

Seeing is believing: what visualising bubbles in the xylem has revealed about plant hydraulic function
Amanda A. Cardoso A * , Cade N. Kane B , Ian M. Rimer B and Scott A. M. McAdam B
A * , Cade N. Kane B , Ian M. Rimer B and Scott A. M. McAdam B
A Department of Crop and Soil Sciences, North Carolina State University, Raleigh, NC 27695, USA.
B Purdue Center for Plant Biology, Department of Botany and Plant Pathology, Purdue University, West Lafayette, IN, USA.
Functional Plant Biology 49(9) 759-772 https://doi.org/10.1071/FP21326
Submitted: 8 November 2021 Accepted: 29 May 2022 Published: 20 June 2022
© 2022 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Maintaining water transport in the xylem is critical for vascular plants to grow and survive. The drought-induced accumulation of embolism, when gas enters xylem conduits, causes declines in hydraulic conductance (K) and is ultimately lethal. Several methods can be used to estimate the degree of embolism in xylem, from measuring K in tissues to directly visualising embolism in conduits. One method allowing a direct quantification of embolised xylem area is the optical vulnerability (OV) technique. This method has been used across different organs and has a high spatial and temporal resolution. Here, we review studies using the OV technique, discuss the main advantages and disadvantages of this method, and summarise key advances arising from its use. Vulnerability curves generated by the OV method are regularly comparable to other methods, including the centrifuge and X-ray microtomography. A major advantage of the OV technique over other methods is that it can be simultaneously used to determine in situ embolism formation in leaves, stems and roots, in species spanning the phylogeny of land plants. The OV method has been used to experimentally investigate the spreading of embolism through xylem networks, associate embolism with downstream tissue death, and observe embolism formation in the field.
Keywords: cavitation, drought, embolism, imaging methods, optical vulnerability, plant hydraulics, vascular plants, xylem network.
Introduction
The transport of water within specialised conduits in the xylem is essential for vascular plant function, supplying water to photosynthetic tissues to enable growth and reproduction (Brodribb et al. 2007; Skelton et al. 2017b; Xiong and Nadal 2020). When stomata on leaves open during the day to facilitate CO2 entry for photosynthesis, water vapour is lost from the leaf interior to the atmosphere via transpiration (Raven 2002; Brodribb et al. 2020b). This evaporation lowers water potential (Ψ) creating negative pressure on the water column and driving a flux of water from hydrated soils to the leaves.
During extended periods of soil drought, especially when associated with high evaporative demand, negative pressure increases within the xylem conduits, increasing the likelihood of air bubbles being pulled into the conduit lumen, expanding and forming an air cavity that blocks water flow though that conduit (i.e. embolism) (Tyree and Sperry 1989a; Kaack et al. 2021). As drought progresses, xylem function is threatened by hydraulic failure, which occurs when embolism spreads throughout the xylem network and causes a progressive decline in hydraulic conductance (K) that can ultimately result in plant death (Sperry and Tyree 1988; Torres-ruiz et al. 2016; Choat et al. 2018; Johnson et al. 2020; Guan et al. 2021).
Xylem vulnerability (or resistance) to hydraulic failure can be quantified by vulnerability curves (VCs) that describe the loss of K (or the accumulation of embolism) as a function of Ψ. Many of the earliest VCs were determined by quantifying the effect of Ψ on K (Sperry et al. 1988; Tyree and Sperry 1989a), with progressive declines in K during drought being attributed to increased embolism. Hydraulic VCs can be constructed in a number of different plant organs (Sperry et al. 1988), gravimetrically (Pockman et al. 1995), through measurements of rehydration kinetics (Brodribb and Cochard 2009) and evaporative flux from leaves (Brodribb and Holbrook 2006), or by measuring flow through xylem driven by centrifugation (i.e. Cavitron) (Cochard et al. 2013).
VCs can also be constructed by methods that associate physical phenomena with a quantification of xylem embolism. These include recordings of acoustic emissions (Tyree and Sperry 1989b) or the volume of air expelled from tissue under vacuum (i.e. the pneumatic method) (Pereira et al. 2016), both being associated with the amount of embolised conduits. There are also methods that directly distinguish between air-filled and fluid-filled conduits; e.g. dye colouration of functional xylem conduits (Hietz et al. 2008), cryo-scanning electron microscopy (cryo-SEM) (Cochard et al. 2000), magnetic resonance imaging (MRI) (Holbrook et al. 2001), X-ray microtomography (micro-CT) (Brodersen et al. 2010), and more recently, the optical vulnerability (OV) technique (Brodribb et al. 2016a).
The OV technique allows for the direct visualisation of embolism-induced changes in light transmission through or reflectance of the xylem while plants desiccate (Brodribb et al. 2016a). This technique can be conducted using relatively common and affordable equipment, which contrasts with other embolism visualisation techniques such as cryo-SEM, MRI and micro-CT, which are limited to researchers that have access to these specialised technologies. Thus, the OV method provides a highly accessible and affordable method for investigating xylem hydraulic function to a global scientific workforce (Cardoso et al. 2020b; Levionnois et al. 2020).
In this review, we conducted a meta-analysis of studies using the OV method that have been published following the development of this method (Brodribb et al. 2016a). First, we provide a detailed description of how the method works and how VCs are constructed using this approach, compare methods for determining cardinal Ψ from an optical VC, and discuss the limitations of this technique. Then, we summarise key insights related to xylem function that have been gained from the use of this method, including: (1) the association between xylem embolism and declines in K; (2) embolism formation and spread; and (3) xylem resistance to embolism across species and organs. Finally, we provide future perspectives on the use of this method in the plant hydraulic field.
Visualising xylem embolism and constructing vulnerability curves using the OV technique
The development of the OV method was the first, simple, and affordable method that could provide a clear visualisation of embolism formation and spread through the xylem network through rapid changes in light transmission associated with embolised, gas-filled xylem (Brodribb et al. 2016a). To date, this method has been applied to visualising drought-induced embolism events in stems (Brodribb et al. 2017; Rodriguez-Dominguez et al. 2018; Gauthey et al. 2020), roots (Rodriguez-Dominguez et al. 2018; Li et al. 2020b), leaves (Brodribb et al. 2016b; Cardoso et al. 2018; Rodriguez-Dominguez et al. 2018; Cardoso et al. 2020a; Chen et al. 2021; Guan et al. 2021; Jacob et al. 2022), and floral parts (Zhang and Brodribb 2017; Bourbia et al. 2020) (see Supplementary Table S1). A key benefit of using the OV method to construct VC is that in most cases, it does not require the use of excised segments, which places it amongst a small number of purely non-invasive techniques for quantifying hydraulic function, making it useful for tracking dynamic embolism formation in intact plants in the field (Wagner et al. 2022). Numerous studies have utilised intact plants when constructing OV method VCs (Skelton et al. 2017a; Cardoso et al. 2018, 2020a; Johnson et al. 2018; Corso et al. 2020; Gauthey et al. 2020; Li et al. 2020b). To date, VCs of a dozen economically important herbaceous species as well as early-diverging species (i.e. ferns, lycophytes, and Polytrichalean bryophytes) have been constructed using intact plants (Skelton et al. 2017a; Cardoso et al. 2018, 2019; Johnson et al. 2018; Brodribb et al. 2020a; Corso et al. 2020; Suissa and Friedman 2021).
