

Effects of grazing season and stocking rate on seed bank in sheep dung on the semiarid Loess Plateau
Shulin Wang A , An Hu A , Jing Zhang A and Fujiang Hou A BA State Key Laboratory of Grassland Agro-Ecosystems, Key Laboratory of Grassland Livestock Industry Innovation, Ministry of Agriculture and College of Pastoral Agriculture Science and Technology, Lanzhou University, Lanzhou, 730000, Gansu, China.
B Corresponding author. Email: cyhoufj@lzu.edu.cn
The Rangeland Journal 41(5) 405-413 https://doi.org/10.1071/RJ19036
Submitted: 8 June 2019 Accepted: 16 October 2019 Published: 12 November 2019
Journal Compilation © Australian Rangeland Society 2019 Open Access CC BY-NC-ND
Abstract
Endozoochorous dispersal of seeds by livestock has long attracted the attention of grassland scientists, but little is known about seed dispersal via Tan sheep dung on the dry grasslands of the Loess Plateau. We investigated the composition of dung seed bank of Tan sheep under summer and winter rotational grazing regimes at different stocking rates (2.7, 5.3 and 8.7 sheep ha–1), and assessed the relationships between seed mass and shape, egested seedling density and species richness and diversity. We also assessed the effects of stocking rate and grazing season on seedling density and species richness and diversity of the dung seed bank. Seeds of 10 pasture species germinated from Tan sheep dung with only two being annual species (Eragrostis pilosa and Chenopodium glaucum). The mean seed mass (±s.e.) was 2.59 ± 0.23 mg, ranging from 0.10 mg (Cleistogenes songorica) to 10.59 mg (Thermopsis lanceolata), and the mean seed shape index (±s.e.) was 0.10 ± 0.02, ranging from 0.03 (Lespedeza bicolor) to 0.19 (Stipa bungeana). Species richness and diversity and seedling density decreased with increasing seed mass and shape index, suggesting that small, round seeds are most suitable for endozoochorous dispersal. Stocking rate had no effect on the number of germinated seeds that collected from summer or winter grazing pastures. Species richness and diversity and seedling density were greater in winter grazing pastures than in summer grazing pastures. Jaccard coefficients of similarity between the Tan sheep dung bank and aboveground vegetation were <0.5 for all stocking rates, indicating their weak relationship, but were significantly higher for winter than summer grazing. Plant seeds on the Loess Plateau could disperse through Tan sheep grazing, endozoochory can increase the heterogeneity of rotationally grazed plant communities, and this dispersal mode is an adaptation of plants to the harsh environment of the semiarid areas.
Additional keywords: endozoochory, grazing intensity, rangeland management, similarity, Tan sheep.
Introduction
Viable seeds in herbivore faeces constitute the dung seed bank – an important source of vegetation renewal and a supplement to the soil seed bank. The composition of the dung seed bank depends in part on pasture composition and the selective feeding of livestock. The passage of plant seeds through the digestive tract of animals affects the success of seed germination (Wang et al. 2017). Seed dispersal following plant consumption by animals (endozoochory) is an important natural means of seed propagation. Myers et al. (2004) reported that more than 70 plant species including native and alien herbs, shrubs and trees are able to germinate from deer faeces collected over one year in North America, and more than 30 seeds from a faecal pellet germinate in all months of the year. After decomposition of dung, seeds in faeces eventually move or are transported into the soil, contributing to the soil seed bank. Livestock manure is therefore significant for pasture seed transmission, soil seed bank composition and seed germination, and has the potential to change the composition of grassland vegetation (Bakker and Olff 2003). Faecal sedimentation and the germination and establishment of seeds from faeces can increase the similarity of plant communities between different types of grazed grassland and promote differences between grassland plants of the same community type (Malo and Suárez 1995). However, the relationship between faecal seed bank and aboveground vegetation, and the mechanisms for maintaining this relationship, remain unclear.
The colour and release height of fruits (or achenes) as well as animal habitat and feeding preferences influence the probability of seeds being swallowed (Picard et al. 2015). Moreover, characteristics such as mass and shape affect seed viability and germination capacity after passage through the digestive tract. Small seeds are more likely than large seeds to survive passage through the digestive tract (Soons et al. 2008; Brochet et al. 2010), since they are less likely to be damaged during chewing (Manzano et al. 2005). Seed shape also influences the passage time through the digestive tract (Picard et al. 2015). Spherical seeds pass the digestive tract more quickly than flatter or elongated seeds, and have a higher germination rate after recovery (Whitacre and Call 2006). The potential for long-distance endozoochory depends on the number of viable seeds consumed and the effects of gut retention on germination.
Stocking rate and grazing season are two important factors affecting the size and composition of the dung seed bank. Stocking rate affects aboveground vegetation composition and the density of reproductive culms (Hu et al. 2019a), which in turn, determines the amount of livestock seed intake. Grazing season affects the effectiveness of endozoochorous seed dispersal, as only seeds mature when consumed are potentially viable when dispersed. Most studies of the dung seed bank have reported that seeds can be dispersed following egestion by livestock (Cosyns et al. 2005; Mouissie et al. 2005; Mancilla‐Leytón et al. 2011), but these studies do not evaluate the effects of grazing practices such as stocking rate and grazing season on the dispersal of viable seeds.
