

Responses of grass productivity traits to bush clearing in semi-arid rangelands in North-West Province of South Africa
Mthunzi Mndela A * , Ignacio C. Madakadze B , Julius T. Tjelele A , Mziwanda Mangwane A B , Florence Nherera-Chokuda C , Sikhalazo Dube D , Abel Ramoelo E and Ngoako L. Letsoalo AA Agricultural Research Council, Irene 0062, Pretoria, South Africa.
B Department of Plant and Soil Sciences, University of Pretoria, Hatfield 0028, Pretoria, South Africa.
C NERPO, Pretoria 0081, Pretoria, South Africa.
D International Livestock Research Institute (ILRI), Mount Pleasant, Harare, Zimbabwe.
E Department of Geography, Geoinformatics and Meteorology, University of Pretoria, Hatfield 0028, South Africa.
The Rangeland Journal 44(1) 33-45 https://doi.org/10.1071/RJ21053
Submitted: 26 October 2021 Accepted: 13 February 2022 Published: 17 March 2022
© 2022 The Author(s) (or their employer(s)). Published by CSIRO Publishing on behalf of the Australian Rangeland Society. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Woody plant encroachment threatens herbaceous plant productivity in many rangelands globally. We evaluated the impact of bush clearing on grass tiller, leaf and biomass production, and tuft sizes in the Kgomo-kgomo and Makapaanstad rangelands in North-West Province, South Africa. In each rangeland, the number of tillers and leaves, tuft sizes and biomass of eight dominant grass species were recorded in bush-cleared and uncleared treatments. The treatment and species interacted significantly (P < 0.001) for tiller and leaf production and tuft sizes. Bush clearing increased tiller production of bunch grasses but not stoloniferous grasses. At Kgomo-kgomo, bunch grasses (Panicum maximum (Jacq.) and Urochloa mosambicensis (Hack.) Dandy] had three to six times more tillers and leaves per plant in the cleared than uncleared treatment. At Makapaanstad, only annual bunch grasses [Brachiaria eruciformis (Sibth. & Sm.) Griseb and Tragus berteronianus (Schult.)] attained twice as many tillers and leaves per plant in the cleared compared to uncleared treatment. Biomass was 1776 ± 159 and 696 ± 159 g m−2 in cleared and uncleared treatments respectively at Kgomo-kgomo and 1358 ± 258 and 1089 ± 258 g m−2 at Makapaanstad. The tufts of bunch grasses were nearly twice as large in the cleared compared with the uncleared treatment at Kgomo-kgomo, whereas only stoloniferous grass tufts increased at Makapaanstad. Overall, bush clearing improved grass productivity and performance, but the responses varied by species.
Keywords: biomass production, bunch and stoloniferous grasses, bush clearing, leaves, South Africa, tillers, tuft sizes, woody plant encroachment.
Introduction
Herbaceous cover and productivity of South African semi-arid rangelands are threatened by rapid and extensive woody-plant encroachment (WPE), with WPE affecting almost 7.3 million hectares (Warren et al. 2018). Effects of WPE on grass productivity are most notable when woody cover and density exceed 40% and 2400 trees ha−1 respectively, in southern Africa (Roques et al. 2001). Tree-grass competition for light, soil water and nutrients favours woody plants over grasses, which reduces grass productivity (Simmons et al. 2008; Hare et al. 2020) and causes a decline in livestock carrying capacity (Hare et al. 2020).
Grasses employ differential evolutionary and life-history strategies to cope with WPE (Solofondranohatra et al. 2018; Holub et al. 2019; Woon et al. 2021). Clonal grasses may persist in bush-encroached rangelands through vegetative reproduction (O’Connor et al. 2020). These grasses exhibit a network of ramets that emerge on stolons or rhizomes and radiate from the main genet to acquire limited resources (Ye et al. 2014; Saixiyala et al. 2017). Conversely, high woody cover induces premature senescence (Bassett et al. 2011) and reduces tiller and leaf production (Volder et al. 2013), reducing grass productivity because of shade-induced reduction in bunch grass meristematic activity (Gomes et al. 2020).
Bush clearing is crucial to modify microclimate and increase soil water and nutrient availability for shade-intolerant bunch grasses (Stephens et al. 2016) and for increasing forage production (Hare et al. 2021). Reduction in woody cover and density facilitates tillering from dormant grass buds (O’Connor et al. 2020). Pierson et al. (1990) found higher tiller and leaf turnover, which translated to higher biomass production under unshaded than a shaded environment. However, most shading studies are conducted in glasshouses and do not mimic field conditions. Hence, a knowledge gap exists regarding how bush clearing alters grass productivity at a tiller and leaf level.
Although a national initiative, Working for Water, directed to control woody plant encroachment and invasion, was launched in 1995 by the Department of Water Affairs and Forestry (Van Wilgen et al. 2020), this programme does not monitor grass performance, such as tillering, leaf production and plant vigour, in response to bush clearing. Thus, this current study intended to address the following questions in semi-arid rangelands: (1) how does bush clearing affect grass tiller, leaf and biomass production, and tuft sizes; and (2) does grass leaf production depend on tillering following bush clearing?