Vulnerability curves based on the OV method are often constructed using fully hydrated plants or organs, given that this method can only detect new embolism events, with a limited capacity to determine the extent of pre-existing embolism in the xylem. Intact plants can be taken to the laboratory and the soil washed from the roots to accelerate plant dehydration (Cardoso et al. 2018). Whenever intact whole plants are not available, such as plants grown in natural settings, long branches exceeding the length of the longest vessel can also be excised under water and transported to the laboratory so that the OV method can be performed in stems (Skelton et al. 2021), leaves (Guan et al. 2021), or in both simultaneously. Excessive transpiration can lead to Ψ decline and embolism events occurring before imaging commences, and to avoid this, transpiration from excised material can be reduced by bagging and maintaining under low light levels (<10 μmol quanta m−2 s−1). To capture images, a range of devices have been used, providing a considerable advantage in resource poor scientific communities. They include stereo microscopes with cameras (Cardoso et al. 2018), custom-made clamps (http://www.opensourceov.org) (Johnson et al. 2020; Avila et al. 2021), flatbed scanners (Hochberg et al. 2017; Lucani et al. 2019), or mobile phone cameras (Petruzzellis et al. 2020), all of which can provide sufficient spatial resolution to capture changes in light transmittance or reflectance on the formation of an embolism (Brodribb et al. 2016a; Cardoso et al. 2018; Rodriguez-Dominguez et al. 2018; Johnson et al. 2020).
Reflected light is generally ideal for imaging stems, while transmitted light is often ideal for leaves (Brodribb et al. 2016b, 2017; Avila et al. 2021). Changes in visual light transmission caused by embolism can be readily observed in intact heterobaric leaves, roots and flower parts. Homobaric leaves can also be successfully imaged when imaged close to the midrib (Brodribb et al. 2016b). When leaves are imaged on excised branches that exceed maximum vessel length, there appears to be little impact of the excised end on the determination of leaf vulnerability (Brodribb et al. 2016b; Zhang and Brodribb 2017; Skelton et al. 2018; Levionnois et al. 2020; Guan et al. 2021). Care should be taken not to harvest individual leaves or leaves into which vessels extend from the excision, as this results in the rapid propagation of embolism through the leaf at very modest Ψ (Guan et al. 2021). In stems, a small area of periderm must be carefully removed so the xylem can be observed (Brodribb et al. 2017; Johnson et al. 2020). Touching the xylem when removing the periderm may induce localised embolism and must be avoided. A layer of adhesive hydrogel applied to the exposed xylem is required to improve light transmission and reduce evaporative water loss (Johnson et al. 2020). Even though debarking the stem to expose the xylem has been questioned, with Venturas et al. (2019) claiming it might damage the xylem and alter the way this tissue will respond to dehydration, the amount of artificial embolism induced by careful bark removal and the application of hydrogel is negligible (Johnson et al. 2020). For species where the periderm cannot be easily removed without damage to the xylem, or in species with a vascular stele such as lycophytes and ferns, thin paradermal sections can be made to expose the xylem before the hydrogel is applied (Cardoso et al. 2020c; Suissa and Friedman 2021).
Materials dehydrating slowly (c. a few days) under dark conditions (to allow whole plant Ψ equilibrium) are used to image the xylem to construct OV curves, and imaging frequency is generally in the range of 1–3 mHz. The Ψ can be assessed using two primary methods, although others are potentially available (Dainese et al. 2022). A psychrometer (ICT PSY Armidale, Australia) is typically attached to the stem and readings are taken at a frequency of 0.5–1.6 mHz. At the same time, a regular pressure chamber can be used to periodically cross-validate these stem Ψ measurements using leaves (Brodribb et al. 2016b; Cardoso et al. 2018; Gauthey et al. 2020; Johnson et al. 2020). Although both measurements of Ψ often match (Gauthey et al. 2020), the reliability of xylem Ψ measured in leaves using the pressure chamber once extensive embolism has accumulated is low (Ritchie and Hinckley 1971). Thus, the use of a stem psychrometer ensures an accurate estimation of Ψ, especially in species in which vulnerability segmentation occurs, whereby leaves are less resistant to embolism than stems (Charrier et al. 2016; Levionnois et al. 2020; Avila et al. 2021). Upon completion of desiccation and the cessation of embolism formation, a regression (mostly but not always linear) between the drying time and Ψ measurements is yielded and used to describe the change in water status as plants dehydrate (Brodribb et al. 2016b; Cardoso et al. 2018).
Image sequences are terminated when all conduits are embolised (Brodribb et al. 2016a). To ensure that all embolism events are captured, images can be taken for several hours after the last embolism event is registered (Cardoso et al. 2018) and finally by severing the organ within 2 cm of the imaged area while still imaging can be used to ensure no additional embolism events were missed. Following data collection, embolism events are identified from the image sequence using a simple image subtraction method described by Brodribb et al. (2016a) and Brodribb et al. (2016b). The most common approach is the use of open-source ImageJ (NIH) software where successive pairs of images are subtracted, yielding the embolism-induced changes in the light transmission between the two images, whenever embolism is present. Noise from image subtraction needs to be next removed through a series of tools provided by ImageJ, such as smothering filters to increase the difference between noise and embolism events and thresholding the image sequence by choosing a threshold that maintains the maximum extent of events and minimisation of background noise. Any potential residual noise and small artefacts must then be manually removed, which is the most time-consuming aspect of this method (see below for common examples of noise and artifacts in image collection in the limitations section). A more detailed description on how to capture, process and analyse image stacks, can be found at http://www.opensourceov.org/. This website also provides users with the OpenSourceOV ImageJ Toolbox; i.e. a special macro for ImageJ (Fiji) containing several functions to facilitate data analysis. The manual image processing of vulnerability curves using ImageJ software is labour intensive, but there are reports of the potential development and utilisation of AI technology which has the potential to greatly enhance the speed of image processing (Y. Sorek, pers. comm.; https://twitter.com/YonatanSorek/status/1417164616044589061). This would be a widely adopted advance as image analysis remains the primary disadvantage of the OV method in terms of human hours and computer processing power for the rapid generation of VC.
