260, 822–832.
| Trends in the activity levels of forest-dwelling vertebrate fauna against a background of intensive baiting for foxes.Crossref | GoogleScholarGoogle Scholar |
Comer, S, Speldewinde, P, Tiller, C, Clausen, L, Pinder, J, Cowen, S, and Algar, D (2018). Evaluating the efficacy of a landscape scale feral cat control program using camera traps and occupancy models. Scientific Reports 8, 5335.
| Evaluating the efficacy of a landscape scale feral cat control program using camera traps and occupancy models.Crossref | GoogleScholarGoogle Scholar | 29593271PubMed |
Descalzo, E, Jiménez, J, Delibes-Mateos, M, Díaz-Ruiz, F, and Ferreras, P (2021). Assessment of methods for detecting an opportunistic and expanding mesocarnivore in southwestern Europe. Journal of Zoology 315, 138–148.
| Assessment of methods for detecting an opportunistic and expanding mesocarnivore in southwestern Europe.Crossref | GoogleScholarGoogle Scholar |
de Tores PJ, Marlow N (2012) The relative merits of predator-exclusion fencing and repeated fox baiting for protection of native fauna: five case studies from Western Australia. In ‘Fencing for Conservation: Restriction of Evolutionary Potential or a Riposte to Threatening Processes?’. (Eds MJ Somers, M Hayward) pp. 21–42. (Springer New York: New York, NY, USA)
Dexter, N, and Murray, A (2009). The impact of fox control on the relative abundance of forest mammals in East Gippsland, Victoria. Wildlife Research 36, 252–261.
| The impact of fox control on the relative abundance of forest mammals in East Gippsland, Victoria.Crossref | GoogleScholarGoogle Scholar |
Doherty, TS, Glen, AS, Nimmo, DG, Ritchie, EG, and Dickman, CR (2016). Invasive predators and global biodiversity loss. Proceedings of the National Academy of Sciences of the United States of America 113, 11261–11265.
| Invasive predators and global biodiversity loss.Crossref | GoogleScholarGoogle Scholar | 27638204PubMed |
Doody, JS, Soanes, R, Castellano, CM, Rhind, D, Green, B, McHenry, CR, and Clulow, S (2015). Invasive toads shift predator–prey densities in animal communities by removing top predators. Ecology 96, 2544–2554.
| Invasive toads shift predator–prey densities in animal communities by removing top predators.Crossref | GoogleScholarGoogle Scholar | 26594710PubMed |
du Preez, BD, Loveridge, AJ, and Macdonald, DW (2014). To bait or not to bait: a comparison of camera-trapping methods for estimating leopard Panthera pardus density. Biological Conservation 176, 153–161.
| To bait or not to bait: a comparison of camera-trapping methods for estimating leopard Panthera pardus density.Crossref | GoogleScholarGoogle Scholar |
Einoder, LD, Southwell, DM, Lahoz-Monfort, JJ, Gillespie, GR, Fisher, A, and Wintle, BA (2018). Occupancy and detectability modelling of vertebrates in northern Australia using multiple sampling methods. PLoS ONE 13, e0203304.
| Occupancy and detectability modelling of vertebrates in northern Australia using multiple sampling methods.Crossref | GoogleScholarGoogle Scholar | 30248104PubMed |
Feit, B, Dempster, T, Jessop, TS, Webb, JK, and Letnic, M (2020). A trophic cascade initiated by an invasive vertebrate alters the structure of native reptile communities. Global Change Biology 26, 2829–2840.
| A trophic cascade initiated by an invasive vertebrate alters the structure of native reptile communities.Crossref | GoogleScholarGoogle Scholar | 32034982PubMed |
Glen, AS, and Dickman, CR (2008). Niche overlap between marsupial and eutherian carnivores: does competition threaten the endangered spotted-tailed quoll? Journal of Applied Ecology 45, 700–707.
| Niche overlap between marsupial and eutherian carnivores: does competition threaten the endangered spotted-tailed quoll?Crossref | GoogleScholarGoogle Scholar |
Glen, AS, Pech, RP, and Byrom, AE (2013). Connectivity and invasive species management: towards an integrated landscape approach. Biological Invasions 15, 2127–2138.