Materials and methods
Site description
The study was conducted at the Makapaanstad and Kgomo-kgomo communal rangelands in the Bojanala District Municipality of the North-West Province in South Africa (Fig. 1). Collectively, these rangelands cover 16 400 ha at 900–1200 m above sea level (Mucina et al. 2006). The average annual rainfall is 459 mm annum−1 (Moerane 2013). The highest rainfall occurs from mid-summer to mid-autumn (December to March) in the study areas (Supplementary Fig. S1). Maximum monthly average temperatures range from 27°C to 34°C in summer and from 20°C to 23°C in winter, and respective minimums range from 15°C to 16°C in summer and from 3°C to 6°C in winter (DIGES 2012). The main soil types are vertisols at Makapaanstad and red-yellow apedal sandy soils at Kgomo-kgomo (Mucina et al. 2006). Vertisols (vertic clays) have a very loose structure (tending to crack when dry and shrink when wet), a high calcium carbonate content and a high cation exchange capacity, whereas red-yellow apedal soils are well drained sandy soils with a high base status (Fey 2010). These rangelands occur in the Central Mixed Bushveld bioregion in Springbokvlakte thornveld (Mucina et al. 2006). The tree layer is dominated by leguminous shrubs, predominantly a mixture of Dichrostachys cinerea (L.) Wight & Arn and Vachellia tortilis (Willd.) Hayne at Kgomo-kgomo (Mndela et al. 2019) and a homogenous layer of Vachellia tenuispina (I. Verd.) Kyal & Boatwr at Makapaanstad (Mndela et al. 2020). The Kgomo-kgomo and Makapaanstad rangelands are encroached at woody plant densities of 4850 and 6900 trees ha−1 respectively (Mndela et al. 2019, 2020). The woody canopy cover is 76% and 82% at Kgomo-kgomo and Makapaanstad rangelands respectively. These rangelands received bush clearing from 2012 to 2017 (Fig. 1c, d), but, for the purpose of this study, we established new experimental plots. Both rangelands are continuously grazed throughout the year by cattle, sheep and goats. However, the experimental plots were fenced immediately after bush clearing in April 2016 to prohibit grazing throughout the study period.

|
Experimental layout and sampling
Pre-treatment layout and species selection
In each rangeland, three blocks of similar woody vegetation type, soil type, and topography were randomly selected using aerial photographs in January 2016. In each block, an area of 0.25 ha was demarcated in which eight 5 × 5 m plots were permanently marked. The eight plots were divided into two rows, 30 m apart, with plots in each row interspaced by 5 m. Three 1.3 × 1.3 m quadrats were sampled in two opposite corners and at the centre of each plot, giving a total of 144 quadrats (3 quadrats × 8 plots × 3 blocks × 2 rangelands).
Herbaceous species were identified in the quadrats to determine species composition and calculate relative abundance (RA) for selection of the eight most dominant grass species in each rangeland. Relative abundance was calculated (Zakaria et al. 2009) as:
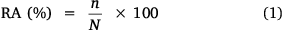
where n = total number of individuals of a species and N = total number of individuals of all species.
The selected grass species were monitored for the responses to bush clearing (Table 1). Of the eight selected species per rangeland, at least four were bunch grasses (Van Oudtshoorn 1999), and the others were stoloniferous. The species were subjectively classified according to the dominant growth habit on the basis of visual judgement during vegetation survey. Thus, although Digitaria eriantha (Stent.) Steud and Panicum coloratum L. are tufted, they relied more on clonal growth through spreading stolons, and so were classified as stoloniferous grasses. The selected species in each rangeland comprised two annuals and six perennials (Table 1). After vegetation assessment, the plots were mowed to 5 cm to avoid the carryover effects of residual biomass.

|
Post-treatment design and sampling
In April 2016, after a pre-treatment vegetation survey, half of each of the 0.25 ha blocks was cleared, and the adjacent half left uncleared, resulting in three replicates of cleared and uncleared treatments. In each replicate per treatment, four 5 × 5 m plots were marked. The three 1.3 × 1.3 m quadrats were sampled in each plot as for the pre-treatment sampling. The experiment was a completely randomised block design.
Tillers and leaves of the eight selected grass species were counted for individuals encountered in the quadrats during peak production in February 2017. The longest and the shortest tuft/crown diameters were measured and averaged to estimate a representative diameter. For stoloniferous grasses, the tiller and leaf counts, and tuft measurements, were conducted in every group of shoots emerging from each ramet attachment to the ground. The directions of the ramets were tracked from the genet within the quadrat and each measurement was conducted from those ramets assigned to the main genet by dividing the diameters of all ramet attachments with the number of attachments from which measurements were taken. Thereafter, grasses were clipped at 5 cm above the ground level and biomass was then sorted by species, oven-dried at 75°C to a constant weight and dry matter per sample determined.