The Loess Plateau in north-western China, one of the most naturally and culturally distinctive regions on earth, is situated in the middle reaches of the Yellow River. It occupies 650 000 km2 and is characterised by windblown loess deposits up to several hundred meters thick (Sun et al. 2001). Approximately 54% of Loess Plateau grassland area is used for grazing livestock, providing the main source of income for the local population of ~16 million people. However, overgrazing has resulted in soil erosion and pasture degradation. Rotational grazing on the Loess Plateau has been encouraged by the local government since the 1980s as the most effective strategy to prevent further degradation (Wang et al. 2018a).
Tan sheep are the traditional and dominant grazing livestock on the Loess Plateau in both summer and winter, but little is known about their seed dispersal by egestion on the dry grasslands. Our preliminary field observations show that a subset of plant seedlings become established from Tan sheep dung (J. Zhang and A. Hu, 35 seedlings in each pile of sheep dung, unpubl. obs. 2015), indicating seeds can pass though the sheep’s gut and be dispersed. The objectives of this study were to determine: (i) the size and species composition of the dung seed bank from Tan sheep and the relationship between the dung seed bank and aboveground vegetation; (ii) the relationships between seed characteristics (i.e. mass and shape) and egested seedling density on the one hand and species richness and diversity on the other; and (iii) how stocking rate and grazing season affect seedling density and the species richness and diversity of the dung seed bank. This research can contribute significantly to a better understanding of the factors involved in grazing management decisions making required for protection and restoration of grasslands.
Materials and methods
Study site
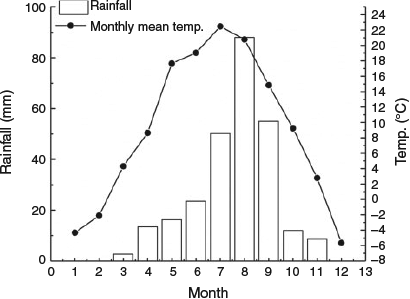
This study site is located in Huan County, eastern Gansu Province, north-western China (37.14°N, 106.84°E) on the Loess Plateau, at an altitude of 1650 m above sea level. The mean daily air temperature is 7.1°C, with a mean annual precipitation of 360 mm and a mean annual potential evaporation of 1993 mm. More than 70% of the rainfall occurs between July and September (Fig. 1). Spring and autumn are typically short, summer hot and humid, and winter long and cold. The soil is classified as sandy, free-draining loess, and the rangeland as a typical temperate steppe. The dominant species at the study site are perennial low shrubs, i.e. Artemisia capillaries Thunb. and Lespedeza davurica (Laxm.) Schindl and perennial grass, Stipa bungeana Trin. (Table 2). The biomass of the three dominant species accounts for 70–90% of the total biomass (Hou et al. 2002; Hu et al. 2019a).
|
Experimental design
The study started on a pasture rotationally grazed by 4-month-old Tan sheep wethers, the traditional sheep breed on the Loess Plateau since 2001. Two flatland areas, ~200 m apart, were selected for summer grazing and early-winter grazing. Vegetation composition and coverage between these two regions were similar. Each area was divided into nine 0.5-ha plots (18 grazing paddocks in total) to give three replicates at each of three stocking rates: 2.7, 5.3 and 8.7 sheep ha–1. Stocking rates were based on data for long-term grass yield, livestock weight and feed intake. Summer grazing started from early June to early September (90 days duration) with three 10-day rotations per replicate and winter grazing from mid-November to late December (45 days duration) with two 8-day rotations per replicate (Chen et al. 2010, 2015; Hu et al. 2019b). To avoid any impact of sheep age on seed ingestion, a new cohort of 4-month-old sheep was used at the beginning of each grazing season. For summer grazing, there was no grazing after early September, and for winter grazing, none after late December.
Because of the local climate on the Loess Plateau, the grazing season is only divided into cold and worm season (Chen et al. 2015). Since the grazing season treatment (summer vs early winter) is not spatially replicated, so ‘grazing season’ could be considered to be pseudoreplicated (Hurlbert 1984).
Dung collection
Samples of freshly deposited sheep dung were collected on two occasions (mid-September and late December 2016) from each plot, and faecal pellets pooled to provide ~2 kg fresh weight per plot. Dung samples bagged in plastic bags were dried for 72 h at 35°C to prevent premature germination. Dried dung pellets were stored in the dark at room temperature.
Germination
Each sample of dry manure was gently compressed without damaging seeds. A 100-g portion was taken from each sample, mixed with ~50 g sterilised sand and spread in a pot at a 2-cm thickness on a 5 cm thick bed of vermiculite. Pots were placed in a greenhouse (70–80% humidity, 15–20°C, 16 h light per day) and watered twice a day from January until June 2017. The temperature regime was chosen to mimic the early-spring germination temperatures of most species typical of the Loess Plateau. Emerging seedlings were recorded and removed as soon as identified or were transplanted into separate pots for later identification. Whenever seedlings were removed, the dung/sand mix was stirred to facilitate germination of remaining buried seeds. Germinated seedlings are referred to here as endozoochorous seedlings (Malo and Suárez 1995).
