

A taxonomic review of the austral genera Asterochaete and Carpha (Cyperaceae, Carpheae)
Jeremy J. Bruhl A # * , Karen L. Wilson B # and Xiufu Zhang A C #
A # * , Karen L. Wilson B # and Xiufu Zhang A C #
A
B
C Present address:
Handling Editor: Daniel Murphy
Abstract
Tribe Carpheae presently includes Carpha and Trianoptiles. Sixteen species were recovered in Carpha Banks & Sol. ex R.Br. sens. lat. by phenetic analyses and these species were separated into two clades that we recognise here as genera Carpha sens. str. and Asterochaete Nees, on the basis of phylogenetic analyses. We present a new taxonomy for the two genera, including eleven species in Asterochaete (A. acuminata X.F.Zhang, A. angustissima (Cherm.) X.F.Zhang, A. borbonica (Steud.) X.F.Zhang, A. capitellata Nees, A. dactyloides (Vahl) K.L.Wilson & J.J.Bruhl, A. discolor T.H.Arnold ex X.F.Zhang, A. eminii (C.B.Clarke) X.F.Zhang, A. filifolia (C.Reid & T.H.Arnold) X.F.Zhang, A. nitens Kunth, A. schlechteri (C.B.Clarke) X.F.Zhang and A. ulugurensis Nelmes ex X.F.Zhang) and five species in Carpha sens. str. (C. alpina R.Br., C. curvata W.M.Curtis, C. nivicola F.Muell., C. rodwayi W.M.Curtis, and C. schoenoides Banks & Sol. ex Hook.f.), with one unassigned name (Carpha aristata Kük.). Asterochaete acuminata, A. discolor and A. ulugurensis are new species; a new combination, A. dactyloides, is made for what was previously called A. glomerata (Nees) Nees. The ranks of Asterochaete angustissima, Carpha schoenoides and C. nivicola are clarified, whereas Carpha bracteosa C.B.Clarke and C. perrieri Cherm. are synonymised with A. capitellata.
Keywords: Africa, Asterochaete, Australia, Carpha, New Guinea, new species, New Zealand, South America, systematics, taxonomy.
Introduction
Carpha Banks & Sol. ex R.Br. was formally described by Brown (1810, p. 230), who took the generic name from a manuscript name of Banks and Solander and included five Australian species in the genus. Since then, four of Brown’s five species have been moved from Carpha and over forty names have been applied to species in this genus in a broad or narrow sense or are now included in other genera (Zhang et al. 2006).
The genus Asterochaete Nees included three southern African species Nees (1834), namely, Asterochaete glomerata Nees, A. angustifolia Nees (later treated as a synonym of Carpha capitellata (Nees) Boeckeler by Boeckeler (1874)) and A. capitellata Nees. However, Nees’ first specific name was illegitimate because the name Schoenus dactyloides Vahl (1805) should have been adopted for that taxon (which was not realised until recently; see discussion below under A. dactyloides (Vahl) K.L.Wilson & J.J.Bruhl), and the other two names were nomina nuda. The following year, Nees (1835) validly published A. capitellata Nees. As detailed in Zhang et al. (2006), subsequent authors have treated Carpha either in a strict sense or in a broader sense including Asterochaete. The only other generic name relevant here is the monotypic African genus Oreograstis K.Schum. (O. eminii K.Schum.), which was moved to the synonymy of Carpha sens. lat. by Clarke (1902). Kükenthal (1939a) also included Trianoptiles Fenzl in Carpha as a separate subgenus, and recent molecular studies have indicate that these taxa are closely related (Zhang et al. 2007). Levyns (1943, 1947) considered that Asterochaete, Carpha and Trianoptiles were separate genera with different floral structures.
Our phenetic analysis of species limits in Carpha sens. lat. (Zhang 2002; Zhang et al. 2006) recovered the following 16 species: C. alpina R.Br., C. curvata W.M.Curtis, C. nivicola F.Muell., C. rodwayi W.M.Curtis, C. schoenoides Banks & Sol. ex Hook.f., C. angustissima Cherm., C. borbonica (Steud.) C.B.Clarke, C. capitellata, C. eminii (K.Schum.) C.B.Clarke, C. filifolia C.Reid & T.H.Arnold, C. glomerata (Thunb.) Nees, C. nitens (Kunth) Kük., C. schlechteri C.B.Clarke, and three new species (given there as C. cf. bracteosa, C. discolor ms and C. ulugurensis ms). Our phylogenetic analyses of Carpha and its relatives using morphological (sampling 16 species of Carpha sens. lat.) and combined morphological and molecular data (sampling eight species of Carpha sens. lat.; Zhang 2002; Zhang et al. 2004a, 2007) placed the 16 species of Carpha in two clades. One clade was composed of all species of Carpha sens. str. (Hooker 1860, 1867; Bentham 1878, 1883; Wilson 1986, 1993, 1994a, 1994b). The other clade included the rest of the species of Carpha sens. lat., which corresponds to the definition of Asterochaete (Nees 1834, 1835; Kunth 1837; Steudel 1855; Levyns 1950). The two clades also appeared in cladistic analyses based on DNA sequence data alone (Zhang 2002; Zhang et al. 2004a, 2007), suggesting that they are two different groups but with a close relationship.
More recent studies using several different molecular regions (Verboom 2006; Muasya et al. 2009; Hinchliff and Roalson 2013; Jung and Choi 2013; Viljoen et al. 2013; Semmouri et al. 2019) have suggested that (a) Carpha sens. str. and Asterochaete form a clade, (b) Trianoptiles is either in the same clade or sister to it, and (c) (Carpha sens. lat. + Trianoptiles) is sister to the rest of tribe Schoeneae or sister to a much larger clade of Cyperaceae. However, in most of these studies, the sample size was small, generally including only one or two species of each putative genus in each study. Zhang et al. (2004a, 2007) included nine putative species of Carpha sens. lat., whereas Semmouri et al. (2019) included a total of five species of Carpha sens. lat. Tribe Carpheae Semmouri & Larridon was established by Semmouri et al. (2019) for Carpha sens. lat. and Trianoptiles, on the basis of molecular data and having a distinctive type of embryo.
Revising the small southern African genus Trianoptiles was not part of our study, but it is clear that it belongs in tribe Carpheae (Zhang et al. 2007; Semmouri et al. 2019; Larridon et al. 2021). We recognise both Asterochaete and Carpha sens. str. (hereafter Carpha), given the results of our phenetic and combined morphological and molecular phylogenetic analyses (Zhang 2002; Zhang et al. 2004a, 2006, 2007), together with the distinct morphological features for both genera. This paper presents a taxonomic treatment of the two genera and their species, with keys to allow identification of plants to genus and to species.
Materials and methods
Plant material
Our study was based on field observations and on specimens collected by us, and on herbarium specimens in the following herbaria: AAU, AK, B, BM, BOL, BR, BRI, C, CANB, CGE, CHR, E, EA, F, FI, G, HBG, HO, K, L, M, MEL, MO, NE, NSW, NU, NY, P, PRE, UPS, US and Z. Specimens cited have been examined unless otherwise indicated by ‘n.v.’ Specimens seen only as high-resolution images on-line, including those at http://plants.jstor.org created in the JSTOR Global Plants project are indicated by an asterisk (‘*’) and superscript hash (‘#’) for images seen on the Global Biodiversity Information Facility portal.
Some species of Carpha (e.g. C. alpina and C. nivicola) are superficially similar and closely sympatric (see descriptions and discussion below). Care is needed to avoid making mixed collections. Details of the character set used in the phenetic analyses and for describing the species in this paper are given in appendix 2 of Zhang et al. (2006). Some extra characters not used in the phenetic analysis have been added here, for example, more details about the perianth bristles.
Light microscopy
Light micrographs of nutlets were taken using a Nikon SMZ25 motorised stereomicroscope with an SHR Plan Apo 1x lens and Nikon DS-Ri2 camera. Images were acquired using NIS-Element AR software by using Z-stack acquisition under bright-field illumination.
Maps
Relevant data were retrieved from the Global Biodiversity Information Facility (https://www.gbif.org/, accessed 18 February 2018), and a few more recent specimens were added subsequently. Doubtful records were removed, as were duplicate and redundant records for the scale of the map. Maps were produced using SimpleMappr (ver. 8 May 2018, D. P. Shorthouse, see https://www.simplemappr.net).
Conservation status
A few species are already listed by IUCN. For other species, we suggest a status based on information about the species and using the IUCN criteria (IUCN Standards and Petitions Committee 2022).
Morphological features and notes for using keys
Culm diameter was measured in the middle third of the culm length. Leaf-blade width was measured in the middle third of several of the longest leaves on each specimen. Inflorescence length was measured from the lowest (proximal) inflorescence node and involucral bract to the top of the uppermost spikelet(s), i.e. ignoring any longer involucral bracts. Leaf and involucral bract measurements and shapes should be examined at around midpoint of length. The uppermost basal leaf on a culm should be examined for the shape of the sheath apex opposite the junction with the blade (lower sheaths are usually split to the base). The apex of the sheath of the proximal (or lowest) involucral bract is usually similar to that of the uppermost leaf.
Presence and size of rhizomes are useful in distinguishing some species in Asterochaete, but they are often absent from perennial rhizomatous specimens, either because they have not been collected or because the specimens are immature.
Leaves are pseudopetiolate (not always conspicuously so) in species of Carpha and obscurely so in species of Asterochaete. As discussed in Bruhl (1995) and Barrett et al. (2019), further investigation is needed to understand this rather unusual morphology found often in tribe Schoeneae.
Leaf sheath colour is described from dried material, i.e. after any green colouration has disappeared, and the leaf blades appear straw-coloured (pale yellow-brown). Many species are more brightly coloured towards the base of the sheath, ranging from medium to dark yellow-brown to dark red-brown.
Spikelet structure varies, as discussed in Zhang et al. (2004b). A reference to relevant figures from that paper, showing the type of spikelet structure found in each species, is added here at the end of the corresponding species’ description. The term ‘glumes’ is used here for fertile and sterile floral bracts (cf. Bruhl 1995) of spikelets as is common usage in Cyperaceae (e.g. Haines and Lye 1983; Wilson 1993; Semmouri et al. 2019; Larridon et al. 2021), although these are not homologous with the glumes of spikelets in Poaceae or with the subtending bracts or prophylls in Cyperaceae (see Bruhl 1995; Angiosperm Phylogeny Website, see https://www.mobot.org/MOBOT/research/APweb/).
Perianth bristle morphology can be assessed in specimens with moderately immature through to mature fruits. The two whorls of bristles can be distinguished by one or more of the following features: the outer whorl is attached lower than the inner whorl; the bases of the outer bristles may overlap the inner; the base of the outer bristles in both genera is usually broader than those of the inner whorl. The upper part of bristles may abscise (notably in A. dactyloides (Vahl) K.L.Wilson & J.J.Bruhl and A. schlechteri (C.B.Clarke) X.F.Zhang), so one should avoid measuring partial bristles when assessing bristle length.
‘Nutlet body’ refers to the main body of the fruit, excluding the persistent stylebase and the gynophore in Carpha. By contrast, nutlet length is measured from the base of the perianth in Asterochaete, because this is usually firmly attached to the fruit and difficult to remove, but excludes the stylebase.
Results and discussion
Morphological characters
We investigated morphological features in detail, and found some of significance at generic level, others at specific level.
At the junction of the leaf sheath and blade, there is no ligule in any species of Asterochaete or Carpha. On the face opposite the attachment of the blade (the ‘sheath front’), the apex of the sheath is not horizontal but has a short vertical gap that is either U-shaped (with more or less parallel sides and a horizontal base to the gap) or V-shaped (with the gap narrowing to a point at the base), with the lateral margins either overlapping near the base or not overlapping. Shape of sheath front apex is best assessed on the uppermost leaves (or, as a proxy, on primary involucral bracts) because the sheath is thin-textured in that region and basal leaf sheaths eventually split much further as the plant matures and expands.
The shape of the sheath front apex in A. schlechteri could be interpreted as either U- or V-shaped in the junction region, but most other species of Asterochaete clearly are U-shaped. Asterochaete dactyloides differs from other species in usually having the base of the ‘U’ slightly thickened and rigid, making the opening resistant to splitting. In A. eminii (C.B.Clarke) X.F.Zhang, the base of the opening is not horizontal but vertically convex like a very short contraligule. V-shaped sheath front apices are found in A. acuminata X.F.Zhang and A. filifolia (C.Reid & T.H.Arnold) X.F.Zhang and occasionally in A. borbonica (Steud.) X.F.Zhang and A. nitens Kunth; in the latter three species the lateral margins may be overlapping, so hiding the base of the opening.
In Carpha, the sheath front apex is often hard to see because the leaves are densely packed at the base of the small plants but are easier to see in the primary involucral bracts. All of these species are shallowly and broadly U-shaped at the sheath apex except for C. schoenoides, which is broadly V-shaped. In C. alpina, the base of the ‘U’ is either horizontal as in the other U-shaped species or is vertically convex like a very short contraligule, the latter being similar to A. eminii.
Bruhl (1995, p. 172) stated that ‘yellow-green anthers are a feature shared by Carpha and Trianoptiles’. Semmouri and Larridon placed considerable importance on anther colour in the diagnosis of their new tribe, Carpheae (Semmouri et al. 2019, p. 33). We concur that anther colour appears to be distinctive for the tribe, although we have observed more variation. In most species of Asterochaete, anthers are pale grey to blackish. An exception to this is A. discolor T.H.Arnold ex X.F.Zhang, which has anthers pale yellowish when immature, becoming slightly greyish when mature. Asterochaete dactyloides has anthers that are pale yellow when immature but pale grey at maturity, and A. schlechteri has anthers that remain pale yellow. In Carpha, anthers range in colour from pale yellow to pale or dark grey.
Anther appendages were not found to be as useful in distinguishing species as Reid and Arnold (1984) suggested for the southern African species. The apical appendage is no more than 0.3 mm long in most species of both genera, only exceeding that length in A. discolor (0.3‒0.8 mm long), A. schlechteri (0.3‒0.5 mm long) and C. rodwayi (0.1‒0.4 mm long). However, we found other distinctions in features of the perianth and fruits (nutlets). The perianth in Asterochaete and Carpha is composed of bristles (Bruhl 1991, 1995), often referred to as hypogynous bristles (e.g. Wilson 1993).
In species of Asterochaete, the three bristles of the inner whorl are usually obviously longer than those of the outer whorl (occasionally subequal in A. borbonica and C. capitellata) (Table 1). All bristles are antrorsely scabrous for their whole length or, near the base, have slightly longer hairs or are glabrous (Fig. 1). The bristles of A. discolor differ in being shortly plumose for up to two-thirds of their length, but those hairs are shorter (0.1–0.5 mm long) than the hairs that make up the plumose bristles in Carpha. In species of Asterochaete, each bristle consists of an upper slender, terete part, a proximal part often still slender but somewhat flattened and broadened, and usually (but variously) a flared base. The proportions and distinctness of each part vary more among than within species. The bristles in A. dactyloides and A. schlechteri differ from all other species in having the basal 1/3–3/5 of their length glabrous and slightly inflated, separated by an obvious abscission point from the scabrous apical section. Frequently, only the glabrous basal section persists in specimens that we have seen.
Nutlets with perianth of perianth bristles in Asterochaete spp.: (a) A. acuminata (B. Sonnenberg 336, NU); (b) A. angustissima (H.U. Stauffer 793, K); (c) A. borbonica (J.J. Bruhl 2977 et al., NE); (d) A. capitellata (J.J. Bruhl 1725, NE); (e) A. capitellata (J.J. Bruhl 1726 & F.C. Quinn, NE); (f) A. dactyloides (B. Sonnenberg 387, NU); (g) A. discolor (E. Esterhuysen 14866, K); (h) A. eminii (R.W. Haines 277, K); (i) A. filifolia (O.M. Hilliard & B.L. Burt 16258, K); (j) A. nitens (J.J. Bruhl 2949 et al., NE); (k) A. schlechteri (J. Browning 823, NU); (l) A. ulugurensis (E.M. Bruce 742, K). a, articulation point or narrowed portion of perianth bristle (top of bristle fallen in a, f and in g where the thin portion of a bristle is seen to lie horizontally in front of stylebase); f, staminal filament (flat, linear and glabrous, in contrast to bristles); s, stylebase above body of nutlet. Scale bars: 0.5 mm. Images by Jeremy J. Bruhl.

| Species | Shape | Surface of bristles (‘Outer’ = bristles in outer whorl; ‘Inner’ = bristles in inner whorl) | Length of hairs (mm) | Length of outer whorl of bristles relative to inner whorl | Length relative to body of nutlet (‘Outer’ = bristles in outer whorl; ‘Inner’ = bristles in inner whorl) | Length of longest bristles (mm) | |
|---|---|---|---|---|---|---|---|
| Asterochaete acuminata | Terete; broadened at base | Antrorsely scabrous (evenly and densely so, especially distally) | <0.05 | Shorter | Outer and inner shorter than nutlet (2/3‒4/5 length), rarely slightly exceeding nutlet | 3.0‒3.7 | |
| A. angustissima | Upper half to two-thirds terete to filiform (and thinner textured); proximally slightly compressed, broadened at base | Distally evenly & shortly antrorsely scabrous; lower half ±glabrous to antrorsely scabrous, with sparse to moderately dense prickle hairs (usually twice as long towards base) | 0.03–0.09 | Shorter (~1/4‒1/2 length) | Outer much shorter than nutlet (~1/3–1/2 as long); inner equalling or exceeding nutlet (to 1.5× as long) | (2.0‒)3‒4.4 | |
| A. borbonica | ±Terete; slightly compressed above the flared base | Antrorsely scabrous | 0.03 | Shorter (rarely subequal) | Outer shorter than nutlet (often <1/2 the length); inner slightly shorter to slightly exceeding nutlet | 1.8‒2.5 | |
| A. capitellata | ±Terete; slightly compressed above the flared base (more so in inner whorl) | Antrorsely scabrous; prickle hairs shortening occasionally towards base to appear ±glabrous | <0.05 | Shorter (rarely only slightly so) | Outer shorter than to equalling nutlet; inner shorter than nutlet (~1/2 the length) to slightly exceeding nutlet (length varies within a whorl) | 1.5‒3.5 | |
| A. dactyloides | Upper part terete (1.4‒2.0 mm long; 2/5‒2/3 length of bristle; breaking off readily); lower part more slender than upper (1.3‒1.7 mm long; 1/3‒3/5 of bristle), slightly to obviously compressed, gradually broadened proximally with base ±further flared | Antrorsely scabrous above; glabrous below joint | 0.03 | Slightly shorter | Outer and inner slightly shorter than to slightly exceeding nutlet, but often only the glabrous basal part of bristles left on specimen | 3.1‒3.8 | |
| A. discolor A | ±Terete, tapering to a filiform apex; slightly broadened proximally; base slightly flared | Sparsely antrorsely scabrous near apex; lower 1/2–2/3 of bristle with dense, antrorse hairs | 0.1‒0.5 | Shorter | Probably exceeding nutlet (only immature fruits seen) | 9.0‒11.5 | |
| A. eminii | Upper half terete to filiform; lower half compressed and broader, slightly flared at base | Outer: antrorsely scabrous. Inner: upper half antrorsely scabrous; lower half with short antrorse hairs on margins of compressed part | <0.03‒0.2 | Shorter (often much shorter) | Outer equalling or slightly exceeding nutlet; inner ≫ nutlet (to ~twice as long) | 3.5‒6.7 | |
| A. filifolia | Terete, tapering to filiform apex; slightly compressed proximally; slightly flared at base | Outer: antrorsely scabrous, with a few short antrorse hairs at very base. Inner: antrorsely scabrous above; lower part sparsely to densely antrorsely hispid to silky, sometimes only at very base, very rarely for 1/3 length of bristle. Hairs in proximal flattened part abaxial (adaxial surface glabrous) | 0.01–0.2 | Shorter | Outer shorter than to slightly exceeding nutlet; inner slightly exceeding nutlet (to 1.5× as long), rarely slightly shorter than nutlet | (3.0‒)4.0–4. 5(‒7.8) | |
| A. nitens | Terete, tapering to filiform apex; inner bristles more obviously flattened & broader proximally, flared at base | Antrorsely scabrous over whole length; slightly longer, softer, sparse hairs towards base (but not plumose). Hairs mostly on margins on flattened proximal portion mostly on the margins | 0.01–0.1 | Shorter | Outer equalling or exceeding nutlet, occasionally shorter than nutlet; inner exceeding nutlet (~1.2‒2× as long) | 2.8‒5.0 | |
| A. schlechteri | Upper part terete, (1.6‒2.3 mm long; 3/5‒2/3 of length of bristle; but breaking off readily); lower part distally more slender than upper (0.7‒1.3 mm long; 1/3‒2/5 of bristle), flattened, scarious, broadening proximally; base slightly flared | Upper part antrorsely scabrous; lower part glabrous | 0.05 | Shorter | Outer shorter than to equalling nutlet; inner slightly shorter than to slightly exceeding nutlet (length often variable within a whorl); but often only the glabrous basal part of bristles left on specimen | 2.3‒3.4 | |
| A. ulugurensis | Terete to trigonous, tapering to a filiform apex; variable near base: often slightly more slender & glabrous for ~0.5 mm, then slightly broadened below; flared at base | Antrorsely scabrous (less dense than the other species); proximally becoming sparsely hairy to glabrous corresponding to a thinner, translucent portion before the flared base | <0.1 | Shorter | Outer shorter than nutlet (1/2‒3/4 of length); inner from 2/3 as long as to slightly exceeding nutlet (to 1.3× as long) | 2.0‒3.0 | |
| Carpha alpina | Flattened for most of length, tapering to short (0.5‒2.7 mm long) terete to filiform apex | Antrorsely scabrous in apical part; plumose margins on flattened part | <0.1‒2.5 | Subequal | Exceeding nutlet (3‒3.5× as long) | 7.5‒12 | |
| C. curvata | Flattened for most of length, tapering to short (1.6‒2.6 mm long) terete to filiform apex | Antrorsely scabrous in apical part; plumose margins on flattened part | <0.1‒1.3 | Usually subequal (1 bristle longer in Fig. 2b) with outer usually slightly shorter | Exceeding nutlet (2.5‒3× as long) | 7.8‒10.5 mm | |
| C. nivicola | Flattened for most of length, tapering to short (0.8‒2.5 mm long) terete to filiform apex | Antrorsely scabrous in apical part; plumose margins on flattened part | <0.1‒2.1 | Subequal | Exceeding nutlet (3.5‒4× as long) | 13‒19 | |
| C. rodwayi | Flattened for most of length, tapering to short (1.5‒2.7 mm long) terete to filiform apex | Antrorsely scabrous in apical part; plumose margins on flattened part | <0.1‒1.7 | Subequal | Exceeding nutlet (3‒4× as long) | 9.8‒13 | |
| C. schoenoides | Flattened for most of length, tapering to short (0.5‒2.5 mm long) terete to filiform apex | Antrorsely scabrous in apical part; plumose margins on flattened part | 0.1‒1.2 | Subequal | Exceeding nutlet (2.5‒3× as long) | 6.5‒9.8 |
Nutlets (a–e) without and (f–j) with perianth in Carpha spp.: (a, f) C. alpina (J.J. Bruhl 1175 & J.B. Williams, NE); (b, f) C. curvata (J.J. Bruhl 1896b & A.C. Rozefelds, NE); (c, h) C. nivicola (X. Zhang & K.L. Wilson, NE); (d, i) C. rodwayi (J.J. Bruhl 1883c, NE); (e, j) C. schoenoides (A.P. Haase s.n., NE 87966). g, gynophore or stalk of ovary; r, remains of perianth after their partial removal to reveal nutlet with gynophore. Scale bars: 0.5 mm (a–e); 1.0 mm (f–j). Images by Jeremy J. Bruhl.

In all species of Carpha, the six perianth bristles in a flower are more or less all the same length, exceeding the length of the fruit (mostly three to four times as long as the body of the fruit), and are plumose on the margins for most of their length, becoming antrorsely scabrous towards the apex (Fig. 2). The bristles are flattened for most of their length, tapering to a terete or filiform scabrous apex that is 0.5–2.7 mm long. The bristles in C. nivicola are the longest: 13–19 mm long; in the other species they are 6.5–12(–13 in C. rodwayi) mm long. The length of the plumose hairs on the bristles varies along a bristle and within a species, but all are in the range 0.5–2.5 mm long.
The stylebase is persistent on the fruit, generally paler and somewhat thickened and rigid in both genera. In Asterochaete, the stylebase is much shorter than the body of the fruit and is hispidulous in most species (Fig. 1). The stylebase is glabrous in A. ulugurensis Nelmes ex X.F.Zhang and A. discolor (but the only material of that is immature) and is occasionally glabrous in A. dactyloides. In Carpha, the stylebase is longer than to as long as the body of the fruit, and is glabrous or occasionally scattered-hispidulous towards the apex (Fig. 2).
Fruit (nutlet) shape in Asterochaete (Fig. 1) varies from narrow-obovoid (e.g. in A. capitellata) through narrow-ellipsoid (several species) to narrow-ovoid (e.g. A. schlechteri). The fruits of species in Carpha are all narrow-ellipsoid to more or less linear (Fig. 2). In species of Asterochaete, a gynophore, or stalk, at the base of the fruit body is absent or no more than ~0.1 mm long (Fig. 1). In Carpha, the gynophore is 0.2–0.7(–1.0) mm long but is essentially concealed under the persistent perianth bristles (Fig. 2).
All species in both genera have a more or less reticulate pattern on the surface of the nutlet, corresponding to the underlying epidermal cells, as shown for several species in Zhang et al. (2006, fig. 3). Epidermal cell size and lateral wall thickness, as seen in surface view, vary more in Asterochaete than in Carpha. The fruits of species of Carpha have surfaces with very small cell outlines that are less discernible, even at higher magnification with a dissecting microscope, than are those of Asterochaete species (apart from A. angustissima (Cherm.) X.F.Zhang). Asterochaete capitellata has the largest cells with the thickest lateral walls. In Asterochaete, A. dactyloides and A. schlechteri are unusual in having rather inflated epidermal cells so that the outer cell walls look convex (colliculate) and are easily scraped or dislodged from the nutlet. Other species of Asterochaete appear to have outer cell walls that are level with the surface or become sunken into the cell lumen, and the epidermis remains more or less intact. When the outer wall is sunken, the central silica cone on the base of the cell may be prominent, described here as puncticulate in surface view.
Generic limits
Debate over recognition of one or two genera for Asterochaete and Carpha is likely to persist until estimates of their phylogeny are based on a comprehensive sample of species of Carpheae. Meanwhile, the set of morphological putative synapomorphies presented here agrees with some of the inadequately sampled molecular phylogenies. It also allows ready recognition of the two genera and their broad geographical separation and, on balance, provides a reasonable basis and workable taxonomic solution that we adopt here. Future work should focus on molecular phylogenetic analysis of a thorough sampling of the two genera (and Trianoptiles) to test our classification, explore character evolution of features discussed above (e.g. anther and bristle characters), and facilitate fine-scale biogeographic analysis.