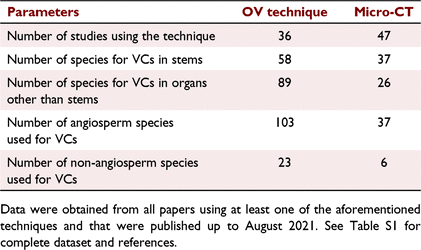
A key benefit of the OV method is that it requires a lower number of biological samples to construct VCs when compared with other methods that directly quantify xylem embolism, particularly those that do so in leaves. By using the OV method, a complete vulnerability curve can be obtained in a single leaf, petal, or stem. Most other methods utilise several organs (often exceeding 20) to construct each VC (Table S1). This is because samples can only be measured once (Savi et al. 2017). Additionally, the OV method can be performed in almost any laboratory (Petruzzellis et al. 2020), contrasting with the cryo-SEM, MRI and micro-CT methods that are geographically restricted to a few locations, or are costly. The combination of both simplicity and reliability contrasts with these other techniques (Cochard et al. 2019; Cardoso et al. 2020b). To date, VCs have been performed in 126 plant species since its development in 2016. When compared with the micro-CT, for instance, which was developed in 2010, the OV method has been utilised in less studies (36 compared to 47) but in considerably more species (126 compared to 43) and to construct VCs of several soft organs not often visualised by micro-CT (e.g. leaves, roots, floral parts in angiosperms as well as stipes and rhizomes in ferns and lycophytes) (Table 1; Table S1).
|
The use of micro-CT to validate embolism resistance quantified through the OV method
Several studies have compared the P50(water potential inducing 50% embolism in the xylem or 50% loss in K, depending on the method) obtained from VCs using the OV method with other hydraulic of imaging-based techniques for determining xylem embolism resistance. There are many papers that report similar P50 values across a diversity of species when embolism is determined using micro-CT and the OV method in stems (Cardoso et al. 2020c; Gauthey et al. 2020; Pratt et al. 2020) and leaves (Emilio et al. 2019; Corso et al. 2020) (closed symbols; Fig. 1a; Supplementary Table 2). In comparing the P50 values obtained using micro-CT and the OV methods for stems and leaves when published in separate studies but in the same species, we also found strong congruence between values (open symbols; Fig. 1a; Table S2). Across leaves and stems of 20 species and 20 studies, the relationship between the micro-CT determined P50 and the OV determined P50 yielded a regression that closely agreed with the 1:1 relationship (Fig. 1a) (r = 0.96, P < 0.0001).

|
The strong agreement between the micro-CT P50 and OV P50 (Fig. 1a), supports the conclusions of Johnson et al. (2020) who performed micro-CT and the OV method in the same stems and found that the OV method reliably captured xylem embolism events to a depth of 0.3 mm into the stem and that this range of analysis is representative of embolism across the entire stem, when small stems are measured. Minimal congruence between these two methods is reported in one study examining the branches Populus trichocarpa Torr. & A. Gray (Venturas et al. 2019) (brown symbol pointed by an arrow in Fig. 1a). However, a number of unconventional approaches were taken in the generation of the OV data in this study. These authors utilised silicone vacuum grease on the exposed and imaged stem surface rather than a transparent hydrogel, which might have resulted in air entry to the xylem or obscured minor embolism events. Secondly, when determining the VC from the image stack, the authors considered all embolism events as equal in value despite observing differences in the amount of pixels per embolism event, which is known to represent differences in the embolised xylem area. This procedure for embolism determination renders the curve an accumulation of embolism event numbers (not sizes) across Ψ, and this curve has no interpretive value for hydraulic function whatsoever. There is also some disagreement between the OV method determined P50 of −8.32 MPa and the micro-CT determined P50 of −6.46 MPa for leaves of Laurus nobilis L. (green symbol pointed by an arrow in Fig. 1a). It is possible, however, that this variation occurred due to a high intra-specific variability in leaf embolism resistance whitin this species, given that leaves were collected from different individuals adapted to different regions in Europe (Klepsch et al. 2018; Lamarque et al. 2018; Rodriguez-Dominguez et al. 2018). A ∼1.50 MPa variation in embolism resistance between leaves in the same canopy has been observed in other Lauraceae species and has been attributed to being the cause of partial leaf and canopy death during drought (Cardoso et al. 2020a). There are also reports of considerable variation across the canopy in branchlet embolism resistance in the highly drought resistant conifer Callitris rhomboidea R. Br. (Johnson et al. 2022), and these differences have also been associated with differences in the lethal Ψ threshold for branch survival under drought.
Limitations to OV method
A primary limitation to performing the OV method is the considerable human hours associated with analysing images to generate the VCs. Although algorithms can be customised to identify and quantify embolism events during imaging analyses (Hochberg et al. 2017, 2019), all images need to be further manually analysed to remove any potential remaining noise originated from movements, insects, tissue drying fronts, bubbles in the hydrogel, and minor changes to light environment. In this regard, automating the analysis of the VCs yielded with the OV method would be a considerable improvement to the utility of this method for the screening of phenotypes across large populations. This, however, might be improbable in the near future given the diverse nature of interference to an image stack.
Another intrinsic limitation to the OV method is that it cannot detect native or pre-existing embolism (Chen et al. 2021), which can impact quantification of relative xylem embolism resistance (Avila et al. 2022), and the translation of embolised xylem area to a percent loss of K. As such, only plants cultivated under well-water conditions, plants that have been previously exposed to drought but are known to refill their xylem (Cardoso et al. 2018), and branches and leaves formed during wet seasons (which are expected to have very low levels of pre-existing embolism) are ideal to construct VCs with the OV method. Recently, it has been suggested that significant native embolism produced by drought is restricted to very extreme events in field grown trees, so the issue of pre-existing embolism on accurate VC determination in field grown plants may be minimal (Guan et al. 2022; Wagner et al. 2022). It is important to maintain the plant material in plastic bags with damp paper towels until the beginning of experiments whenever collecting branches in the field to construct VCs in order to prevent excessive water loss and any potential embolism prior to analysis (Skelton et al. 2021). Although the OV method cannot directly quantify the native embolism level, it has been very recently demonstrated that it is possible to indirectly assess the level of native embolism with high confidence by utilising the VCs obtained using this method and measurements of the lowest Ψ experienced by the plant in the field (Chen et al. 2021).