Aboveground vegetation
To compare dung seed bank species with the aboveground vegetation, herbaceous species were identified during the peak growth season (mid-August 2016). During this period, aboveground vegetation has the highest species richness (~25 species) and thus provides the greatest representation of grassland productivity. Species richness (number of plant species) and species density were recorded in each of six quadrats (1 m × 1 m) that were laid out in the middle of each of the 18 grazing paddocks.
Seed mass and shape index
Between mid-June and mid-November 2016 (to match the grazing period), mature seeds (representative of the range of seed size and shape) were manually collected from at least 20 individual plants from each of the 27 common grassland species identified in the aboveground vegetation (12 families, Table 2). All are native and represent both current and historical species of the Loess Plateau. In addition, owing to limited precipitation and low species richness in this semiarid area, harsh environmental conditions have weakened the ability of Tan sheep to graze on selected/preferred plants (Chen et al. 2010), implying that all 27 plant species were considered to be consumed equivalently. Seeds were stored at room temperature under dry and dark conditions (Peco et al. 2006).
Seeds without appendages were weighed (100 seeds per species, accuracy ± 0.01 mg). Ten seeds per species were randomly selected for measurement of length, width and height using an electronic Vernier caliper under a stereomicroscope (Nikon SMZ 1500, Shanghai, China, 25 µm precision). The seed shape index (I), i.e. divergence from sphericity, was calculated according to Wang et al. (2017) as the variance of the three main dimensions:
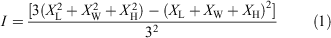
where XL, XW, XH are seed length, width and height, respectively, divided by seed length. I value ranges from 0 and 1 and I = 0 for spherical seeds and I >0 for flatter or elongated seeds.
Diversity and similarity indices
The number of emerged seedlings (seedlings g–1 dung) and species richness (species g–1 dung) were determined basing on data collected from five replicates from each dung sample. For each plot, a Shannon–Wiener diversity index (H′) was calculated as:
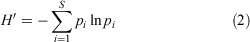
where pi is the relative proportion of species of the total community (in this study, community refers to the dung seed bank) and s is the total number of species for each dung sample.
The Jaccard coefficient of similarity (SJ) was used to test the similarities in species composition between the dung seed bank and the aboveground vegetation for each stocking rate under summer and winter rotational grazing regimes:

where a is the number of common species presenting in both dung seed bank and aboveground vegetation at the same stocking rate in the same grazing season and b is the total number of species.
Data analysis
Data were analysed using Statistical Package for the Social Sciences (SPSS) for Windows (ver. 25.0, SPSS, Inc., Chicago, IL, USA). Data were checked for normal distribution using the Shapiro-Wilk test. Data of dung seed bank species richness, species diversity, or seedling density were log(10)-transformed to pursue the assumption of normality and homogeneity of variances. Dung seed bank species richness, species diversity, or seedling density in relation to single-seed mass or seed shape index were determined by multiple regression analyses, except the seed species richness in relation to mass, where an exponential decline model was used (Fig. 2). Data on Jaccard coefficient of similarity (Fig. 3) and dung seed bank species richness, species diversity, seedling density (Fig. 4) in different grazing seasons affected by stocking rate were analysed using analysis of variance with least significant difference for multiple comparisons. The level of significance used was P < 0.05.

|
|
|
Structural equation modelling (SEM) was utilised to estimate the contribution of stocking rate and grazing season to the aboveground vegetation composition and dung seed bank, i.e. seedling density, species richness and species diversity. The primary advantage of SEM is its ability to evaluate hypothesised and complex causal relationships between variables by translating these into a pattern of expected statistical relationships in the data (Grace 2006). Also, the core theory of structural equation model analysis is the covariance of variables and the pseudorepetition (some treatment lack of replication, e.g. grazing season) could also use this analytical method (Deng and Qiu 2015). SEM analyses were performed using AMOS 22 (Arbuckle 2010). When using the AMOS 22 software for analysis, grazing season is named as pseudoreplicated, and summer grazing season and winter grazing season defined as a character string, the software automatically giving the statistical results. The chi-square test was used to evaluate the fit of the model. The model has a good fit when 0 ≤ χ2/df ≤ 2 and 0.05 < P ≤ 1. A large P-value (>0.05) indicates that the covariance structure of the data does not differ significantly from the expected model (Grace 2006).
Results
Germinated species from dung
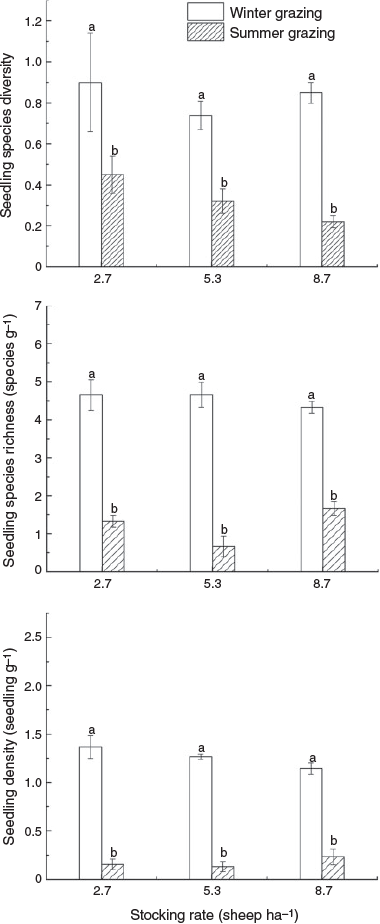
Of 27 plant species identified from field vegetation, only 10 species (two annual and eight perennial species) from Tan sheep dung germinated successfully, with an average seedling density of 0.72 g–1 dung. Four species, Potentilla bifurca, E. pilosa, Dodartia orientalis and L. bicolor, germinated from both summer and winter grazing dung samples. The remainder (A. capillaris, Gueldenstaedtia verna, C. glaucum, Potentilla multifida, C. songorica and L. secalinus) germinated only from winter grazing dung samples. In both summer and winter grazing systems, seedling density (g–1 dung) decreased when stocking rate increased, with the exception of L. bicolor, a native dominant species (Table 1).