Biogeography
The two genera show overall distribution patterns similar to those of numerous other Southern Hemisphere taxa (Fig. 3). Many species occur in isolated high-altitude or high-latitude habitats, often referred to as ‘sky islands’ or ‘habitat islands’ (e.g. Venn et al. 2017; Brochmann et al. 2021), both in the tropics and temperate zones. Asterochaete is found in southern Africa (from near sea level to 1500 m in altitude) and on high mountains of Tropical East Africa, Madagascar and Réunion. Carpha ranges from high-altitude areas in south-eastern mainland Australia and New Guinea to a wider range of altitudes in the higher latitudes of Tasmania and New Zealand and down to close to sea level in Tierra del Fuego, South America. The other member of tribe Carpheae, Trianoptiles, is confined to wetlands of the Western Cape Province of South Africa.
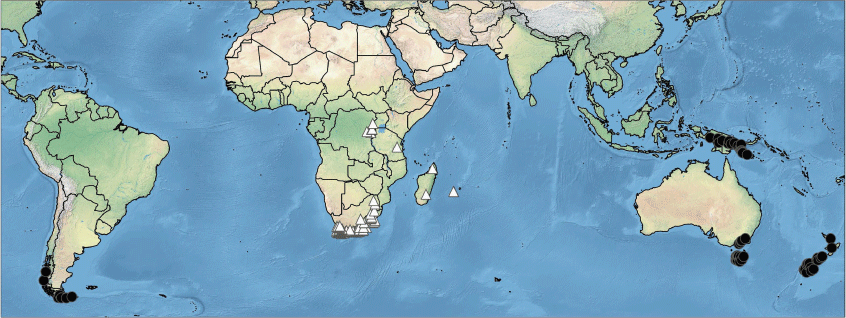
The African species of Asterochaete occur in the fynbos of the Greater Cape Floristic Region, the high-altitude grasslands of the Drakensberg escarpment and Highveld, and a few species occur isolated at high altitude on ‘sky islands’ in Tropical East Africa as well as on a few mountains in Madagascar and Réunion. The fynbos of the Greater Cape Region is suggested to have been evolving throughout the Cenozoic (Verboom et al. 2015; van Santen and Linder 2020). By comparison, Drakensberg and Highveld were more recently uplifted and therefore the high-altitude grassland there is more recent (Gehrke and Linder 2009; Linder and Verboom 2015), as is the tropical Afro-alpine flora of the high mountains of Tropical East Africa, which are mainly volcanic and date from the Miocene or Pliocene (Gehrke and Linder 2014; Brochmann et al. 2021; Kandziora et al. 2022). The Afro-alpine flora has been assessed as being not only relatively young but also of small extent, with limited genetic variation at the population level, suggesting that many species may be vulnerable to changing climate and increasing human impact (Brochmann et al. 2021).
In Australasia (Australia, New Guinea and New Zealand) and the Patagonian region of southern South America, all of the species of Carpha are at high altitude or high latitude. The Australasian alpine regions are all considered to be of relatively recent geological origin (Ollier 1986), unlike the Southern Andes Cordillera of Patagonia (Bouza and Bilmes 2020).
The biogeography of the Carpha clade has not yet been investigated in detail, but long-distance dispersal is likely to be involved, at least for the species of the high-altitude mountains; see, for example, Wardle et al. (2001) on New Zealand–South American links including Carpha spp.; Jordan (2001) on Tasmanian–New Zealand links including Carpha alpina; Verboom (2006) and Viljoen et al. (2013) on the origins of tribe Schoeneae; Gehrke and Linder (2014) on the tropical Afro-alpine flora. Vicariance could have been possible in the case of Australasia–South America, given the apparent link via Antarctica until c. 30 Ma (Sanmartín and Ronquist 2004; Sanmartín et al. 2007; Elliott et al. 2021) but that would be near the oldest estimated age for Carpha species (Wardle et al. 2001) and therefore is perhaps less likely than more recent long-distance dispersal. Such dispersal events have probably been very rare. Possible dispersal agents are hypothesised to be birds and wind (Muñoz et al. 2004; Sanmartín and Ronquist 2004; Green et al. 2023) but much more research is needed on diaspore dispersal and subsequent establishment (Nathan et al. 2008; Wu et al. 2023). Another avenue to be explored is molecular analysis of the species involved to assess similarity between the distant populations, as has been performed for Abrotanella Cass. (Asteraceae; Wagstaff et al. 2006).
Taxonomy
A key to the genera in tribe Carpheae is provided for completeness (updating that in Larridon et al. 2021), but Trianoptiles is not further considered in this taxonomic section. Table 2 provides an index of names published in Asterochaete and Carpha.
| 1 | Plants perennial without amphicarpy; perianth bristles 6, subequal in length or with 3 shorter than the nutlet and 3 that are subequal to or exceed the length of the nutlet...2 Plants annual with frequent amphicarpy; perianth bristles absent from basal spikelets, 6 in aerial spikelets with 3 very reduced and 3 longer than the nutlet, often broadly flattened towards the base; the apex usually with a shorter bristle on either side of the central bristle...Trianoptiles |
| 2 | Only lower sterile glumes persistent, upper fertile glumes deciduous; perianth bristles antrorsely scabrous throughout their length or scabrous above and slightly plumose or glabrous towards the base, differing in length, the inner whorl of 3 longer than the outer whorl of 3, filiform to terete for most their length, only obviously flattened or enlarged at the base; nutlet sessile to subsessile on a very short gynophore; persistent stylebase much shorter than the nutlet body (Africa; Fig. 3)...Asterochaete Glumes all persistent; perianth bristles plumose for at least half of their length (the apical section is antrorsely scabrous for up to 2.7 mm), all subequal, and flat for much of their length, tapering to terete or filiform near the apex; nutlet with an obvious gynophore; persistent stylebase longer then the nutlet body (up to 2.5 times as long) (Australia, New Guinea, New Zealand, South America; Fig. 3)...Carpha |
| Asterochaete acuminata X.F.Zhang | |
| A. angustifolia Nees, nom. nud. = A. capitellata | |
| A. angustissima (Cherm.) X.F.Zhang | |
| A. arundinacea (Sol. ex Vahl) Kunth = Tetraria arundinacea (Sol. ex Vahl) T.Koyama | |
| A. borbonica (Steud.) X.F.Zhang | |
| A. capitellata Nees | |
| A. dactyloides (Vahl) K.L.Wilson & J.J.Bruhl | |
| A. discolor T.H.Arnold ex X.F.Zhang | |
| A. elongata Kunth = Costularia melicoides (Poir.) C.B.Clarke | |
| A. eminii (C.B.Clarke) X.F.Zhang | |
| A. filifolia (C.Reid & T.H.Arnold) X.F.Zhang | |
| A. glomerata (Nees) Nees = A. dactyloides | |
| A. glomerata var. minor Nees = A. capitellata | |
| A. ludwigii Hochst. = A. capitellata | |
| A. nitens Kunth | |
| A. schlechteri (C.B.Clarke) X.F.Zhang | |
| A. tenuis Kunth = A. nitens | |
| A. ulugurensis Nelmes ex X.F.Zhang | |
| Carpha alpina R.Br. | |
| C. alpina var. schoenoides (Banks & Sol. ex Hook.f.) Kük. = C. schoenoides | |
| C. alpina var. subacaulis Kük. = C. nivicola | |
| C. andina Phil. = Schoenus andinus (Phil.) H.Pfeiff. | |
| C. angustissima Cherm. = A. angustissima | |
| C. antarctica (Hook.f.) C.B.Clarke = Schoenus antarcticus (Hook.f.) Dusén | |
| C. aristata Kük. = nomen dubium | |
| C. arundinacea Boeckeler = Tetraria arundinacea (Sol. ex Vahl) T.Koyama | |
| C. arundinacea Brongn. = Costularia pilisepala (Steud.) J.Kern | |
| C. aubertii Nees ex Boeckeler = A. nitens | |
| C. aubertii var. explicatior C.B.Clarke, nom. inval. = Costularia melicoides (Poir.) C.B.Clarke. | |
| C. avenacea R.Br. = Cyathochaeta avenacea (R.Br.) Benth. | |
| C. borbonica (Steud.) C.B.Clarke = A. borbonica | |
| C. bracteosa C.B.Clarke = A. capitellata | |
| C. capensis (Steud.) M.Martens ex Steud. = Trianoptiles capensis (Steud.) Harv. | |
| C. capitellata (Nees) Boeckeler = A. capitellata | |
| C. capitellata var. bracteosa (C.B.Clarke) Kük. = A. capitellata | |
| C. clandestina R.Br. = Cyathochaeta clandestin (R.Br.) Benth. | |
| C. costularioides C.B.Clarke = Costularia melicoides (Poir.) C.B.Clarke | |
| C. curvata W.M.Curtis | |
| C. deusta R.Br. = Ptilothrix deusta (R.Br.) K.L.Wilson | |
| C. diandra R.Br. = Cyathochaeta diandra (R.Br.) Nees | |
| C. elongata (Kunth) Boeckeler = Costularia melicoides (Poir.) C.B.Clarke | |
| C. eminii (K.Schum.) C.B.Clarke = A. eminii | |
| C. eminii var. angustissima (Cherm.) Kük. = A. angustissima | |
| C. filifolia C.Reid & T.H.Arnold = A. filifolia | |
| C. glomerata Nees = A. dactyloides | |
| C. graciliceps C.B.Clarke = Mesomelaena graciliceps (C.B.Clarke) K.L.Wilson | |
| C. hexandra Nees = Cyathocoma hexandra (Nees) Browning | |
| C. junciformis Boeckeler = Tricostularia undulata (Thwaites) J.Kern | |
| C. laxa (Hook.f.) H.Pfeiff. = Schoenus rhynchosporoides (Steud.) Kük. | |
| C. nitens (Kunth) Kük. = A. nitens | |
| C. nivicola F.Muell. | |
| C. novae-zelandiae Gand. = C. alpina | |
| C. paniculata Phil. = Schoenus rhynchosporoides (Steud.) Kük. | |
| C. perrieri Cherm. = A. capitellata | |
| C. rodwayi W.M.Curtis | |
| C. schlechteri C.B.Clarke = A. schlechteri | |
| C. schoenoides Banks & Sol. ex Hook.f. | |
| C. schweinfurthiana Boeckeler = Coleochloa schweinfurthiana (Boeckeler) Nelmes | |
| C. solitaria (C.B.Clarke) H.Pfeiff. = Trianoptiles solitaria (C.B.Clarke) Levyns | |
| C. tasmanica Gand. = C. alpina | |
| C. urvilleana Gaudich. ex Boeckeler = Costularia pilisepala (Steud.) J.Kern | |
| C. viridis Phil., nom. nud. = C. schoenoides | |
| Chaetospora alpina (R.Br.) F.Muell. = Carpha alpina | |
| C. dactyloides (Vahl) Dietr. = A. dactyloides | |
| Cyclocampe aubertii (Nees ex Boeckeler) Pax = A. nitens | |
| Elynanthus borbonicus Steud. = A. borbonica | |
| Ficinia trinkleriana H.Pfeiff. = A. capitellata | |
| Oreograstis eminii K.Schum. = A. eminii | |
| Rhynchospora boivinii Boeckeler = Carpha nitens | |
| Schoenus dactyloides Vahl = Asterochaete dactyloides | |
| S. glomeratus L. = Rhynchospora glomeratus (L.) Vahl | |
| S. glomeratus Thunb. = A. dactyloides |
Names in bold and italic are accepted names; those in italic only are synonyms and invalid names that appear in the literature. This table does not include names of fungi published under the later homonym Asterochaete (Patouillard) Bondartsev & Singer.
Asterochaete Nees, Linnaea 9 (3): 300 (1834)
Type: Asterochaete glomerata Nees (= Asterochaete dactyloides (Vahl) K.L.Wilson & J.J.Bruhl)
Oreograstis K.Schum. in A. Engler (ed.), Pflanzenw. Ost-Afrikas C, 127 (1895). Type: Oreograstis eminii K.Schum.
Perennial herbs with short rhizome. Culms erect, more or less terete to trigonous in cross-section, unnoded or 1–5-noded. Leaves basal and cauline, eligulate, not obviously pseudopetiolate. Inflorescence narrow, panicle-like with mostly slender branches, with spikelets loosely arranged to densely clustered. Involucral bracts leaf-like. Spikelets compressed, with 1 or 2 bisexual flowers (lower flower male in A. schlechteri); glumes (floral bracts) 4–6(–7), distichously arranged, the lower ones empty, persistent, approximately half the length of the upper glumes (which are deciduous); the uppermost glume sometimes linear, empty. Perianth consisting of 6 bristles in 2 whorls, falling with the nutlet, whitish to dark golden brown, antrorsely scabrous, sometimes with shorter or slightly longer hairs or glabrous in the lower half (often only at the very base), and mostly with the inner whorl obviously longer than the outer whorl. Stamens 3; anthers pale grey to blackish in most species, pale yellow when immature in A. dactyloides, A. discolor and A. schlechteri (remaining so at maturity in the latter two species), with a short pyramidal to conical apical appendage. Style slender, continuous with the ovary, lower part slightly enlarged near base, persistent on the nutlet; stigmas 3. Nutlet trigonous in cross-section, sessile or subsessile on a very short gynophore, with surface reticulate, sometimes puncticulate.
A genus of 11 species, found at high altitude in eastern Africa, Madagascar and the Mascarenes, and from low to high altitudes in southern Africa (Fig. 3, 4). Most species are relatively narrow endemics, but A. capitellata (as currently understood) is widespread (in South Africa, Zimbabwe and Madagascar), whereas A. dactyloides is found from the Western Cape to Eastern Cape regions of South Africa.
Distribution of Asterochaete spp.: (a) A. acuminata (circle), A. discolor (stars); (b) A. angustissima (stars), A. borbonica and A. nitens (inverted triangles), A. capitellata (circles), A. eminii (triangle), A. ulugurensis (square); (c) A. dactyloides (circles); (d) A. filifolia (circles), A. schlechteri (stars).
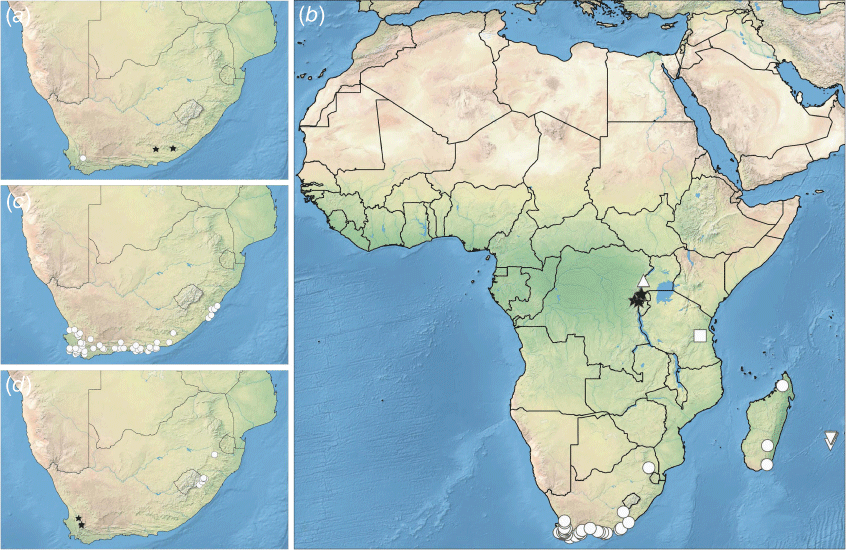
The phylogeny and biogeography of Asterochaete species need further study to elucidate them more fully. The distribution of its species is similar to that found in other plant groups in Africa such as Disa P.J.Bergius (Orchidaceae), Iridieae p.p. (Iridaceae), the Pentaschistis Stapf clade (Poaceae), the family Restionaceae, and Senecio L. and related genera (Asteraceae) (Linder 2003; Galley et al. 2007; Galbany-Casals et al. 2014; Kandziora et al. 2016). In those groups, the evidence suggests that the lineages originated in the Greater Cape Floristic Region and spread from there to the Afro-alpine regions of Tropical East Africa (Galley et al. 2007; Galbany-Casals et al. 2014; Gerhke and Linder 2014) via the Drakensberg Range (Kandziora et al. 2016) and, in some cases, spreading farther afield to Madagascar, Réunion and other areas. Verboom et al. (2015) suggested that the topographic heterogeneity of the Cape Region has promoted the diversity of species found there, whereas Cramer et al. (2014) and van Santen and Linder (2020) proposed a strong role for the oligotrophic soils of that region in defining the composition of the flora. The East African Rift Western Branch mountains are estimated to vary in age from 12.6 to 3 Ma, and the Afro-alpine flora is considered to be mostly of recent origin, probably no more than 10–5 Ma (Gehrke and Linder 2014; Kandziora et al. 2022). These mountains are mostly of volcanic origin (e.g. Virunga–Kahuzi-Biéga mountains), except for Rwenzori, which consists mostly of a Precambrian metamorphic basement block, with some areas covered with Neogene volcaniclastics (Brochmann et al. 2021). The Eastern Arc Mountains (including the Ulugurus) are also Precambrian crystalline basement blocks geologically (Burgess et al. 2007), reaching high altitudes but not high enough to be considered Afro-alpine (Brochmann et al. 2021). The summit of the Lukwangale Plateau is described as having subalpine communities by Pócs (1976).
| 1 | Leaf blades without an obvious abaxial midrib or keel at mid-length of the blade, thickly crescentiform in cross-section or thickly channelled or ellipsoid with small channel on one side or semicircular, or ±trigonous and strongly angled abaxially (slender specimens of A. capitellata, e.g. Reid 1807, or A. angustissima, e.g. Troupin 14282, may occasionally have a mixture of V-shaped and slightly thickened U-shaped leaves, see below)...2 Leaf blades with an obvious abaxial midrib or keel at least at mid-length, V-shaped to thinly U-shaped to nearly flat in cross-section when fresh (often folded when dried)...5 |
| 2 | Perianth bristles hispid to shortly plumose (hairs up to ~0.5 mm long) for lower half of length, antrorsely scabrous above; older leaves strongly curling in upper half; glumes mixed in colour, mainly mid- to dark red-brown, but pale yellow-brown near base and with conspicuous hyaline to whitish margins (~0.2–0.3 mm wide); anther apical appendage 0.3–0.5 mm long...A. discolor Perianth bristles antrorsely scabrous throughout their length (only the outer whorl in A. filifolia) or smooth near base and antrorsely scabrous above, occasionally with a few longer hairs at base (the inner whorl in A. filifolia hispid to shortly plumose (hairs up to 0.4 mm long) only at the base or rarely for 1/3 of their length; scabrous above); leaves not strongly curling with age; glumes more or less uniformly pale yellow-brown to mid-yellow-brown, with hyaline margins ranging from <0.1 to 0.3 mm wide at the widest point; anther apical appendage 0.1–0.3 mm long...3 |
| 3 | Involucral bracts with much broadened bases partly sheathing the clusters of spikelets; lowest involucral bract (and usually several others) longer than the inflorescence; leaf blades 1.5–2.5 mm in diameter...A. acuminata Involucral bracts not markedly broadened at the base; lowest involucral bract shorter than to longer than the inflorescence; leaf blades ±filiform, 0.5–0.8 mm in diameter...4 |
| 4 | Inner whorl of perianth bristles antrorsely scabrous for whole length; involucral bracts shorter than to ~1/3 longer than inflorescence; nutlets 1.7–2.0 mm long; leaf sheaths dull, pale yellow-brown with mid- to dark yellow-brown patches below; apex of uppermost leaf sheath rather broadly V-shaped (regularly or obliquely) opposite the junction with the blade...A. borbonica Inner whorl of bristles hispid to shortly plumose (hairs up to 0.2 mm long) near the base (rarely for 1/3 of the length), antrorsely scabrous above; the involucral bracts 3 times longer than to rarely subequalling the inflorescence; nutlets 3.5–4.3 mm long; leaf sheaths shining, golden brown to very dark brown at the base; apex of uppermost leaf sheath narrowly and regularly V-shaped opposite the junction with the blade, with margins overlapping near base of the ‘V’...A. filifolia |
| 5 | Bristles with slender glabrous basal part 0.8–1.7 mm long (1/3 to 3/5 of length), upper part (disarticulating from basal part so often missing from specimens) somewhat more robust and antrorsely scabrous; robust plants (0.8–)1–3 m tall; leaf blades 5–25 mm wide at mid-length...6 Bristles antrorsely scabrous for all of their length, or with a few longer hairs (up to 0.2 mm long) or glabrous for 0.2–0.6 mm near the base, and not disarticulating; more or less slender plants 20–80 (rarely up to 130) cm tall; leaf blades 1–5 mm wide at mid-length...7 |
| 6 | Spikelets 6–9 mm long, with 2 bisexual flowers; fertile glumes 5.5–7.5 mm long, 1.5–2.8 mm wide; nutlets 2.5–3.4 mm long; leaf blades 7–24 mm wide; apex of leaf sheath U-shaped (usually broadly so, sometimes narrowly so) opposite junction with blade; longest perianth bristles 3.1–3.8 mm long; anthers pale grey at maturity...A. dactyloides Spikelets 4–5 mm long, with 1 male flower below the bisexual flower; fertile glumes 3.3–4.3 mm long, 0.5–1.8 mm wide; leaf blades 4.5–8 mm wide; apex of leaf sheath V-shaped to narrowly U-shaped opposite junction with blade; nutlets 2.0–2.5 mm long; longest bristles 2.3–3.4 mm long; anthers pale yellow at maturity...A. schlechteri |
| 7 | Inflorescence branches ending in dense hemispherical to ovoid clusters of numerous pale to mid-yellow-brown spikelets; nutlets with large (~0.1 mm in diameter) epidermal cell outlines...A. capitellata Inflorescence branches ending in sparse to moderately dense, oblong clusters with few to numerous red-brown to dark red-brown spikelets (if paler yellow-brown, then only 1–5(–10) spikelets per cluster); nutlets with small to very small (≤0.05 mm in diameter) epidermal cell outlines...8 |
| 8 | Nutlets 1.5–1.8 mm long (Réunion)...A. nitens Nutlets 2.0–3.0 mm long (Tropical East Africa)...9 |
| 9 | Longest perianth bristles (usually 3 of them) much longer than the nutlet body (up to 2 times as long), 3.5–6.7 mm long; spikelets predominantly yellow-brown...A. eminii Longest perianth bristles shorter than to slightly longer than the nutlet body (0.5–1.5 times as long), 2.0–4.4 mm long; spikelets predominantly red-brown...10 |
| 10 | Plants 20–50 cm high; inflorescence branches slender and drooping; leaves 0.8–1.5 mm wide; perianth bristles with hairs slightly longer towards the base of the bristles than at their apex; nutlets yellow-brown...A. angustissima Plants 70–90 cm high; inflorescence branches rigid and erect; leaves 2.2–5.5 mm wide; bristles with hairs similar over whole length of bristles, but often glabrous at the base; nutlets mid- to dark red-brown...A. ulugurensis |
Asterochaete acuminata X.F.Zhang, sp. nov.
Planta formam A. capitellatae sens. str. simulans sed in characteribus foliorum, bracteolarum et inflorescentiarum differt. Folia crassa canaliculata ecarinata. Bracteolae basi dilatatae ovatae, apice longe acuminato, inflorescentiam vel capitula superantes 2–10 cm. Spiculae in capitula oblonga dispositae, non in capitula ovata dense congesta.
Type: South Africa: Eastern Cape: Hogsback, alt. 1700 m, 24 Nov. 1994, B. Sonnenberg 301 (holo: NU; iso: GRA n.v., NE 66170).
[Carpha sp. A: Archer in Bredenkamp, Fl. Eastern Cape Province 3: 1702 (2019)].
Tufted perennial, 35–70 cm high; rhizome apparently short. Culms 1-noded, the node bearing a single leaf, rarely unnoded. Leaves crowded at the base of the culms; blades straight, 30–60 cm long, 0.9–2.5 mm wide; proximal half thickly channelled in cross-section; distal half thickly channelled to subcircular or elliptical in cross-section; sheaths pale yellow-brown with patches of dark yellow-brown. Inflorescence with several more or less short internodes, 3–12 cm long; spikelets 18–100 in one or few oblong clusters. Involucral bracts several, 5–21 cm long, broadened at the base and 3–6 mm wide, partly sheathing the spikelet clusters, with a very long acuminate apex, several overtopping the inflorescence by 2–10 cm, ~1.2–3 times as long as (rarely subequalling) the inflorescence. Spikelets 6–8 mm long, 2-flowered; glumes 5 or 6, pale yellow-brown to mid-brown; lowest 3 or 4 glumes empty, persistent; the upper 2 fertile, deciduous; lowest glumes shorter than the rest; the proximal fertile glume 5–6.7 mm long, 1.8–2 mm wide; the second fertile glume 4.7–6 mm long, 0.9–1.1.8 mm wide; axis elongated above proximal fertile glume and adnate to the base of that glume. Bristles: outer 3 bristles shorter than (occasionally only ~1/8 as long as) to rarely slightly exceeding the body of the nutlet; the inner 3 bristles shorter than to slightly exceeding the nutlet body, 3.0–3.7 mm long. Anthers pale to dark grey, 2.0–2.7 mm long (excluding appendage), with apical appendage 0.1–0.3 mm long, 0.1 mm wide. Stylebase 0.9–1.9 mm long, 0.15–0.2 mm wide, hispidulous with hairs extending to upper ridges of nutlet. Nutlets 2 per spikelet, narrow-ellipsoid to ellipsoid or narrow-obovoid, 2.8–3.5 mm long, 0.9–1.1 mm wide, mid-orange-brown at maturity, surface finely reticulate and puncticulate (Fig. 1a, 5; see also Zhang et al. 2004b, fig. 7k; Zhang et al. 2006, fig. 3b as C. bracteosa).
Holotype of Asterochaete acuminata (B. Sonnenberg 301, NU). Reproduced with permission of Bews Herbarium, University of KwaZulu-Natal, Pietermaritzburg.