Bush clearing method
Woody plants in the cleared treatment were cut 9 cm above the soil surface by using chainsaws and loppers and removed. Stumps were treated with Browser® herbicide, containing picloram (4-amino-3,5,6-trichloro-2-pyridinecarboxylic acid) as the active ingredient (240 g L−1). The chemical was mixed with crop oil for cut stump treatment and water for resprout control at the recommended concentration of 1%. A minor woody plant resprouting occurred during the rainy season. Resprouts were sprayed using the herbicide after vegetation sampling at the end of February 2017 to avoid chemical interference with herbaceous vegetation.
Statistical analysis
The analysis was conducted in SAS 9.4 (SAS Institute Inc., Cary, NC, USA, 2009). The univariate analysis was performed using Kolmogorov–Smirnov and Levene’s tests to assess data normality and homoscedasticity respectively (Pearce and Derrick 2019). Tiller and leaf numbers were transformed using square root ( ), whereas log10(x + 1) transformation was applied for tuft size. Thereafter, two-way ANOVAs were generated using mixed models, with paired treatments (n = 2) and grass species (n = 8) and their interactions included as fixed factors, whereas a block (n = 3) was used as a random factor. The plots were nested within the blocks. The model was expressed as follows:
), whereas log10(x + 1) transformation was applied for tuft size. Thereafter, two-way ANOVAs were generated using mixed models, with paired treatments (n = 2) and grass species (n = 8) and their interactions included as fixed factors, whereas a block (n = 3) was used as a random factor. The plots were nested within the blocks. The model was expressed as follows:
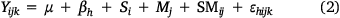
where, Y = dependent variables (numbers of tillers and leaves, biomass and tuft size), μ = overall mean, β = effect of the hth block, Si = fixed effect of the ith species, Mj = fixed effect of the Jth treatment (bush-cleared and uncleared), SMij = effect of the species by treatment interaction and εijk = experimental error associated with block, species and treatment.
Although the rangelands (n = 2) experience similar climatic conditions, there was no basis to compare them because of differences in soil and vegetation types. The means were separated using a paired Student’s t-test, with significant differences noted at α = 0.05. The means and standard errors were back-transformed to the original scale after analysis. We further conducted linear regression analysis to ascertain the bivariate relationships of tiller and leaf production for each grass species by using JMP software (SAS Institute 2015). We used root mean-square error (RMSE) and coefficient of determination (r2) to measure the precision with which tiller production predicted leaf production.
Results
Tiller production
At Kgomo-kgomo, treatment and grass species interacted significantly (F7,40 = 21.23, P < 0.001) on tiller number per plant of grass species. The number of tillers of P. maximum (t = 10.14, P < 0.001), U. mosambicensis (t = 12.77, P < 0.001) and S. pappophoroides (t = 6.03, P < 0.001) were three- to six-fold higher in the cleared than uncleared treatment and T. berteronianus (t = 3.18, P = 0.005) and A. barbicollis (t = 6.27, P < 0.001) had twice as many tillers in the cleared treatment (Fig. 2). Bush clearing increased tiller numbers of D. eriantha, but had no significant effects (P = 0.370) on other stoloniferous species (E. pseudoschlerantha and C. dactylon; Fig. 2).

|
Treatment and species also interacted significantly (F7,40 = 2.81, P = 0.026) on grass tiller production at Makapaanstad. Bush clearing significantly altered tiller production of annual bunch grasses [B. eruciformis (t = 2.04, P = 0.04) and T. berteronianus (t = 3.15, P = 0.004) Fig. 2]. The number of tillers of B. eruciformis and T. berteronianus were 1.5–1.9-fold higher in cleared compared to uncleared treatment. There was no treatment effect on other species (P = 0.076; Fig. 2).
Leaf production
The treatment and grass species interacted significantly (F7,40 = 28.51, P < 0.001) on leaf numbers of grasses at Kgomo-kgomo. The bunch grasses (Panicum maximum, S. pappophoroides and U. mosambicensis) had significantly (P < 0.001) more leaves in cleared than uncleared treatment. Among stoloniferous grasses, E. pseudoschlerantha (t = 2.78, P = 0.011) and D. eriantha (t = 2.84, P = 0.011) responded positively to clearing, whereas C. dactylon responded negatively (Fig. 3). Tragus berteronianus (t = 5.15, P < 0.001) and A. barbicollis (t = 5.95, P < 0.001) had double the number of leaves in cleared compared with uncleared treatment (Fig. 3). The treatment by species interaction on leaf production was also significant at Makapaanstad (F7,40 = 5.68, P < 0.001). Aristida bipartita had significantly (t = 2.44, P = 0.022) more leaves in cleared than uncleared treatment (Fig. 3). The stoloniferous grasses (C. dactylon and B. insculpta) had 1.5–1.9-fold more leaves and annual bunch grasses (T. berteronianus and B. eruciformis) had two to three times more leaves in cleared than uncleared treatment (Fig. 3).