|
Seed mass and shape index
Twenty-seven plant species were identified in the pasture vegetation. Approximately 70% were perennials, including two shrubs (L. bicolor and A. capillaris) (Table 2). The mean seed mass (±s.e.) of identified species was 2.59 ± 0.23 mg, ranging from 0.10 mg (C. songorica) to 10.59 mg (Thermopsis lanceolata), with 18 species (67% of total) having a seed mass of >1 mg. The mean seed shape index (±s.e.) was 0.10 ± 0.02, ranging from 0.03 (L. bicolor) to 0.19 (S. bungeana), with 13 species (48% of the total) having a seed shape index of >0.10

|
Dung seed species richness, species diversity and seedling density in relation to seed mass and shape index
As seed mass increased, dung seed bank species richness, species density and seedling diversity significantly decreased, indicating that large ingested seeds had a lower germination capacity than small ingested seeds (Fig. 2a). Similarly, as seed shape index increased, dung seed bank species richness, species density and seedling diversity significantly decreased (Fig. 2b), indicating that flatter or elongated seeds ingested by Tan sheep had a lower germination capacity than spherical seeds.
Similarity of dung seed bank and aboveground vegetation
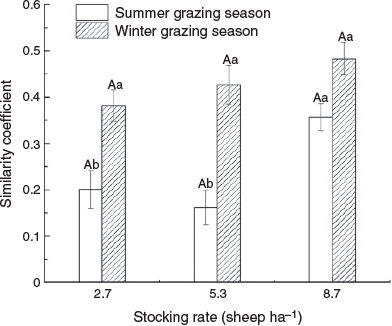
The Jaccard coefficients of similarity were <0.5 for all three stocking rates (Fig. 3). The similarity in winter grazing pastures was significantly higher than in summer grazing pastures (P < 0.05), with no significant difference detected between stocking rates in winter grazing pastures (P > 0.05) (Fig. 3). The similarity at 8.7 sheep ha–1 was significantly higher than that at 2.7 and 5.3 sheep ha–1 in summer grazing pastures (P < 0.05) (Fig. 3).
Seed species richness, species diversity and seedling density in relation to grazing season and stocking rate
Dung seed bank species richness, species diversity and seedling density were significantly higher in winter grazing pastures than summer grazing ones at all three stocking rates (P < 0.01) (Fig. 4; Table 3). However, stocking rate and the interaction of grazing season and stocking rate had no significant impact on species richness, species diversity and seedling density (P > 0.05) (Fig. 4; Table 3).

|
Regulation of dung seed bank by stocking rate and grazing season
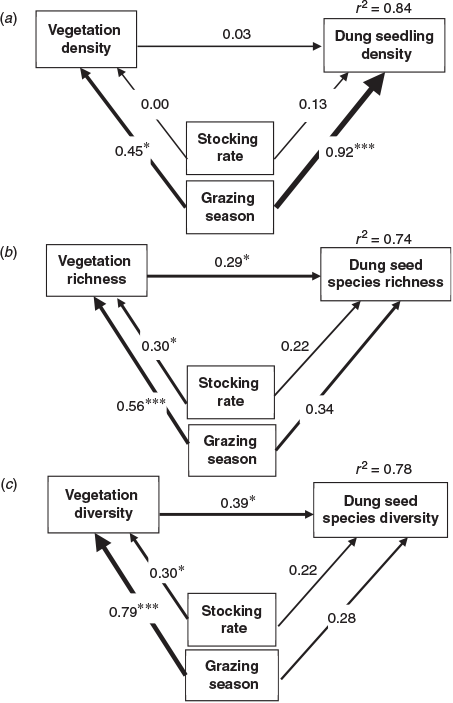
Stocking rate had no significant indirect (through vegetation density) or direct effect on seedling density (standardised path coefficients = 0.03 and 0.13, respectively, both P > 0.05; Fig. 5a). Both direct and indirect effects of grazing season on seedling density were significantly positive (standardised path coefficients = 0.92 and 0.45; P < 0.001 and P < 0.05, respectively, see Fig. 5a).
|
Stocking rate and grazing season had no significant direct effect on dung seed bank species richness (0.22 and 0.34, respectively, both P > 0.05), but both had significant indirect effects on dung seed bank species richness through species richness of aboveground vegetation (0.29; P < 0.05) (Fig. 5b).
Stocking rate and grazing season also had no significant direct effect on species diversity of the dung seed bank (0.22 and 0.28, respectively, both P > 0.05), but both had indirect significant effects on species diversity of the dung seed bank through diversity of the aboveground vegetation (0.39; P < 0.05) (Fig. 5c).