Similar to other methods that directly assess embolism resistance, the OV method is liable to error due to open vessel artifacts if the branch or leaf being measured is shorter than the longest vessel (Guan et al. 2021). Hence, knowledge of maximum vessel length is required prior to harvesting branches for image analysis, so that images can be captured beyond the length of the longest open vessel (Martin-StPaul et al. 2014; Torres-Ruiz et al. 2014). During analysis, there is a number of major classes of artefacts that appear in image subtractions that could be confused for embolism events to an untrained eye (e.g. movement, drying from post embolism, and insects), although embolism events are readily apparent in image subtractions (Fig. 2).

|
Movement
During the process of capturing images, stems and leaves might move if not firmly secured. In some species with large volume changes in stems or leaves during dehydration, movement will occur during imaging regardless of how secure the sample is, particularly prior to the formation of embolism (Johnson et al. 2018). If there is a sudden movement of the sample, it could appear as a widespread change, or localised changes, during image subtraction. This can be avoided, when possible, by making sure the sample is secured during imaging. An alternative is to increase the imaging rate, although this results in trade-off with processing time. Imaging at a frequency of 1.0 Hz could eliminate most movement noise while still capturing embolism events, at the expense of considerable processing power and time to analyse the larger image stacks. If movements do occur while taking imaging at a lower frequency, they are often easy to recognise as distinct from embolism events, as the movement will affect large or localised regions of the sample area in one image rather than events in individual xylem elements (Fig. 3). The hydrogel, or bubbles in the hydrogel, may also move with time in the imaged area, but the movement is very slow and can easily be recognised in the raw images.

|
Drying front post embolism
The image subtractions may also identify a sequential series of colour changes even if the sample is firmly secured during the final stages of drying after most, if not all embolism events have occurred. This results from a rapidly moving drying front, often observed in leaves as the mesophyll desiccates. A drying front has a tendency to spread from a location and radiates across the imaged area in each sequential subtraction (Fig. 4).

|
Insects
If the plant material is collected from plants grown in field settings, the presence of insects may be recognisable during image subtractions and it needs to be deleted from image sequences. Smaller bugs can also move onto a leaf or stem, attracted by the light used for imaging. Insects are easily distinguishing from embolism events during image subtraction because they appear as small circles that occur in a random nature in single frames. Additionally, the organisms are visible and easy to be recognised in the raw images (Fig. 5). It is also possible that larger insects may appear and fully obscure the field of view. Phloem feeding insects that can move onto the leaf during imaging may be observed feeding, and there are reports that such feeding can cause an embolism events (Kim 2013). To reduce the impact of insects on OV curve collection, damp paper towels can gently clean both leaf surfaces, or insecticide can be used to greatly reduce insect load.

|
Light intensity
The visualisation of embolism using the OV method is based on changes in light, either transmitted or reflected, meaning that any change in ambient or applied light intensity could cause an artefact across the whole sample or in one region of the imaged sample. This could happen, for example, if using a microscope to image, and the light levels in the room fluctuate. To avoid this error, a cylindrical or conical shield can be placed over the sample to reduce the effect of any sudden changes in ambient light intensity. Light must also be constant and consistent, which can be challenging to maintain when using a flatbed scanner. It is also important to appropriately set the light intensity or camera exposure time at the beginning of an experiment, to avoid over saturation of the vessels at high light levels, thus rendering embolism events undetectable in the images, or not visible at low light intensities. Also, specific requirements for intensity need to be evaluated for each species individually.
Limitations to analysis
There are some species in which generating VC using the OV method is not possible. Our research group have had issues measuring both leaves and stems of certain species, and we assume that these issues can be attributed to two broad anatomical causes. First, wood colour is one major limitation when assessing stem vulnerability. Once the periderm is removed most species have greenish or yellowish sap wood that changes in colour on embolism formation; i.e. a change that is apparent to the naked eye and after imaging. In contrast, other species including Aesculus glabra Willd. and Magnolia salicifolia Maxim. have very pale sapwood, so that embolism is very challenging to visualise even when using directed light and high-resolution microscopic imaging. Second, thick, homobaric leaves also pose a challenge for generating VCs using the OV method. Leaf veins require the transmission of light to capture embolism events and if a leaf has no bundle-sheath extensions (like in homobaric leaves) and is thick, very little light will transmit through the veins. We have found this to be an issue in some species such as Ginkgo biloba L.. A simple test to check whether a leaf will be appropriate for image analysis is to hold it up to the sun or high light source and see if midribs and minor order veins are visible. If so, then OV vulnerability curves should be accessible. In the case of homobaric leaves, one option is to generate OV curves for leaves from imaging the midrib solely (Lucani et al. 2019).
Another intrinsic limitation of the OV method remains in understanding how much the hydraulic vulnerability, especially in leaves or soft organs, is impacted by the percentage of cumulative embolised area of the xylem. Although, most data collected so far demonstrate a close association between the OV P50 and the hydraulic P50 (Fig. 1b; Supplementary Table 2), this may not occur in all species (see below).
Issues with determining cardinal water potentials from OV curves
Most plant hydraulic physiologists are interested in comparing the embolism resistance of genotypes or species. A key metric for this comparison is P50 but other cardinal Ψ such as Pe (the Ψ of first embolism), P12 (the Ψ when 12% of xylem is embolised or conductivity is lost) and P88 (the Ψ when 88% of the xylem is embolised or conductivity is lost) are often used. P88 is often associated with leaf or canopy death in angiosperms (Brodribb and Cochard 2009). A key advantage of the OV method is that these cardinal Ψ values can be taken directly from VCs. However, issues can arise when trying to extract Ψ values for a particular % embolism when individual embolism events span that threshold. Often values are taken from the Ψ when an embolism event spanned the given cardinal threshold. Curve fitting is commonly used to determine cardinal Ψ from invasively generated VCs but it is unnecessary and can be even problematic for determining cardinal Ψ from OV curves (Fig. 6). Using a dataset of vulnerability curves from six leaves of Persea americana Mill., we show that mean Ψ at P50 taken directly from the curves is −2.24 ± 0.08 MPa. This value is similar to the P50 estimated from a fitted Weibull model −1.91 ± 0.01 MPa and to the mean of individually fitted Weibull functions to the data obtained on individual leaves (−2.24 ± 0.08 MPa) (Table 2). Differences, however, arose when comparing P12 and P88 values for fitted curves with the absolute values of embolism resistance (Table 2). Thus, we recommend that mean Ψ and standard errors are calculated and plotted rather than using fitted curves for OV curves as per Avila et al. (2021).