| Connectivity and invasive species management: towards an integrated landscape approach.Crossref | GoogleScholarGoogle Scholar |
Griffiths, AD, and McKay, JL (2007). Cane toads reduce the abundance and site occupancy of Merten’s water monitor (Varanus mertensi). Wildlife Research 34, 609–615.
| Cane toads reduce the abundance and site occupancy of Merten’s water monitor (Varanus mertensi).Crossref | GoogleScholarGoogle Scholar |
Guarino, F (2002). Spatial ecology of a large carnivorous lizard, Varanus varius (Squamata: Varanidae). Journal of Zoology 258, 449–457.
| Spatial ecology of a large carnivorous lizard, Varanus varius (Squamata: Varanidae).Crossref | GoogleScholarGoogle Scholar |
Hayward, MW, Boitani, L, Burrows, ND, Funston, PJ, Karanth, KU, MacKenzie, DI, Pollock, KH, and Yarnell, RW (2015). FORUM: ecologists need robust survey designs, sampling and analytical methods. Journal of Applied Ecology 52, 286–290.
| FORUM: ecologists need robust survey designs, sampling and analytical methods.Crossref | GoogleScholarGoogle Scholar |
Hayward MW, Somers MJ (2012) An introduction to fencing for conservation. In ‘Fencing for Conservation’. (Eds MJ Somers, MW Hayward) pp. 1–6. (Springer: New York, NY, USA)
Hines JE (2006) Program PRESENCE. Available at http://www.mbrpwrc.usgs.gov/software/doc/presence/presence.html
Hradsky, BA, Kelly, LT, Robley, A, and Wintle, BA (2019). FoxNet: an individual-based model framework to support management of an invasive predator, the red fox. Journal of Applied Ecology 56, 1460–1470.
| FoxNet: an individual-based model framework to support management of an invasive predator, the red fox.Crossref | GoogleScholarGoogle Scholar |
Hu, Y, Gillespie, G, and Jessop, TS (2019). Variable reptile responses to introduced predator control in southern Australia. Wildlife Research 46, 64–75.
| Variable reptile responses to introduced predator control in southern Australia.Crossref | GoogleScholarGoogle Scholar |
Imansyah, MJ, Jessop, TS, Ciofi, C, and Akbar, Z (2008). Ontogenetic differences in the spatial ecology of immature Komodo dragons. Journal of Zoology 274, 107–115.
| Ontogenetic differences in the spatial ecology of immature Komodo dragons.Crossref | GoogleScholarGoogle Scholar |
Jessop, TS, Anson, JR, Narayan, E, and Lockwood, T (2015). An introduced competitor elevates corticosterone responses of a native lizard (Varanus varius). Physiological and Biochemical Zoology 88, 237–245.
| An introduced competitor elevates corticosterone responses of a native lizard (Varanus varius).Crossref | GoogleScholarGoogle Scholar | 25860823PubMed |
Jessop, TS, Ariefiandy, A, Forsyth, DM, Purwandana, D, White, CR, Benu, YJ, Madsen, T, Harlow, HJ, and Letnic, M (2020). Komodo dragons are not ecological analogs of apex mammalian predators. Ecology 101, e02970.
| Komodo dragons are not ecological analogs of apex mammalian predators.Crossref | GoogleScholarGoogle Scholar | 31984486PubMed |
Jessop TS, Gillespie GR, Letnic M (2016) Examining multi-scale effects of the invasive fox on a large varanid (Varanus varius White, 1790) mesopredator. In ‘Proceedings of the 2015 Interdisciplinary World Conference on Monitor Lizards’. (Ed. M Cota) pp. 221–236. (Institute for Research and Development, Suan Sunandha Rajabhat University: Thailand)
Jessop, TS, Holmes, B, Sendjojo, A, Thorpe, MO, and Ritchie, EG (2021). Assessing the benefits of integrated introduced predator management for recovery of native predators. Restoration Ecology 29, e13419.
| Assessing the benefits of integrated introduced predator management for recovery of native predators.Crossref | GoogleScholarGoogle Scholar |
Jessop, TS, Kearney, MR, Moore, JL, Lockwood, T, and Johnston, M (2013). Evaluating and predicting risk to a large reptile (Varanus varius) from feral cat baiting protocols. Biological Invasions 15, 1653–1663.