Discussion
Germinated species from dung
Of the 10 plant species that germinated from Tan sheep dung, only two were annuals (E. pilosa and C. glaucum; Table 1). Annuals rely only on seeds for reproduction, and thus tend to produce a larger number of seeds (Smith and Fretwell 1974) with ‘r-selected’ life-history characteristics (plants produce large numbers of offspring to increase the number of surviving offspring) (Mac-Arthur and Wilson 1967). However, plants with higher number of seeds also tend to have higher seed abortion rates and poor seed tolerance (Lönnberg and Eriksson 2013), which contributes to the low seed survival found for annual plants in animal faeces. In fact, the composition of the dung seed bank is determined by the species of seed ingested by livestock, and selective feeding of livestock has an important impact on this process. To date, few studies on selective feeding and the gut environment of sheep on the Loess Plateau have been conducted (Chen et al. 2010).
The size of the dung seed bank was affected by livestock species, amount of seed intake and the physical and chemical properties of the faeces (Milotić and Hoffmann 2016), as well as seed traits (Pakeman et al. 2002). We explored the efficiency of endozoochory for temperate (e.g. Loess Plateau) grassland species, where the processes of mastication and gut passage appear to impose a high seed loss, as high as >80% (Wang et al. 2017). The mean seed density was 0.72 g–1 dung, slightly lower than that (0.8 g–1) reported by Kuiters and Huiskes (2010). High seed consumption rate suggests the size of the dung seed bank was small (Yu et al. 2013). However, the cost of endozoochory may be offset by the possibility of dispersed seeds travelling long distances (e.g. C. songorica and L. bicolor in our research) without investment in other specialised structures (e.g. wings or pappus; Peco et al. 2006).
Effect of seed traits on the dung seed bank
All seeds germinated from Tan sheep dung were classified as ‘small seed’ species according to the criteria of Thompson et al. (1993). Mean seed mass ranged from 0.10 mg (C. songorica) to 4.5 mg (L. secalinus) (Table 2). In addition, seeds the 10 germinated species were approximately spherical and small, with seed shape indices ranging from 0.03 (L. bicolor) to 0.09 (C. songorica) (present study; Wang et al. 2018b). Previous studies have indicated that small size is an important characteristic that aids seed survival following ingestion, allowing seeds to avoid molar grinding (Kuiters and Huiskes 2010) and survive after passing the digestive tract (Pakeman et al. 2002). Small seed size and round shape lead to shorter retention time in the digestive tract, reducing the seeds exposure to microbial attack in the rumen. It is not surprising that as egested seed mass or seed shape index increased, species richness, species diversity and seedling density decreased (Fig. 2), indicating that larger (single-seed mass >10 mg) and elongated (shape index >0.1) seeds had lower germination after egestion. Some researchers have suggested viable small seeds are more likely to be present in egested herbivore dung than large seeds, because of the negative relationship between seed size and seed production (i.e. small seeds are produced in greater numbers) rather than the better adaptation of small seeds to survive gut passage (Eriksson and Jakobsson 1998; Pakeman et al. 2002; Couvreur et al. 2005).
The similarity between dung seed bank and aboveground vegetation
The relationship between the dung seed bank and aboveground vegetation can be influenced by selective feeding of livestock (Bagchi and Ritchie 2010), physical and chemical properties of faeces (Milotić and Hoffmann 2016) and microhabitat properties of sites at which seeds are egested with dung (Calviño-Cancela and Martín-Herrero 2009). In the present study, the similarity of species between the soil seed bank and aboveground vegetation was low (<0.5; Fig. 3), because 10 species detected from aboveground vegetation were absent from the dung seed bank. Nonetheless, ~37% of pasture plant species could germinate after passage. In semiarid environments such as the Loess Plateau, seed dispersal within the dung pellet provides significant protection for seed survival until sufficient rainfall for germination and establishment. This mode of germination represents an adaptation of plant seeds to the harsh environment of the Loess Plateau and other semiarid grasslands.
Effect of stocking rate and grazing season on dung seed bank
Stocking rate had no significant effect on species density and richness in sheep dung as well as the subsequent seedling diversity, due to the decrease in forage intake as stocking rate increased (Langlands and Bennett 1973). In addition, more seeds are buried through increased trampling at high stocking rates (Willms and Quinton 1995), leaving fewer seeds on the parent plants to be consumed by sheep.
The dung seed bank plays an important role in vegetation regeneration, with seeds in egested sheep dung contributing significantly to it. However, the value of this contribution depends on the rate and degree of decomposition of the dung, which often is hard and dry. After defecation, the main factor influencing on the condition of dung is trampling by livestock (Liu et al. 2011). Chen (2015) reported that higher stocking rate boshes dung decomposition. Precipitation also affects dung decomposition. For example, in the Chihuahuan Desert, dung and other organic matter decompose rapidly during the rainy season (Whitford et al. 1982). Seedling emergence in arid environments is primarily related to patterns of soil water availability (Winkel and Roundy 1991). On the Loess Plateau, more than 60% of rainfall is concentrated in July to September (Fig. 1) (Hou et al. 2002). This period coincides with summer rotational grazing, when many seeds are transferred from dung to the soil seed bank, and wet conditions are more amenable to seed germination and plant establishment.