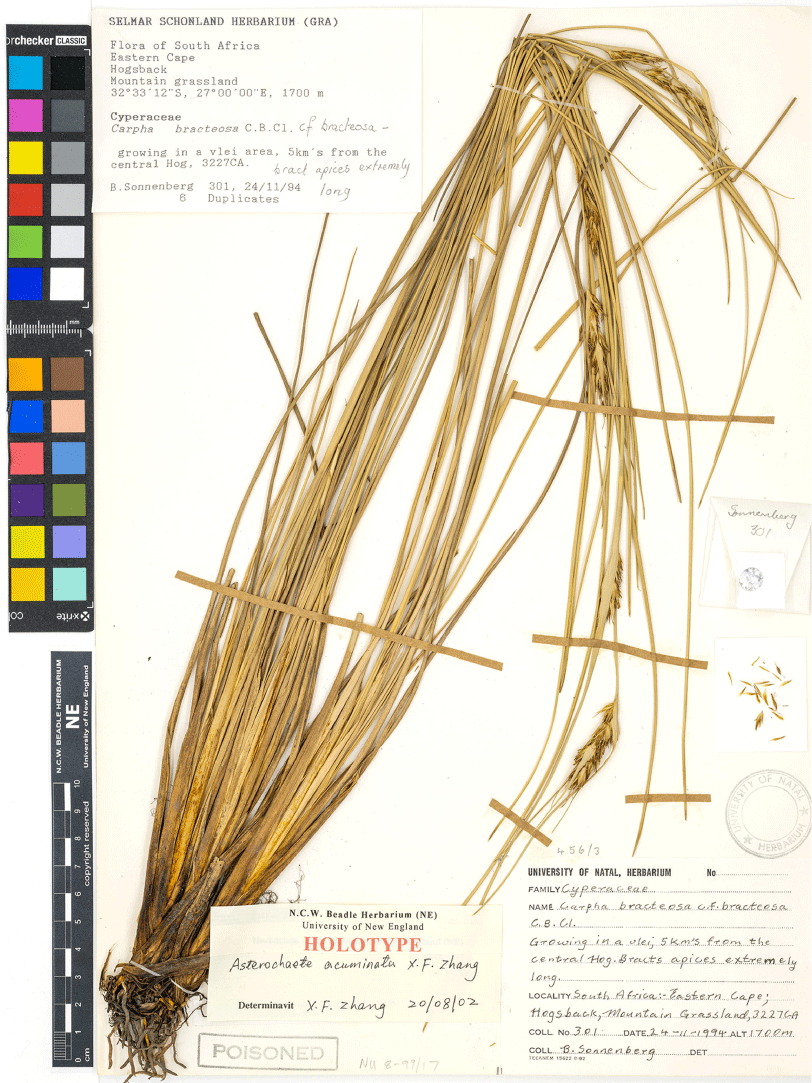
In Eastern Cape Province, South Africa (Fig. 4a). In marshes and on rocky river banks in montane grasslands, at 1300–1700 m in altitude, where it may be a co-dominant in its community (Brand et al. 2013).
The specific epithet reflects the distinctive shape of the involucral bracts, which are broadened and more or less ovate at the base, tapering gradually to the long-acuminate apex, one of the key distinguishing features of this species.
Asterochaete acuminata resembles A. capitellata, but differs in characters of the leaves, involucral bracts, inflorescences and nuts. Its leaves are thick, the proximal half thickly crescentiform in cross-section, the distal half thickly crescentiform to subcircular in cross-section, whereas the leaves of A. capitellata are thin and V-shaped in cross-section. Its bracts are ovate at the base with a long acuminate apex, 2–10 cm longer than the inflorescence or heads of spikelets, whereas the bracts of A. capitellata range from ovate to only slightly broadened at the base and are the same length as or only slightly longer than the heads of spikelets. The spikelets of A. acuminata are in one or few oblong heads rather than in ovoid heads as in A. capitellata. Its nutlet surface is finely reticulate (epidermal-cell outlines ~0.05 mm in diameter) and puncticulate, whereas the nutlet surface of A. capitellata is reticulate (cell outlines ~0.1 mm in diameter) and not or rarely puncticulate (Zhang et al. 2006, fig. 3a).
Asterochaete filifolia also has relatively long involucral bracts (the proximal bracts are usually 1.5–3 times longer than the inflorescence) and single-noded culms, but differs from A. acuminata in having narrow bases to the involucral bracts (including the bracts subtending the cluster(s) of spikelets); spikelets in one very dense conical cluster (rarely two clusters); leaves filiform, 0.5–0.8 mm in diameter; and nutlets 3.5–4.3 mm long.
Two specimens of this species, MacOwan 1616 and 2187 (see Zhang et al. 2006, fig. 14), were cited as Carpha bracteosa by Clarke (1898) when he described that species. The application of that name is fixed in this paper by lectotypifying it (see below under A. capitellata) on the third specimen cited by Clarke, which is A. bracteosa as traditionally understood. More recent specimens of this new species were also identified as C. bracteosa by other workers. These specimens were temporarily named as Carpha cf. bracteosa in our analyses (Zhang 2002; Zhang et al. 2006).
Of limited distribution but does occur in national parks; we suggest a status of ‘Data Deficient’ (IUCN Standards and Petitions Committee 2022).
South Africa: Eastern Cape: Uitenhage Division, Great Winterhoek Mountains, The Cockscomb, 30 Nov. 1958, E. Esterhuysen 28040 (K 000480712, PRE n.v.); in summo monte Boschberg, alt. 4800 ft [~1460 m], Somerset Division, Dec. [?year; received at Kew March 1871], P. MacOwan 1616 (K, Cibachrome print in NE, SAM 24258-1, SAM 24258-2); in paludosis summi Boschberg, 4500 ft [~1370 m], Nov.–Dec. [?year; received at Kew 1874], Somerset Division, P. MacOwan 2187 (K, Cibachrome print in NE); Amatole Mountains, below Gaikas Kop, 1 Dec. 1983, H.D. Furness 347 & P.B. Phillipson (K, UFH n.v.); Robertson Falls, 1500 m, 26 Nov. 1994, B. Sonnenberg 336 (NU); ridge W of Mount Thomas, 4500 ft [~1370 m], 11 Nov. 1948, R. Storey 3682 (K, PRE n.v.).
Asterochaete angustissima (Cherm.) X.F.Zhang, comb. nov.
Carpha angustissima Cherm., Bull. Soc. Bot. France 82: 341 (1935). Carpha eminii var. angustissima (Cherm.) Kük., Feddes Repert. 47: 210 (1939). Type citation: ‘Massif du Kahuzi (O. du lac Kivu), 2400 m, février 1929 (Humbert 7722); volcan Karisimbi (NE. du lac Kivu), 3300 m., juin 1929 (H. Humbert 8586).’ Type: Democratic Republic of Congo: Massif du Kahuzi (O. du Lac Kivu), 2400 m, Feb. 1929, H. Humbert 7722 (lecto, here designated: P 742029; isolecto: B 10 0000961, BR 461836*, BR 8644640*, BR 8644657*, P 00461836, P 00461977, P 00461837). Residual syn: Rwanda: Volcan Karisimbi (au N.-E. du lac Kivu), 3300 m, June 1929, H. Humbert 8586, (B 10 0000959, BR 5441426*, BR 5578160*, BR 5577514*, BR 5578160*, K (fragment ex B in packet on sheet of Purseglove 2199, with notes by Nelmes on the packet and sheet); P 00461978, P 00461838 (2 sheets), US 01117924).
Tufted perennial, 25–50 cm high; rhizome short to long. Culms unnoded, rarely 1-noded with a single cauline leaf. Leaves crowded at the base of the culms; blades straight, 15–35 cm long, 0.8–1.5(–2.0) mm wide, more or less flat to V-shaped or U-shaped in cross-section, V-shaped near apex, with a slightly thickened midrib or keeled abaxially (very slender leaves may have a poorly developed midrib), usually smooth but sometimes sparsely scabrous; sheaths dull mid- to dark orange to red-brown below, pale yellow-green towards apex. Inflorescence 6–35 cm long, with 2–4 internodes; spikelets 12–35 loosely arranged in small clusters (not dense). Involucral bracts several; lowest equalling to slightly exceeding the inflorescence, 15–30 cm long, to 1.5 mm wide. Spikelets 5.1–6.5 mm long, with 2 (very rarely 1) bisexual flowers; glumes 5, pale yellow-brown to mid-brown; lowest 3 glumes empty, persistent; the upper two fertile, deciduous, very rarely only the upper one fertile and the uppermost one sterile; the lowest 2 glumes shorter than the rest; the proximal fertile glume 4.6–5.5 mm long, 1.1–2 mm wide, the second fertile glume 3.5–5 mm long, 0.5–1.4 mm wide; the axis elongated above proximal fertile glume and adnate to the base of that glume. Bristles: outer 3 bristles shorter than inner 3 and much shorter than the body of the nutlet, lower part of bristles slightly compressed and antrorsely scabrous with sparse slightly longer hairs near the base (to twice as long right at the base), and upper half antrorsely scabrous, OR outer bristles evenly and shortly antrorsely scabrous over their whole length; the inner 3 (2.0–)3–4.4 mm long, as long as to exceeding the body of the nutlet by up to ~1/2 of the length; lower part of bristles in inner whorl more or less flattened and antrorsely scabrous with sparse slightly longer hairs near the base (up to twice as long right at the base). Anthers greyish, 1.5–4 mm long (excluding appendage), with apical appendage 0.02–0.15 mm long, 0.05–0.15 mm wide. Stylebase 0.7–1.1 mm long, 0.15–0.3 mm in diameter, glabrous at base, very sparsely hispidulous, not extending onto the apex of the nutlet. Nutlets 2 per spikelet, very rarely 1, ellipsoid to narrow-ellipsoid, 2.6–2.9 mm long, 0.8–1.1 mm in diameter, pale yellow-brown to mid-orange-brown at maturity, ribs not paler, surface reticulate and puncticulate. (Fig. 1b; see Zhang et al. 2004b, fig. 7k (rarely e) for spikelet structure; Haines and Lye 1983, fig. 663; Hoenselaar et al. 2010, fig. 55).
Tropical East Africa, on Mount Karisimbi, Mount Mgahinga, Mount Muhavura in the Virunga Mountains along the border between Uganda, Rwanda and the Democratic Republic of Congo; also on Mount Bigugu (SW Rwanda) and Mount Kahuzi (D.R. Congo) (Fig. 4b). In alpine bogs and heaths at ~2400‒3300 m in altitude.
Asterochaete angustissima was previously treated either as a species of Carpha (Chermezon 1935; Haines and Lye 1983) or as a variety of C. eminii (=Asterochaete eminii) (Kükenthal 1939b; Napper 1964), which is in the nearby Rwenzori Mountains in western Uganda. Our studies (Zhang 2002; Zhang et al. 2006) indicated that A. angustissima would be better treated as a species rather than a variety of A. eminii. It differs from A. eminii by usually having two flowers in each spikelet, with the axis elongated above the proximal fertile glume and adnate to the fertile glume base, the shorter perianth bristles (inner whorl (1.8–)3–4.4 mm long, and the outer whorl even shorter), and the leaf blades slender and usually canaliculate to thinly but stiffly crescentiform in cross-section, 0.7–1.6(–2.0) mm wide. A. eminii has one-flowered spikelets, with the axis not elongated above fertile glumes, longer perianth bristles (inner whorl 3.5–6.7 mm long), and the leaf blades V-shaped in cross-section, 2.0–4.0 mm wide at mid-length. Plants of A. angustissima are generally more slender, with few and shorter spikelets in the inflorescence, with shorter, paler glumes, and leaf sheaths that are more brightly coloured.
The third tropical montane African species, A. ulugurensis (q.v.), endemic to the Uluguru Mountains in Tanzania, is generally more robust in its parts than are A. angustissima and E. eminii, with darker glumes.
Enquiries at STR (where Chermezon’s main herbarium is held) failed to find either of the Humbert collections cited in the protologue (Françoise Deluzarche 2004, pers. comm.). In P, there are sheets of these collections that are annotated by H. Chermezon so it is reasonable to choose one of these as lectotype. The sheet of Humbert 7722 that is labelled by Chermezon as ‘Carpha angustissima H.Chermezon’ is chosen because it is labelled as having been in ‘Hb. Chermezon’ (printed label) and has been annotated by Chermezon. There is a probable isolectotype in B (10 0000961) that has not been annotated by Chermezon. In P, there is another sheet of Humbert 7722 (P 00742030) that differs in collecting details; according to the printed label it was collected ‘vers 3000 m’ in March (‘Mars’) 1929. Probable duplicates of this sheet are in B (10 0000960) and NSW (384362) but these differ in having the date as ‘Février – Mars’. Given these different details, we do not consider any of these variant sheets to be isolectotypes. The two sheets of Humbert 8586 in P have not been determined by Chermezon in his own handwriting, although on one sheet he has added habitat information. The sheet in B (10 0000959) has been determined by Chermezon.
Of limited distribution but some populations are in national parks; listed as ‘Endangered’ (Beentje 2017, as Carpha angustissima), which seems appropriate given that the high-altitude habitat is limited and predicted to become even more so with changing climate and increasing human impact.
Rwanda: Mount Bigugu, 2870 m, Prefecture Cyangugu, 4 Mar. 1972, P. Bamps 3297 (BR 13663056*, P 199387); Forêt de Nyungwe, sommet du Mont Ibigugu, alt. 2954 m, Prefecture Cyangugu, 1972, G. Bouxin 1360 (BR 13663117*, P 199385, L 486009); Mont Bigugu, Terr. Shangugu, 2900 m, 26 Mar. 1956, R. Christiansen 1642 (BR 13663063*, P 03471933); Parc des Volcans, piste vers la selle Ngahinga-Muhavura, Préf. Ruhengeri, 2840 m, 31 July 1974, P. Van der Veken 10449 (BR 13663094*, P 01948986).
Uganda: Kigezi District: crater of Gahinga Mountains, 3000 m, Bufumbira County, 24 Apr. 1970, A.B. Katende & K. Lye K207B (K); Gahinga-Muhavura Saddle, 3050 m, Bufumbira County, 24 Apr. 1970, K.A. Lye 5289 & A. Katende (EA, K, P 03471934); Muhenva-Mgahenga Saddle, 10,000 ft [~3000 m], Sep. 1946, J.W. Purseglove 2199 (BR 13662998*, EA, K); Western Province, Virunga-Kette, Gahinga, ~3400 m, 15 Nov. 1954, H.U. Stauffer 793 (BR 24800396*, K, P 03471932).
Democratic Republic of Congo: Mount Kahuzi (Kivu), 3000 m, 25 May 1958, A. Meurillon 626 (BR 13663124*, K); Mt Kahuzi, 3300 m, Province Kivu, 29 Dec. 1971, G. Troupin 14282 (BR 13663148*, LWI 444303451 n.v., P 00199386); Kivu, Parc National du Kahuzi-Biega, Mont Kahuzi, alt. 3200 m, 29 Dec. 1971, P. Van der Veken 8882 (AAU, BR 25390414V*).
Asterochaete borbonica (Steud.) X.F.Zhang, comb. nov.
Elynanthus borbonicus Steud., Syn. Pl. glumac. 2: 155 (1855) (as E.? borbonicus). Carpha borbonica (Steud.) C.B.Clarke in T.A. Durand & H. Schinz (eds), Consp. Fl. Afr. 5: 655 (1894). Type citation: ‘Ins. Borbonia’. Type: Réunion: s. loc., [no date], H. Giraudy in herb. Steudel [no other details] (lecto, here designated: P 552881); Ile Bourbon, 1847, H. Giraudy (isolecto: P 592877).
Erect perennial, 30–50 cm high; rhizome short, slender. Culms 1–2-noded, each node bearing a single leaf. Leaves crowded at the base of the culms; blades straight, up to 20 cm long, 0.4–0.7 mm in diameter, nearly filiform, thickly U-shaped to rigidly crescentiform in cross-section, not or scarcely keeled except near apex; sheaths pale yellow-brown above, dark brown towards base. Inflorescence 1.5–11 cm long, usually with 3 short internodes; spikelets 15–30 in 1–3 dense oblong clusters. Involucral bracts several, up to 17 cm long, shorter than to slightly exceeding the inflorescence, up to 0.7 mm wide. Spikelets 5.0–6.0 mm long, with 1 or 2 bisexual flowers; glumes 5 or 6, pale yellow-brown to dark yellow-brown or red-brown; lowest 3 or 4 glumes empty, persistent, the upper 2 fertile, deciduous, or the uppermost one sterile; the lowest 2 or 3 glumes shorter than the rest; the proximal fertile glume 4.5–5.0 mm long, 1.3–1.5 mm wide, the second fertile glume ~4.5 mm long, ~1 mm wide; axis elongated above proximal fertile glume and adnate to the base of that glume. Bristles: outer 3 bristles shorter than or rarely subequal to inner 3 (often 1 less than half of the length of the fruit), the inner 3 1.8–2.5 mm long, slightly shorter than to slightly exceeding the body of the nutlet. Anthers greyish, 1.3–1.5 mm long (excluding appendage), with apical appendage ~0.1 mm long, ~0.1 mm wide. Stylebase ~0.6 mm long, ~0.15 mm in diameter, very shortly hispidulous to verruculose. Nutlets 1 or 2 per spikelet, ellipsoid, 1.7–2.0 mm long, 0.7–0.8 mm in diameter, brown at maturity, surface regularly reticulate. (Fig. 1c; see Zhang et al. 2004b, fig. 7e, k for spikelet structure).
Endemic to the Mascarene Island of Réunion (previously called ‘Ile Bourbon’), in the Indian Ocean (Fig. 4b). In montane shrubland in deep clay-loam soil on basalt at high altitude (~1950–2300 m).
Asterochaete borbonica is more slender than A. nitens, the other species on Réunion (also endemic there). It differs from that species principally in being shortly rhizomatous, with more or less filiform leaves crescentiform in cross-section and with shorter perianth bristles. Some specimens are morphologically intermediate (e.g. Bruhl 2984 and 2986), with leaves U-shaped in cross-section and up to ~2 mm wide without prominent midrib abaxially, and with longer perianth bristles (up to 5 mm long). Further study is needed to determine whether these names represent extremes of one taxon.
Asterochaete borbonica is a slender species with nearly filiform leaf blades that are thickly channelled in cross-section, 0.5–0.7 mm wide and lacking an obvious midrib abaxially. The uppermost leaf at the base of a culm (and the lowest primary involucral bract) has the junction of the leaf sheath and blade marked by an elongated V-shaped opening in the sheath opposite the attachment of the blade, with the margins of the V often slightly overlapping towards its base or narrowly U-shaped.
The inflorescences in A. borbonica are slender and short (up to 11 cm long). The perianth bristles are shorter than or subequal to the fruit body, 1.8–2.5 mm long, often with some bristles less than half the length of the body. The bristles are more or less terete in cross-section, covered densely by short, rigid hairs and of equal diameter throughout their length, tapering abruptly to the relatively much-enlarged, short, smooth basal section. The nutlets are obovoid to ellipsoid, and the persistent stylebase is short (<0.5 mm long) and not clearly differentiated from the upper part of the style (Fig. 1c).
Asterochaete nitens is a generally taller and more robust species with leaf blades broadly V-shaped in cross-section, 2.5–5 mm wide and with an obvious midrib abaxially. The opening at the sheath–blade junction of the uppermost leaf is mostly deeply U-shaped, often asymmetrically so, but V-shaped openings, with the margins overlapping or not, are occasionally found on the same plant as the more common U-shape (e.g. J.J. Bruhl 2976). The inflorescences of A. nitens are slender but usually more elongated (20–50 cm long). The perianth bristles are either all much longer than the fruit body (up to twice as long), 2.8–5.0 mm long, or with the outer three bristles occasionally shorter than the body. The bristles are more or less flat in cross-section towards the base, covered with more or less sparse longer, softer hairs. They are filiform near the apex, expanding gradually over their length to be relatively much broadened and flat for the lower third of their length above the short, smooth basal section, which is more obvious than in A. borbonica. The nutlet is ellipsoid to obovoid, with the stylebase thickened compared with the upper part of the style and approximately twice as long (up to ~0.7 mm long) as in A. borbonica. The nutlet is usually slightly larger than in A. borbonica but with similar reticulate surface (Fig. 1c).
These differences are slight but appear to hold for most specimens seen. However, there are a few slender specimens that resemble A. borbonica but have bristle and nutlet features similar to those in A. nitens (e.g. Bruhl 2979), and conversely a few coarser specimens similar to A. nitens but with bristle and nutlet features similar to those in A. borbonica (e.g. some of the five plants collected as Bruhl 2953a‒e from a population on the plateau at Saint Joseph, Foc-Foc, SW of Piton de la Fournaise). Further investigation is needed to clarify whether this indicates hybridisation between the two species or that there is a single variable species on the island.
Both type sheets bear labels in Steudel’s script. The lectotype bears the usual ‘Herbarium Steudel’ label. The isolectotype has a label bearing a manuscript diagnosis in Steudel’s script.
Of limited distribution but occurs in national parks; we suggest a status of ‘Vulnerable’ (IUCN Standards and Petitions Committee 2022), pending further investigation of its taxonomic status, and considering that the high-altitude habitat is limited and predicted to become even more so with changing climate.
Réunion: Bourbon, [no date; received at Kew Oct. 1875], I.B. Balfour s.n. (K, fragment B 10 0000966 p.p.); overlooking Piton de la Fournaise, Nez Coupé de Sainte Rose, 1942 m, 14 Jan. 2011, J.J. Bruhl 2977, J. Hivert & S. Pichillou (BOL, BRI, K, MO, NE 104059, NSW, P, REU); Grand Bénard, environs du Maido, 2100 m, 22 Nov. 1968, T. Cadet 1461 (P 00592876); Plaine des Salazes, 2200–2400 m, 14 Feb. 1969, T. Cadet 1953 (P 592875); Ile de la Réunion, 21 July1875, G. de l’Isle 410 bis (K, P 00592874); Plaine des Caffres, Ile Bourbon, 1837, J.M.C. Richard 660 (NSW 538940).
Asterochaete capitellata Nees, Linnaea 10: 194 (1835)
Carpha capitellata (Nees) Boeckeler, Linnaea 38: 266 (1874). Type citation: ‘In Cap. b. Sp. Forbes in Herb. Lindleyii; e districtu Zwellendam Herb. Eckl.’ Type: South Africa: Cape of Good Hope, [no date], [J.?] Forbes in Herb. Lindley (lecto, here designated: CGE 00003).
Asterochaete angustifolia Nees, Linnaea 9: 300 (1834), nom. inval., nom. nud.
Asterochaete capitellata Nees, Linnaea 9: 300 (1834), nom. inval., nom. nud.
Asterochaete glomerata var. β minor Nees, Linnaea 10: 194 (1835). Type citation: ‘Ad ‘waterfall’ [prope ‘Tulbagh’, alt. II-V (Worcester)] cum forma communi.’ [Ecklon]. Type: n.v.
Asterochaete tenuis Kunth, Enum. Pl. 2: 312 (1837). Type citation: ‘Cap. b. Spei.’ [= Caput Bonae Spei]. Type: South Africa: Cape of Good Hope: numerous probable syntypes seen (see Nomenclatural Notes below).
Asterochaete ludwigii Hochst. in F. Krauss, Flora 28: 759‒760 (1845) in footnote. Type citation: ‘Inter plantas capenses exsiccatas a Barone de Ludwig anno 1837 benigne communicatas e genere Asterochaete Nees novam speciem inveni.’ Type: South Africa: Cape of Good Hope, [no date], C.F.H. von Ludwig s.n. [presented to Hochstetter in 1837] (holo (2 sheets): TUB-007204, TUB-007205).
Carpha bracteosa C.B.Clarke [in T.A. Durand and H. Schinz, Consp. Fl. Afr. 5: 656 (1894), nom. nud.], in W.T. Thiselton-Dyer, Fl. Capensis 7: 270 (1898). Carpha capitellata var. bracteosa (C.B.Clarke) Kük., Feddes Repert. 47: 117 (1939). Type citation: ‘Coast Region: Worcester Division; in the valley of the Breede River, near Bains Kloof, 800 ft [~240 m], Bolus, 2867! Central Region: Somerset Division; marshy places on the summit of Bosch Berg, 4500 ft [~1370 m], MacOwan, 1616! 2187!.’ Type: South Africa: Cape Province: in valle fluvii Breede prope ‘Bains’s Kloof’, Worcester Division, ~800 ft [~240 m], Oct. 1873, H. Bolus 2867 (lecto, here designated: K (Zhang et al. 2006, fig. 13).
Ficinia trinkleriana H.Pfeiff., Herbarium no. 54: 34 (1920). Type citation: ‘In paludosis pr. Mitchels Pass (Schlechter 8970!).’ Type: South Africa: Mitchels Pass, 1000 ft [~300 m], Worcester District, Cape Province, 11 Sep. 1896, R. Schlechter 8970 (syn: B 10 0000974 (fragment), BR 24800877*, ?BREM, n.v., K, NSW 387363, P 003471828, P 003471830, PRE 0101841-0, US00680245, Z 80373).
Carpha perrieri Cherm., Bull. Soc. Bot. France 69: 721 (1922). Type citation: ‘Massif d’Andringitra (Perrier de la Bâthie, 14555).’ Type: Madagascar: Massif de l’Andringitra, audessus de 2500 m, Feb. 1922, H. Perrier de la Bâthie 14555 (lecto, here chosen: P 00199383; probable isolecto: B 10 0000969, G n.v., K, MO 176397*, P 00199384).
Erect perennial, 20–50(–70) cm high; rhizome short to long. Culms 1- or 2-noded, each node with a single leaf, or unnoded. Leaves crowded at the base of the culms; blades straight, 13–36 cm long, 1.3‒4 mm wide, V-shaped to U-shaped in cross-section, with midrib obvious abaxially (rarely not obvious, e.g. in Reid 1807), scabrous only near the apex; leaf sheaths pale yellow-brown to darker orange-brown. Inflorescence 5–25(–35) cm long, with 2–5 short to long internodes; spikelets ~50–100, in a few (2–8) dense ovoid clusters. Involucral bracts several, occasionally strongly broadened at base, up to 25 cm long, 3‒7 mm wide across broadened base, shorter or slightly longer than the spikelet clusters. Spikelets 5–7.5 mm long, with 1 or 2 bisexual flowers; glumes 5 (rarely 4), pale yellow-brown to dark yellow-brown; lowest 3 (rarely 2) glumes empty, persistent; the upper 2 usually fertile (but the uppermost one sometimes sterile), deciduous; lowest glumes shorter than the rest; the proximal fertile glume 4.4–6.2 mm long, 1.5–2.5 mm wide; the upper fertile glume 4.2–6.5 mm long, 1–2.3 mm wide; axis elongated above proximal fertile glume and adnate to the base of that glume. Bristles scabrous over whole length or nearly smooth at base; outer 3 bristles shorter (rarely only slightly so) than inner 3; the inner three 1.5–3.5 mm long, shorter than to slightly exceeding nutlet body (the length varies within a whorl). Anthers greyish, 1.8–2.9 mm long, with apical appendage 0.1–0.3 mm long, ~0.1 mm wide. Stylebase 0.3–1.0 mm long, 0.15– 0.3 mm in diameter, hispidulous with hairs often also on the upper ribs on the nutlet. Nutlets 1 or 2 per spikelet, narrow-obovoid to narrow-ellipsoid, 2.0–2.7 mm long, 0.8–1.4 mm in diameter, mid- to dark brown at maturity, surface reticulate, occasionally puncticulate (Fig. 1d, e; also Zhang et al. 2004b, fig. 4d, e as C. bracteosa, 5a, 7e, k, for spikelet structure; Zhang et al. 2006, fig. 3a; Clarke 1904, pl. 10 as ‘C. bracteosa’; Clarke 1909, t. 77, fig. 2; Chermezon 1937, fig. XXI 5–7 as ‘C. perrieri’).