|

|
What we have learned about water transport in the xylem by utilising the OV method
Associating xylem embolism with declines in hydraulic conductance
Xylem embolism has long been known to reduce K (Sperry et al. 1988; Tyree and Sperry 1989a). Thus, methods that quantify declines in K were believed to measure embolism formation (Sperry et al. 1988; Cochard et al. 1992, 1999, 2005, 2013; Stout and Sala 2003; Stiller 2009; Awad et al. 2010). The development of reliable methods directly quantifying xylem embolism, mainly the micro-CT and more recently, the OV method, have allowed researchers to test whether declines in K are always the result of xylem embolism formation, irrespective of the plant organ or species. The OV P50 for the stems of several species closely matches the P50 obtained with the Cavitron (in situ flow centrifuge), which assess declines in K (Brodribb et al. 2017). In leaves, the hydraulic P50 determined by rehydration kinetics also matches the OV determined P50 in several plant species (Brodribb et al. 2016b; Skelton et al. 2017a; Corso et al. 2020). A strong agreement between the OV method-determined P50 and the hydraulic method-P50 can be observed across the literature (Fig. 1b; Table S2).
The main progress attributed to methods directly quantifying xylem embolism (including the OV) relies in demonstrating that although xylem embolism will always result in decreased K, the later might also occur prior to the formation of embolism as a result of extra-xylary events. Early studies confirmed these reductions to K outside of the xylem utilising cryo-SEM (Cochard et al. 2004; Zhang et al. 2014) and micro-CT (Bouche et al. 2016) in the leaves of conifers (Taxus baccata L., Pinus spp., and Podocarpus grayi de Laub.), which can experience considerable shrinkage and deformation of transfusion tracheids during drought, resulting in K declines in the absence of xylem embolism (Brodribb and Holbrook 2005). In addition, recent studies have demonstrated with measurements of evaporative flux determined leaf K and micro-CT that leaves from a number of woody angiosperm species experience declines in leaf K decoupled from xylem embolism (Scoffoni et al. 2017), a result also demonstrated for leaves of Caragana spp. (green symbols pointed by black arrows in Fig. 1b) by comparing data from the OV method with evaporative flux determined VCs (Yao et al. 2021). Finally, by comparing VCs obtained hydraulically, by rehydration kinetics, and the OV method, the shoot K of two Selaginella spp. have been recently demonstrated to decline considerably prior to embolism formation in the stem (brown symbols pointed by red arrows in Fig. 1b) (Cardoso et al. 2020c). In the later study, the utilisation of micro-CT revealed that an air-filled lacuna formed in the stems when plants of Selaginella were dehydrated, disconnecting the central vascular stele from the cortex and likely explaining the relatively hydrated declines in the shoot K (Cardoso et al. 2020c).
In the case of soft organs, such as leaves, flower parts and fine roots, which are more likely prone to shrinkage (Scoffoni et al. 2014; Bouche et al. 2016) and observations that the K of leaves and roots relies on the conductance of several other tissues other than the xylem, including the activity of membrane aquaporins (Sade et al. 2014; Buckley et al. 2015; Moshelion et al. 2015), it is possible that outside xylary events taking place prior to xylem embolism will influence the hydraulic P50 of a number of other species. Therefore, we believe that the combination of hydraulic and observation xylem methods is a powerful approach to fully understand the sequence of physiological events taking place in plant organs during drought. Additionally, only by combining these two approaches, we will be able to thoroughly understand why agreement between the OV P50 and the hydraulic P50 is observed for some species, but not others.
Regarding potential artifacts associated with the OV method that could potentially result in contrasting hydraulic and OV P50 values, two recent studies assessing the stems of P. trichocarpa (Venturas et al. 2019) and Quercus douglasii Hook & Arn. (Pratt et al. 2020) found less negative P50 using hydraulic methods than the P50 obtained by the OV method (Pratt et al. 2020). The authors then suggested that the OV method overestimated the xylem resistance of Q. douglasii stems because this method was believed to only sample a small portion of the outer xylem tissue, which can be more resistant to embolism than the xylem of more internal xylem regions. In this study the OV curves matched micro-CT curves, which instead points to issues with the determination of embolism resistance by hydraulic means in species with long vessels (Choat et al. 2010; Cochard et al. 2010, 2013; Jansen et al. 2015; Torres-Ruiz et al. 2017).
Embolism formation and spread
In addition to insights into the threshold Ψ for embolism formation, the OV method further provides a high spatial and temporal resolution to explore the mechanisms of embolism spread throughout the xylem network. In a study utilising the OV technique, Johnson et al. (2020) demonstrated that more than 80% of the stem embolism events occurred in single vessels in three dry forest angiosperm with low connectivity xylem networks, suggesting a close association between vessel arrangement and embolism spread. In another study utilising the OV method fully embolised, excised petioles, resulted in more vulnerable leaf xylem because of an enhanced spreading of embolism in six woody angiosperm species (Guan et al. 2021). Finally, the OV method can map the spread of embolism in leaves across spatial and temporal scales for the first time, revealing that larger vein orders present the first embolism events thus leading to a widespread embolism of minor veins (Brodribb et al. 2016a). This general sequence of xylem embolism throughout the leaf veins was further confirmed for several species with different venation patterns (Brodribb et al. 2016b; Skelton et al. 2017a; Johnson et al. 2018; Avila et al. 2021).
The widespread embolism formation in leaves has been associated with leaf mortality in a study that directly assessed the level of embolism in the petiole of drought-induced necrotic leaves using the cryo-SEM (Cardoso et al. 2020a). In this study, a positive strong correlation was obtained between petiole embolism level and foliar necrotic area in a set of leaves spanning different levels of necrosis and embolism and the variation in leaf mortality across canopy was linked to an intra-specific variability in the leaf P50 obtained through the OV method (Cardoso et al. 2020a). The OV method has been used to provide the first foundational evidence confirming the connection between leaf hydraulic failure and tissue damage during water stress by exposing plants to drought, or leaves to severe evaporative stress, and associating the leaf area damaged by dehydration with the direct observation of embolism in the veins irrigating that region of the leaf using the OV method (Brodribb et al. 2021).