As only matured seeds consumed by livestock have the potential to be viable after passing the gut, grazing season has an important impact on the dung seed bank. Results of SEM showed that both direct and indirect effects of grazing season on seedling density were significantly positive (standardised path coefficients = 0.92 and 0.45; P < 0.001 and P < 0.05, respectively, Fig. 5a). On the Loess Plateau, most plant species mature before winter, and seeds retained through the winter are ingested by foraging sheep and egested in their faeces (mean 0.72 seed g–1 dung). Malo and Suárez (2000) found that the seed content of rabbit dung samples peaked at the end of spring, coinciding with the maximum density of certain mature species (e.g. Caryophyllaceae).
In conclusion, the findings of our study suggest that the foraging activities of Tan sheep have the potential to contribute to the gathering of plant seeds under traditional rotational grazing on the Loess Plateau. This is especially true in a winter grazing system, when most plants still retain seeds. Although the cost of passage through the gut of Tan sheep is undoubtedly high for dry-fruited plant species (Traveset et al. 2002), endozoochory may enhance species colonisation through the potential benefits of directional dispersal. This mode of dispersal may increase the heterogeneity of plant communities via the rotational grazing of sheep on the Loess Plateau.
Conflicts of interest
The authors declare no conflicts of interest.
Acknowledgements
This study was financially supported by the Strategic Priority Research Program of the Chinese Academy of Sciences (grant no. XDA20100103), the National Natural Science Foundation of China (no. 31672472) and the Program for Changjiang Scholars and Innovative Research Team in University (IRT_17R50). We thank Dr Coby Jean Hoogendoorn, Dr Xiongzhao He and Roger Davies for their suggestions and improvements to the language in this manuscript.
References
Arbuckle, J. (2010). ‘IBM SPSS Amos 19 User’s Guide.’ (Amos Development Corporation: Crawfordville, FL, USA.)Bagchi, S., and Ritchie, M. E. (2010). Introduced grazers can restrict potential soil carbon sequestration through impacts on plant community composition. Ecology Letters 13, 959–968.
| 20482575PubMed |
Bakker, E. S., and Olff, H. (2003). Impact of different-sized herbivores on recruitment opportunities for subordinate herbs in grasslands. Journal of Vegetation Science 14, 465–474.
| Impact of different-sized herbivores on recruitment opportunities for subordinate herbs in grasslands.Crossref | GoogleScholarGoogle Scholar |
Brochet, A. L., Guillemain, M., Gauthier-Clerc, M., Fritz, H., and Green, A. J. (2010). Endozoochory of Mediterranean aquatic plant seeds by teal after a period of desiccation: determinants of seed survival and influence of retention time on germinability and viability. Aquatic Botany 93, 99–106.
| Endozoochory of Mediterranean aquatic plant seeds by teal after a period of desiccation: determinants of seed survival and influence of retention time on germinability and viability.Crossref | GoogleScholarGoogle Scholar |
Calviño-Cancela, M., and Martín-Herrero, J. (2009). Effectiveness of a varied assemblage of seed dispersers of a fleshy-fruited plant. Ecology 90, 3503–3515.
| Effectiveness of a varied assemblage of seed dispersers of a fleshy-fruited plant.Crossref | GoogleScholarGoogle Scholar | 20120817PubMed |
Chen, H. (2015). Decomposition of dung and herbage litter in typical steppe – Tan sheep rotational grazing system in the eastern Gansu Loess Plateau. Masters thesis, Lanzhou University, China. [in Chinese with English abstract.]
Chen, X. J., Hou, F. J., Matthew, C., and He, X. Z. (2010). Stocking rate effects on metabolizable energy intake and grazing behaviour of Tan sheep in steppe grassland on the Loess Plateau of Northwest China. The Journal of Agricultural Science 148, 709–721.
| Stocking rate effects on metabolizable energy intake and grazing behaviour of Tan sheep in steppe grassland on the Loess Plateau of Northwest China.Crossref | GoogleScholarGoogle Scholar |
Chen, J., Hou, F., Chen, X., Wan, X., and Millner, J. (2015). Stocking rate and grazing season modify soil respiration on the Loess Plateau, China. Rangeland Ecology and Management 68, 48–53.
| Stocking rate and grazing season modify soil respiration on the Loess Plateau, China.Crossref | GoogleScholarGoogle Scholar |
Cosyns, E., Claerbout, S., Lamoot, I., and Hoffmann, M. (2005). Endozoochorous seed dispersal by cattle and horse in a spatially heterogeneous landscape. Plant Ecology 178, 149–162.
| Endozoochorous seed dispersal by cattle and horse in a spatially heterogeneous landscape.Crossref | GoogleScholarGoogle Scholar |
Couvreur, M., Cosyns, E., Hermy, M., and Hoffmann, M. (2005). Complementarity of epi- and endozoochory of plant seeds by free ranging donkeys. Ecography 28, 37–48.
| Complementarity of epi- and endozoochory of plant seeds by free ranging donkeys.Crossref | GoogleScholarGoogle Scholar |
Deng, S. Y., and Qiu, Q. H. (2015). Current situation and prospects for study and application on structural equation model. Jiangsu Science and Technology Information 24, 76–78.