|
Biomass
Treatment by species interaction was not significant (P = 0.180), but treatment (P = 0.014) and species had a significant (P < 0.001) effect on biomass at Kgomo-kgomo. The cleared treatment had twice the biomass of the uncleared treatment (Fig. 4). Panicum maximum had a significantly higher biomass on both an area and per plant basis than did other grass species (Fig. 5). Cynodon dactylon and A. barbicollis had the lowest biomass on both per plant and an area basis compared with the other species (Fig. 5). Eragrostis pseudoschlerantha, D. eriantha and U. mosambicensis had a comparable biomass on both per plant and an area basis (Figs 4, 5). On per plant basis, Schmidtia pappophoroides had the lowest biomass; however, on per area basis, its biomass was comparable to other species except for P. maximum and U. mosambicensis (Fig. 5).

|

|
Likewise, treatment and species had no significant (P = 0.235) interaction at Makapaanstad. However, the treatment (P = 0.011) and species (P < 0.0001) independently had a significant effect on biomass, with cleared treatment having a higher biomass than uncleared treatment on per area basis (Fig. 4). In contrast, when biomass per plant was averaged across species, cleared treatment had a lower biomass than did uncleared treatment (Fig. 4). On an area basis, E. lehmanniana generally produced more biomass than did other species (Fig. 5), whereas on per plant basis, A. bipartita attained a higher biomass (Fig. 5). However, these differences were not always significant.
Relationships between tiller and leaf production
At both Kgomo-kgomo and Makapaanstad, there were significant positive relationships (P < 0.001) between tiller and leaf production (Figs 6, 7). The relationships were strongest in cleared relative to uncleared treatment in both sites for stoloniferous grasses (Figs 6, 7), with their slopes (2–6 leaves tiller−1) being steeper than for bunch grasses (1–3 leaves tiller−1; Table 2). Leaf production was predicted from tillers with better accuracy for stoloniferous grasses than for bunch grasses in cleared treatment (Table 2). Except for A. bipartita and T. berteronianus at Makapaanstad, leaves of bunch grasses were better predicted from tillers (Table 2) and correlated more strongly in uncleared than cleared treatment in both rangelands (Figs 6, 7).

|

|

|
Tuft size
There was a significant treatment by species interaction on tuft size of grass species at Kgomo-kgomo (F7,42 = 1.45, P < 0.01). Tufts of U. mosambicensis (t = 3.27, P = 0.003), P. maximum (t = 2.83, P = 0.009) and T. berteronianus were two-fold larger in cleared than uncleared treatments (Fig. 8). At Makapaanstad, there was also a significant treatment by species interaction on tuft sizes (F7,42 = 3.81, P = 0.005). Bush clearing significantly (P = 0.003) increased only the tuft sizes of stoloniferous species (Fig. 8). However, P. coloratum tufts were significantly (P = 0.039) smaller in the cleared than in uncleared treatment (Fig. 8). Conversely, D. eriantha (t = 2.33, P = 0.028) and C. dactylon (t = 3.83, P < 0.001) respectively had two- and six-fold larger tufts in the cleared treatment (Fig. 8).

|
Discussion
The impact of bush clearing on tiller production varies by grass species
The effects of clearing on tiller production were species-dependent, depending largely on growth habits at Kgomo-kgomo and the lifespan of selected grass species at Makapaanstad. Bunch grasses responded most positively to treatment, most likely because stoloniferous grasses acquire, through clonal ramets, resources in fertile islands under tree canopies and between trees (Saixiyala et al. 2017). This result is consistent with Lett and Knapp (2003) who recorded low tiller densities of a rhizomatous grass following shrub control. Conversely, greater bunch grass tillering in the cleared treatment confirmed that bush encroachment can suppress grass productivity. Similarly, Pierce et al. (2019) recorded more axillary tillers in the grass growing in bush-cleared than uncleared shrublands.
At Kgomo-kgomo, clearing altered tiller production of shade-intolerant species and of P. maximum and S. pappophoroides (Fig. 2), the shade-tolerant perennial grasses (Van Oudtshoorn 1999), suggesting that tillering in uncleared treatments is largely governed by below-ground competition. In support, Simmons et al. (2008) reported that below-ground competition between Prosopis glandulosa and Nassella leucotricha was a more important determinant of grass tillering than was light. However, this did not seem to be true for E. lehmanniana and A. bipartita at Makapaanstad because they did not show a positive response to clearing.
Eragrostis lehmanniana and A. bipartita proliferated in dense swards in the clumps of encroaching shrubs. This could indicate a facilitative relationship between the encroacher shrub and these grass species, perhaps through soil fertility amelioration and protection from grazers (Pierce et al. 2019). Only annual-grass tiller production (B. eruciformis and T. berteronianus) was responsive to clearing at Makapaanstad. However, tuft sizes were comparable between treatments, indicating a partial reliance on aerial tillering.