At high altitude in Zimbabwe (only one record, at ~1900 m in the Mutasa District) and Madagascar (1200–2600 m from several massifs); at a wide range of altitudes (180–1500 m) in southern Africa, from KwaZulu-Natal to the Western Cape Province (Fig. 4b). In marshy places.
Asterochaete capitellata is a variable, widespread species and needs more detailed study. Clare Archer (née Reid) noted on some specimens in NU (e.g. Robinson 1976 from Zimbabwe and Getliffe, Hearne & Kerfoot 56 from the Transvaal) that these more northern specimens of this species have three of the perianth bristles much longer than the other three, whereas in Cape specimens the bristles are more or less equal in length. We have not found this to be a consistent difference in the material examined.
The variation in plant size and in glume colour over the geographic range of this species needs to be examined more closely too. The sole Zimbabwean specimen seen, Robinson 1976, is particularly slender, especially its inflorescence branches (Browning et al. 2020, fig. 14.64). Two slender specimens from KwaZulu-Natal (Linder 5516, BOL 60368) and Western Cape (Esterhuysen 18381, K) have less dense spikelet clusters and are generally less robust and rigid, but the fruit and bristles are similar to those of A. capitellata. Molecular study of populations will probably be needed to understand all of the variation in this species.
One obvious variant with very broad bases to the inflorescence bracts, found in the Cape Province of South Africa, was described as C. bracteosa by Clarke. Plants of this form can be found growing with more typical plants, e.g. Bruhl 1725 (broad bases) and 1726 (typical), and can be distinguished by their bracts and somewhat more robust appearance.
Carpha bracteosa was treated as a variety of C. capitellata by Kükenthal (1939a) and later as conspecific with C. capitellata by Levyns (1950) and Archer (2000), whereas other authors (Clarke 1894, 1898; Reid and Arnold 1984; Browning and Guthrie 1994) have recognised it as a distinct species. Archer (2000) treated C. bracteosa as a synonym of A. capitellata. In our phenetic analyses (Zhang 2002; Zhang et al. 2006), the specimens of C. bracteosa and C. capitellata always formed a group, but with the specimens of C. bracteosa consistently forming a subgroup. These results suggest that C. bracteosa is only slightly different from typical C. capitellata, differing in having involucral bracts that are ovate towards the base rather than leaf-like bracts that are only slightly broader near the base (Zhang et al. 2006, fig. 13). These two forms are not isolated geographically and apparently have the same habitat in the Cape Province. No other character variation seems to correlate with the broad bracts of C. bracteosa, so it seems unwise to recognise this variant formally before further investigation. Robust plants are found also in Madagascar.
Carpha perrieri was described by Chermezon (1922) from Madagascar. He regarded it as close to C. capitellata but differing from that species by its slightly bigger glumes, its longer perianth bristles, its larger stylebase, and its narrower nutlet. We found that glume, perianth bristle and stylebase length and nutlet diameter of C. perrieri overlap with those of typical C. capitellata (Zhang et al. 2006, table 7). Specimens of the two species grouped together in the phenetic analyses, and C. perrieri does not have any unique characters to distinguish it from C. capitellata; therefore C. perrieri is here included in C. capitellata.
The CGE sheet of Asterochaete capitellata is the only sheet seen of the Forbes and Ecklon syntypes mentioned in the protologue and is here selected as the lectotype. The specimen consists of a complete tuft with leaves, inflorescence and a young rhizomatous shoot at the base. It has been annotated by Nees as ‘Asterochaete capitellata N ab E’. Ecklon’s specimens are found in numerous herbaria, but we have seen no specimen of this species labelled (by a code number, as detailed by Drège 1847; also Glenn and Germishuizen 2010) as coming from the Zwellendam (now Swellendam) district.
We have not seen any specimen recognisable as a type of Asterochaete glomerata var. β minor, but from the protologue description, this is probably merely a smaller plant of A. capitellata. Both Clarke (1898, p. 270) and Kükenthal (1939a, p. 116) cited this name as a synonym of A. capitellata. At the same time, Kükenthal cited an Ecklon specimen from ‘Waterfall bei Tulbagh’ as Carpha capitellata var. β bracteosa, so the Nees varietal name may apply to this form of A. capitellata with broad involucral bracts (see below).
Typification of Asterochaete tenuis is problematic. Kunth’s private herbarium ended up in B (Stafleu and Cowan 1979, p. 693; Hiepko 1987) but at least part of it seems to have been lost in World War II, and we have not seen any material of this species that is obviously recognisable as coming from Kunth’s herbarium or has been annotated by Kunth. We are not choosing a lectotype because further study could find authentic material from Kunth’s herbarium and meantime there is no ambiguity about the taxon to which the probable syntypes belong. There is a fragmentary specimen (loose glumes and nuts) of this taxon in B, Drège 2449, (‘zum Elefant rivier’, 10 Nov 1820) B10 0000968a, mounted on a sheet with fragments (a spikelet cluster and loose glumes and nutlets) from a more recent specimen of this species, Meebold 11724. This Drège material may have come from Kunth’s herbarium, but the label on the fragment was written by Kükenthal, so it is impossible to tell. He does not cite this specimen in his paper (Kükenthal 1939a). We have seen various sheets of C. F. Drège collections of this species, some numbered (e.g. Drège 1840 (in K) and 2031 (P 00461843) from Zuurberg Range, Alexandria Division, 2000–3000 ft (~600–900 m), and Drège 2449 (P00461845 and the fragmentary specimen B10 0000968) from Rooderberg 4000–5000 ft (~1200–1500 m) and some sheets without collection numbers, e.g. in K (2 sheets) and P 00461844. The latter sheets, along with the sheet of Drège 1840 in K, have the name ‘Asterochaete tenuis Kunth a’ written on them. Note that Drège codified his collecting localities by the letters a, b, c, etc. following the taxon name; see the full list of his collecting localities in South Africa in Drège (1843).
A Drège specimen in K (Zuurberg Range, Alexandria Division, 2000–3000 ft, ~609–914 m, C.F. Drège 1840, no date, stamped as coming from Bentham’s herbarium) has been annotated by C. B. Clarke as ‘Carpha capitellata forma tenuis; this differs from type capitellata in having 1 nut only’. Contrary to this statement, we found one or two nutlets per spikelet in the specimens of Drège 1840.
Asterochaete ludwigii has sometimes been regarded as a nomen nudum but the description in a long footnote in Krauss (1845) is more than adequate for valid publication. Krauss attributes the name and description to Hochstetter.
The lectotypification of the name Carpha bracteosa effected here preserves the traditional usage of that name. When Clarke (1898) published his new species C. bracteosa, he cited three specimens (Bolus 2867, MacOwan 1616 and 2187), all with involucral bracts broadened towards the base. Of the three syntypes, only Bolus 2867 belongs to this taxon as traditionally recognised and, therefore, Bolus 2867 is selected as lectotype here to maintain the application of the name. The other two syntypes, MacOwan 1616 and 2187 (Zhang et al. 2006, fig. 3b, 14), are considered to belong to the new species Asterochaete acuminata. C. capitellata differs from A. acuminata in having leaves with a keel, V-shaped in cross-section, involucral bracts more or less equal in length to the inflorescence, spikelets in ovoid clusters, and the nutlet surface reticulate.
Hans Heinrich Pfeiffer worked in Bremen for part of his career and his herbarium is at least partly in BREM, as are some of Schlechter’s African collections (Metzing 1999) but no Schlechter 8970 sheet is present in BREM (Michaela Grein, pers. comm.). We are not choosing a lectotype for Ficinia trinkleriana because it is possible that a specimen annotated by Pfeiffer will be found and, furthermore, all the syntypes and protologue agree as to which taxon they represent so there is no ambiguity in the application of the name. The type collection belongs to the broad-based form of A. capitellata.
There are two sheets in P of Perrier de la Bâthie 14555 with the same collection details and labelled as Carpha perrieri; one is from Chermezon’s herbarium (P 00199383), the other (P 00199384) has a general Paris herbarium label. Both bear adequate specimens of this same taxon, but neither has been annotated by Chermezon. The sheet from Chermezon’s herbarium has the more obvious connection with him and is therefore chosen as the lectotype of Carpha perrieri. None of the apparent replicates seen in other herbaria has been annotated by Chermezon. The sheets GOET 010743 and NSW 491742 of Perrier de la Bâthie 14555 bear the date February 1923, which may have been a slip of the pen. The material looks similar to that on the other sheets of this number dated February 1922, and may have been part of the same original collection.
Widespread in South Africa, including in national parks, so listed as ‘Least Concern’ (Contu et al. 2013, as Carpha capitellata). However, it has very restricted known distributions in Madagascar and Zimbabwe, so we suggest a status of ‘Data Deficient’ or possibly ‘Vulnerable’ (IUCN Standards and Petitions Committee 2022) for populations in those countries.
Madagascar: Massif de l’Andringitra (Iratsy), vallées de la Riambava et de l’Antsifotra et montagnes environnantes, 2000–2500 m, 27 Nov.–8 Dec. 1924, H. Humbert 3878 (B10 0000970, P 01880322, P 01948583); crête et rochers siliceux du sommet, Massif de l’Andohahelo, 1800–1979 m, 21–22 Oct. 1928, H. Humbert 6146928 (BM, BR 24800914*, K, P 00199390, P 00199391, P 00199393, US 1528711), 1200–1800 m, 18–26 Oct. 1928 (P 00199394), 1700–1850 m, 21–22 Oct. 1928 (B 10 0000972); Massif de Tsaratanana, 2300‒2800 m, Nov.–Dec. 1937, H. Humbert 18404 (P 01880323, P01880324, P 03471827); Massif de Tsaratanana, 2600 m, Nov. 1966, P. Morat 2307 (NSW 996706, P 01948582, P 00199389).
South Africa: Eastern Cape Province: Hopewell, ~600 ft [~180 m], Bathurst District, 30 Oct. 1964, J.P.H. Acocks 23507 (PRE); Zuurberg Range, Alexandria Division, [no date], C.F. Drège 1840 (K, two sheets); marshy places near Komgha, 2000 ft [~600 m], Sep. ?year [received at K Aug 1893], Komgha Division, H.G. Flanagan 920 (K); Humansdorp, 650 ft [~200 m], Oct. 1930, H.G. Fourcade 4476 (K); 7 miles [~11 km] E of Humansdorp along P.E. road, 21 Dec. 1943, H.G. Fourcade 6336 (BOL 63166); in humidis pr. [prope] Howisons Point, 1800 ft [~1490 m], [no date], P. MacOwan 351 (MEL 1543862).
Mpumalanga Province: Wakkerstroom, Oshoek District, ~1800 m, 3 Nov. 1979, N.J. Devenish 1821 (US); Mount Sheba Nature Reserve, 1960 m, Mpumalanga, Oct. 1975, F. Getliffe, [initial unknown] Hearne & O. Kerfoot 56 (NU). Lydenburg District, Steenkampsberg Pass near layby, 3 Dec. 1992, C. Reid 1807 (US). Western Cape Province: off R323, 200 m along Langkloof Road, 6 Dec. 1996, J.J. Bruhl 1718 & F.C. Quinn (BOL, CANB, GRA, K 486710, MO, NE 76468, NSW 599812, PRE); Franschhoek Pass, Route 45 between Villiersdorp and Franschhoek, 305 m, 7 Dec. 1996, J.J. Bruhl 1726 & F.C. Quinn (BOL, EIU, GENT, K, MO, NE 76340, NSW, NU, P, PRE); Worcester Division, SE slopes on Fonteintjietberg, 5500 ft [~1675 m], 26 Nov. 1944, E. Esterhuysen 10977 (BOL 63169); Kenilworth, Sep. 1938, M. R. Levyns 6846 (P 0371832); Cape Infanta, Oct. 1947, M.R. Levyns 8391 (K); Swartberg, 4900 ft [~1490 m], 7 Jan. 1975, M.F. Thompson 2282 (PRE).
Zimbabwe: Gairesi Ranch, 6 miles [~10 km] N of Troutbeck, Inyanga, 6200 ft [~1890 m], 21 Nov. 1956, E.A. Robinson 1976 (BR 24800884*, IRLCS n.v., K n.v., LISC n.v., NRGH n.v., NU, SRGH n.v.).
South Africa: Eastern Cape: valley N side of Outeniquas near Joubertina, Uniondale Division, 5 Nov. 1944, E. Esterhuysen 10611 (BOL 63167, PRE). Western Cape Province: Franschhoek Pass, Route 45 between Villiersdorp and Franschhoek, 305 m, 7 Dec. 1996, J.J. Bruhl 1725 & F.C. Quinn (BOL, CANB, GENT, K, MO, NE 76339, NSW 834199, NU); Du Toits Kloof, 19 Oct. 1947, M.R. Levyns 8624 (BOL 63165); Somerset West, Stellenbosch Division, 25 Oct. 1942, R.N. Parker 3744 (K); Jakkalsvlei, Jonkershoek, ~1300–2300 ft [~390–700 m], Stellenbosch District, 26 Sep. 1963, H.C. Taylor 5220 (K, PRE), ~1300 ft [~390 m], 14 Oct. 1964, H.C. Taylor 5988 (NU).
Asterochaete dactyloides (Vahl) K.L.Wilson & J.J.Bruhl, comb. nov.
Schoenus dactyloides Vahl, Enum. Pl. 2: 224 (1805). Chaetospora dactyloides (Vahl) Dietr., Spec. Pl. 6th edn, 2: 32 (1833). Type citation: ‘Habitat in Cap. b. spei.’
Type: South Africa: e Cap. b. spei [Cape of Good Hope], [no date], ded. Burman (holo: C10001535).
Schoenus glomeratus Thunb., Prodr. Fl. Capens. 1: 17 (1794), nom. illeg. [non S. glomeratus L. (1753)]; Carpha glomerata Nees, Linnaea 7: 529 (1832), nom. illeg.; Asterochaete glomerata Nees, Linnaea 10: 194 (1835).
Type: as for S. dactyloides Vahl.
Asterochaete glomerata Nees, Linnaea 9: 300 (1834), nom. inval., nom. nud.
Erect perennial, 1–3 m high; rhizome stout. Culms triquetrous to trigonous in cross-section, 1–3-noded, each node bearing a single leaf. Leaves crowded at the base of the culms; blades straight, 40–105 cm long, (7–)11–25 mm wide, V-shaped in cross-section, keeled abaxially, scabrous on keel near apex; sheaths pale yellow-brown. Inflorescence with 4–9 internodes; spikelets (300–)500–3000 in many (10–60) dense ovoid clusters. Involucral bracts several, lowest much shorter than inflorescence, 15–45 cm long, up to 22 mm wide. Spikelets 6.2–9 mm long, with 2 bisexual flowers; glumes 6, pale yellow-brown; lowest 3 glumes empty, persistent; the next 2 fertile, and the uppermost one empty, deciduous; the lowest 2 glumes shorter than the rest; the proximal fertile glume 5.8–7.5 mm long, 1.8–2.8 mm wide, the second fertile glume 5.0–7 mm long, 1.5–2 mm wide; axis elongated above proximal fertile glume and adnate to the base of that glume. Bristles: upper part (2/5–2/3) of each bristle terete, antrorsely scabrous, breaking off readily, lowest 1/3–3/5 much more slender, smooth; outer 3 bristles slightly shorter than inner 3; the inner 3 from slightly shorter than to slightly exceeding the nutlet body, 3.1–3.8 mm long. Anthers yellow when immature but pale greyish at maturity, 2.4–4.2 mm long (excluding appendage), with apical appendage 0.2–0.3 mm long, ~0.1 mm wide. Stylebase 0.3–1.0 mm long, 0.1–0.3 mm in diameter. Nutlets 2 per spikelet, narrow-ellipsoid to obovoid, 2.1–3.4 mm long, 0.9–1.1 mm in diameter, dark brown at maturity, surface finely reticulate (Fig. 1f; see Zhang et al. 2004b, fig. 7q for spikelet structure; Hooker 1883, pl. 1435; Clarke 1909, t. 76, fig. 7–9).
In southern parts of South Africa (KwaZulu-Natal and Eastern and Western Cape provinces) (Fig. 4c). In marshes, on stream banks and low-lying flats in near-coastal regions, at 90–500 m in altitude (Bruhl 1706 and Sonnenberg 387 seem to be at exceptionally high altitude for this species, at 1249–1500 m near Robertson Falls). This species was reported for Mount Uluguru in Tanzania by Haines and Lye (1983) and Hoenselaar et al. (2010), but that refers to the taxon here published as Asterochaete ulugurensis (q.v.).
This is the biggest species in the genus and has many more dense clusters of spikelets in the inflorescence than any other species. It resembles the somewhat smaller A. schlechteri, and the two species share a synapomorphy in their unusual perianth bristles, with the upper 2/3 or so of each bristle being terete and antrorsely scabrous, whereas the lower part of the bristle is much more slender and smooth. The upper part is readily broken off. Both species have a more elongated apical appendage on the anthers than in the other species.
There have been different views on how to distinguish A. dactyloides (universally referred to as C. glomerata or A. glomerata in past literature) and A. schlechteri, essentially based on the former being more robust than the latter. When Clarke (1904, p. 37) published his new species Carpha schlechteri, he described it as follows: ‘C. glomerata Nees affinis; differt: foliis angustioribus (vix 5 mm. latis); inflorescentia graciliore, spiculis minoribus; nuce grosse conspicue hexagono-reticulata levi (neque, ut in C. glomerata, minute obscure reticulata)’. He did not mention the actual size of the spikelet and nutlet of A. schlechteri in his description but he did say that the spikelets were smaller. Later, Clarke (1908) noted that A. schlechteri had leaves 4–5 mm wide, and nutlets 2–2.5 mm long. Examination of the type collection (R. Schlechter 10010) found the leaves to be 4.5–6 mm wide at their mid-length. Occasional specimens such as J. Browning 823 match the type and have nutlets 2.2–2.4 mm long but with leaves up to 8 mm wide.
Leaf width is a useful measure of the robustness of C. dactyloides, given that complete herbarium specimens are very rare because of the large size of the plants. The leaf width of A. dactyloides was first described by Boeckeler (1874, p. 265) as ‘7–12 lin. lat.’ (~14.8–25.4 mm wide), whereas Clarke (1898) described it as 1/4 inch (~6.4 mm) or more. Kükenthal (1939a) considered that A. dactyloides had leaves 5–12 mm wide, spikelets 6–8 mm long and nutlets 3.5 mm long, whereas A. schlechteri had leaves 4–5 mm wide, spikelets 4 mm long and nutlets 2.5 mm long. Levyns (1950) described the leaves of A. dactyloides as 2 cm wide, and Gordon-Gray (1995) as 15–28 mm wide. Reid and Arnold (1984) distinguished A. dactyloides by its more robust stature, and its leaves being 15–28 mm wide, whereas the leaves of A. schlechteri were 5–8 mm wide. Our examination of specimens found leaves to be usually 11‒25 mm wide (rarely some leaves as narrow as 7 mm wide) in A. dactyloides v. 4.5‒6 mm wide (rarely some leaves up to 8 mm wide) in A. schlechteri.
Our study also found spikelet characters useful for distinguishing A. dactyloides and A. schlechteri (Zhang 2002; Zhang et al. 2006). Asterochaete dactyloides has two bisexual flowers per spikelet, both of which mature a nutlet, and the axis is elongated above the proximal fertile glume, whereas A. schlechteri has spikelets with a proximal male flower, one bisexual flower, and the axis does not elongate above the proximal fertile glume. A. dactyloides has longer spikelets (6.2–9 mm long), longer inner-whorl bristles (3.1–3.8 mm long) and longer nutlets (2.5–3.4 mm long) than those of A. schlechteri (spikelets 4–5 mm long, inner-whorl bristles 2.3–3.4 mm long, nutlets 2.2–2.4 mm long).
Geographically speaking, A. dactyloides is much more widespread, occurring from near sea level to ~1400 m across south-eastern and southern South Africa, whereas A. schlechteri is found only at high altitude (900–1370 m) in the Koue Bokkeveld Mountains of the Western Cape Province. They are not known to grow together; however, further fieldwork is needed to investigate their distributions and habitats.
Vahl’s Schoenus dactyloides has been overlooked as the correct name for this species despite his providing a full description, probably because he cited S. glomeratus Thunb. as a synonym, and therefore his name has been incorrectly treated as illegitimate. However, Thunberg’s name (dating from 1794) is a later homonym of S. glomeratus L. (= Rhynchospora glomerata (L.) Vahl) from 1753. Vahl obviously recognised that two different taxa were involved because he included both Schoenus dactyloides and Rhynchospora glomerata in his publication (Vahl 1805, pp. 224 and 234 respectively). Under Art. 6.13 of the International Code of Nomenclature for algae, fungi, and plants (Turland et al. 2018), Vahl’s name could be treated either as publication of a new species or as a replacement name. We choose to treat this as a new species because the protologue description is clear and typification is clearcut. The holotype collection from the Cape of Good Hope from Burman’s herbarium that was given to Vahl is now in C as sheet C10001535. Carpha glomerata Nees is illegitimate because it cites the earlier name Schoenus dactyloides as a synonym.
Widespread, including in national parks; listed as ‘Least Concern’ (Foden and Potter 2010, as Carpha glomerata).
South Africa: Eastern Cape Province: 23 km from Uitenhage on road to Groendaldam, Oct. 1975, T.H. Arnold 1065 (PRE); Hogsback Range, N side of Robinsons Fall Creek, at top of falls, 1240 m, 1 Dec. 1996, J.J. Bruhl 1706 & F.C. Quinn (NE 76453, NU); along R102, N of Witelsbos, 210 m, 4 Dec. 1996, J.J. Bruhl 1711 & F.C. Quinn (K, MO, NE 76464A, NSW 6586779, NU); along R102, S of Witelsbos, 0.5 km N of Elands R crossing, 220 m, 4 Dec. 1996, J.J. Bruhl 1712 & F.C. Quinn (BOL, CANB, CHR, GENT, GRA, K, MO, NE 76460, NSW 599810, NU, PRE); Port Elizabeth to Storms River Road, past Humansdorp turn-off, 12 Feb. 1982, F. Getliffe 1142 (NU); Kabusie Forest, Stutterheim, 20 Oct 1980, O.M. Hilliard and B.L. Burtt 13204 (NU); 7 km uphill from Robertson Falls, 1500 m, 26 Nov. 1994, B. Sonnenberg 387 (NU); Witelsbosch SAFCOL Forests, 1 Feb. 1996, B. Sonnenberg 458 (NU). KwaZulu-Natal Province: Vernon Crookes Nature Reserve, 400‒500 m, Port Shepstone District, 1 Oct. 1989, J. Browning 228 (NU); Alexandra District, Station Dumisa, Farm Friedenau, Umgaye, 600 m, 22 June 1909, H. Rudatis 661 (NSW 996707, P 01948981, P 03471866, PRE n.v.); Farm Etheldale, adjacent Mtamvuna Forest Reserve, ~370 m, Port Shepstone District, 5 Sep. 1971, C.J. Ward 7196 (NU 54603, PRE n.v.). Western Cape Province: 32 km from Riversdale, 12 km from turnoff along road to Langekloof, Cape, Oct. 1975, T.H. Arnold 1041 (PRE); Umgegend von Hopefield, Weg nach Theefontein, Oct. 1885, F. Bachmann 866 (Z 80381*); Simonstown, Kogelberg Forest Reserve, junction of Louw and Palmiert Rivers, 300 ft [~90 m], Cape, 21 Nov. 1969, C. Boucher 911 (PRE); Devon Valley, Fransmanskraal Farm, 14 Nov 1978, C. Boucher 4091 (US); Verlorenvlei, Clanwilliam, 24 Jan. 1996, J. Browning 803 (NU); Franschhoek Pass, Route 45 between Villiersdorp and Franschhoek, 320 m, 7 Dec. 1996, J.J. Bruhl 1729 & F.C. Quinn (BOL, NE 76343, NSW 834200).
Asterochaete discolor T.H.Arnold ex X.F.Zhang, sp. nov.
Planta A. capitellatae Nees et A. dactyloidei in structura inflorescentiarum, spicularum et florum simulans, sed in characteribus foliorum, squamarum coloribus et setis hypogynis differt. Folia trigona, conspicue crispa. Squamae discoloria. Setae hypogynae in dimidio distali antrorse scabrae, in dimidio proximali plumosae.
Type: South Africa: Western Cape Province: Worcester Div.: damp cliff face below shale band E side of Milner Peak, Hex River Mountains, alt. 4500 ft [~1370 m], 18 Dec 1948, E. Esterhuysen 14866 (holo: BOL 63171; iso: K (2 sheets)).
Tufted perennial ~35 cm high, with fibrous remains of old leaf sheaths at base; rhizome probably very short (complete plant bases not seen). Culms unnoded. Leaves basal, erect to spreading but curling as they age, thickly channelled or hemispherical to trigonous in cross-section, midrib not obvious below but more or less keeled towards apex, up to 20 cm long, 1.5–1.7 mm wide; sheaths mid- to dark yellow-brown. Inflorescence up to 15 cm long, with several (usually 2) nodes; composed of 2 or 3 ovoid to conical dense clusters of spikelets. Involucral bracts much shorter than inflorescence, up to 8 cm long, ~1 mm wide. Spikelets ~10 mm long, with 2 bisexual flowers; glumes 6, mixed in colour, mainly red-brown to dark red-brown in colour, but pale yellow-brown at base and with hyaline to whitish margins 0.2–0.3 mm wide; lowest 4 glumes empty, shorter than the fertile upper 2; proximal fertile glume 7.5–9.5 mm long, 1.7–2 mm wide; upper fertile glume 8.0–9.0 mm long, 1.2–2 mm wide; axis not elongated above proximal fertile glume. Bristles (slightly immature) lower half (up to 2/3) plumose with soft hairs up to 0.5 mm long, upper part antrorsely scabrous; outer 3 bristles more slender and shorter than inner 3; inner 3 9.0–11.5 mm long. Anthers pale yellow to pale yellow-grey at maturity, 3.0–4.5 mm long (excluding appendage), with apical appendage linear-acuminate, 0.3–0.8 mm long, ~0.1 mm wide. Stylebase (immature) ~0.3 mm long, glabrous. Nutlet (only seen immature) trigonous in cross-section, narrow-ellipsoid, probably pale grey-brown, with paler longitudinal ribs (Fig. 1g, 6).