| Evaluating and predicting risk to a large reptile (Varanus varius) from feral cat baiting protocols.Crossref | GoogleScholarGoogle Scholar |
Jessop, TS, Smissen, P, Scheelings, F, and Dempster, T (2012). Demographic and phenotypic effects of human mediated trophic subsidy on a large Australian lizard (Varanus varius): meal ticket or last supper? PLoS ONE 7, e34069.
| Demographic and phenotypic effects of human mediated trophic subsidy on a large Australian lizard (Varanus varius): meal ticket or last supper?Crossref | GoogleScholarGoogle Scholar | 22509271PubMed |
Jessop, TS, Urlus, JA, Lockwood, T, and Gillespie, GR (2010). Preying possum: assessment of the diet of lace monitors (Varanus varius) from coastal forests in southeastern Victoria. Biawak 4, 59–63.
Kéry, M, and Schmidt, BR (2008). Imperfect detection and its consequences for monitoring for conservation. Community Ecology 9, 207–216.
| Imperfect detection and its consequences for monitoring for conservation.Crossref | GoogleScholarGoogle Scholar |
Kinnear, JE, Onus, ML, and Sumner, NR (1998). Fox control and rock-wallaby population dynamics — II. An update. Wildlife Research 25, 81–88.
| Fox control and rock-wallaby population dynamics — II. An update.Crossref | GoogleScholarGoogle Scholar |
Kinnear, JE, Sumner, NR, and Onus, ML (2002). The red fox in Australia—an exotic predator turned biocontrol agent. Biological Conservation 108, 335–359.
| The red fox in Australia—an exotic predator turned biocontrol agent.Crossref | GoogleScholarGoogle Scholar |
Legge, S, Woinarski, JCZ, Burbidge, AA, Palmer, R, Ringma, J, Radford, JQ, Mitchell, N, Bode, M, Wintle, B, Baseler, M, Bentley, J, Copley, P, Dexter, N, Dickman, CR, Gillespie, GR, Hill, B, Johnson, CN, Latch, P, Letnic, M, Manning, A, McCreless, EE, Menkhorst, P, Morris, K, Moseby, K, Page, M, Pannell, D, and Tuft, K (2018). Havens for threatened Australian mammals: the contributions of fenced areas and offshore islands to the protection of mammal species susceptible to introduced predators. Wildlife Research 45, 627–644.
| Havens for threatened Australian mammals: the contributions of fenced areas and offshore islands to the protection of mammal species susceptible to introduced predators.Crossref | GoogleScholarGoogle Scholar |
Lindenmayer, D, Woinarski, J, Legge, S, Southwell, D, Lavery, T, Robinson, N, Scheele, B, and Wintle, B (2020). A checklist of attributes for effective monitoring of threatened species and threatened ecosystems. Journal of Environmental Management 262, 110312.
| A checklist of attributes for effective monitoring of threatened species and threatened ecosystems.Crossref | GoogleScholarGoogle Scholar | 32250795PubMed |
Lindenmayer, DB, and Likens, GE (2010). The science and application of ecological monitoring. Biological Conservation 143, 1317–1328.
| The science and application of ecological monitoring.Crossref | GoogleScholarGoogle Scholar |
Lindenmayer, DB, Wood, J, MacGregor, C, Foster, C, Scheele, B, Tulloch, A, Barton, P, Banks, S, Robinson, N, Dexter, N, O’Loughlin, LS, and Legge, S (2018). Conservation conundrums and the challenges of managing unexplained declines of multiple species. Biological Conservation 221, 279–292.
| Conservation conundrums and the challenges of managing unexplained declines of multiple species.Crossref | GoogleScholarGoogle Scholar |
Long, RA, Donovan, TM, Mackay, P, Zielinski, WJ, and Buzas, JS (2007). Comparing scat detection dogs, cameras, and hair snares for surveying carnivores. Journal of Wildlife Management 71, 2018–2025.