Eriksson, O., and Jakobsson, A. (1998). Abundance, distribution and life histories of grassland plants: a comparative study of 81 species. Journal of Ecology 86, 922–933.
| Abundance, distribution and life histories of grassland plants: a comparative study of 81 species.Crossref | GoogleScholarGoogle Scholar |
Grace, J. B. (2006). ‘Structural Equation Modeling and Natural Systems.’ (Cambridge University Press: Cambridge, UK.)
Hou, F. J., Xiao, J. Y., and Nan, Z. B. (2002). Eco-restoration of abandoned farmland in the Loess Plateau. Chinese Journal of Applied Ecology 13, 923–929.
Hu, A., Zhang, J., Chen, X., Chang, S., and Hou, F. (2019a). Winter grazing and rainfall synergistically affect soil seed bank in semiarid area. Rangeland Ecology and Management 72, 160–167.
| Winter grazing and rainfall synergistically affect soil seed bank in semiarid area.Crossref | GoogleScholarGoogle Scholar |
Hu, A., Zhang, J., Chen, X. J., Millner, J. P., Chang, S. H., Bowatte, S., and Hou, F. J. (2019b). The composition, richness, and evenness of seedlings from the soil seed bank of a semi-arid steppe in northern China are affected by long-term stocking rates of sheep and rainfall variation. The Rangeland Journal 41, 23–32.
| The composition, richness, and evenness of seedlings from the soil seed bank of a semi-arid steppe in northern China are affected by long-term stocking rates of sheep and rainfall variation.Crossref | GoogleScholarGoogle Scholar |
Hurlbert, S. H. (1984). Pseudoreplication and the design of ecological field experiments. Ecological Monographs 54, 187–211.
| Pseudoreplication and the design of ecological field experiments.Crossref | GoogleScholarGoogle Scholar |
Kuiters, A. T., and Huiskes, H. P. J. (2010). Potential of endozoochorous seed dispersal by sheep in calcareous grasslands: correlations with seed traits. Applied Vegetation Science 13, 163–172.
| Potential of endozoochorous seed dispersal by sheep in calcareous grasslands: correlations with seed traits.Crossref | GoogleScholarGoogle Scholar |
Langlands, J. P., and Bennett, I. L. (1973). Stocking intensity and pastoral production: II. Herbage intake of Merino sheep grazed at different stocking rates. The Journal of Agricultural Science 81, 205–209.
| Stocking intensity and pastoral production: II. Herbage intake of Merino sheep grazed at different stocking rates.Crossref | GoogleScholarGoogle Scholar |
Liu, X. M., Chen, H. Y., Zheng, R., Wu, Y., Arengaowa, , and Wang, R. R. (2011). Decomposition of sheep and cattle dung in a Steppe in Inner Mongolia, China. Chinese Journal of Applied and Environmental Biology 17, 791–796.
Lönnberg, K., and Eriksson, O. (2013). Rules of the seed size game: contests between large-seeded and small-seeded species. Oikos 122, 1080–1084.
| Rules of the seed size game: contests between large-seeded and small-seeded species.Crossref | GoogleScholarGoogle Scholar |
Mac-Arthur, R. H., and Wilson, E. O. (1967). ‘The Theory of Island Biogeography.’ (Princeton University Press: Princeton, NJ, USA.)
Malo, J. E., and Suárez, F. (1995). Establishment of pasture species on cattle dung: the role of endozoochorous seeds. Journal of Vegetation Science 6, 169–174.
| Establishment of pasture species on cattle dung: the role of endozoochorous seeds.Crossref | GoogleScholarGoogle Scholar |
Malo, J. E., and Suárez, F. (2000). Herbivore dunging and endozoochorous seed deposition in a Mediterranean Dehesa. Journal of Range Management 53, 322–328.
| Herbivore dunging and endozoochorous seed deposition in a Mediterranean Dehesa.Crossref | GoogleScholarGoogle Scholar |
Mancilla‐Leytón, J. M., Fernández‐Alés, R., and Vicente, A. M. (2011). Plant–ungulate interaction: goat gut passage effect on survival and germination of Mediterranean shrub seeds. Journal of Vegetation Science 22, 1031–1037.
| Plant–ungulate interaction: goat gut passage effect on survival and germination of Mediterranean shrub seeds.Crossref | GoogleScholarGoogle Scholar |
Manzano, P., Malo, J. E., and Peco, B. (2005). Sheep gut passage and survival of Mediterranean shrub seeds. Seed Science Research 15, 21–28.
| Sheep gut passage and survival of Mediterranean shrub seeds.Crossref | GoogleScholarGoogle Scholar |
Milotić, T., and Hoffmann, M. (2016). How does gut passage impact endozoochorous seed dispersal success? in a spatially heterogeneous landscape. Plant Ecology 178, 149–162.