Xylem resistance to embolism across different species and organs
The OV method has been utilised to study more angiosperm species than any other plant group (Table 1; Fig. 7; Table S3), likely due to the larger number of species, economic importance of this lineage of land plants, and ease with which the OV method can obtain VCs in long-vesseled angiosperm species, which contrasts with prior methods that are restricted to short-vesseled and tracheid-bearing species (Choat et al. 2016; Scoffoni et al. 2017; Sergent et al. 2020). When comparing average data on the mean stem P50 of gymnosperms (n = 16) with of those of angiosperms (n = 60), we obtain statistically similar values (−4.71 and −4.81 MPa, respectively) (Fig. 7a), with Acacia aneura F. Muell. (an angiosperm) being the most resistant species in the dataset (P50 of nearly −12 MPa) (Gauthey et al. 2020). This dataset contrasts with the commonly held view that gymnosperms are more resistant to embolism than angiosperms due to the presence of torus-margo pit membranes (Maherali et al. 2004; Choat et al. 2012). It is possible that the conclusions from previous studies demonstrating gymnosperms outcompeting angiosperms in terms of embolism resistance have been draw from meta-analyses including underestimated values of P50 for several angiosperm species, and an over-sampling of mesic temperate species, that were obtained with invasive-methods (Maherali et al. 2004; Choat et al. 2012). It should be noted that we only provide a partial dataset that does not include data from a wide diversity of gymnosperm species including Callitris tuberculata R. Br., the most embolism resistant tree species so far studied (Larter et al. 2015), and that additional studies and meta-analyses are necessary in order to more accurately compare the P50 of angiosperms and gymnosperms.

|
In addition to stem vulnerability, over 100 values (n = 114) of leaf P50 have been obtained with the OV in the last 5 years (Fig. 7; Table S3). Although xylem embolism in the stem is believed to be the main driver for plant mortality (Adams et al. 2017; Choat et al. 2018; Hammond et al. 2019), leaves are the key organs responsible for photosynthesis, with any embolism having major impacts on plant productivity (Brodribb et al. 2021). Leaves also represent one of the main resistors to water flow within plants (Sack and Holbrook 2006). The large amount of data collected for angiosperm leaves using the OV method allows us to confirm a general trend towards lower embolism resistance in leaves (P50 of −3.84 MPa) when compared with stems (P50 of −4.81 MPa) (Fig. 7b), as proposed by the hydraulic vulnerability segmentation hypothesis (Tyree and Ewers 1991). However, studies determining the P50 of different organs with the OV method within the same species (Table S4), yield contrasting results, suggesting instead that hydraulic vulnerability segmentation is species-specific. It has been observed that many species do present such segmentation across organs (Zhang and Brodribb 2017; Rodriguez-Dominguez et al. 2018; Skelton et al. 2018; Bourbia et al. 2020; Li et al. 2020b; Avila et al. 2021), while many other studies using either the micro-CT or the OV method do not report any vulnerability segmentation (Skelton et al. 2017a; Losso et al. 2019; Corso et al. 2020; Li et al. 2020a; Smith-Martin et al. 2020; Peters et al. 2020; Skelton et al. 2021).
Because of the low sample number of species in which roots (P50 of −6.34 MPa, n = 2) and petals (P50 of −2.72 MPa, n = 4) in which embolism resistance using the OV method has been observed (Fig. 7b), it is hard to draw conclusions about the general vulnerability of these organs relative to other organs. In fact, roots have been observed to be more (Rodriguez-Dominguez et al. 2018), less (Peters et al. 2020) or equally resistant (Li et al. 2020b) compared to stems, depending on the species. Likewise, petals and flowers (in which embolism resistance was first assessed using the OV) have been demonstrated to display higher or similar P50 than leaves across different species (Zhang and Brodribb 2017; Bourbia et al. 2020). Conversely, a recent study utilised the OV method to assess the xylem vulnerability to embolism in peduncles of tomato and found that they are considerably more resistant than leaves, allowing this annual species to maintain fruit development during drought (Harrison Day et al. 2022).
Finally, the newly developed OV method has allowed researchers to determine variation in embolism resistance across different leaves (Fig. 8; Table S5). Until the OV was developed, no other method could determine the P50 of a single leaf, so the variation of this trait across leaves could not be assessed. Recent studies using the OV method have found that there can be a consideable variability in leaf P50 between leaves in the same species (Fig. 8). Such large intra-specific variation in the leaf P50 can result in heterogenous leaf damage across canopies (Cardoso et al. 2020a).

|
Concluding remarks
Because of its reliability associated with simplicity and accessibility, the OV method has been extensively utilised by the plant hydraulic community since its development in 2016. Through this method, researchers have confirmed several hydraulic mechanisms such as a threshold Ψ for embolism formation that is both species- and organ-specific and that embolism spread depends on the xylem conductivity and the presence of adjacent gas source. This method has also allowed researchers to demonstrate other hydraulic events for the first time such as: (1) mapping embolism spread throughout the leaf vein network; (2) intraspecific variability of leaf P50 (and even across the canopy); and (3) the direct association between leaf xylem embolism and leaf mortality. This method also provided researchers with the first measurements of embolism in floral parts and it is likely to continue being utilised to understand how embolism forms, spreads, and impacts all the plant organs (including soft ones) as well as early-diverging species such as ferns and lycophytes that have been neglected in the past.
Supplementary material
Supplementary material is available online.
Data availability
The data that support this review are available in the article and accompanying online supplementary material.
Conflicts of interest
The authors declare no conflicts of interest.
Declaration of funding
This work was supported by the USDA National Institute of Food and Agriculture (Hatch Project 1014908).