| Comparing scat detection dogs, cameras, and hair snares for surveying carnivores.Crossref | GoogleScholarGoogle Scholar |
Long RA, MacKay P, Ray J, Zielinski W (2012) ‘Noninvasive Survey Methods for Carnivores.’ (Island Press: Washington, DC, USA)
Lunn, DJ, Thomas, A, Best, N, and Spiegelhalter, D (2000). WinBUGS – a Bayesian modelling framework: concepts, structure, and extensibility. Statistics and Computing 10, 325–337.
| WinBUGS – a Bayesian modelling framework: concepts, structure, and extensibility.Crossref | GoogleScholarGoogle Scholar |
MacKenzie DI, Nichols JD, Royle JA, Pollock KH, Bailey L, Hines JE (2017) ‘Occupancy Estimation and Modeling: Inferring Patterns and Dynamics of Species Occurrence.’ (Elsevier: Oxford, UK)
MacKenzie D, Nichols J, Royle J, Pollock K, Bailey L, Hines J (2006) ‘Occupancy Estimation and Modelling.’ (Academic Press: Burlington, MA, USA)
Marlow, NJ, Thomas, ND, Williams, AAE, Macmahon, B, Lawson, J, Hitchen, Y, Angus, J, and Berry, O (2015). Cats (Felis catus) are more abundant and are the dominant predator of woylies (Bettongia penicillata) after sustained fox (Vulpes vulpes) control. Australian Journal of Zoology 63, 18–27.
| Cats (Felis catus) are more abundant and are the dominant predator of woylies (Bettongia penicillata) after sustained fox (Vulpes vulpes) control.Crossref | GoogleScholarGoogle Scholar |
Martin, J, McIntyre, CL, Hines, JE, Nichols, JD, Schmutz, JA, and MacCluskie, MC (2009). Dynamic multistate site occupancy models to evaluate hypotheses relevant to conservation of Golden Eagles in Denali National Park, Alaska. Biological Conservation 142, 2726–2731.
| Dynamic multistate site occupancy models to evaluate hypotheses relevant to conservation of Golden Eagles in Denali National Park, Alaska.Crossref | GoogleScholarGoogle Scholar |
Mattfeldt, SD, Bailey, LL, and Grant, EHC (2009). Monitoring multiple species: estimating state variables and exploring the efficacy of a monitoring program. Biological Conservation 142, 720–737.
| Monitoring multiple species: estimating state variables and exploring the efficacy of a monitoring program.Crossref | GoogleScholarGoogle Scholar |
McCarthy MA (2007) ‘Bayesian Methods for Ecology’. (Cambridge University Press: Cambridge, UK)
McGrath, T, Guillera-Arroita, G, Lahoz-Monfort, JJ, Osborne, W, Hunter, D, and Sarre, SD (2015). Accounting for detectability when surveying for rare or declining reptiles: turning rocks to find the Grassland Earless Dragon in Australia. Biological Conservation 182, 53–62.
| Accounting for detectability when surveying for rare or declining reptiles: turning rocks to find the Grassland Earless Dragon in Australia.Crossref | GoogleScholarGoogle Scholar |
Mcilroy, JC, King, DR, and Oliver, AJ (1985). The sensitivity of Australian animals to 1080 poison VIII.* Amphibians and reptiles. Wildlife Research 12, 113–118.
| The sensitivity of Australian animals to 1080 poison VIII.* Amphibians and reptiles.Crossref | GoogleScholarGoogle Scholar |
Michael, DR, Cunningham, RB, Donnelly, CF, and Lindenmayer, DB (2012). Comparative use of active searches and artificial refuges to survey reptiles in temperate eucalypt woodlands. Wildlife Research 39, 149–162.
| Comparative use of active searches and artificial refuges to survey reptiles in temperate eucalypt woodlands.Crossref | GoogleScholarGoogle Scholar |
Moll, RJ, Redilla, KM, Mudumba, T, Muneza, AB, Gray, SM, Abade, L, Hayward, MW, Millspaugh, JJ, and Montgomery, RA (2017). The many faces of fear: a synthesis of the methodological variation in characterizing predation risk. Journal of Animal Ecology 86, 749–765.