The impact of bush clearing on leaf production varies by grass species
Leaf production responses were governed by interactions between treatment and species. Stoloniferous-grass leaf production, e.g. C. dactylon, responded positively to clearing regardless of limited tillering. Results suggest that leaf production of stoloniferous grasses, to some degree, is a function of a higher leaf production per tiller. The emergence of spaced clustered shoots having more phytomers along the stolons is common in stoloniferous grasses (da Silva et al. 2015). Furthermore, our data indicated that stoloniferous grasses produced three to six leaves tiller−1 (Table 2), suggesting that these species invested more in higher leaf production per tiller. This is a common strategy in grasses, especially where competition among plants is high (Irving 2015). Unlike bunch grasses, stoloniferous species do not exhibit self-shading because of the lateral spread of ramets (Solofondranohatra et al. 2018). As expected, bunch grasses produced numerous leaves in the cleared relative to the uncleared treatment. This highlights that grasses employ differential response strategies to clearing.
Leaf production of grasses depends largely on tillering
Leaf production was positively related to tiller production in both cleared and uncleared treatments (Figs 6, 7). Bunch grasses produced more leaves in the cleared treatment, which led to dense and compact swards. Hence, leaves at the bottom of these grass swards were shaded, with some senesced leaves, which presumably increased prediction error (RMSE) for bunch grasses (see Table 2). Similarly, young tillers of bunch grasses may suffer similar effects of shading (Irving 2015) and intra-tiller competition within the sward (Tomlinson and O’Connor 2004; Gomes 2020). Thus, if leaf mortality surpasses leaf emergence, the linear correlation between tillering and leaf production is likely to be weakened for bunch grasses. Hence, well timed lenient defoliation to ensure access to light for all vegetative sward components may be necessary (Pereira et al. 2017).
Conversely, with space stoloniferous grasses spread effectively through clonal ramets to access limited resources (Saixiyala et al. 2017). Thus, as indicated by steeper slopes (Table 2), stoloniferous grasses produced more leaves per tiller than did bunch grasses that relied largely on more tillers–more leaves relationships. The production strategy of stoloniferous grasses is more important for grazers because it ensures higher leaf than stem material. The higher leafiness of grasses not only serves as an indicator of productivity, but also of forage quality and palatability (O’Sullivan 2009). For bunch grasses, a good fit between tillers and leaves was obtained in uncleared treatments at Kgomo-kgomo. At low light levels, characteristic of the uncleared treatment, grasses tend to have a low leaf area that reduces self-shading and intra-tiller competition (Irving 2015).
Impact of bush clearing on grass biomass
Clearing increased grass biomass. Hare et al. (2021) reported similar results in Ethiopian rangelands, as did Smit (2005) in South Africa. Increased biomass is associated with a change in microclimate and increased resource availability, including soil nutrients and water, in cleared rangelands (Ding and Eldridge 2019). The higher biomass in the cleared treatment could be attributed to increased tiller and leaf production of grasses, largely bunch grasses. However, lack of interaction between treatment and species indicated that responses to clearing were not dependent on species type. Highly productive species (P. maximum) produced more biomass independent of the treatment (Fig. 5), despite a higher leaf and tiller production in the cleared than the uncleared treatment. This could perhaps be ascribed to higher plant densities of this species in the uncleared treatment, which is likely to have negated the positive effects of clearing on tiller and leaf production. At Makapaanstad, E. lehmanniana attained higher biomass on per area basis, whereas A. bipartita had a greater biomass per plant (Fig. 5). Tiller production of A. bipartita did not increase and leaf numbers increased, highlighting that biomass per plant was a function of a higher leaf production. The erect growth of bunch grasses promotes tillering from basal and axillary buds, which, subsequently, increases biomass production (Pereira et al. 2017). Even when space is limited for lateral tillering, stem elongation of bunch grasses allows a higher light interception, leading to a higher performance than for stoloniferous grasses (Scasta et al. 2015). This is mainly facilitated by partitioning of more photoassimilates to shoots in bunch grasses (Gomes et al. 2020).
Impact of bush clearing on tuft sizes of grasses
Grass tuft responses were explained largely by treatment and species interaction. At Kgomo-kgomo for example, the high tiller-producing grasses, P. maximum and U. mosambicensis, had larger tufts in cleared treatments, suggesting that an increase in basal tillering increased the tuft size, which may help rehabilitate bare areas (Pierce et al. 2019). Angassa (2002) reported similar findings. However, there were no differences in tuft sizes of T. berteronianus and A. barbicollis between treatments, despite higher tiller production in the cleared treatment, perhaps a consequence of both aerial and basal tillering, becaus the former does not contribute to an increase in tuft width. This implies that T. berteronianus and A. barbicollis are not reliable for increasing ground cover via vegetative growth of tufts, suggesting that land managers would need alternative restoration options, such as, for example, seeding with local adaptable species. Only the stoloniferous grasses increased in tuft sizes at Makapaanstad, despite their tillers being similar between cleared and uncleared treatments. Stolons radiate to long distances, with each stolon having multiple attachments on the ground from which shoots emerge (Da Silva et al. 2015). Thus, assigning all measurements taken from multiple ramet attachments to the main genet could explain why stoloniferous grasses had larger tufts. It is possible that some ramet attachments occurred in resource-rich patches, which is likely to have increased individual basal tiller sizes instead of tiller numbers.