Holotype of Asterochaete discolor (E. Esterhuysen 14866, BOL). Reproduced with permission of Bolus Herbarium, University of Cape Town.

Known only from one collection from a damp cliff face below a shale band on the eastern side of Milner Peak at 1370 m in altitude, in the Western Cape Province of South Africa (Fig. 4a).
The specific epithet is from the manuscript name written on the two isotype sheets in K by South African botanist Trevor H. Arnold (1947‒). It reflects the conspicuous mixed colours of the glumes, one of the key distinguishing features of this species.
This species resembles Asterochaete capitellata and A. dactyloides in the structure of the inflorescence, spikelets and flowers, but differs in its leaves, which are thickened, channelled and more or less trigonous in cross-section, the fibrous old leaf bases around the tufts, the conspicuously coloured glumes, and the perianth bristles, which are plumose in the lower half (similar to those of Carpha sens. str. but the hairs are shorter and less dense), and antrorsely scabrous in the upper half. The leaves and longer involucral bracts become more or less uncinate with age and curl somewhat over their whole length. Its distribution overlaps with that of A. schlechteri but the species are not known to be sympatric. It differs in many features from A. schlechteri, including stature (A. discolor much more slender and smaller), leaf lamina cross-section (thick and channelled to hemispherical to trigonous in A. discolor; V-shaped in cross-section in A. schlechteri); inflorescence with one or two nodes in A. discolor; usually many-noded in A. schlechteri); spikelets length (~10 mm long in A. discolor; 4–5 mm long in A. schlechteri); perianth and nutlet morphology (cf. Fig. 1g, k).
Known only from the type locality; we suggest a status of ‘Data Deficient’ or possibly ‘Vulnerable’ (IUCN Standards and Petitions Committee 2022).
Asterochaete eminii (K.Schum.) X.F.Zhang, comb. nov.
Oreograstis eminii K.Schum. in A. Engler (ed.), Pflanzenw. Ost-Afrikas C, 127 (1895); Carpha eminii (K.Schum.) C.B.Clarke in W.T. Thiselton-Dyer, Fl. Trop. Afr. 8: 483 (1902). Type citation: ‘R. bei 3100 m. – Stuhlm. 2439’. Type: Uganda: Ra Nssoro [Rwenzori Mountains], 3100 m, 13 July 1891, Dr Stuhlmann 2439 (syn: B 10 0000975 (fragments of spikelets).
Tufted perennial, 45–110 cm high; rhizome short. Culms erect to spreading, with 1 or 2 nodes, each bearing a single leaf, or unnoded. Leaves crowded at the base of the culms; blades more or less straight, 20–75 cm long, 2.0–4.0 mm wide, V-shaped (narrowly to nearly flat) in cross-section, midrib obvious below, glabrous, strongly keeled and scabrous towards apex; sheaths pale yellow-brown above to dark orange-brown below. Inflorescence with 3–6 internodes, 10–60 cm long; spikelets 27–130 loosely clustered. Involucral bracts several, up to 70 cm long, to 4 mm wide. Spikelets 6–8 mm long, with 1 bisexual flower; glumes 4 or 5, pale to dark yellow-brown; lowest 3 glumes empty, persistent, the next one fertile, deciduous, the uppermost one sterile when spikelet has 5 glumes; lowest 2 glumes shorter than the rest; the fertile one 5.5–7.5 mm long, 1–1.5 mm wide; axis not elongated above fertile glumes. Bristles: outer 3 bristles as long as or exceeding the nutlet body, much shorter than inner 3, antrorsely scabrous; the inner 3 3.5–6.7 mm long, exceeding the nutlet body (to twice as long); lower part of bristles in inner whorl compressed and with slightly longer spreading hairs along the margins but shorter again right at base, upper half antrorsely scabrous. Anthers greyish to black, 2.8–4.0 mm long (excluding appendage), with apical appendage 0.1–0.2 mm long, 0.1–0.15 mm wide. Stylebase 0.3–2.3 mm long, 0.15–0.2 mm in diameter, glabrous or very sparsely hispidulous. Nutlet 1 per spikelet, narrow-ellipsoid, 2.8–3 mm long, 0.8–0.9 mm in diameter, mid-grey-brown with paler longitudinal ribs at maturity, surface reticulate and puncticulate. (Fig. 1h; see Zhang et al. 2004b, fig. 7a, b for spikelet structure; Napper 1964, fig. 17; Haines and Lye 1983, fig. 662).
Endemic to the Rwenzori (formerly Ruwenzori) Mountains straddling the border of the Western Province of Uganda and the Democratic Republic of Congo (Fig. 4b). In mountain bogs and on stream banks at high altitude (~2700‒3750 m).
Kükenthal (1939b) treated Carpha angustissima as a variety of A. eminii, and this was followed by Napper (1964), but our analyses (Zhang 2002; Zhang et al. 2006) indicated that it should be maintained as a separate species, in agreement with Chermezon (1935) and Haines and Lye (1983).
These two species are allopatric; Asterochaete eminii is found on the Rwenzori Mountains (west of Kampala) in western Uganda and A. angustissima on the Mgahinga Mountains in south-western Uganda. A. eminii differs from A. angustissima in being slightly more robust and by usually having 1-flowered spikelets with the axis not elongated above the fertile glumes and not adnate to the fertile glume base, and the glumes pale to dark yellow-brown; the leaf blades V-shaped to nearly flat in cross-section, 2.0–4.0 mm wide; and longer perianth bristles (the inner whorl 3.5–6.7 mm long). A. angustissima has two flowers per spikelet with the axis elongated above the fertile glumes and adnate to the fertile glume base, and the glumes pale yellow-brown; the leaf blades thinly crescentiform in cross-section, 0.7–1.6 mm wide; and shorter perianth bristles (the inner whorl (2.0–)3–4.4 mm long).
The third African tropical montane species is A. ulugurensis (q.v.), found in the Uluguru Mountains of the Eastern Arc in Tanzania.
The fragments (glumes, anthers and an immature nutlet with associated bristles much longer than the fruit) in B 10 0000975 were apparently removed by Kükenthal from the main type specimen for examination during his work on this group. The label of this sheet is in Kükenthal’s script. This material is identifiable as A. eminii. The main specimen was probably destroyed in B during World War II. A complete duplicate of Stuhlmann 2439 may yet be found in another herbarium, so we refrain from designating the fragments in B as lectotype.
Of very restricted distribution but occurs in conservation reserves; listed as ‘Vulnerable’ (Luke 2010) but ‘Endangered’ may be more appropriate given that the high-altitude habitat is limited and predicted to become even more so with changing climate and increasing human impact.
Uganda: Bujuka, Mount Ruwenzori, 11,500 ft [~3500 m], Aug. 1931, C.W.L. Fishlock & G.L.R. Hancock 109 (K); Mount Ruwenzori, near Nyamileju Hut, 10,500 ft [~3200 m], 30 Dec. 1968, R.W. Haines 277 (BR 13662936*, K); Bujuku Valley, near Bigo camp, Mt Ruwenzori, 3400 m, 24 Mar. 1948, O. Hedberg 435 (EA, K, LD); Mijusi Valley, Mount Ruwenzori, 3850 m, 30 Mar. 1948, O. Hedberg 598 (EA, K); Kasese, Busongora County, above Bigo camp, 3050 m, 21 Jan. 1981, A. Katende 3003 (BR 13662929*, MO, PRE, US 3398837); Muteinda, Mount Ruwenzori, 11,900 ft [~3625 m], 25 Dec. 1961, J.P. Loveridge 120 (K); near Nyamileju (2nd) Hut, Mount Ruwenzori, 3300 m, 30 Dec. 1968, K.A. Lye 1249 (EA); above Kichucha, Mount Ruwenzori, 3000 m, 31 July 1953, H.O. Osmaston 3210 (K); Butahu Valley, Kitandara, Mount Ruwenzori, 3500 m, Aug. 1953, H.O. Osmaston 3788 (13662912*, K); Mount Ruwenzori, 12,500 ft [~3810 m], Aug. 1938, J.W. Purseglove P270 (K); Mount Ruwenzori, 12,500 ft [~3810 m], 21 Feb. 1906, A.F.R. Wollaston (BM 000092183).
Democratic Republic of Congo [sic on label]: affluent de la Mososa (Cascade), E de Mahungu (Ruwenzori), 3180 m, 23 May 1953, G.F. De Witte 9104 (BR 13662950, K, P 01724486).
Asterochaete filifolia (C.Reid & T.H.Arnold) X.F.Zhang, comb. nov.
Carpha filifolia C.Reid & T.H.Arnold, Bothalia 15(1&2): 139, fig. 2a–h, 3, 4a (1984). Type citation: ‘Natal, 2929 (Underberg): ‘Storm Heights’, Mpendhle District (–BC), Hilliard & Burtt 11791 (PRE, holo; E; NU)’. Type: South Africa: KwaZulu-Natal Province: Mpendhle District, ‘Storm Heights’, ~7000 ft [~2130 m], 16 Dec. 1978, O.M. Hilliard & B.L. Burtt 11791 (holo: PRE 0649543-0*; isotypes: E n.v., MO 1515484*, NH 191146-0*, NU 0016081-0*).
Tufted perennial, 30–70 cm high; rhizome absent or very short. Culms unnoded or rarely 1-noded and bearing a single cauline leaf. Leaves crowded at the base of the culms; blades filiform, 17–40 cm long, 0.4–0.7 mm in diameter, thickly crescentiform (without abaxial keel or midrib) or more or less circular in cross-section; sheaths mid- to dark yellow-brown, often shining. Inflorescence with 1 or 2 very short internodes, 2–6 cm long; 9–25 spikelets in 1 very dense narrow obconical to oblong head, rarely in 2 clearly separated heads. Involucral bracts several, lowest usually 1.5–3 times longer than the inflorescence, rarely approximately as long as inflorescence, 3–11 cm long, to 2 mm wide. Spikelets 8–10 mm long, with 2 bisexual flowers; glumes 4 or 5, pale to mid-yellow-brown; lowest 2 or 3 glumes empty, persistent, the upper 2 fertile, deciduous; lowest 1 or 2 glumes shorter than the rest; the proximal fertile glume 7.2–9.5 mm long, 1.4–2.4 mm wide, the second fertile glume 4.3–8 mm long, 1–1.3 mm wide; axis elongated above proximal fertile glume and adnate to the base of that glume. Bristles filiform near apex, terete below; outer 3 bristles slightly shorter than to slightly exceeding the nutlet body, shorter than the inner 3, antrorsely scabrous with a few longer hairs at very base; the inner 3 usually longer than the nutlet body (up to 1.5 times as long), (3.0–)4.0–4.5(–7.8) mm long, sparsely to densely hispid (up to 0.2 mm long) in lower part (usually only at base, very rarely for 1/3 of length) and the upper part antrorsely scabrous. Anthers greyish, 2.7–4.0 mm long (excluding appendage), with apical appendage 0.1–0.3 mm long, 0.1 mm wide. Stylebase 0.5–1.7 mm long, 0.15–0.4 mm in diameter, hispidulous with hairs extending to ribs on apex of nutlet. Nutlets 2 per spikelet, narrow-ellipsoid, 3.3–4.3 mm long, 0.8–1.1 mm in diameter, mid- to dark brown with paler longitudinal ribs at maturity, surface reticulate and often puncticulate. (Fig. 1i; see Zhang et al. 2004b, fig. 7k for spikelet structure; Hilliard and Burtt 1987, pl. 9, fig. C; Gordon-Gray 1995, fig. 16, 17A).
Restricted to the sandstone and basalt formations of the upper Karoo System in the Drakensberg Mountains of South Africa (Eastern Cape, KwaZulu-Natal, Free State and Mpumalanga provinces) and Lesotho (Fig. 4d; Reid and Arnold (1984, fig. 1). In marshes and along stream banks at high altitude (1800–2800 m) in the Drakensberg Grassland (Bastervoetpad) (Archer 2019). Co-dominant with Bulbostylis schoenoides (Kunth) C.B.Clarke in one community (Brand et al. 2013).
The features of this species are compared with the other South African species by Reid and Arnold (1984), and a key is given there. This species differs from other South African species in being tufted and having the following features: unnoded or single-noded culms; inflorescence of one very dense obconical cluster (rarely two clusters) of spikelets; filiform leaves (0.5–0.8 v. 1.5–24 mm wide in other southern African species), and a longer nutlet (3.3–4.3 v. 2.0–3.5 mm long in other southern African species). The overall colour of plants of this species is distinctively golden brown, particularly the rather shiny or glossy leaf sheaths. Some tussocks show obvious signs of regenerating after fire.
Its glumes are distichous to subdistichous in their arrangement rather than spiral as Reid and Arnold (1984) indicated, and spikelets have two bisexual flowers without an empty uppermost glume (the uppermost glume, described by Reid and Arnold (1984) as sometimes subtending a male flower or empty, was not found by Browning and Guthrie (1994) – also not found in this study). The specimen Devenish 1067 is unusual in the inner whorl of bristles being short-plumose for ~1/3 of their length, rather than only near the base of these bristles.
Although we did not examine the holotype, its image is on Global Plants JSTOR, the protologue has a clear illustration, and we examined other specimens cited in the protologue, such as Devenish 1067 and 1821, Hilliard & Burtt 9788, 13462, 16258 and 16677, Hoener 2138, and Killick & Vahrmeijer 3595.
Widespread, including in national parks, so not considered to be at risk: listed as ‘Least Concern’ (Raimondo et al. 2009, as Carpha filifolia).
South Africa: Eastern Cape: Elliot–Maclear district boundary, Bastervoetpad, ~7200 ft [~2195 m], 15 Feb. 1983, O.M. Hilliard & B.L. Burtt 16677 (K, NU 3500285). KwaZulu-Natal: Giants Castle Nature Reserve, Drakensberg Mountains, near top of Giants Ridge track, 2200 m, 24 Nov. 1996, J.J. Bruhl 1701 & F.C. Quinn (CANB, K, MO, NE 76458, NSW 599809, NU); path from Loteni Nature Reserve to Redi, Mpendhle District, 7700 ft [~2345 m], Sani Pass, ~8000 ft [~2435 m], Underberg District., 22 Mar. 1977, O.M. Hilliard & B.L. Burtt 9788 (K, 2 sheets, NU 3500280, E, P 03471846, PRE); Cobham Forest Reserve, ~6800 ft [~2070 m], Underberg District, 16 Feb. 1979, O.M. Hilliard 12509 & B.L. Burtt (K, NU 3500282, PRE 0557011-0); path from Loteni Nature Reserve to Redi, 26 Dec. 1982, O.M. Hilliard & B.L. Burtt 16098 (E n.v., K, P); Highmoor Forest Reserve, SE of Giants Castle,7700 ft [~2345 m], Mpendhle District, 6 Jan. 1983, O.M. Hilliard & B.L. Burtt 16258 (E n.v., K, NU 3500279); vicinity of Tarn Cavek above Bushman’s Nek, ~8000 ft [~2435 m], Underberg District, 20 Nov. 1983, O.M. Hilliard & B.L. Burtt 16813 (E n.v., K, PRE n.v.); vicinity of Tarn Cavek above Bushman’s Nek, ~8000 ft [~2435 m], Underberg District, 22 Jan. 1984, O.M. Hilliard & B.L. Burtt 17489 (E n.v., K); Little Berg, Highmoor Forest Station, 6500 ft [~1980 m], 14 Jan. 1966, D.J.B. Killick & J. Varhmeijer 3595 (PRE). Mpumalanga Province: Wakkerstroom, Oshoek District, 6600 ft [~2010 m], 14 Nov. 1963, N.J. Devenish 1067 (K, PRE); Wakkerstroom, Oshoek District, ~1800 m, 3 Nov. 1979, N.J. Devenish 1821 (K, PRE n.v.).
Lesotho: Base of Matadeng Pass, Sehlabathebe National Park, 25 Jan. 1995, J. Browning 696 (NE 87962, NU 3500299); between Pocha and Tsoelikana River, ~2300 m, Sehlabathebe National Park, 10 Jan. 1979, F.K. Hoener 2138 (K, NU 23103-0*, PRE); Sehlabathebe National Park, 5 Jan. 1990, C. Schwabe 0171 (K – 2 sheets, NU 3500296).
Asterochaete nitens Kunth, Enum. Pl. 2: 313 (1837) (as A. ?nitens)
Carpha nitens (Kunth) Kük., Feddes Repert. 47: 209 (1939). Type citation: ‘Schoenus nitens Willd. herb. n. 1119– Mauritius.’ Type: Mascarenes: in insula Mauritii, [no date], Aubert du Petit-Thouars in Herb. Willd. 1119 (holotype B–W*; probable isotype B 10 0000965 (fragments: a spikelet and some glumes and nuts with attached bristles), P 552879).
Carpha aubertii Nees, Linnaea 9: 300 (1834), nom. inval., nom. nud.
Carpha aubertii Nees ex Boeckeler, Linnaea 38: 270 (1874); Cyclocampe aubertii (Nees ex Boeckeler) Pax in H.G.A. Engler and K.A.E. Prantl (eds), Nat. Pflanzenfam. 2(2): 115 (1887). Type citation: ‘Schoenus nitens Willd. herb. n. 1119– Mauritius.’ Type: Mascarenes: in insula Mauritii, [no date], Aubert du Petit-Thouars in herb. Willd. 1119 (holotype B–W*; probable isotype B 10 0000965 (fragments: a spikelet and some glumes and nuts with attached bristles), P 552879, i.e. the same type as for Asterochaete nitens Kunth).
Rhynchospora boivinii Boeckeler, Linnaea 38: 403 (1874). Type citation: ‘Bourbon. Boivin. herb. no. 999.’ Type: Réunion: Brulé du Grand Bénard, Ile Bourbon, 16 Feb. 1847, L.H. Boivin 999 (lecto, here designated: P 00552910; possible isolecto: B 10 0000966 p.p. (fragments), P 00592871).
Tufted perennial, (20–)40–70(–130) cm high; rhizome short or not obvious. Culms (1–)2–5-noded, each node bearing a single leaf. Leaves crowded at the base of the culms; blades straight, 20–55 cm long, 2.5–5.0 mm wide, broadly V-shaped to nearly flat in cross-section, with scabrous keel towards apex abaxially and glabrous midrib below; sheaths pale yellow-brown with darker streaks. Inflorescence (10–)20–50 cm long, with 1–5 internodes 1–30 cm long; spikelets 50–125, in 2–10 oblong to ovate clusters 1–2 cm long, ~1 cm in diameter, on slender branches 1–20 cm long. Involucral bracts several, up to 27 cm long, up to 3.3 mm wide, shorter than the inflorescence. Spikelets 5–6.6 mm long, with 2 (rarely 1) bisexual flowers; glumes 5–7, yellow-brown to red-brown; lowest 3 or 4 glumes empty, persistent; the upper 2 fertile, deciduous, rarely only the upper one fertile; sometimes with an uppermost empty glume; lowest 2 or 3 glumes shorter than the rest; the proximal fertile glume 4.5–5.4 mm long, 1.4–2 mm wide, the second fertile glume 4.5–5.1 mm long, 1–1.3 mm wide; the axis elongated above proximal fertile glume and adnate to the base of that glume. Bristles filiform near apex, terete below and then conspicuously flattened near base, antrorsely scabrous over whole length (slightly longer, softer, sparse hairs near base); outer 3 bristles equalling to exceeding the nutlet body but still shorter than the inner 3; the inner 3 from slightly exceeding the nutlet body to twice as long as it, 2.8–5.0 mm long. Anthers greyish, 1.3–2.0 mm long (excluding appendage), with apical appendage ~0.1 mm long, ~0.1 mm wide. Stylebase 0.5–0.8 mm long, 0.1–0.2 mm in diameter, glabrous or sparsely hispidulous, hairs occasionally extending to apex of nutlet. Nutlets 2 per spikelet, rarely 1, ellipsoid to narrow-ellipsoid or obovoid, 1.5–1.8 mm long, ~0.7 mm in diameter, mid- to dark brown at maturity, surface coarsely reticulate (Fig. 1j; see Zhang et al. 2004b, fig. 7e, k, q for spikelet structure; Zhang et al. 2006, fig. 15).
Endemic to the island of Réunion in the Indian Ocean (Fig. 4b). Growing in dry low shrubland, in skeletal soils on laval rock pavements, and by roadsides on moist rock faces, at middle to high altitude (~1200–2300 m). The plants in the more exposed habitats are not as tall as those from more protected habitats but are similar in their leaves and other features.
This is one of two species growing on Réunion, both at high altitude. A. nitens differs most obviously from A. borbonica in having broader leaf blades that are keeled and broadly V-shaped to nearly flat in cross-section, a longer inflorescence, and with inner perianth bristles longer than in A. borbonica (2.8–5.0 v. 1.8–2.5 mm). See further discussion under A. borbonica, including the status of these taxa. Apparent morphological intermediates are listed below.
The type of Asterochaete nitens is labelled as coming from Mauritius (‘île Maurice’ in French) rather than Réunion, which, together with Rodrigues, are known collectively as the Mascarene Islands. However, this was presumably a mistake in labelling, as noted by Jean Raynal on the isotype in P, which is labelled ‘Ins. Borbon’ and ‘Ile de Bourbon’ (the old name for Réunion). The type is very much more likely to have come from Réunion, which has peaks rising to 3000 m in altitude, whereas Mauritius is a much lower island, with its highest peaks reaching only 800 m in altitude. All other known specimens come from high-altitude areas on Réunion.
The probable isotype in P is labelled as coming from the ‘Herbier Petit-Thouars’ but with the locality given as ‘Ile de Bourbon’ (=Réunion). An unknown person has pencilled ‘Carpha aubertii’ on the sheet, and Jean Raynal annotated it ‘Double de l’échantillon Asterochaete ?nitens hb. Willdenow 1119 qui porte l’indication ‘Maurice’ érronée’. That is, Raynal believed that the specimen had not come from Mauritius. The fragments in B were apparently removed by Kükenthal from the parent specimen for examination during his work on this group. The label on the B 10 0000965 sheet is in Kükenthal’s script; it states that the fragments were from du Petit-Thouars’ herbarium, suggesting the sheet in P was the source.
Some have considered that Hooker (1883, in Icon. Pl. 15: t. 1434) made a new combination Cyclocampe nitens (Kunth) Hook.f., but his note discussing the relationships of Costularia arundinacea stated that he had not seen Asterochaete nitens and merely ‘assumed’ (his word) that it belonged in Cyclocampe. This does not seem adequate for a new combination.
The lectotype sheet of Rhynchospora boivinii in P was examined by KLW in May 2006. It was determined earlier by Jean Raynal as A. borbonica but KLW re-determined it as A. nitens on the basis that it has the habit of A. nitens and the perianth bristles are slightly less than 3 mm long and longer than the nutlet, which is a feature of A. nitens rather than A. borbonica. There is a fragmentary possible isotype or isolectotype of R. boivinii in B: Insel Bourbon, Boivin 999 on sheet B 10 0000966. These fragments (glumes and one fruit with perianth bristles exceeding the fruit) in a paper packet were probably removed by Kükenthal from the collection in P for examination during his study of this group. The label on this B sheet is in Kükenthal’s script, ending with ‘6.II.1935 vid. G. Kükenthal’. There is another small packet on the sheet B 10 0000966 that contains glumes and nutlets of A. borbonica (with bristles shorter than the fruit). It has been annotated by Kükenthal as ‘leg. I.B. Balfour’. These fragments are probably from a Balfour specimen in K that was annotated by Kükenthal in October 1935, in which case the fragments were presumably removed by him then.
Of limited distribution but some populations are in national parks. We suggest a status of ‘Least Concern’ (IUCN Standards and Petitions Committee 2022).
Réunion: Pelouse altimontaine du Plateau Kerveguen, 16 Nov. 1973, F. Badré 964 (P 03471835, P 00592878); Bas de Bellecombe, 20 Dec. 1973, F. Badré 1116 (P 01948936); Bourbon [Réunion], [no date; received at Kew Oct. 1875], I.B. Balfour s.n. (K); Roches Plates path, Rivière des Remparts, 7000 ft [~2130 m], 16 Nov. 1967, C. Barclay 501 (K); Rivière des Remparts, Roches Plates path, 5100 ft [~1550 m], 20 Nov. 1968, C. Barclay 1251 (K); Piton Fougères east of Dos D’ane [sic], 4400 ft [~1340 m], 21 Nov. 1970, C. Barclay 1920 (K); Rampe de la Grande Montée, 4800 ft [~1360 m], 22 Nov. 1970, C. Barclay 1966 (K); Gite du vulcan, 2200–2300 m, June 1957, J. Bosser 11441 bis (P 03471834); SW of Piton de la Fournaise, 2255 m, 13 Jan. 2011, J.J. Bruhl 2949, J. Férard, L. Gigord, H. Hivert & K.L. Wilson (BOL, CANB, K, NE 101298, NSW, NU, P, PRE, REU); SW of Piton de la Fournaise, 2263 m, 13 Jan. 2011, J.J. Bruhl 2953, J. Férard, L. Gigord, H. Hivert & K.L. Wilson (BOL, BRI, CANB, CHR, EA, GENT, HO, K, M, MEL, MO, NE 101958, NSW, NY, P, PRE, REU, SI); SW rim of Piton de la Fournaise, 2265 m, 13 Jan. 2011, J.J. Bruhl 2955, J. Férard, L. Gigord, H. Hivert & K.L. Wilson (BOL, CANB, EA, K, NE 104037, NSW, P, PRE, REU); La Plaine des Cafres, Route du Volcan, 1904 m, 13 Jan. 2011, J.J. Bruhl 2963, J. Férard, L. Gigord, H. Hivert & K.L. Wilson (BOL, BRI, CANB,K, MEL, NE 104045, NSW, P, PRE, REU); La Plaine des Cafres, Route du Volcan, 1904 m, 13 Jan. 2011, J.J. Bruhl 2964, J. Férard, L. Gigord, H. Hivert & K.L. Wilson (BOL, BRI, CANB, K, MEL, NE 104046, NSW, P, PRE, REU); La Plaine des Cafres, Route du Volcan, 1904 m, 13 Jan. 2011, J.J. Bruhl 2966, J. Férard, L. Gigord, H. Hivert & K.L. Wilson (BOL, CANB, K, NE 104048, NSW, NY, P, PRE, REU); La Plaine des Cafres, Route du Volcan, 1904 m, 13 Jan. 2011, J.J. Bruhl 2967, J. Férard, L. Gigord, H. Hivert & K.L. Wilson (BOL, BRI, CANB, EA, K, NE 104049, NSW, NY, P, REU); La Plaine des Cafres, Route du Volcan, 1904 m, 13 Jan. 2011, J.J. Bruhl 2968, J. Férard, L. Gigord, H. Hivert & K.L. Wilson (K, NE 104050, NSW, P, REU); La Plaine des Cafres, Route du Volcan, 1904 m, 13 Jan. 2011, J.J. Bruhl 2972, J. Férard, L. Gigord, H. Hivert & K.L. Wilson (K, NE 104054, NSW, P, REU); La Plaine des Cafres, near Col Bellevue, 1627 m, 13 Jan. 2011, J.J. Bruhl 2976, J. Férard, L. Gigord, H. Hivert & K.L. Wilson (K, NE 104058, NSW, P, REU); above Piton de la Fournaise, Nez Coupé de Sainte Rose, 14 Jan. 2011, J.J. Bruhl 2985, J. Hivert & S. Pichillou (BOL, BRIP, CANB, EA, K, NE 104067, NSW, P, PRE, REU); above Piton de la Fournaise, Nez Coupé de Sainte Rose, 14 Jan. 2011, J.J. Bruhl 2986, J. Hivert & S. Pichillou (BOL, EA, K, NE 104068, NSW, P, PRE, REU); Plateau des Basaltes (sur la Plaine des Remparts), 2400 m, 31 May 1966, T. Cadet 452 (P 03471842, P 03471843); Plaine des Palmistes, bord du la Route N3, 1400 m, 16 Dec. 1968, T. Cadet 1787 (P 00592868); Plaines des Salazes, 2400 m, 14 Feb. 1970, T. Cadet 1955 (P 00592869); Cirque de Cilaos, au dessous couteau Kerveguen, 2000 m, 16 Nov. 1973, T. Cadet 4507 (P 00592870); Sentier de la Roche Ecrite, above Saint Denis, ~1200 m, 25 Nov. 1973, M.J.E. Coode 4186 & F. Badré (K, P 03471845); hauts de St Leu, 22 Oct. 1875, G. De L’Isle 619 (P 0059287); Rempart de Bellecombe, 3 Dec. 1970, F. Friedmann 870 (P 00592879); Piton de la Fournaise, Volcano, Plaine des Sables, ~2200 m, 9 Nov. 1966, H-J. Schlieben 10904 (PRE).