| The many faces of fear: a synthesis of the methodological variation in characterizing predation risk.Crossref | GoogleScholarGoogle Scholar | 28390066PubMed |
Moore, HA, Champney, JL, Dunlop, JA, Valentine, LE, and Nimmo, DG (2020). Spot on: using camera traps to individually monitor one of the world’s largest lizards. Wildlife Research 47, 326–337.
| Spot on: using camera traps to individually monitor one of the world’s largest lizards.Crossref | GoogleScholarGoogle Scholar |
Murray AJ, Poore RN, Dexter N (2006) Project Deliverance – the response of “critical weight range” mammals to effective fox control in mesic forest habitats in far East Gippsland, Victoria. Victorian Department of Sustainability and Environment, Melbourne, Vic., Australia.
Nichols, JD, Bailey, LL, O’Connell, AF, Talancy, NW, Campbell Grant, EH, Gilbert, AT, Annand, EM, Husband, TP, and Hines, JE (2008). Multi-scale occupancy estimation and modelling using multiple detection methods. Journal of Applied Ecology 45, 1321–1329.
| Multi-scale occupancy estimation and modelling using multiple detection methods.Crossref | GoogleScholarGoogle Scholar |
Nichols, JD, Hines, JE, Mackenzie, DI, Seamans, ME, and Gutiérrez, RJ (2007). Occupancy estimation and modeling with multiple states and state uncertainty. Ecology 88, 1395–1400.
| Occupancy estimation and modeling with multiple states and state uncertainty.Crossref | GoogleScholarGoogle Scholar | 17601132PubMed |
Nichols, JD, and Williams, BK (2006). Monitoring for conservation. Trends in Ecology & Evolution 21, 668–673.
| Monitoring for conservation.Crossref | GoogleScholarGoogle Scholar |
Otto, CRV, and Roloff, GJ (2011). Using multiple methods to assess detection probabilities of forest-floor wildlife. The Journal of Wildlife Management 75, 423–431.
| Using multiple methods to assess detection probabilities of forest-floor wildlife.Crossref | GoogleScholarGoogle Scholar |
Phillips, RB, and Winchell, CS (2011). Reducing nontarget recaptures of an endangered predator using conditioned aversion and reward removal. Journal of Applied Ecology 48, 1501–1507.
| Reducing nontarget recaptures of an endangered predator using conditioned aversion and reward removal.Crossref | GoogleScholarGoogle Scholar |
Purwandana, D, Ariefiandy, A, Imansyah, MJ, Rudiharto, H, Seno, A, Ciofi, C, Fordham, DA, and Jessop, TS (2014). Demographic status of Komodo dragons populations in Komodo National Park. Biological Conservation 171, 29–35.
| Demographic status of Komodo dragons populations in Komodo National Park.Crossref | GoogleScholarGoogle Scholar |
Purwandana, D, Ciofi, C, Imansyah, MJ, Ariefiandy, A, Rudiharto, H, and Jessop, TS (2021). Prey preferences and body mass most influence movement behavior and home range area of Komodo Dragons. Ichthyology & Herpetology 109, 92–101.
| Prey preferences and body mass most influence movement behavior and home range area of Komodo Dragons.Crossref | GoogleScholarGoogle Scholar |
Radford, JQ, Woinarski, JC, Legge, S, Baseler, M, Bentley, J, Burbidge, AA, Bode, M, Copley, P, Dexter, N, Dickman, CR, Gillespie, G, Hill, B, Johnson, CN, Kanowski, J, Latch, P, Letnic, M, Manning, A, Menkhorst, P, Mitchell, N, Morris, K, Moseby, K, Page, M, and Ringma, J (2018). Degrees of population-level susceptibility of Australian terrestrial non-volant mammal species to predation by the introduced red fox (Vulpes vulpes) and feral cat (Felis catus). Wildlife Research 45, 645–657.
| Degrees of population-level susceptibility of Australian terrestrial non-volant mammal species to predation by the introduced red fox (Vulpes vulpes) and feral cat (Felis catus).Crossref | GoogleScholarGoogle Scholar |
Read, JL, and Scoleri, V (2015). Ecological implications of reptile mesopredator release in arid South Australia. Journal of Herpetology 49, 64–69.