Conclusions
Species’ response to clearing in terms of tiller production varied with the species type, with clearing altering the tiller production of bunch but not stoloniferous grasses. However, leaf production of stoloniferous grasses increased despite tiller production not increasing in the cleared treatment. The increase in per plant biomass, more so for bunch grasses, signifies that bush clearing is important for increasing forage production in bush-encroached rangelands, through enhancement of plant performance, including increased tiller and leaf production. Thus, management following clearing should be directed towards promoting tillering, especially that leaf production increases linearly with tiller production.
Data availability
The data used in this study will be made available on request and discussion with the corresponding author.
Conflicts of interest
Authors declare that they have no competing interest.
Declaration of funding
This research was funded by ARC (grant no. MNM 3014) and National Research Foundation (NRF) of South Africa (grant no. 97887).
Supplementary material
Supplementary material is available online.
Acknowledgements
Authors thank Mr T. Magandana, T. Matlala and B. Bonzana for tiller and leaf counting as well as plant census. We further thank Makapaanstad bush control project manager (Mr F. Mpuloane) for availing equipment for bush control. We also thank the colleagues who dedicated their time to read and provide valuable comments for the enhancement of this paper.
References
Angassa, A (2002). The effect of clearing bushes and shrubs on range condition in Borana, Ethiopia. Tropical Grasslands 36, 69–76.Bassett, IE, Paynter, Q, and Beggs, JR (2011). Effect of artificial shading on growth and competitiveness of Alternanthera philoxeroides (alligator weed). New Zealand Journal of Agricultural Research 54, 251–260.
| Effect of artificial shading on growth and competitiveness of Alternanthera philoxeroides (alligator weed).Crossref | GoogleScholarGoogle Scholar |
Da Silva, SC, Sbrissia, AF, and Pereira, LET (2015). Ecophysiology of C4 forage grasses: understanding plant growth for optimising their use and management. Agriculture 5, 598–625.
| Ecophysiology of C4 forage grasses: understanding plant growth for optimising their use and management.Crossref | GoogleScholarGoogle Scholar |
DIGES (2012) Proposed construction of 100MW solar power plant on portion 6 and 7 of farm Bezuiidenhoutskraal 96 JR within Moretele Local Municipality of Bojanala Platinum District Municipality, North West Province. Draft scoping report. Department of Environmental Affairs. www.diges.co.za. [Accessed 01 June 2021]
Ding, J, and Eldridge, DJ (2019). Contrasting global effects of woody plant removal on ecosystem structure, function and composition. Perspectives in Plant Ecology, Evolution and Systematics 39, 125460.
| Contrasting global effects of woody plant removal on ecosystem structure, function and composition.Crossref | GoogleScholarGoogle Scholar |
Fey M (2010) ‘Soils of South Africa.’ 1st edn. (Cambridge University Press: Cambridge, UK)
Gomes, FJ, Pedreira, BC, Santos, PM, Bosi, C, and Pedreira, CGS (2020). Shading effects on canopy and tillering characteristics of continuously stocked palisadegrass in a silvopastoral system in the Amazon biome. Grass and Forage Science 75, 279–290.
| Shading effects on canopy and tillering characteristics of continuously stocked palisadegrass in a silvopastoral system in the Amazon biome.Crossref | GoogleScholarGoogle Scholar |
Hare, ML, Xu, X, Wang, Y, and Gedda, AI (2020). The effects of bush control methods on encroaching woody plants in terms of dieoff and survival in Borana rangelands, southern Ethiopia. Pastoralism: Research, Policy and Practice 10, 1–4.
| The effects of bush control methods on encroaching woody plants in terms of dieoff and survival in Borana rangelands, southern Ethiopia.Crossref | GoogleScholarGoogle Scholar |
Hare, ML, Xu, XW, Wang, YD, Yuan, Y, and Gedda, AE (2021). Do woody tree thinning and season have effect on grass species’ composition and biomass in a semi-arid savanna? The case of a semi-arid savanna, southern Ethiopia. Frontiers in Environmental Science 9, 692239.
| Do woody tree thinning and season have effect on grass species’ composition and biomass in a semi-arid savanna? The case of a semi-arid savanna, southern Ethiopia.Crossref | GoogleScholarGoogle Scholar |
Holub, P, Klem, K, Linder, S, and Urban, O (2019). Distinct seasonal dynamics of responses to elevated CO2 in two understorey grass species differing in shade‐tolerance. Ecology and Evolution 9, 13663–13677.