Réunion: above Piton de la Fournaise, Nez Coupé de Sainte Rose, 14 Jan. 2011, J.J. Bruhl 2979, J. Hivert & S. Pichillou (BOL, K, NE 104061, NSW, P, PRE, REU [arching, slender tussock perennial herb; perianth segments exceeding nutlet, typical of A. nitens.]; above Piton de la Fournaise, Nez Coupé de Sainte Rose, 14 Jan. 2011, J.J. Bruhl 2980, J. Hivert & S. Pichillou (BOL, BRI, CANB, CHR, EA, K, MEL, NE 104062, NSW, P, PRE, REU [much more slender than usual for A. nitens, but with nutlets more typical of A. nitens]; above Piton de la Fournaise, Nez Coupé de Sainte Rose, 14 Jan. 2011, J.J. Bruhl 2983, J. Hivert & S. Pichillou (BOL, CANB, K, MO, NE 104065, NSW, P, PRE, REU) [as slender as typical A. borbonica, but with nutlets more typical of A. nitens].
Asterochaete schlechteri (C.B.Clarke) X.F.Zhang, comb. nov.
Carpha schlechteri C.B.Clarke in E.A.J. de Wildeman, Pl. Nov. Hort. Thenensis 1: 37, t. 9 (1904). Type citation: ‘Afrique australe. – Région austro-occidentale: Koude Bokkeveld, Gydow à 4500 pieds (R. Schlechter, 17 janvier 1897).’ Type: South Africa: Western Cape Province: Koude Bokkeveld, Gydow, 4500 ft [~1370 m], 17 Jan. 1897, R. Schlechter 10010 (lecto, here designated: BR 8644664*; isolecto: B 10 000976 (fragments of inflorescence), BOL 63205, E 200182*, GENT, K, L 0819901*, NSW 181489, P 00199388, P 00461848, P 00461848, PRE (two sheets), S-G-6592*, WAG 0000846*, Z 000052364*).
Erect perennial, 80–130 cm high; rhizome long. Culms triquetrous to trigonous in cross-section, with 1 or 2 nodes, each node bearing a single leaf. Leaves crowded at the base of the culms; blades straight, 40–60 cm long, 4.5–6(–8) mm wide, V-shaped in cross-section, keeled abaxially, scabrous above, smooth below; sheaths pale to dark yellow-brown. Inflorescence 35–60 cm long, with 4–7 internodes up to 20 cm long; spikelets 120–500 in 6–20 dense ovoid clusters. Involucral bracts much shorter than inflorescence, up to 33 cm long, up to 6 mm wide. Spikelets 4–5 mm long, with 2 flowers, lower one male, upper one bisexual; glumes 5–7, pale yellow-brown to reddish brown; lowest 3 or 4 glumes empty, persistent; the adjacent two above fertile, sometimes an empty uppermost glume present, deciduous; lowest 2 or 3 glumes shorter than the rest; the proximal fertile glume 3.5–4.2 mm long, 1.2–1.8 mm wide, the second fertile glume 3.3–4.3 mm long, 0.5–1.5 mm wide; the axis not elongated above the proximal fertile glume. Bristles: upper part (3/5‒2/3) of each bristle terete, breaking off readily, antrorsely scabrous, lowest 1/3–2/5 much more slender, smooth; outer 3 bristles shorter than to equalling the body of the nutlet, shorter than inner 3; the inner 3 2.3–3.4 mm long, slightly shorter than to slightly exceeding the nutlet body (length often variable within a whorl). Anthers pale yellow, 1.6–2.5 mm long (excluding appendage), with apical appendage 0.3–0.5 mm long, ~0.1 mm wide. Stylebase 0.2–0.5 mm long, 0.1–0.2 mm in diameter, glabrous or minutely hispidulous or verruculose. Nutlet 1 per spikelet, narrow-ellipsoid to ellipsoid, 2.2–2.5 mm long, 0.7–1.0 mm in diameter, dark brown at maturity, surface reticulate. (Fig. 1k; see Zhang et al. 2004b, fig. 7h, o, q for spikelet structure; Reid and Arnold 1984, fig. 2.2e, 4b).
Endemic to the Koue Bokkeveld mountains of the Ceres District of the Western Cape Province, South Africa (Fig. 4d; Reid and Arnold 1984; Archer and Muasya 2012). Only known from a few collections, so the distribution may be broader than currently known. In marshes and on sandy flats near streams, at 900–1370 m in altitude.
Carpha schlechteri was first described by Clarke (1904, p. 37) as ‘C. glomerata Nees affinis; differt: foliis angustioribus (vix 5 mm. latis); inflorescentia graciliore, spiculis minoribus; nuce grosse conspicue hexagono-reticulata levi (neque, ut in C. glomerata, minute obscure reticulata)’. That is, he considered it to differ from A. dactyloides in its narrower leaves, its fewer clusters of spikelets in the inflorescence and larger reticulations on the nutlet surface. We found that the reticulations on the nutlet surface of C. schlechteri are not obviously larger than those of A. dactyloides, and that spikelet cluster number in the two species overlaps slightly. The nutlet surface in A. schlechteri is unique in the genus in having the outer epidermal cell walls reticulate and colliculate, forming a layer that sloughs off readily as a single sheet to expose the inner walls of the epidermal cells as a reticulate pattern as seen in the other species in this genus. The epidermal layer is similar in A. dactyloides but not as obvious nor as readily sloughed off as in A. schlechteri.
As mentioned above under Asterochaete dactyloides, that species and A. schlechteri are similar morphologically, including having the same unusual form of perianth bristles, but A. schlechteri has the proximal flower male and produces only one nutlet per spikelet; the axis is not elongated above the proximal fertile node; spikelets are 4–5 mm long; the proximal fertile glume is 3.5–4.2 mm long and 1.2–1.8 mm wide, whereas the second fertile glume is 3.3–4.3 mm long and 0.5–1.5 mm wide; the nutlets are 2.1–2.4 mm long; and the inner-whorl bristles are 2.3–3.4 mm long. By contrast, A. dactyloides has two bisexual flowers in each spikelet, both developing nutlets; the axis is elongated above the proximal fertile node; the spikelets are 6.2–9 mm long; the proximal fertile glume is 5.8–7.5 mm long and 1.8–2.8 mm wide and the second fertile glume is 5.5–7 mm long and 1.5–2 mm wide; the nutlets are 2.5–3.4 mm long; and inner-whorl bristles 3.1–3.8 mm long.
Asterochaete schlechteri is found only at high altitude in the Koue Bokkeveld Mountains of the Western Cape Province, whereas A. dactyloides has a much wider distribution, often at a lower altitude, across southern South Africa.
The sheet in BR, annoted by Clarke with a draft description, is selected as lectotype. This specimen is labelled as being originally in the herbarium of the Hortus Thenensis (‘Horti Thenensis Herbarium’) in the town of Tienen (Tirlemont in French and latinised as Thenen), which was owned by Léon van den Bossche. As van den Bossche wrote in the preface to part 1 (de Wildeman 1904), he commissioned Emile de Wildeman (director of the Jardin Botanique de l’Etat de Bruxelles) to write the Plantae novae vel minus cognitae ex Herbario Horti Thenensis, which in practice meant that de Wildeman also edited contributions by specialists in particular groups, including Clarke (1904). In the preface, van den Bossche stated that the Schlechter specimens being used to describe the new species were in his personal herbarium. This means that, in the case of Carpha schlechteri, it could be argued that the BR specimen is the holotype of the name. However, if one takes the publication literally, Clarke’s protologue text does not state explicitly which herbarium the type collection is in, so Article 9.1 (Turland et al. 2018) does not apply. As well as the sheet in BR, Clarke also annotated the sheets BOL 63205 and the K sheet, indicating that he had seen more than one of the sheets of Schlechter 10010, which implies that these are syntypes (Art. 9.6) and that it is therefore appropriate to select a lectotype.
Collected only twice in the past 80 years, most recently in 1996; it is regarded as ‘Data Deficient’ (Raimondo et al. 2009, as Carpha schlechteri), but it is probably ‘Endangered’ (IUCN Standards and Petitions Committee 2022).
Asterochaete ulugurensis Nelmes ex X.F.Zhang, sp. nov.
Planta Asterochaete nitenti Kunth simulans sed in characteribus inflorescentiarum differt. Spiculae in capitula oblonga dense congestae, non laxe dispositae. Asterochaete borbonica (Steud.) X.F.Zhang simulans, sed in characteribus foliorum differt: folia plana carinata 1.7–5.5 mm lata, non perangusta canaliculata ecarinata.
Type: Tanzania: Morogoro District: Lukwangule, Uluguru Mountains, 8 Dec. 1969, T. Pócs 3766 (holo: K; iso: EA).
[Carpha glomerata auct. non (Nees) Nees: R.W. Haines & K.A. Lye, Sedges & Rushes East Africa 322 (1983) p.p.; K. Hoenselaar et al., Fl. Trop. East Africa, Cyperaceae 366 (2010); J.-P. Lebrun & A.L. Stork, Trop. African Fl. Pl. 11: 81 (2020)]
Tufted perennial, 70–80(–90) cm high; rhizome very short. Culms unnoded or 1-noded and bearing a single leaf. Leaves crowded at the base of the culms; blades straight, 40–65 cm long, 2.2–5.5 mm wide, narrowly to broadly V-shaped, to V-shaped near apex in cross-section, keeled abaxially (more strongly near apex), scabrous near apex only, smooth below; sheaths pale yellow-brown. Inflorescence 9–30 cm long, with 4–6 short internodes ~2–7 cm long; spikelets ~100–200, in 3–6 dense oblong clusters. Involucral bracts 10–27 cm long, up to 4.5 mm wide. Spikelets 5–7 mm long, with 1 or 2 bisexual flowers; glumes 5, dark yellow-brown to red-brown; lowest 3 glumes empty, persistent, either both the upper 2 fertile or the uppermost one sterile, both glumes deciduous; lowest 2 glumes shorter than the rest; the proximal fertile glume 4.3–5.2 mm long, 1–1.8 mm wide, the second fertile glume 4.1–4.7 mm long, 1–1.3 mm wide; the axis elongated above proximal fertile glume and adnate to the base of that glume. Bristles terete to trigonous above, antrorsely scabrous (sparsely so towards base), with the often slightly more slender, glabrous, basal part (~1/5–1/4 of length), somewhat flattened and flared at base; outer 3 bristles shorter than the nutlet body (1/2‒3/4 the length), shorter than inner 3 (shortest often ~1 mm long); the inner 3 from 2/3 as long as to slightly exceeding the nutlet body (up to 1.3 times as long), 2.0–3.0 mm long. Anthers greyish to blackish, 1.4–2.2 mm long (excluding appendage), with apical appendage 0.1–0.2 mm long, ~0.1 mm wide. Stylebase ~0.2 mm long, 0.1–0.3 mm in diameter, glabrous or occasionally minutely verruculose or hispidulous. Nutlets 1 or 2 per spikelet, narrow-ellipsoid to narrow-obovoid, 2.0–2.6 mm long, 0.5–0.7 mm in diameter, dark red-brown at maturity, surface reticulate. (Fig. 1l; see Zhang et al. 2004b, fig. 7e, k, for spikelet structure). (Fig. 7.)
Specimen of Asterochaete ulugurensis (E.M. Bruce 742, K). Reproduced with permission of The Board of Trustees of the Royal Botanic Gardens, Kew.

Endemic to the Lukwangule Plateau in the Uluguru Mountains, Morogoro District of Tanzania (Fig. 4b). In alpine peat bogs or marshes (at 2300–2500 m in altitude) in grassland.
The specific epithet refers to the locality of this species, and is the manuscript name written on a specimen in K (E.M. Bruce 742; see Fig. 7) by the late Ernest Nelmes, a cyperologist at the Royal Botanic Gardens Kew. The type was labelled as ‘Carpha ulugurensis Nelmes’ by the late Sheila Hooper, and another specimen in K (Bidgood et al. 232) was also so labelled by David Simpson in 1987.
The specimens of this species were provisionally named as Carpha ulugurensis and C. cf. nitens in our previous analyses (Zhang 2002; Zhang et al. 2006). This species resembles A. nitens (which is endemic on Réunion) but differs from A. nitens in inflorescence features, number of culm nodes, and nutlet length. Its culms are one-noded or unnoded, whereas A. nitens has (1–)2–3-noded culms. Its nutlets are longer than those of A. nitens (2.0–2.6 v. 1.5–1.8 mm), with a very short persistent stylebase. The epidermal cells of the fruits of A. ulugurensis are mostly slightly longitudinally elongated, whereas those of A. nitens are isodiametric or slightly horizontally elongated. The stylebase is very short and glabrous, whereas it is better developed in A. borbonica and A. nitens, with a hispidulous surface. It was illustrated as Carpha glomerata in Haines and Lye (1983, fig. 664, 665). These authors stated that the illustrations were based on specimen E.M. Bruce 742 from Mount Uluguru.
This is one of three tropicalpine species in eastern Africa, each endemic to high peaks in a different mountain range. It differs from A. eminii in having more numerous spikelets, with one or two flowers per spikelet, shorter glumes (4.1–5.2 v. 5.5–7.5 mm in A. eminii) and nutlets (2.0–2.6 mm long v. 2.8–3 mm in A. eminii), inner bristles shorter than to slightly exceeding the nutlet body (much longer than the body in A. eminii), and anthers 1.4–2.2 mm long (2.8–4.0 mm in A. eminii).
The isotype bears more complete collecting information than does the holotype. The collectors and number are given on the isotype as ‘T. Pócs, P. & K. Csontos, BJH [B.J. Harris] 3766’, and the habitat is given as ‘peat bogs with Sphagnum’ at an altitude of 2385 m.
Of limited distribution but some populations are in national parks; we suggest a status of ‘Vulnerable’ (IUCN Standards and Petitions Committee 2022), but ‘Endangered’ may be more appropriate given that the high-altitude habitat is limited and predicted to become even more so with changing climate and increasing human impact.
Tanzania: South Ulugurus, Lukwangule Plateau, 2400 m, 14 Mar. 1986, S. Bidgood, J. Lovett, J.L. Paul & T. Poćs 232 (K); Ulugurus, Lukwangule Plateau, 8200 ft [~2500 m], 30 Jan. 1935, E.M. Bruce 742 (BM, BR 24800907*, K, NU (fragment with photocopy of K sheet and notes by J. Browning), P).
Carpha Banks & Sol. ex R.Br., Prodr. 230 (1810)
Type: Carpha alpina R.Br. (designated by T.Koyama, J. Fac. Sci. Uni. Tokyo, Sect. 3, Bot. 8, 77 (1961)).
Perennial herbs with short rhizome. Culms erect, trigonous to more or less terete in cross-section, unnoded or 1-noded. Leaves basal and cauline, eligulate, pseudopetiolate, not always conspicuously so. Inflorescence condensed panicle-like with short slender branches, spikelets forming 1–3 loosely obovoid or obconical to oblong clusters, or sometimes not forming obvious clusters. Involucral bracts leaf-like. Spikelets compressed, with 1 or 2 bisexual flowers; glumes (floral bracts) 5–6(–7), distichously arranged, persistent, the lower ones empty, approximately half the size of the upper glumes, the uppermost glume usually linear, empty. Perianth consisting of 6 bristles in 2 whorls, falling with the nutlet, pale to dark golden brown, flattened and plumose along the margins for most of their length, terete to filiform and antrorsely scabrous near the apex, more or less equal in length. Stamens 3, rarely 2; anthers linear, pale yellow to grey, with a short pyramidal to conical apical appendage. Style slender, continuous with the ovary, lower part enlarged and persistent on the nutlet; stigmas 3. Nutlet trigonous in cross-section, on an obvious gynophore, with surface minutely reticulate (only clearly visible with magnification), occasionally minutely puncticulate.
A genus of five species, found at high altitude in south-eastern Australia (at lower altitude in Tasmania), and at high altitude in New Guinea, in New Zealand and temperate South America (Fig. 3, 8; Wilson 1986, fig. 3).
Distribution of Carpha spp.: (a) C. alpina; (b) C. curvata; (c) C. nivicola; (d) C. rodwayi; (e) C. schoenoides.
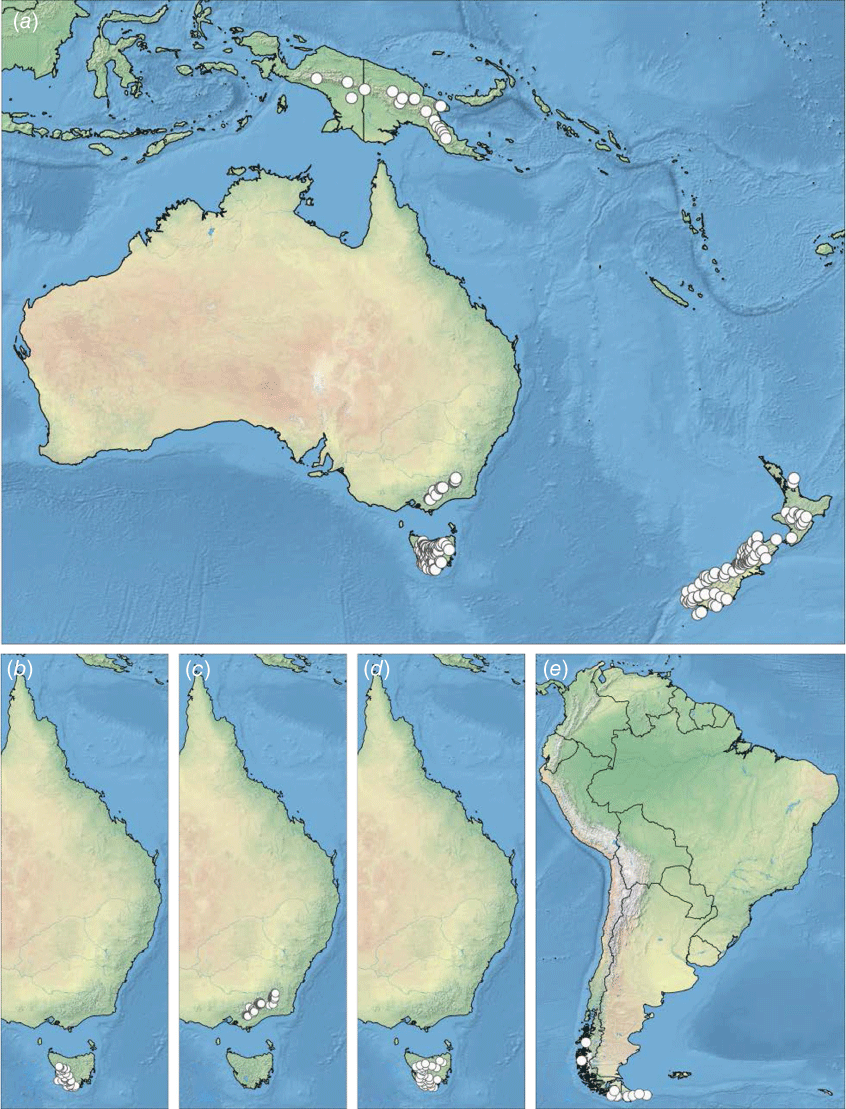
The phylogeny of Carpha is not clear and further molecular studies are needed. Biogeographically, the distribution of its species suggests long-distance dispersal (Wilson 1986), particularly the presence of C. alpina in tropicalpine habitats at a high altitude in New Guinea, alpine areas of south-eastern mainland Australia, and at more varied altitudes in New Zealand and Tasmania. Dispersal between Australia and New Zealand could have occurred in either direction (Winkworth et al. 2002). The presence of C. schoenoides in temperate South America also suggests long-distance dispersal, given that the two continents have been isolated for so long (Wardle et al. 2001).
| 1 | Spikelets 2-flowered; nutlet epidermis reticulate, not puncticulate...C. schoenoides Spikelets 1-flowered; nutlet epidermis reticulate or not reticulate, puncticulate...2 |
| 2 | Upper third of leaves conspicuously curled on drying; terminal flowering cluster loosely oblong to obconical; nutlet surface reticulate and puncticulate...C. curvata Leaves straight throughout their length (tips may curl sometimes on drying in C. alpina); terminal flowering cluster loosely obconical; nutlet surface puncticulate, not reticulate...3 |
| 3 | Plants forming extensive mats; leaf blades rigidly erect, apex ±pungent ...C. rodwayi Plants tufted; leaf blades not rigidly erect, not pungent...4 |
| 4 | Spikelets 8–12.5 mm long; nutlet 2.2–3.5 mm long, 0.5–1.2 mm in diameter; perianth bristles 7.5–12 mm long; leaf blades 0.5–2.0 mm wide...C. alpina Spikelets 13.5–19 mm long; nutlet 3.5–4.5 mm long, 0.9–1.4 mm in diameter; perianth bristles 13–19 mm long; leaf blades 1.5–3 mm wide...C. nivicola |
Carpha alpina R.Br., Prodr. Fl. Nov. Holl. 230 (1810)
Rhynchospora alpina (R.Br.) Spreng., Systema Vegetabilium 1: 195 (1824); Chaetospora alpina (R.Br.) F.Muell., Fragm. 9: 39 (1875), p.p. Type citation: ‘D’ [= Tasmanian regions visited by Robert Brown]. Type: Australia: Tasmania: on the summit of Table Mountain [now called Mt Wellington], near the Derwent River, Mar.–Apr. 1804, R. Brown (Bennett 6020) (lecto, designated by K. L. Wilson & J. J. Bruhl in D. J. Mabberley & D. T. Moore, Robert Brown Handb. 115 (2022): BM 000990989; probable isolecto: BM 00092170, BM 000990990, BRI 015597, BRI AQ342063, E 00686463, E 00686464*, F 0093183, FI-W 012213, G 00191591, G-G-167760-1, K, MEL 49294, NSW 687664, NY 0021813, P 00603248, P 00603249, P 00603251, P 00603252, P 00603253).
Carpha tasmanica Gand., Bull. Soc. Bot. France 66: 297 (1920). Type citation: ‘Tasmania, in m. Wellington (Lucas!).’ Type: Australia: Tasmania: Mt Wellington, Jan. 1901, A.H.S. Lucas s.n. (syn: BRI-AQ340667; LY, n.v. (photo in NSW; McGillivray 1973); NSW 120927).
Carpha novae-zelandiae Gand., Bull. Soc. Bot. France 66: 297 (1919). Type citation: ‘Nova Zelandia, in Maungatua (Thomson!)’. Type: New Zealand: Maungatua, [no date], Thomson s.n. (holo: LY?, n.v.).
Tufted perennial, 6–35 cm high. Culms 1-noded, with a single cauline leaf, or unnoded. Leaves crowded at the base of the culms; blades more or less straight, 3–20 cm long, 0.5–2 mm wide, sometimes tips tending to curl on drying, thinly but stiffly crescentiform in cross-section; sheaths pale to medium yellow-brown in lower half. Inflorescence with 1–4 short internodes, 2–16 cm long; 3–26 spikelets in a loose to more or less dense obconical cluster, rarely not forming an obvious cluster. Involucral bracts several, (1.5–)3–15 cm long, up to 1.9 mm wide. Spikelets 8–12.5 mm long, with 1 (very rarely 2) bisexual flowers; glumes 5–6(–7), pale yellow-brown; lowest 3–4(–5) glumes empty; lowest 2–3(–4) glumes shorter than the rest; the fertile one 7–10.5 mm long, 1–2.1 mm wide; the uppermost one linear, sterile or very rarely fertile; axis not elongated above proximal fertile glume. Bristles 7.5–12 mm long, plumose with hairs up to 2.5 mm long; distal scabrous filiform zone 0.5–2.7 mm long. Anthers pale yellow-grey, 1.1–2.9 mm long (excluding appendage), with an apical appendage 0.1–0.3 mm long, ~0.1 mm wide. Stylebase 2.8–5.0 mm long, 0.15–0.4 mm in diameter, sparsely antrorsely hispidulous for its whole length. Nutlet 1 per spikelet, with a gynophore 0.3–0.6 mm long, the body narrow-ellipsoid, 2.2–3.5 mm long, 0.5–1.1(–1.2) mm in diameter, mid- to dark yellow-brown at maturity with 3 longitudinal ribs slightly paler or not paler, surface minutely reticulate and puncticulate, epidermal cell outlines more or less isodiametric, apparently not in longitudinal columns (not easy to see without high magnification). (Fig. 2a, f; see Zhang et al. 2004b, fig. 7b (rarely 7f) for spikelet structure; Zhang et al. 2006, fig. 3d; Hooker 1877, pl. 1216; Clarke 1909, t. LXXVI, fig. 1–4; Kern 1974, fig. 74; Costin et al. 1979, pl. 103; Wilson 1994a, fig. 55f.)