| Ecological implications of reptile mesopredator release in arid South Australia.Crossref | GoogleScholarGoogle Scholar |
Ritchie, EG, and Johnson, CN (2009). Predator interactions, mesopredator release and biodiversity conservation. Ecology Letters 12, 982–998.
| Predator interactions, mesopredator release and biodiversity conservation.Crossref | GoogleScholarGoogle Scholar | 19614756PubMed |
Saunders, GR, Gentle, MN, and Dickman, CR (2010). The impacts and management of foxes Vulpes vulpes in Australia. Mammal Review 40, 181–211.
| The impacts and management of foxes Vulpes vulpes in Australia.Crossref | GoogleScholarGoogle Scholar |
Smissen, PJ, Melville, J, Sumner, J, and Jessop, TS (2013). Mountain barriers and river conduits: phylogeographical structure in a large, mobile lizard (Varanidae: Varanus varius) from eastern Australia. Journal of Biogeography 40, 1729–1740.
| Mountain barriers and river conduits: phylogeographical structure in a large, mobile lizard (Varanidae: Varanus varius) from eastern Australia.Crossref | GoogleScholarGoogle Scholar |
Stewart, FEC, Volpe, JP, and Fisher, JT (2019). The debate about bait: a red herring in wildlife research. The Journal of Wildlife Management 83, 985–992.
| The debate about bait: a red herring in wildlife research.Crossref | GoogleScholarGoogle Scholar |
Stokeld, D, Frank, ASK, Hill, B, Choy, JL, Mahney, T, Stevens, A, Young, S, Rangers, D, Rangers, W, and Gillespie, GR (2015). Multiple cameras required to reliably detect feral cats in northern Australian tropical savanna: an evaluation of sampling design when using camera traps. Wildlife Research 42, 642–649.
| Multiple cameras required to reliably detect feral cats in northern Australian tropical savanna: an evaluation of sampling design when using camera traps.Crossref | GoogleScholarGoogle Scholar |
Thompson, GG, and Thompson, SA (2007). Usefulness of funnel traps in catching small reptiles and mammals, with comments on the effectiveness of the alternatives. Wildlife Research 34, 491–497.
| Usefulness of funnel traps in catching small reptiles and mammals, with comments on the effectiveness of the alternatives.Crossref | GoogleScholarGoogle Scholar |
Thompson W (2013) ‘Sampling Rare or Elusive Species: Concepts, Designs, and Techniques for Estimating Population Parameters.’ (Island Press: Washington, DC, USA)
Twigg, LE, and King, DR (1991). The impact of fluoroacetate-bearing vegetation on native Australian fauna: a review. Oikos 61, 412–430.
| The impact of fluoroacetate-bearing vegetation on native Australian fauna: a review.Crossref | GoogleScholarGoogle Scholar |
Walsh, JC, Wilson, KA, Benshemesh, J, and Possingham, HP (2012). Unexpected outcomes of invasive predator control: the importance of evaluating conservation management actions. Animal Conservation 15, 319–328.
| Unexpected outcomes of invasive predator control: the importance of evaluating conservation management actions.Crossref | GoogleScholarGoogle Scholar |
Willson, JD, Winne, CT, and Todd, BD (2011). Ecological and methodological factors affecting detectability and population estimation in elusive species. The Journal of Wildlife Management 75, 36–45.
| Ecological and methodological factors affecting detectability and population estimation in elusive species.Crossref | GoogleScholarGoogle Scholar |
Woinarski, JCZ, Braby, MF, Burbidge, AA, Coates, D, Garnett, ST, Fensham, RJ, Legge, SM, McKenzie, NL, Silcock, JL, and Murphy, BP (2019). Reading the black book: the number, timing, distribution and causes of listed extinctions in Australia. Biological Conservation 239, 108261.
| Reading the black book: the number, timing, distribution and causes of listed extinctions in Australia.Crossref | GoogleScholarGoogle Scholar |
Yoccoz, NG, Nichols, JD, and Boulinier, T (2001). Monitoring of biological diversity in space and time. Trends in Ecology & Evolution 16, 446–453.
| Monitoring of biological diversity in space and time.Crossref | GoogleScholarGoogle Scholar |