| Distinct seasonal dynamics of responses to elevated CO2 in two understorey grass species differing in shade‐tolerance.Crossref | GoogleScholarGoogle Scholar | 31938473PubMed |
Irving, LJ (2015). Carbon assimilation, biomass partitioning and productivity in grasses. Agriculture 5, 1116–1134.
| Carbon assimilation, biomass partitioning and productivity in grasses.Crossref | GoogleScholarGoogle Scholar |
Lett, MS, and Knapp, AK (2003). Consequences of shrub expansion in mesic grassland: resource alterations and graminoid responses. Journal of Vegetation Science 14, 487–496.
| Consequences of shrub expansion in mesic grassland: resource alterations and graminoid responses.Crossref | GoogleScholarGoogle Scholar |
Mndela, M, Madakadze, IC, Nherera-Chokuda, F, and Dube, S (2019). Dynamics of the soil seed bank over the short-term after bush clearing in a semi-arid shrubland in Springbokvlakte thornveld of South Africa. South African Journal of Botany 125, 298–309.
| Dynamics of the soil seed bank over the short-term after bush clearing in a semi-arid shrubland in Springbokvlakte thornveld of South Africa.Crossref | GoogleScholarGoogle Scholar |
Mndela, M, Madakadze, IC, Nherera-Chokuda, F, and Dube, S (2020). Is the soil seed bank a reliable source for passive restoration of bush-cleared semi-arid rangelands of South Africa? Ecological Processes 9, 1.
| Is the soil seed bank a reliable source for passive restoration of bush-cleared semi-arid rangelands of South Africa?Crossref | GoogleScholarGoogle Scholar |
Moerane R (2013) The impact of training using a structured primary animal health care model on the skills of rural small-scale farmers. Master’s Thesis, University of Pretoria, Pretoria, South Africa.
Mucina L, Rutherford MC, Palmer AR, Milton SJ, Scott L, Lloyd JW, van der Merwe B, Hoare DB, Bezuidenhout H, Vlok JHJ, Euston-Brown DIW, Powrie LW, Dold AP (2006) Nama-Karoo Biome. In ‘The vegetation of South Africa, Lesotho and Swaziland’. (Eds L Mucina, MC Rutherford) pp. 324–347. (SANBI: Pretoria, South Africa)
O’Connor, RC, Taylor, JH, and Nippert, JB (2020). Browsing and fire decreases dominance of a resprouting shrub in woody encroached grassland. Ecology 101, e02935.
| Browsing and fire decreases dominance of a resprouting shrub in woody encroached grassland.Crossref | GoogleScholarGoogle Scholar | 31746458PubMed |
O’Sullivan D (2009) Pasture management for the inland Burnett. p. 5. Project Report. State of Queensland.
Pearce, J, and Derrick, B (2019). Preliminary testing: the devil of statistics? Reinvention: An International Journal of Undergraduate Research 12, e02935.
| Preliminary testing: the devil of statistics?Crossref | GoogleScholarGoogle Scholar |
Pereira, LET, Paiva, AJ, Geremia, EV, and Da Silva, SC (2017). Contribution of basal and aerial tillers to sward growth in intermittently stocked elephant grass. Grassland Science 64, 108–117.
| Contribution of basal and aerial tillers to sward growth in intermittently stocked elephant grass.Crossref | GoogleScholarGoogle Scholar |
Pierce, NA, Archer, SR, Bestelmeyer, BT, and James, DK (2019). Grass–shrub competition in arid lands: an overlooked driver in grassland–shrubland state transition? Ecosystems 22, 619–628.
| Grass–shrub competition in arid lands: an overlooked driver in grassland–shrubland state transition?Crossref | GoogleScholarGoogle Scholar |
Pierson, EA, Mack, RN, and Black, RA (1990). The effect of shading on photosynthesis, growth, and regrowth following defoliation for Bromus tectorum. Oecologia 84, 534–543.
| The effect of shading on photosynthesis, growth, and regrowth following defoliation for Bromus tectorum.Crossref | GoogleScholarGoogle Scholar | 28312971PubMed |
Roques, KG, O’Connor, TG, and Watkinson, AR (2001). Dynamics of shrub encroachment in an African savanna: relative influences of fire, herbivory, rainfall and density dependence. Journal of Applied Ecology 38, 268–280.
| Dynamics of shrub encroachment in an African savanna: relative influences of fire, herbivory, rainfall and density dependence.Crossref | GoogleScholarGoogle Scholar |
Saixiyala, DY, Yang, D, Zhang, S, Liu, G, Yang, X, Huang, Z, and Ye, X (2017). Facilitation by a spiny shrub on a rhizomatous clonal herbaceous in thicketization-grassland in northern China: increased soil resources or shelter from herbivores. Frontiers in Plant Science 8, 809.
| Facilitation by a spiny shrub on a rhizomatous clonal herbaceous in thicketization-grassland in northern China: increased soil resources or shelter from herbivores.Crossref | GoogleScholarGoogle Scholar |
SAS Institute (2015) JMP: version12: scripting guide. SAS Institute Inc., Cary, NC, USA.