References
Adams HD, Zeppel MJB, Anderegg WRL, Hartmann H, Landhäusser SM, Tissue DT, et al. (2017) A multi-species synthesis of physiological mechanisms in drought-induced tree mortality. Nature Ecology and Evolution 1, 1285–1291.| A multi-species synthesis of physiological mechanisms in drought-induced tree mortality.Crossref | GoogleScholarGoogle Scholar | 29046541PubMed |
Avila RT, Cardoso AA, Batz TA, Kane CN, DaMatta FM, McAdam SAM (2021) Limited plasticity in embolism resistance in response to light in leaves and stems in species with considerable vulnerability segmentation. Physiologia Plantarum 172, 2142–2152.
| Limited plasticity in embolism resistance in response to light in leaves and stems in species with considerable vulnerability segmentation.Crossref | GoogleScholarGoogle Scholar | 33942915PubMed |
Avila RT, Guan X, Kane CN, Cardoso AA, Batz TA, DaMatta FM, Jansen S, Mcadam SAM (2022) Xylem embolism spread is largely prevented by interconduit pit membranes until the majority of conduits are gas-filled. Plant, Cell & Environment 45, 1204–1215.
| Xylem embolism spread is largely prevented by interconduit pit membranes until the majority of conduits are gas-filled.Crossref | GoogleScholarGoogle Scholar |
Awad H, Barigah T, Badel E, Cochard H, Herbette S (2010) Poplar vulnerability to xylem cavitation acclimates to drier soil conditions. Physiologia Plantarum 139, 280–288.
| Poplar vulnerability to xylem cavitation acclimates to drier soil conditions.Crossref | GoogleScholarGoogle Scholar | 20210873PubMed |
Bouche PS, Delzon S, Choat B, Badel E, Brodribb TJ, Burlett R, Cochard H, Charra-Vaskou K, Lavigne B, Li S, Mayr S, Morris H, Torres-Ruiz JM, Zufferey V, Jansen S (2016) Are needles of Pinus pinaster more vulnerable to xylem embolism than branches? New insights from X-ray computed tomography. Plant, Cell & Environment 39, 860–870.
| Are needles of Pinus pinaster more vulnerable to xylem embolism than branches? New insights from X-ray computed tomography.Crossref | GoogleScholarGoogle Scholar |
Bourbia I, Carins-Murphy MR, Gracie A, Brodribb TJ (2020) Xylem cavitation isolates leaky flowers during water stress in pyrethrum. New Phytologist 227, 146–155.
| Xylem cavitation isolates leaky flowers during water stress in pyrethrum.Crossref | GoogleScholarGoogle Scholar | 32130731PubMed |
Brodersen CR, McElrone AJ, Choat B, Matthews MA, Shackel KA (2010) The dynamics of embolism repair in xylem: in vivo visualizations using high-resolution computed tomography. Plant Physiology 154, 1088–1095.
| The dynamics of embolism repair in xylem: in vivo visualizations using high-resolution computed tomography.Crossref | GoogleScholarGoogle Scholar | 20841451PubMed |
Brodribb TJ, Cochard H (2009) Hydraulic failure defines the recovery and point of death in water-stressed conifers. Plant Physiology 149, 575–584.
| Hydraulic failure defines the recovery and point of death in water-stressed conifers.Crossref | GoogleScholarGoogle Scholar | 19011001PubMed |
Brodribb TJ, Holbrook NM (2005) Water stress deforms tracheids peripheral to the leaf vein of a tropical conifer. Plant Physiology 137, 1139–1146.
| Water stress deforms tracheids peripheral to the leaf vein of a tropical conifer.Crossref | GoogleScholarGoogle Scholar | 15734905PubMed |
Brodribb TJ, Holbrook NM (2006) Declining hydraulic efficiency as transpiring leaves desiccate: two types of response. Plant, Cell & Environment 29, 2205–2215.
| Declining hydraulic efficiency as transpiring leaves desiccate: two types of response.Crossref | GoogleScholarGoogle Scholar |
Brodribb TJ, Feild TS, Jordan GJ (2007) Leaf maximum photosynthetic rate and venation are linked by hydraulics. Plant Physiology 144, 1890–1898.
| Leaf maximum photosynthetic rate and venation are linked by hydraulics.Crossref | GoogleScholarGoogle Scholar | 17556506PubMed |
Brodribb TJ, Bienaimé D, Marmottant P (2016a) Revealing catastrophic failure of leaf networks under stress. Proceedings of the National Academy of Sciences of the United States of America 113, 4865–4869.
| Revealing catastrophic failure of leaf networks under stress.Crossref | GoogleScholarGoogle Scholar | 27071104PubMed |
Brodribb TJ, Skelton RP, McAdam SAM, Bienaimé D, Lucani CJ, Marmottant P (2016b) Visual quantification of embolism reveals leaf vulnerability to hydraulic failure. New Phytologist 209, 1403–1409.
| Visual quantification of embolism reveals leaf vulnerability to hydraulic failure.Crossref | GoogleScholarGoogle Scholar | 26742653PubMed |
Brodribb TJ, Carriqui M, Delzon S, Lucani C (2017) Optical measurement of stem xylem vulnerability. Plant Physiology 174, 2054–2061.
| Optical measurement of stem xylem vulnerability.Crossref | GoogleScholarGoogle Scholar | 28684434PubMed |
Brodribb TJ, Carriquí M, Delzon S, McAdam SAM, Holbrook NM (2020a) Advanced vascular function discovered in a widespread moss. Nature Plants 6, 273–279.
| Advanced vascular function discovered in a widespread moss.Crossref | GoogleScholarGoogle Scholar | 32170283PubMed |
Brodribb TJ, Sussmilch F, McAdam SAM (2020b) From reproduction to production, stomata are the master regulators. The Plant Journal 101, 756–767.
| From reproduction to production, stomata are the master regulators.Crossref | GoogleScholarGoogle Scholar | 31596990PubMed |
Brodribb T, Brodersen CR, Carriqui M, Tonet V, Rodriguez Dominguez C, McAdam S (2021) Linking xylem network failure with leaf tissue death. New Phytologist 232, 68–79.
| Linking xylem network failure with leaf tissue death.Crossref | GoogleScholarGoogle Scholar | 34164816PubMed |
Buckley TN, John GP, Scoffoni C, Sack L (2015) How does leaf anatomy influence water transport outside the xylem? Plant Physiology 168, 1616–1635.
| How does leaf anatomy influence water transport outside the xylem?Crossref | GoogleScholarGoogle Scholar | 26084922PubMed |
Cardoso AA, Brodribb TJ, Lucani CJ, DaMatta FM, McAdam SAM (2018) Coordinated plasticity maintains hydraulic safety in sunflower leaves. Plant, Cell & Environment 41, 2567–2576.
| Coordinated plasticity maintains hydraulic safety in sunflower leaves.Crossref | GoogleScholarGoogle Scholar |
Cardoso AA, Randall JA, McAdam SAM (2019) Hydraulics regulate stomatal responses to changes in leaf water status in the fern Athyrium filix-femina. Plant Physiology 179, 533–543.
| Hydraulics regulate stomatal responses to changes in leaf water status in the fern Athyrium filix-femina.Crossref | GoogleScholarGoogle Scholar | 30538169PubMed |
Cardoso AA, Batz TA, McAdam SAM (2020a) Xylem embolism resistance determines leaf mortality during drought in Persea Americana. Plant Physiology 182, 547–554.