Mouissie, A. M., Vos, P., Verhagen, H. M. C., and Bakker, J. P. (2005). Endozoochory by free-ranging, large herbivores: ecological correlates and perspectives for restoration. Basic and Applied Ecology 6, 547–558.
| Endozoochory by free-ranging, large herbivores: ecological correlates and perspectives for restoration.Crossref | GoogleScholarGoogle Scholar |
Myers, J. A., Vellend, M., Gardescu, S., and Marks, P. L. (2004). Seed dispersal by white-tailed deer: implications for long-distance dispersal, invasion, and migration of plants in eastern North America. Oecologia 139, 35–44.
| Seed dispersal by white-tailed deer: implications for long-distance dispersal, invasion, and migration of plants in eastern North America.Crossref | GoogleScholarGoogle Scholar | 14740288PubMed |
Pakeman, R. J., Digneffe, G., and Small, J. L. (2002). Ecological correlates of endozoochory by herbivores. Functional Ecology 16, 296–304.
| Ecological correlates of endozoochory by herbivores.Crossref | GoogleScholarGoogle Scholar |
Peco, B., Lopez-Merino, L., and Alvir, M. (2006). Survival and germination of Mediterranean grassland species after simulated sheep ingestion: ecological correlates with seed traits. Acta Oecologica 30, 269–275.
| Survival and germination of Mediterranean grassland species after simulated sheep ingestion: ecological correlates with seed traits.Crossref | GoogleScholarGoogle Scholar |
Picard, M., Papaïx, J., Gosselin, F., Picot, D., Bideau, E., and Baltzinger, C. (2015). Temporal dynamics of seed excretion by wild ungulates: implications for plant dispersal. Ecology and Evolution 5, 2621–2632.
| Temporal dynamics of seed excretion by wild ungulates: implications for plant dispersal.Crossref | GoogleScholarGoogle Scholar | 26257875PubMed |
Smith, C. C., and Fretwell, S. D. (1974). The optimal balance between size and number of offspring. American Naturalist 108, 499–506.
| The optimal balance between size and number of offspring.Crossref | GoogleScholarGoogle Scholar |
Soons, M. B., Vlugt, C. V. D., Lith, B. V., Heil, G. W., and Klaassen, M. (2008). Small seed size increases the potential for dispersal of wetland plants by ducks. Journal of Ecology 96, 619–627.
| Small seed size increases the potential for dispersal of wetland plants by ducks.Crossref | GoogleScholarGoogle Scholar |
Sun, Y. B., Zhou, J., and An, Z. S. (2001). The late Cenozoic eolian deposits in the loess plateau and the aridity of eolian dust source region. Earth Science Frontiers 8, 77–81.
Thompson, K., Band, S. R., and Hodgson, J. G. (1993). Seed size and shape predict persistence in soil. Functional Ecology 7, 236–241.
| Seed size and shape predict persistence in soil.Crossref | GoogleScholarGoogle Scholar |
Traveset, A., Verdú, M., Levey, D. J., Silva, W. R., and Galetti, M. (2002). A meta-analysis of the effect of gut treatment on seed germination. Seed Dispersal and Frugivory Ecology Evolution and Conservation 91, 334–339.
Wang, S., Lu, W., Narkes, W., Ma, C., Zhang, Q., and Wang, C. (2017). Recovery and germination of seeds after passage through the gut of Kazakh sheep on the north slope of the Tianshan Mountains. Seed Science Research 27, 43–49.
| Recovery and germination of seeds after passage through the gut of Kazakh sheep on the north slope of the Tianshan Mountains.Crossref | GoogleScholarGoogle Scholar |
Wang, L., Gan, Y. T., Wiesmeier, M., Zhao, G. Q., Zhang, R. Y., Han, G. D., Siddique, K. H. M., and Hou, F. J. (2018a). Grazing exclusion – an effective approach for naturally restoring degraded grasslands in Northern China. Land Degradation & Development 10, 1–18.
Wang, S., Lu, W., Chen, Y., and Jing, P. (2018b). Effect of sheep digestive tract on the recovery and germination of seeds of fifteen leguminous plants in the northern Xinjiang region, China. Acta Phytoecologica Sinica 42, 185–194.
Whitacre, M. K., and Call, C. A. (2006). Recovery and germinability of native seed fed to cattle. Western North American Naturalist 66, 121–128.
| Recovery and germinability of native seed fed to cattle.Crossref | GoogleScholarGoogle Scholar |
Whitford, W. G., Steinberger, Y., and Ettershank, G. (1982). Contributions of subterranean termites to the ‘economy’ of Chihuahuan desert ecosystems. Oecologia 55, 298–302.
| Contributions of subterranean termites to the ‘economy’ of Chihuahuan desert ecosystems.Crossref | GoogleScholarGoogle Scholar | 28309968PubMed |
Willms, W. D., and Quinton, D. A. (1995). Grazing effects on germinable seeds on the fescue prairie. Journal of Range Management 48, 423–430.
| Grazing effects on germinable seeds on the fescue prairie.Crossref | GoogleScholarGoogle Scholar |
Winkel, V. K., and Roundy, B. A. (1991). Effects of cattle trampling and mechanical seedbed preparation on grass seedling emergence. Journal of Range Management 44, 176–180.
| Effects of cattle trampling and mechanical seedbed preparation on grass seedling emergence.Crossref | GoogleScholarGoogle Scholar |
Yu, X., Xu, C., Wang, F., Shang, Z., and Long, R. (2013). Levels of germinable seed in Topsoil and Yak dung on an alpine meadow on the North-East Qinghai-Tibetan Plateau. Journal of Integrative Agriculture 12, 2243–2249.
| Levels of germinable seed in Topsoil and Yak dung on an alpine meadow on the North-East Qinghai-Tibetan Plateau.Crossref | GoogleScholarGoogle Scholar |