At high altitude in south-eastern Australia (New South Wales, Victoria and Tasmania), New Zealand (North Island, South Island, Stewart Island (and the Auckland Islands: Johnson and Campbell 1975), and New Guinea; occasionally found near sea level in southern Tasmania, New Zealand and the Auckland Islands (Fig. 8a). In bogs, fens and other wet alpine and subalpine habitats (Fig. 8).
There is variation among the specimens under this name that should be further investigated. For example, a specimen from Tasmania (Mount Sprent, J. Kirkpatrick s.n., Dec 1981, HO 91834) with short relatively dark yellow-brown glumes may represent a new taxon or possibly a hybrid with C. curvata or C. rodwayi (q.v.). Variation in New Zealand has not been investigated.
The species has a very broad distribution, including in national parks, so is not considered to be at imminent risk. However, the alpine habitat is limited and predicted to become even more so with changing climate, so the status is likely to change in the next few decades. The species is already listed as ‘Endangered’ in Victoria (Department of Environment, Land, Water and Planning 2021).
Indonesia: West Irian: Northern slopes of Mount Wilhelmina [now Mount Trikora], 4160 m, Sep. 1938, L.J. Brass 10078 & E. Meyer-Drees (BM 92168); Dugundugu Plateau, 0.5 km N of the Noorolwand, Carstenz Mountains, 4080 m, 26 Jan. 1972, G. Hope ANU 16047 (CANB 229944); terraces after passing Mount Trikora Pass to the west, alt. 3790 m, 9 Aug. 1984, J-M. Mangen 1160 (L 1367808); Carstenz Meadow, extrémité S, alt. 3400 m, 29 Apr. 1973, J. Raynal 17381 (BR 35477143*, P 01926637).
Papua-New Guinea: Central District: Mount Scratchley, Tapini Subdistrict, ~11,000 ft [~3350 m], 7 May 1971, M. Coode & P. Stevens NGF 46317 (NSW1002190; A, BISH, BOGOR, BRI, CANB, K, L, LAE, PHN, SING, US, all n.v.); Mount Albert Edward, between E and W domes, 3600 m, Goilalal Subdistrict, 20 June 1974, J. Croft et al. [names not specified on label] LAE 61353 (NSW 462097; A, BISH, BM, BOGOR, BRI, CANB, E, K, L, LAE, M, PNH, US, all n.v.); Mount Dickson, Goilala Subdistrict, alt. ~11,500 ft [~3500 m], 11 Feb. 1964, T.G. Hartley 12996 (CANB, K). Eastern Highlands District: Mount Wilhelm, SW of Lake Aunde, 3978 m, 25 Jan. 2015, J.J. Bruhl 3303, P. Dua, P.P. Homot & M. Lovave (CANB, CHR, K, LAE, NE 101699, NSW); Mount Wilhelm, ~3550 m, Chimbu Province, 5 July 1980, P. Goetghebeur 3544 & E. Coppejans (GENT n.v., NSW 462100); Mount Knutsford, 1889, W. MacGregor (MEL 2066102). East Sepik District: Burgess Mountain, 3675 m, 24 Mar. 1977, J.F. Veldkamp & A. Vines 7441 (HBG, L n.v.). Morobe District: Mount Sarawaket, 11–12,000 ft [~3350–3650 m], 6 Apr. 1937, M.S. Clemens 6051A (HBG); summit region Mount Sarawaket, 3800 m, 15 Dec. 1975, J.R. Croft & Y. Lelean LAE 68440 (K, NSW 1002188; A, BISH, BRI, CANB, E, L, LAE, M, all n.v.). Southern Highlands District: Mount Giluwe, ~12,300 ft [~3750 m], 14 Aug. 1961, R. Schodde 1827 (CANB 107745, L 1367821#, WAG 17771449#). West Sepik District: Telefomin Subdistrict, S slope of Mount Scorpion, Star Mountains, alt. 3400 m, 25 May 1975, J.R. Croft & G.S. Hope LAE 68020 (K, NSW 1002187, US#; A, BO, BRI, CANB, L, LAE, SING, all n.v.).
Australia: New South Wales: Southern Tablelands: Tate East Ridge (near Gills Knob), Kosciuszko National Park, 10 Feb. 1978, J. Thompson 2947 (CANB, NSW 462091); 1 km from Snowy River bridge, 10 m from river, Kosciuszko National Park, 24 Jan. 2000, X. Zhang 13 & K.L. Wilson (K, NE 72986, NSW 593159). Victoria: Snowfields: ~500 m N of Mount Jim, alt. 1780 m, Bogong High Plains, 26 Feb. 1982, R.J. Adair 1620 (MEL 693734); Bogong National Park, 1.5 km beyond turnoff to Wallace’s Hut towards Omeo, 7 Feb. 1985, E.H. Norris 356 (NSW 462089); Bogong High Plains, Mount Nelse snowpatch, 1880 m, 14 Feb. 1981, H. van Rees 281 (MEL). Tasmania: Ben Lomond region: Hamilton Crags, Ben Lomond National Park, 1400 m, 28 Mar. 1986, A.M. Gray 676 (HO 328073). Central Highlands: Mount Inglis, 1200 m, 24 Feb. 1983, A. Moscal 1955 (HO 100424); King William Saddle, 830 m, 4 Feb. 1984, A. Moscal 6135 (HO 400516); Forth River track, 25 Mar. 1992, K.L. Wilson 8359 (NSW 254555). South West: Lake Dobson, 50 m NE of SE corner, 1027 m, 15 Feb. 2000, J.J. Bruhl 1886 & F.C. Quinn (G, HO 533181, L 3777949, NE 71826, NSW 658680, UPS); West Coast Region, 3 Feb. 1985, S.J. Jarman 244 (HO 326461); Mount Wellington, Jan 1901, A.H.S. Lucas s.n. (NSW 120927); start of Lake Osborne track, Hartz Mountains National Park, 26 Feb. 1986, K.L. Wilson 6643 (HO 506627, MO, NE 70799, NSW 432013, RSA).
New Zealand: North Island: National Park Railway Station, Volcanic Plateau, 790 m, 16 Mar. 1978, R.O. Gardner 1866 (AK n.v., NSW 817178, MO n.v.); Taurewa, Central Plateau, 800 m in altitude, 18 Jan. 1973, E.M. Miller s.n. (AKU n.v., AAU); Tongariro National Park, 26 Jan. 1950, C.B. Trevarthen s.n. (NSW 462094). South Island: Wairau Mountains, Nelson, 4000 ft [~1200 m], Jan. 1878, T.F. Cheeseman (NY); Westport, ridge above German Terraces, 19 Mar. 2001, D.S. Glenny 8413 (CHR, NSW 817181); Mount Arthur, 23 Mar. 1987, P.D. Hind 4814 & C. Sproule (NSW 219224); Lake Misery, [no date], T. Kirk 456 (BM 92171); Arthurs Pass, 10 Mar. 1956, J.D. Lovis 1351 (BM 92172, BR 9507159*); below Sealy Lake, Mount Cook District, 12 Jan. 1956, J.D. Lovis 1077 (BM 000092173); near Lake Sylvester, 12 Jan. 1962, R. Melville 5904, E. Melville & H. Talbot (AK 155582, K, NSW 817179). Stewart Island: Mount Anglem, 17 Feb. 1962, R. Melville 6372 (AK 155584, K n.v., NSW 462095).
Carpha curvata W.M.Curtis, Brunonia 7(2): 303 (1984)
Type citation: ‘Holotypus: J. & M. Davies s.n., Eliza plateau [sic], South-west National Park, east of Lake Pedder, 25. i. 1982, HO 49444: isotype: NSW.’ Type: Australia: Tasmania: Eliza Plateau, South-West National Park, east of Lake Pedder, 25 Jan. 1982, J. & M. Davies s.n. (holo: HO 49444; iso: NSW 687892).
Tufted perennial, 18–40 cm high. Culms 1-noded with a single cauline leaf, or unnoded. Leaves crowded at the base of the culms; blades more or less straight, 15–55 cm long, 1.2–2.2 mm wide, thinly but stiffly crescentiform in cross-section, upper third of leaf blade conspicuously curled with age and when dried; sheaths rich yellow-brown in lower half. Inflorescence with 2–4 short internodes, 3.5–20 cm long; 15–35 spikelets in 1 or 2 loosely oblong to obconical clusters. Involucral bracts several, up to 17 cm long, up to 2.5 mm wide. Spikelets 8.5–11 mm long, with 1 bisexual flower; glumes 5 or 6, pale to mid yellow-brown; lowest 3 or 4 glumes empty, lowest 2 or 3 glumes shorter than the rest; the fertile one 7.9–10.1 mm long, 1.3–2 mm wide; the uppermost one linear, sterile; axis not elongated above proximal fertile glume. Bristles 7.8–10.5 mm long, plumose with hairs up to 1.3 mm long; distal scabrous zone 1.6–2.6 mm long. Anthers pale yellow, 2.3–4.5 mm long (excluding appendage), with apical appendage 0.1–0.2 mm long, 0.1–0.15 mm wide. Stylebase 2.0–4.2 mm long, 0.2–0.4 mm in diameter, minutely antrorsely hispidulous towards apex, glabrous towards base, or sparsely hispidulous throughout. Nutlet 1 per spikelet, with a gynophore 0.3–0.5 mm long, the body narrow-ellipsoid, 2.8–3.5 mm long, 0.8–1.2 mm in diameter, pale orange-brown to dark red-brown at maturity with 3 slightly paler longitudinal ribs, surface very minutely reticulate and slightly puncticulate, cells more or less isodiametric, more or less in longitudinal columns (Fig. 2b, f; see Zhang et al. 2004b, fig. 2a, 7b for spikelet structure).
Endemic to the Central Highlands and South West regions of Tasmania, Australia (Fig. 8b). On mountain slopes, and streamsides in shrubby heathland, to sea level around Bathurst Harbour.
Widespread, including in national parks, so not considered to be at imminent risk; we suggest a status of ‘Least Concern’ (IUCN Standards and Petitions Committee 2022). However, the alpine habitat is limited and predicted to become even more so with changing climate, so the status is likely to change in the next few decades.
Australia: Tasmania: Central Highlands: above Lake Ewart, 950 m, 5 Feb. 1987, A.M. Buchanan 9948 (HO 122194). South West: Mount Anne National Park, 75 m NE of Mount Eliza, 1274 m, 16 Feb. 2000, J.J. Bruhl 1894 & A.C. Rozefelds (HO, NE 71843, NSW 599815); Mount Anne National Park, track below Mount Eliza and hut, 1173 m, 16 Feb. 2000, J.J. Bruhl 1895 & A.C. Rozefelds (CANB, HO, K, MO, NE 71844, NSW 658774); Mount Anne National Park, Mount Anne track below Mount Eliza hut, 768 m, 16 Feb. 2000, J.J. Bruhl 1896A & A.C. Rozefelds (HO, K, L, MO, NE 71845, PRE); Schnells Ridge, 27 Jan. 1998, A.M. Buchanan 15043 (HO, NSW 470935); Hamilton Range, ~880 m, 26 Feb. 1986, S.J. Jarman s.n. (HO 412117); summit of Elliot Range, 15 Jan. 1985, S.J. Jarman & G. Kantvilas s.n. (AD, CANB, HO 443230, MEL, NSW 487467); Mt Sprent, Dec. 1981, J. Kirkpatrick s.n. (HO 91835); Mount Hesperus, 6 Feb. 1982, A.V. Ratkowsky s.n. (HO 53801).
Carpha nivicola F.Muell., Defin. Austral. Pl. 34 (June‒July 1855)
Type citation: ‘On the highest summits of the Australian Alps, near swamps.’ Type: Australia: New South Wales: Southern Tablelands: Australian Alps, 1853, F. Mueller (lecto, here designated: MEL 2295059; isolecto: MEL 2295060).
Carpha alpina var. subacaulis Kük., Feddes Repert. 47: 113 (1939). Type citation: ‘Neu-Süd-Wales: Mt Kosciusko, Baumgrenze bis 7000’, I. 1899 (J. H. Maiden!).’ Type: Australia: New South Wales: Southern Tablelands: Mt Kosciuszko, Baumgrenze bis [tree-line to] 7000 ft [~2130 m], Jan. 1899, J.H. Maiden 4 (syn: B, NSW 636993).
[Chaetospora alpina auct. non (R.Br.) F.Muell.: F.Mueller, Fragm. 9: 39 (1875), p.p.]
[Carpha alpina auct. non R.Br.: G.Bentham, Fl. Austral. 7: 381 (1878), p.p.; H.H.Pfeiffer, Feddes Repert. 29: 178 (1931), p.p.; G.Kükenthal, Feddes Repert. 47: 112 (1939), p.p.]
Tufted perennial, 8–40 cm high. Culms unnoded, or rarely 1-noded with a single cauline leaf. Leaves crowded at the base of the culms; blades more or less straight, 5–23 cm long, 1.5–2.8 mm wide, thinly but stiffly crescentiform in cross-section; sheaths mid- to dark yellow-brown towards the base, occasionally with darker patches. Inflorescence with 1–3 short internodes, 2–11 cm long; 9–25 spikelets in a single obconical cluster or head. Involucral bracts several, up to 9.5 cm long, up to 2 mm wide. Spikelets 13.5–19 mm long, with 1 bisexual flower; glumes 5 or 6, pale yellow-brown; lowest 3 or 4 glumes empty, lowest 2 or 3 glumes shorter than the rest; the fertile one 12.5–17 mm long, 2–3.1 mm wide; the uppermost one linear, sterile; axis not elongated above proximal fertile glume. Bristles 13–19 mm long, plumose with hairs to 2.1 mm long; distal scabrous zone 0.8–2.5 mm long. Anthers pale yellow to pale grey, 2.6–4 mm long (excluding appendage), with apical appendage 0.05–0.15 mm long, 0.05–0.1 mm wide. Stylebase 6–9.8 mm long, 0.2–2 mm in diameter, glabrous, or occasionally glabrous towards base and minutely antrorsely hispidulous towards apex. Nutlet 1 per spikelet, with a gynophore 0.5–0.7(–1.0) mm long, the body narrow-ellipsoid to ellipsoid, 3.5–4.5 mm long, 0.9–1.4 mm in diameter, pale yellow-brown at maturity with 3 longitudinal ribs slightly paler, surface minutely reticulate, slightly puncticulate, cells isodiametric or slightly elongated longitudinally (Fig. 2c, h; see Zhang et al. 2004b, fig. 7b for spikelet structure; Costin et al. 1979, pl. 102; Wilson 1994a, fig. 55d, e).
Only in high-altitude areas of south-eastern Australia: New South Wales and Victoria (Fig. 8c). In bogs, fens and other wet alpine and subalpine habitats.
Mueller (1875) changed his mind and included his species Carpha nivicola in C. alpina 20 years after he first published C. nivicola. This was accepted by some subsequent authors (Bentham 1878; Pfeiffer 1931; Kükenthal 1939a). However, other authors (Blake 1940; Costin et al. 1979; Thompson 1981; Thompson and Gray 1981; Wilson 1993, 1994a, 1994b) have regarded Carpha nivicola as being separate from C. alpina. Our studies (Zhang 2002; Zhang et al. 2006) have confirmed that Carpha nivicola should be treated as a species. Carpha nivicola differs from C. alpina in having larger spikelets, flowers and nuts, and wider leaves.
There are two sheets in MEL that could be considered as lectotype. Both are labelled as coming from the ‘Australian Alps’ and both probably come from the same trip to the Mount Kosciuszko region by F. Mueller in 1853; indeed, both are morphologically similar and could come from the same collection. However, they differ in their labels and neither can satisfactorily be recognised as a holotype, despite the earlier labels to that effect by S. T. Blake and K. L. Wilson. MEL 2295059 bears two fertile tufts and a sterile tuft of this taxon. The handwritten label (‘Carpha nivicola/ferd Mueller, Australian Alps’) is not in Mueller’s script; it is in the same elegant, large script seen on other 19th century specimens in MEL and appears to have been written for display purposes. J. H. Willis has annotated the label with ‘1853’. The other sheet, MEL 2295060, bears one fertile tuft and one sterile tuft. Its label is in Mueller’s script: ‘Carpha alpina/Australian Alps’; J. H. Willis has also annotated this with ‘1853’. The first sheet, MEL 2295059, is here selected as lectotype because its label has the name of the new taxon and the collector’s name, as well as there being more material on the sheet.
The species has a reasonably broad distribution, including in national parks, so it not considered to be at imminent risk. However, the alpine habitat is limited and predicted to become even more so with changing climate, so the status is likely to change in the next few decades. The species is already listed as Endangered in Victoria (Department of Environment, Land, Water and Planning 2021).
Australia: New South Wales: Southern Tablelands: 500 m W of Seamans Hut, 1950 m, Kosciuszko National Park, 19 Feb. 1980, B. Barnsley 1287, J. Rymer & R. Jackson (CBG 8001431); ~2 km along Summit Road from Charlottes Pass, 1900 m, Kosciuszko National Park, 23 Dec. 1999, J.J. Bruhl 1872 & L.A. Craven (BOL, CANB 591878, CHR, HO 541646, K, MO, NE 70655, NSW 834196); Mount Kosciuszko, along track to summit, ~1 km beyond Charlottes Pass, Kosciuszko National Park, 19 Feb. 1990, M.G. Corrick 10667 (MEL 1578959); Daners Gap, Kosciusko region, 24 Feb. 1961, M. Gray 5035 (CANB n.v., NSW 462102); Snowy River bridge below Seamans Hut, Kosciuszko area, 6 Mar. 1968, M. Gray 6201 & C. Totterdell (CANB 478753, MEL 275470, NE 75489, NSW 687745); Mount Stilwell, Kosciuszko National Park, 18 Jan. 1951, L.A.S. Johnson & E. Constable s.n. (NSW 19610); Rawson Pass, 0.5 km E of Mount Kosciusko, Kosciuszko National Park, 19 Mar. 1974, A.N. Rodd 1613 (NSW 248289); Merritts Creek Valley, Kosciuszko National Park, 1950 m, 28 Feb. 1983, J. Thompson 4500 (NE 70795, NSW 433249); below Blue Lake, Kosciusko Plateau, 5 Feb. 1946, J.H. Willis s.n. (MEL 2066099); just E of Merrits Creek bridge, 1930 m, Kosciuszko National Park, 24 Jan. 2000, X. Zhang 14 & K.L. Wilson (CANB 559967, HO, MO, NE 72987, NSW 593158, NY); Schlink-Hilton Hut, Dicky Cooper Creek, 1800 m, Kosciuszko National Park, 25 Jan. 2000, X. Zhang 18 & K.L. Wilson (HO, MEL, NE 72991, NSW). Victoria: Snowfields: Sun Valley, Bogong High Plains, alt. 1610 m, 1 Mar. 1982, R.J. Adair 1644 (MEL 649163, NSW 462085); Wellington Plain, ~2.4 km NW of Mount Wellington, 7 Jan. 1973, A.C. Beauglehole 15748 & E.A. Chesterfield (MEL, NSW 462086); valley north of Basalt Hill, 7 km from Alpine Resorts Commission, Falls Creek Tourist Road, Bogong High Plains, 1617 m, 6 Feb. 1996, M.L. Cupper 08 (MEL 2066100); Bogong High Plains above Falls Creek, 1640 m, 21 Dec. 1999, J.R. Hosking 1830 (CANB 530728, MEL 2116813, NE 72565, NSW 442009); Bogong High Plains, Pretty Valley, 19 Jan. 1988, N.G. Walsh 1974, P.S. Short & M.C. Looker (MEL 1578805).
Carpha rodwayi W.M. Curtis, Brunonia 7(2): 302 (1984)
Type citation: ‘Holotypus: J. Davies s.n., Eliza plateau, south-west [sic] Tasmania, 4. ii. 1982, HO 53808.’ Type: Australia: Tasmania: South West: Mount Eliza Plateau, 4 Feb. 1982, J. Davies s.n. (holo: HO 53808; iso: NSW 687893).
Plants forming extensive mats, 7–12 cm high. Culms unnoded, rarely 1-noded with a single cauline leaf. Leaves crowded at the base of the culms; blades rigid, 2–4 cm long, 0.7–1.6 mm wide, thinly but stiffly crescentiform in cross-section; sheaths rich yellow-brown towards base. Inflorescence with 1 or 2 short internodes, 2–4 cm long; 4–9 spikelets in a loose cluster. Involucral bracts several, up to 2.6 cm long, up to 2 mm wide. Spikelets 10.5–11.5 mm long, with 1 bisexual flower; glumes 5, pale yellow-brown; lowest 3 glumes empty, lowest 2 glumes shorter than the rest; the fertile one 9–10.8 mm long, 1–1.9 mm wide; the uppermost one linear, sterile; axis not elongated above proximal fertile glume. Bristles 9.8–13 mm long, plumose with hairs 0.9–1.7 mm long; distal scabrous zone 1.5–2.7 mm long. Anthers pale yellow to pale grey, 2.9–3.3 mm long (excluding appendage), with apical appendage 0.1–0.4 mm long, 0.1–0.15 mm wide. Stylebase 3.0–6.5 mm long, 0.2–0.4 mm in diameter, glabrous. Nutlet 1 per spikelet, with a gynophore 0.3–0.5 mm long, the body narrow-ellipsoid, 3–3.5 mm long, 0.9–2 mm in diameter, pale orange-brown to mid-yellow-brown at maturity with 3 longitudinal ribs slightly paler or not paler, surface minutely reticulate, slightly puncticulate, cells more or less isodiametric. (Fig. 2d, i; see Zhang et al. 2004b, fig. 7b for spikelet structure.)
Endemic to the Central Highlands and South West regions of Tasmania, Australia (Fig. 8d). Forming extensive mats on the margins of alpine and subalpine pools, and in permanently wet montane areas.
Some specimens (e.g. Kirkpatrick s.n. (HO 91834) and Jarman s.n. (HO 411185) both determined to be C. rodwayi) appear to be morphologically intermediate between this species and C. alpina (e.g. Bruhl 1878A–E (NE 71803–71808) and Wilson 6643 (NE 70799, NSW 432013) both determined to be C. alpina). Further study is needed to clarify the limits of these species.
The species has a limited distribution but it occurs mostly in national parks, so is not considered to be at imminent risk; we suggest a status of ‘Least Concern’ (IUCN Standards and Petitions Committee 2022). However, the alpine habitat is limited and predicted to become even more so with changing climate, so the status is likely to change in the next few decades.
Australia: Tasmania: Central Highlands: Western Bluff, 1380 m, 2 Feb. 1983, A. Moscal 1640 (HO 62812). South West: NE edge of Mawson Plateau, just below main plateau, 1316 m, 15 Feb. 2000, J.J. Bruhl 1881A (CANB, HO 533168, K, NE 71815, NSW 658771); Mount Anne National Park, rise S side of saddle, below Mount Anne towards Mount Eliza, 1240 m, 16 Feb. 2000, J.J. Bruhl 1890A & A.C. Rozefelds (BM, HO, K, MO, NE 71834, NSW 658773, PRE); 0.5 km south-west of Nevada Peak, 25 Feb. 1990, P. Collier 4562 (HO 121972); Southwest National Park, Mount Eliza–Mount Anne saddle, 1250 m, 21 Feb. 1989, J.R. Croft 10237 & M. M. Richardson (AK, CBG 8904462, NSW 62088); Mount Mawson, Mount Field National Park, 26 Feb. 1977, A.T. Dobson 77245 (HO 30509); K Col, Mount Field, 10 Feb. 1960, W. D. Jackson s.n. (HO 24187, K).
Carpha schoenoides Banks & Sol. ex Hook.f., Fl. Antarct. 1(2): 362, t. 148 (1847)
Carpha alpina var. schoenoides (Banks & Sol. ex Hook.f.) Kük., Feddes Repert. 47: 113 (1939). Type citation: ‘Southern parts of Tierra del Fuego; Good Success Bay, Banks & Solander; Hermite Island, Cape Horn, J. D. H.’ Type: Tierra del Fuego, Jan. 1769, J. Banks & D. Solander (lecto, here designated: BM 000092178). Residual syn: Hermite Island, Cape Horn, 1839–1843, J.D. Hooker (BM 000092175); No. 71 Hermite Isd, [no date], J.D. H. [Hooker] (K).
Carpha viridis Phil., Anales Univers. Chile 27: 322 (1865). Type citation: ‘In palude quadam montium litoralium prov, Valdivia ‘Cordillera pelada’ dictarum invenimus.’
Type: Chile: Cordillera Pelada, Prov. Valdivia, Jan. 1865, F. Philippi s.n. (syn: SGO 000000804*, SGO 000000805*).
[Carpha alpina auct. non R.Br.: F.Philippi, Catal. Pl. Chilens. 306 (1881); C.B.Clarke, Bot. Jahrb. 30: 40 (1901); H.H.Pfeiffer, Feddes Repert. 29: 178 (1931), p.p.; Gunckel, Annales Mus. Hist. Nat. Valparaiso 4, 28 (1971); D.M.Moore, Fl. Tierra del Fuego 329 (1983).]
Tufted perennial, 18–42 cm high. Culms unnoded. Leaves crowded at the base of the culms; blades more or less straight, up to 20 cm long, 0.7–1.6 mm wide, thinly but stiffly crescentiform to thickly canaliculate in cross-section with an abaxial midrib; sheaths pale yellow-brown. Inflorescence with 1–3 short internodes, 3–10 cm long; 4–9 spikelets in a loose cluster or head, or not forming obvious heads. Involucral bracts several, up to 14 cm long, up to 1.4 mm wide. Spikelets 8.2–10 mm long, with 2 bisexual flowers; glumes 5 or 6, pale yellow-brown; lowest 3 glumes empty, lowest 2 glumes shorter than the rest; the proximal fertile glume 7.3–8.5 mm long, 1.7–2.4 mm wide; the second fertile glume 5.7–7.8 mm long, 0.9–1.5 mm wide; when spikelet has 6 glumes, the uppermost one linear, sterile; axis not elongated above proximal fertile glume. Bristles 6.5–9.8 mm long, plumose with hairs up to 1.2 mm long; distal scabrous zone 0.5–2.5 mm long. Anthers pale yellow to pale to dark greyish, 1.4–3.1 mm long (excluding appendage), with apical appendage 0.1–0.3 mm long, 0.05–0.1 mm wide. Stylebase 2.5–5 mm long, 0.15–0.3 mm in diameter, glabrous. Nutlets 2 per spikelet, with a gynophore 0.2–0.4 mm long, the body narrow-ellipsoid, 3–3.5 mm long, 0.7–1 mm in diameter, dark yellow-brown to dark grey-brown at maturity with 3 slightly paler longitudinal ribs, surface minutely reticulate, rarely minutely puncticulate, cells isodiametric to slightly elongated longitudinally (Fig. 2e, j; see Zhang et al. 2004b, fig. 7f, i, m, p, r for spikelet structure; Zhang et al. 2006, fig. 3c; Barros 1969, fig. 42a, b).