Scasta, JD, Engle, DM, Fuhlendorf, SD, Redfearn, DD, and Bidwell, TG (2015). Meta-analysis of exotic forages as invasive plants in complex multi-functioning landscapes. Invasive Plant Science and Management 8, 292–306.
| Meta-analysis of exotic forages as invasive plants in complex multi-functioning landscapes.Crossref | GoogleScholarGoogle Scholar |
Simmons, MT, Archer, SR, Teague, WR, and Ansley, RJ (2008). Tree (Prosopis glandulosa) effects on grass growth: an experimental assessment of above- and belowground interactions in a temperate savannah. Journal of Arid Environments 72, 314–325.
| Tree (Prosopis glandulosa) effects on grass growth: an experimental assessment of above- and belowground interactions in a temperate savannah.Crossref | GoogleScholarGoogle Scholar |
Smit, GN (2005). Tree thinning as an option to increase herbaceous yield of an encroached semi-arid savanna in South Africa. BMC Ecology 5, 4.
| Tree thinning as an option to increase herbaceous yield of an encroached semi-arid savanna in South Africa.Crossref | GoogleScholarGoogle Scholar | 15921528PubMed |
Solofondranohatra, CL, Vorontsova, MS, Hackel, J, Besnard, G, Cable, S, Williams, J, Jeannoda, V, and Lehman, CER (2018). Grass functional traits differentiate forest and savanna in the Madagascar central highlands. Frontiers in Ecology and Evolution 6, 184.
| Grass functional traits differentiate forest and savanna in the Madagascar central highlands.Crossref | GoogleScholarGoogle Scholar |
Stephens, GJ, Johnston, DB, Jonas, JL, and Paschke, MW (2016). Understory responses of mechanical treatment of Pinyon–Juniper in north-western Colorado. Rangeland Ecology & Management 69, 351–359.
| Understory responses of mechanical treatment of Pinyon–Juniper in north-western Colorado.Crossref | GoogleScholarGoogle Scholar |
Tomlinson, KW, and O’Connor, TG (2004). Control of tiller recruitment in bunchgrasses: uniting physiology and ecology. Functional Ecology 18, 489–496.
| Control of tiller recruitment in bunchgrasses: uniting physiology and ecology.Crossref | GoogleScholarGoogle Scholar |
Van Oudtshoorn F (1999) ‘Guide to grasses of southern Africa’, (Briza Publications: Pretoria, South Africa)
Van Wilgen BW, Measey J, Richardson DM, Wilson JR, Zengeya TA (Eds) (2020) Biological invasions in South Africa. In ‘Invading nature – Springer Series in Invasion Ecology’. pp. 48–51. (Springer Nature: Cham, Switzerland)
| Crossref |
Volder, A, Briske, DD, and Tjoelker, MG (2013). Climate warming and precipitation redistribution modify tree–grass interactions and tree species establishment in a warm-temperate savanna. Global Change Biology 19, 843–857.
| Climate warming and precipitation redistribution modify tree–grass interactions and tree species establishment in a warm-temperate savanna.Crossref | GoogleScholarGoogle Scholar | 23504841PubMed |
Warren K, Hugo W, Wilson H (2018) Preliminary report and data on bush encroachment and land cover change, released to DEA, DEA consultants, and selected collaborators. Policy brief. Department of Environmental Affairs. Available at www.dffe.gov.za. [Accessed 20 January 2021]
Woon, JS, Atkinson, D, Adu-Bredu, S, Eggleton, P, and Parr, CL (2021). Termites have developed wider thermal limits to cope with environmental conditions in savannas. bioRxiv. , .
| Termites have developed wider thermal limits to cope with environmental conditions in savannas.Crossref | GoogleScholarGoogle Scholar |
Ye, D, Hu, Y, Song, M, Pan, X, Xie, X, Liu, G, Ye, X, and Dong, M (2014). Clonality-climate relationships along latitudinal gradient across China: adaptation of clonality to environments. PLoS One 9, e94009.
| Clonality-climate relationships along latitudinal gradient across China: adaptation of clonality to environments.Crossref | GoogleScholarGoogle Scholar | 24709992PubMed |
Zakaria, M, Rajpar, MN, and Sajap, SA (2009). Species diversity and feeding guilds of birds in Paya Indah Wetland Reserve, Peninsular Malaysia. International Journal of Zoological Research 5, 86–100.
| Species diversity and feeding guilds of birds in Paya Indah Wetland Reserve, Peninsular Malaysia.Crossref | GoogleScholarGoogle Scholar |