Occurring in temperate South America: in Patagonia (southern Chile and Argentina); south from ~40°S to Tierra del Fuego (Fig. 8e). In open marshes, on stream edges, open ground, and along forest roads.
After Carpha schoenoides was described by Hooker (1847), it was synonymised with C. alpina by Philippi (1881), who was followed in this by Clarke (1901), Pfeiffer (1931), Gunckel (1971) and Moore (1983). Kükenthal (1939a) treated it as a variety of C. alpina and was followed in this by Barros (1969). Our studies (Zhang 2002; Zhang et al. 2006, 2007) showed that C. schoenoides is better treated as a species. It differs from C. alpina in having two flowers in each spikelet, both developing nutlets that have an epidermis that is reticulate, not puncticulate (Zhang et al. 2007, fig. 3c), and in having somewhat shorter bristles with plumose hairs only up to 1.2 mm long (up to 2.5 mm in C. alpina). This species is geographically disjunct from other species of Carpha and probably results from long-distance dispersal from Australasia (Wilson 1986; Wardle et al. 2001; Viljoen et al. 2013).
The BM specimen from Hermite Island is on a sheet with another specimen consisting of two individual tufts collected by E. C. Reed in January 1872 from a different locality (Cord.[illera] Pelada, in Valdivia Province). The sheet in K is stamped as belonging originally to Hooker’s Herbarium, and bears Hooker’s manuscript label (‘No. 71, Hermite Isd, J.D.H.’), as well as pencilled drawings of the plant, and also two tufts collected by E. C. Reed in Jan 1872 from Cordillera Pelada. This would be appropriate to be lectotype except that the material is very depauperate, as is the apparent duplicate in BM. The Banks and Solander specimen in BM is in better condition and bears an original collecting label annotated ‘Carpha schoenoides’. It is therefore selected as lectotype.
We have seen only photographs of the two syntypes of Carpha viridis at SGO, so we are not selecting a lectotype. Both appear to belong to Carpha schoenoides.
Of limited distribution but occurs in national parks, so not considered to be at risk; we suggest a status of ‘Least Concern’ (IUCN Standards and Petitions Committee 2022).
Chile: summit of Cordillera de la Carpa, ~950 m, Province Osorno, 1–3 Feb. 1958, W.J. Eyerdam 10586A (K, NY); mainland, opposite Puerto Eden, 21 Dec. 1958, E.J. Godley 755a (BM 92177); Cordillera Pelada de la Cordillera de la Costa, 800 m, 22 Jan. 1924, P.A. Hollermayer 1334 (NY); Cord[illera] Pel[ada], Vald[ivia], presented July 1909, T. King (E 00514487); Isla Navarino, Pto Williams, Valle del Róbalo, 400 m, 14 Feb. 2005, A. Perez Haase BCN 27007 (NE 97980, NSW); in Province Valdivia, Cordillera Pelada, [no date], Philippi 981 (P 132670); Orange Harbour, 1838–42, US Exploring Expedition (K).
Argentina: Tierra del Fuego: Ruta J, between Estancia Moat and Estancia Puerto Rancho, 28 Dec. 2004, J.J. Bruhl 2243 & A. Plos (BOL, CHR, EIU, GENT, MO, NE 86464, NSW 930386, PRE, SI); Rio Azopardo, 200 m, 6 Mar. 1896, P. Dusén s.n. (AK 29810, BM 2176, MO 1626156); Bahia Aguirre, hills to E of Spaniard Harbor, 14 Feb. 1968, R.N.P. Goodall 1387 (US 2617434); mountain to NE of Rio Tristen, at head of Rio Tristen Valley, 28 Apr. 1974, R.N.P. Goodall 4811 (MO 2572272); Estancia Moat, 5 m, 30 Jan 2005, A. Perez Haase BCN 26993 (NE 87966, NSW 1002185); Bahía Aguirre, ~5 km E of Puerto Espagnol, 30–80 m, 14 Feb. 1968, D.M. Moore 1835 (K); W side of Río Varela, 500–600 m, 17 Feb. 1968, D.M. Moore 1925 (K, MO 2150322); Rio Fortune, 5 Mar. 1909, C. Skottsberg (B 10 0000962); Bahia Thetis, Mar. 1941, A.C. Umana 144 (NY).
Carpha aristata Kük., Feddes Repert. 47: 112 (1939)
Type citation: ‘Japan: Kiu-Schiu, bei Nagasaki (Schotmüller! in herb. mus. berol. pro Trianoptiles capensis Fenzl).’ Type: ?B, n.v.
Kükenthal (1939a) based this species on a poor (‘dürftiges’) specimen in B labelled as coming from Nagasaki, Kyushu, in Japan. Ohwi (1944, pp. 25–26) cited this species on the basis of Kükenthal’s description, but he suspected this species was not native to Japan and wrote ‘Hab. Kiushiu: Nagasaki (Schottmüller, non vid.). An planta certe in Japonia indigena?’ He also cited this species in Flora of Japan (Ohwi 1965, p. 212), but he noted that this species was ‘reported in Kyushu (Nagasaki) by Kükenthal.’ Toyama (1980, p. 269) also listed this species in The Flora of Nagasaki Prefecture, but he noted that the information came from Ohwi. Thus, no-one has apparently found this species growing in Japan.
The Berlin herbarium (B), where Kükenthal cited the type as being stored under the name Trianoptiles capensis, was contacted to check for the type or any other specimen labelled as Carpha aristata, but none was found. One of us (KLW) had not found any such specimen previously when she examined specimens of Cyperaceae in B.
We have also contacted some Japanese herbaria about specimens of this species. No such specimen has been located in Japanese herbaria, and it was suggested that Carpha aristata was probably an introduced plant at that time or that it may be extinct (H. Ohba, pers. comm., 2000). Another possible explanation is that the specimen was not collected in Japan. The label for this specimen may have been mixed up with collections from one of Schottmüller’s other collecting points during his long sea voyage as Second Botanist on the Prussian Expedition to East Asia (1860–1862), which visited Java, Singapore, the Philippines, China, Cape Town in South Africa, Rio de Janeiro in Brazil, and Madeira on its way to and from Japan (van Steenis Kruseman 1950).
Kükenthal (1939a) included this taxon in his key to Carpha sens. lat. (keyed out against A. borbonica, A. eminii and A. nitens) and gave a full description so he had obviously seen a complete specimen. It is possible that this specimen belongs to some African sedge species, and probably in or near Carpha sens. lat. because Kükenthal had an excellent knowledge of the family. However, we have not been able to match this with any other species by comparing the description of Carpha aristata with all of the species examined in our study and in a range of sedge floras, including species in other genera from South America, southern Africa and Asia.
Data availability
Most data that support this study are available as indicated in Zhang et al. (2004a, 2006, 2007); unpubl. data will be shared upon reasonable request to the corresponding author.
Conflicts of interest
Jeremy Bruhl is an associate editor for Australian Systematic Botany but did not at any stage have editor-level access to this manuscript while in peer review, as is the standard practice when handling manuscripts submitted by an editor to this journal. Australian Systematic Botany encourages its editors to publish in the journal and they are kept totally separate from the decision-making processes for their manuscripts. The authors have no further conflicts of interest to declare.
Declaration of funding
This study was undertaken while X. Zhang was the recipient of an Overseas Postgraduate Research Scholarship and a University of New England (UNE) Research Scholarship. She also acknowledges financial support from the N.C.W. Beadle Fund (Botany, UNE) and Joyce W. Vickery Scientific Research Fund (Linnean Society of New South Wales). The Australian Biological Resources Study provided support for K. L. Wilson and J. J. Bruhl as Australian Botanical Liaison Officers at the Royal Botanic Gardens Kew.
Acknowledgements
We acknowledge access to the facilities of N.C.W. Beadle Herbarium (NE) and the National Herbarium of New South Wales (NSW), crucial to the success of this study. The authors thank then Associate Professor Dr N. Prakash (UNE) for his valuable comments, and the directors and staff of AAU, B, BM, BOL, BRI, CANB, CGE, CHR, E, EA, F, FI, G, HBG, HO, K, MEL, MO, NU, NY, P, PE, PRE, US and Z for access to collections or loans. We also thank Mme Françoise Deluzarche at STR for information about Chermezon’s specimens in that herbarium; Dr Mats Hjertson at UPS for images of a Thunberg type specimen; Associate Professor Tony Verboom, Dr Cornelia Klak and Mr Dean Phillips at BOL for an image of the holotype of Asterochaete discolor; Dr Michaela Grein for information about specimens in BREM; Dr Tony Dold for information about specimens in GRA; Professor Muthama Muasya and Mr Dean Phillips for information about specimens in BOL; Dr Hidetoshi Nagamasu (Kyoto University Museum) for a copy of Toyama (1980); and Mr Alastair Wilson for translating key German publications. J. J. Bruhl thanks Dr Jane Browning and NU for assistance with fieldwork and extended access to facilities; the late Dick Brummitt for discussion of types; the Royal Botanic Gardens Kew for prolonged access to collection and library facilities as Australian Botanical Liaison Officer, and the University of New England for study leave. J. J. Bruhl and K. L. Wilson thank Dr Luc Gigord and his colleagues at the Conservatoire Botanique National de Mascarin, Réunion, for their assistance with fieldwork and laboratory resources. We also thank the co-collectors of the study group over many years (see specimens cited) and two reviewers for their comments.
References
Barrett RL, Wilson KL, Bruhl JJ (2019) Anthelepis, a new genus for four mainly tropical species of Cypereaceae from Australia, New Caledonia and South-East Asia. Australian Systematic Botany 32, 269-289.
| Crossref | Google Scholar |
Beentje HJ (2017) Carpha angustissima. In ‘The IUCN Red List of Threatened Species 2017’. e.T185599A84269279. (International Union for Conservationof Nature and Natural Resources) Available at https://www.iucnredlist.org/species/185599/84269279
Blake ST (1940) Notes on Australian Cyperaceae. III. Proceedings of the Royal Society of Queensland 51, 32-50.
| Crossref | Google Scholar |
Boeckeler O (1874) Die Cyperaceen des Königlichen Herbariums zu Berlin. Linnaea 38, 223-544 [In German and Latin].
| Google Scholar |
Brand RF, du Preez PJ, Brown LR (2013) High altitude montane wetland vegetation classification of the Eastern Free State, South Africa. South African Journal of Botany 88, 223-236.
| Crossref | Google Scholar |
Brochmann C, Gizaw A, Chala D, Kandziora M, Eilu G, Popp M, Pirie MD, Gehrke B (2021) History and evolution of the afroalpine flora: in the footsteps of Olov Hedberg. Alpine Botany 132, 65-87.
| Crossref | Google Scholar |
Browning J, Guthrie I (1994) Studies in Cyperaceae in southern Africa. 22: spikelet structure in African species of Carpha R.Br. South African Journal of Botany 60, 148-151.
| Crossref | Google Scholar |
Bruhl JJ (1991) Comparative development of some taxonomically critical floral/inflorescence features in Cyperaceae. Australian Journal of Botany 39, 119-127.
| Crossref | Google Scholar |
Bruhl JJ (1995) Sedge genera of the world: relationships and a new classification of the Cyperaceae. Australian Systematic Botany 8, 125-305.
| Crossref | Google Scholar |
Burgess ND, Butynski TM, Cordeiro NJ, Doggart NH, Fjeldså J, Howell KM, Kilahama FB, Loader SP, Lovett JC, Mbilinyi B, Menegon M, Moyer DC, Nashanda E, Perkin A, Rovero F, Stanley WT, Stuart SN (2007) The biological importance of the Eastern Arc Mountains of Tanzania and Kenya. Biological Conservation 134, 209-231.
| Crossref | Google Scholar |
Chermezon MH (1922) Sur quelques Cypéracées nouvelles de Madagascar. Bulletin de la Société Botanique de France 69, 719-725 [In French].
| Crossref | Google Scholar |
Chermezon H (1935) Cypéracées récoltées par M. Humbert au Congo belge oriental. Bulletin de la Société Botanique de France 82, 334-346 [In French].
| Crossref | Google Scholar |
Clarke CB (1901) Cyperaceae (praeter Caricinas) Chilenses. Beiblatt zu den Botanischen Jahrbüchern 30, 39-41.
| Google Scholar |
Clarke CB (1908) New genera and species of Cyperaceae. Bulletin of Miscellaneous Information, Kew Add. Ser. 8, 1-196.
| Google Scholar |
Contu S, Foden W, Potter L (2013) Carpha capitellata. In ‘The IUCN Red List of Threatened Species 2013’. T44393092A44448622. (International Union for Conservation of Nature and Natural Resources) Available at https://www.iucnredlist.org/species/44393092/44448622
Department of Environment, Land, Water and Planning (2021) Flora and Fauna Guarantee Act 1988 – Threatened list 2022: September 2022. (State of Victoria: Melbourne, Vic., Australia) Available at https://www.environment.vic.gov.au/__data/assets/pdf_file/0031/536089/FFG-Threatened-List-September-2022.pdf
Drège JF (1843) Zwei pflanzengeographische Documente. Flora 26(II, Suppl.), 1-230 [In German].
| Crossref | Google Scholar |
Drège JF (1847) Standörter-Verzeichniss der von C.L. Zeyher in Südafrika gesammelten Pflanzen. Linnaea 19, 853-889 [In German].
| Google Scholar |
Elliott TL, van Mazijk R, Barrett RL, Bruhl JJ, Joly S, Muthaphuli N, Wilson KL, Muasya AM (2021) Global dispersal and diversification of the genus Schoenus (Cyperaceae) from the Western Australian biodiversity hotspot. Journal of Systematics and Evolution 59, 791-808.
| Crossref | Google Scholar |
Foden W, Potter L (2010) Carpha glomerata. In ‘The IUCN Red List of Threatened Species 2010’. e.T185331A8390566. (International Union for Conservation of Nature and Natural Resources) Available at https://www.iucnredlist.org/species/185331/8390566
Galbany-Casals M, Unwin M, Garcia-Jacas N, Smissen RD, Susanna A, Bayer RJ (2014) Phylogenetic relationships in Helichrysum (Compositae: Gnaphalieae) and related genera: incongruence between nuclear and plastid phylogenies, biogeographic and morphological patterns, and implications for generic delimitation. Taxon 63, 608-624.
| Crossref | Google Scholar |
Galley C, Bytebier B, Bellstedt DU, Linder HP (2007) The Cape element in the Afrotemperate flora: from Cape to Cairo? Proceedings of the Royal Society B: Biological Sciences 274, 535-543.
| Crossref | Google Scholar | PubMed |
Gehrke B, Linder HP (2009) The scramble for Africa: pan-temperate elements on the African high mountains. Proceedings of the Royal Society B: Biological Sciences 276, 2657-2665.
| Crossref | Google Scholar | PubMed |
Gehrke B, Linder HP (2014) Species richness, endemism and species composition in the tropical Afroalpine flora. Alpine Botany 124, 165-177.
| Crossref | Google Scholar |
Green AJ, Lovas‐Kiss Á, Reynolds C, Sebastián‐González E, Silva GG, van Leeuwen CHA, Wilkinson DM (2023) Dispersal of aquatic and terrestrial organisms by waterbirds: a review of current knowledge and future priorities. Freshwater Biology 68, 173-190.
| Crossref | Google Scholar |
Gunckel H (1971) Revision sistematica de las especies Chilenas de la subfamilia de las Rhynchosporoides de las Ciperaceas. Annales del Museo de Historia Natural Valparaiso 4, 15-52 [In Spanish].
| Google Scholar |
Hiepko P (1987) The collections of the Botanical Museum Berlin-Dahlem (B) and their history. Englera 7, 219-252.
| Google Scholar |
Hinchliff CE, Roalson EH (2013) Using supermatrices for phylogenetic inquiry: an example using the sedges. Systematic Biology 62, 205-219.
| Crossref | Google Scholar | PubMed |
IUCN Standards and Petitions Committee (2022) Guidelines for using the IUCN Red List categories and criteria. Version 15.1. (Standards and Petitions Committee, International Union for Conservation of Nature and Natural Resources) Available at https://www.iucnredlist.org/documents/RedListGuidelines.pdf
Johnson PN, Campbell DJ (1975) Vascular plants of the Auckland Islands. New Zealand Journal of Botany 13, 665-720.
| Crossref | Google Scholar |
Jordan GJ (2001) An investigation of long-distance dispersal based on species native to both Tasmania and New Zealand. Australian Journal of Botany 49, 333-340.
| Crossref | Google Scholar |
Jung J, Choi H-K (2013) Recognition of two major clades and early diverged groups within the subfamily Cyperoideae (Cyperaceae) including Korean sedges. Journal of Plant Research 126, 335-349.
| Crossref | Google Scholar | PubMed |
Kandziora M, Kadereit JW, Gehrke B (2016) Frequent colonization and little in situ speciation in Senecio in the tropical alpine-like islands of eastern Africa. American Journal of Botany 103, 1483-1498.
| Crossref | Google Scholar | PubMed |
Kandziora M, Gehrke B, Popp M, Gizaw A, Brochmann C, Pirie MD (2022) The enigmatic tropical alpine flora on the African sky islands is young, disturbed, and unsaturated. Proceedings of the National Academy of Sciences 119, e2112737119.
| Crossref | Google Scholar | PubMed |
Krauss CF (1845) Pflanzen des Cap- und Natal-Landes, gesammelt und zusammengestellt von Dr Ferdinand Krauss (Fortsetzung). Flora 28, 753-764 [In German].
| Google Scholar |
Kükenthal G (1939a) Vorarbeiten zu einer Monographie der Rhynchosporoideae. 6. Feddes Repertorium 47, 101-119 [In German].
| Google Scholar |
Kükenthal G (1939b) Vorarbeiten zu einer Monographie der Rhynchosporoideae. 7. Feddes Repertorium 47, 209-216 [In German].
| Google Scholar |
Larridon I, Zuntini AR, Léveillé‐Bourret É, Barrett RL, Starr JR, Muasya AM, Villaverde T, Bauters K, Brewer GE, Bruhl JJ, Costa SM, Elliott TL, Epitawalage N, Escudero M, Fairlie I, Goetghebeur P, Hipp AL, Jiménez‐Mejías P, Sabino Kikuchi IAB, Luceño M, Márquez‐Corro JI, Martín‐Bravo S, Maurin O, Pokorny L, Roalson EH, Semmouri I, Simpson DA, Spalink D, Thomas WW, Wilson KL, Xanthos M, Forest F, Baker WJ (2021) A new classification of Cyperaceae (Poales) supported by phylogenomic data. Journal of Systematics and Evolution 59, 852-895.
| Crossref | Google Scholar |
Levyns MR (1943) A revision of Trianoptiles Fenzl. Journal of South African Botany 9, 21-26.
| Google Scholar |
Levyns MR (1947) Tetraria and related genera, with special reference to the flora of the Cape Peninsula. Journal of South African Botany 13, 73-93.
| Google Scholar |
Linder HP (2003) The radiation of the Cape Flora, Southern Africa. Biological Reviews 78, 597-638.
| Crossref | Google Scholar | PubMed |
Linder HP, Verboom GA (2015) The evolution of regional species richness: the history of the Southern African flora. Annual Review of Ecology, Evolution and Systematics 46, 393-412.
| Crossref | Google Scholar |
Luke WRQ (2010) Carpha eminii. In ‘The IUCN Red List of Threatened Species 2010’. e.T185319A8388165. (International Union for Conservation of Nature and Natural Resources) Available at https://www.iucnredlist.org/species/185319/8388165
McGillivray DJ (1973) Michel Gandoger’s names of Australian plants. Contributions from the New South Wales National Herbarium 4, 31-359.
| Google Scholar |
Metzing D (1999) Das Herbarium des Bremer Uebersee-Museums Geschichte, Sammlungen und Bedeutung. Abhandlungen des Naturwissenschaftlichen Vereins zu Bremen 44, 771-790 [In German].
| Google Scholar |
Muasya AM, Simpson DA, Verboom GA, Goetghebeur P, Naczi RFC, Chase MW, Smets E (2009) Phylogeny of Cyperaceae based on DNA sequence data: current progress and future prospects. The Botanical Review 75, 2-21.
| Crossref | Google Scholar |
Muñoz J, Felicísimo AM, Cabezas F, Burgaz AR, Martínez I (2004) Wind as a long-distance dispersal vehicle in the Southern Hemisphere. Science 304, 1144-1147.
| Crossref | Google Scholar | PubMed |
Napper D (1964) Cyperaceae of East Africa II. Journal of the East Africa Natural History Society and Coryndon Museum 24, 23-46.
| Google Scholar |
Nathan R, Schurr FM, Spiegel O, Steinitz O, Trakhtenbrot A, Tsoar A (2008) Mechanisms of long-distance seed dispersal. Trends in Ecology & Evolution 23, 638-647.
| Crossref | Google Scholar | PubMed |
Nees CG (1834) Uebersicht der Cyperaceengattungen. Linnaea 9, 273-306 [In German].
| Google Scholar |
Nees CG (1835) Cyperaceae capenses, secundum novissiimas Ecklonii collectiones. Linnaea 10, 129-270 [In Latin].
| Google Scholar |
Ohwi J (1944) Cyperaceae Japonicae II. A synopsis of the Rhynchosporoideae and Scirpoideae of Japan, including the Kuriles, Saghalin, Korea and Formosa. Memoirs of the College of Science, Kyoto Imperial University, Series B 18, 1-182.
| Google Scholar |
Pfeiffer H (1931) Decas Cyperacearum criticarum vel emendatarum III. Repertorium Novarum Specierum Regni Vegetabilis 29, 171-186 [In German].
| Crossref | Google Scholar |
Pócs T (1976) Vegetation mapping in the Uluguru Mountains (Tanzania, East Africa). Boissiera 24, 477-498.
| Google Scholar |
Reid C, Arnold TH (1984) Cyperaceae. Bothalia 15, 139-142.
| Crossref | Google Scholar |
Sanmartín I, Ronquist F (2004) Southern Hemisphere biogeography inferred by event-based models: plant versus animal patterns. Systematic Biology 53, 216-243.
| Crossref | Google Scholar | PubMed |
Sanmartín I, Wanntorp L, Winkworth RC (2007) West Wind Drift revisited: testing for directional dispersal in the Southern Hemisphere using event-based tree fitting. Journal of Biogeography 34, 398-416.
| Crossref | Google Scholar |
Semmouri I, Bauters K, Léveillé-Bourret É, Starr JR, Goetghebeur P, Larridon I (2019) Phylogeny and systematics of Cyperaceae, the evolution and importance of embryo morphology. The Botanical Review 85, 1-39.
| Crossref | Google Scholar |
Thompson J (1981) A key to the plants of the subalpine and alpine zones of the Kosciusko region. Telopea 2, 219-297.
| Crossref | Google Scholar |
Thompson J, Gray M (1981) A check-list of the subalpine and alpine species found in the Kosciusko region of New South Wales. Telopea 2, 299-346.
| Crossref | Google Scholar |
Turland NJ, Wiersema JH, Barrie FR, Greuter W, Hawksworth DL, Herendeen PS, Knapp S, Kusber W-H, Li D-Z, Marhold K, May TW, McNeill J, Monro AM, Prado J, Price MJ, Smith GF (Eds) (2018) ‘International Code of Nomenclature for algae, fungi, and plants (Shenzhen Code) adopted by the Nineteenth International Botanical Congress Shenzhen, China, July 2017. Regnum Vegetabile Vol. 159.’ (Koeltz Botanical Books: Glashütten, Germany)
van Santen M, Linder HP (2020) The assembly of the Cape flora is consistent with an edaphic rather than climatic filter. Molecular Phylogenetics and Evolution 142, 106645.
| Crossref | Google Scholar | PubMed |
van Steenis Kruseman MJ (1950) Cyclopaedia of collectors. In ‘Flora Malesiana. Vol. 1, Ser. 1’. (Noordhoff-Kolle N.V.: Djakarta, Indonesia) Available at http://www.nationaalherbarium.nl/FMCollectors/ [Verified 18 April 2024]
Verboom GA (2006) A phylogeny of the schoenoid sedges (Cyperaceae: Schoeneae) based on plastid DNA sequences, with special reference to the genera found in Africa. Molecular Phylogenetics and Evolution 38, 79-89.
| Crossref | Google Scholar | PubMed |
Verboom GA, Bergh NG, Haiden SA, Hoffmann V, Britton MN (2015) Topography as a driver of diversification in the Cape Floristic Region of South Africa. New Phytologist 207, 368-376.
| Crossref | Google Scholar | PubMed |
Viljoen J-A, Muasya AM, Barrett RL, Bruhl JJ, Gibbs AK, Slingsby JA, Wilson KL, Verboom GA (2013) Radiation and repeated transoceanic dispersal of Schoeneae (Cyperaceae) through the Southern Hemisphere. American Journal of Botany 100, 2494-2508.
| Crossref | Google Scholar | PubMed |
Wagstaff SJ, Breitwieser I, Swenson U (2006) Origin and relationships of the austral genus Abrotanella (Asteraceae) inferred from DNA sequences. Taxon 55, 95-106.
| Crossref | Google Scholar |
Wardle P, Ezcurra C, Ramírez C, Wagstaff S (2001) Comparison of the flora and vegetation of the southern Andes and New Zealand. New Zealand Journal of Botany 39, 69-108.
| Crossref | Google Scholar |
Wilson KL (1994b) New taxa and combinations in the family Cyperaceae in eastern Australia. Telopea 5, 589-625.
| Google Scholar |
Winkworth RC, Wagstaff SJ, Glenny D, Lockhart PJ (2002) Plant dispersal N.E.W.S. from New Zealand. Trends in Ecology & Evolution 17, 514-520.
| Crossref | Google Scholar |
Wu Z-Y, Milne RI, Liu J, Nathan R, Corlett RT, Li D-Z (2023) The establishment of plants following long-distance dispersal. Trends in Ecology & Evolution 38, 289-300.
| Crossref | Google Scholar | PubMed |
Zhang X, Marchant A, Wilson KL, Bruhl JJ (2004a) Phylogenetic relationships of Carpha and its relatives (Schoeneae, Cyperaceae) inferred from chloroplast trnL intron and trnL–trnF intergenic spacer sequences. Molecular Phylogenetics and Evolution 31, 647-657.
| Google Scholar |
Zhang X, Wilson KL, Bruhl JJ (2004b) Sympodial structure of spikelets in the tribe Schoeneae (Cyperaceae). American Journal of Botany 91, 24-36.
| Google Scholar |
Zhang X, Wilson KL, Bruhl JJ (2006) Species limits in Carpha (Schoeneae, Cyperaceae) based on phenetic analyses. Australian Systematic Botany 19, 437-465.
| Crossref | Google Scholar |
Zhang X, Bruhl JJ, Wilson KL, Marchant A (2007) Phylogeny of Carpha and related genera (Schoeneae, Cyperaceae) inferred from morphological and molecular data. Australian Journal of Botany 20, 93-106.
| Google Scholar |