| Xylem embolism resistance determines leaf mortality during drought in Persea Americana.Crossref | GoogleScholarGoogle Scholar | 31624082PubMed |
Cardoso AA, Billon L-M, Borges AF, Fernández-de-Uña L, Gersony JT, Güney A, Johnson KM, Lemaire C, Mrad A, Wagner Y, Petit G (2020b) New developments in understanding plant water transport under drought stress. New Phytologist 227, 1025–1027.
| New developments in understanding plant water transport under drought stress.Crossref | GoogleScholarGoogle Scholar | 32662102PubMed |
Cardoso AA, Visel D, Kane CN, Batz TA, García Sánchez C, Kaack L, Lamarque LJ, Wagner Y, King A, Torres-Ruiz JM, Corso D, Burllet R, Badel E, Cochard H, Delzon S, Jansen S, McAdam SAM (2020c) Drought-induced lacuna formation in the stem causes hydraulic conductance to decline before xylem embolism in Selaginella. New Phytologist 227, 1804–1817.
| Drought-induced lacuna formation in the stem causes hydraulic conductance to decline before xylem embolism in Selaginella.Crossref | GoogleScholarGoogle Scholar | 32386326PubMed |
Charrier G, Torres-Ruiz JM, Badel E, Burlett R, Choat B, Cochard H, Delmas CEL, Domec J-C, Jansen S, King A, Lenoir N, Martin-StPaul N, Gambetta GA, Delzon S (2016) Evidence for hydraulic vulnerability segmentation and lack of xylem refilling under tension. Plant Physiology 172, 1657–1668.
| Evidence for hydraulic vulnerability segmentation and lack of xylem refilling under tension.Crossref | GoogleScholarGoogle Scholar | 27613852PubMed |
Chen YJ, Maenpuen P, Zhang YJ, Barai K, Katabuchi M, Gao H, Kaewkamol S, Tao LB, Zhang JL (2021) Quantifying vulnerability to embolism in tropical trees and lianas using five methods: can discrepancies be explained by xylem structural traits? New Phytologist 229, 805–819.
| Quantifying vulnerability to embolism in tropical trees and lianas using five methods: can discrepancies be explained by xylem structural traits?Crossref | GoogleScholarGoogle Scholar | 32929748PubMed |
Choat B, Drayton WM, Brodersen C, Matthews MA, Shackel KA, Wada H, McElrone AJ (2010) Measurement of vulnerability to water stress-induced cavitation in grapevine: a comparison of four techniques applied to a long-vesseled species. Plant, Cell & Environment 33, 1502–1512.
| Measurement of vulnerability to water stress-induced cavitation in grapevine: a comparison of four techniques applied to a long-vesseled species.Crossref | GoogleScholarGoogle Scholar |
Choat B, Jansen S, Brodribb TJ, Cochard H, Delzon S, Bhaskar R, Bucci SJ, Feild TS, Gleason SM, Hacke UG, Jacobsen AL, Lens F, Maherali H, Martínez-Vilalta J, Mayr S, Mencuccini M, Mitchell PJ, Nardini A, Pittermann J, Pratt RB, Sperry JS, Westoby M, Wright IJ, Zanne AE (2012) Global convergence in the vulnerability of forests to drought. Nature 491, 752–755.
| Global convergence in the vulnerability of forests to drought.Crossref | GoogleScholarGoogle Scholar | 23172141PubMed |
Choat B, Badel E, Burlett R, Delzon S, Cochard H, Jansen S (2016) Noninvasive measurement of vulnerability to drought-induced embolism by X-ray microtomography. Plant Physiology 170, 273–282.
| Noninvasive measurement of vulnerability to drought-induced embolism by X-ray microtomography.Crossref | GoogleScholarGoogle Scholar | 26527655PubMed |
Choat B, Brodribb TJ, Brodersen CR, Duursma RA, López R, Medlyn BE (2018) Triggers of tree mortality under drought. Nature 558, 531–539.
| Triggers of tree mortality under drought.Crossref | GoogleScholarGoogle Scholar | 29950621PubMed |
Cochard H, Cruiziat P, Tyree MT (1992) Use of positive pressures to establish vulnerability curves. Plant Physiology 100, 205–209.
| Use of positive pressures to establish vulnerability curves.Crossref | GoogleScholarGoogle Scholar | 16652947PubMed |
Cochard H, Lemoine D, Dreyer E (1999) The effects of acclimation to sunlight on the xylem vulnerability to embolism in Fagus sylvatica L. Plant, Cell & Environment 22, 101–108.
| The effects of acclimation to sunlight on the xylem vulnerability to embolism in Fagus sylvatica L.Crossref | GoogleScholarGoogle Scholar |
Cochard H, Bodet C, Améglio T, Cruiziat P (2000) Cryo-scanning electron microscopy observations of vessel content during transpiration in walnut petioles. Facts or artifacts? Plant Physiology 124, 1191–1202.
| Cryo-scanning electron microscopy observations of vessel content during transpiration in walnut petioles. Facts or artifacts?Crossref | GoogleScholarGoogle Scholar | 11080296PubMed |
Cochard H, Froux F, Mayr S, Coutand C (2004) Xylem wall collapse in water-stressed pine needles. Plant Physiology 134, 401–408.
| Xylem wall collapse in water-stressed pine needles.Crossref | GoogleScholarGoogle Scholar | 14657404PubMed |
Cochard H, Damour G, Bodet C, Tharwat I, Poirier M, Améglio T (2005) Evaluation of a new centrifuge technique for rapid generation of xylem vulnerability curves. Physiologia Plantarum 124, 410–418.
| Evaluation of a new centrifuge technique for rapid generation of xylem vulnerability curves.Crossref | GoogleScholarGoogle Scholar |
Cochard H, Herbette S, Barigah T, Badel E, Ennajeh M, Vilagrosa A (2010) Does sample length influence the shape of xylem embolism vulnerability curves? A test with the Cavitron spinning technique. Plant, Cell & Environment 33, 1543–1552.
| Does sample length influence the shape of xylem embolism vulnerability curves? A test with the Cavitron spinning technique.Crossref | GoogleScholarGoogle Scholar |
Cochard H, Badel E, Herbette S, Delzon S, Choat B, Jansen S (2013) Methods for measuring plant vulnerability to cavitation: a critical review. Journal of Experimental Botany 64, 4779–4791.
| Methods for measuring plant vulnerability to cavitation: a critical review.Crossref | GoogleScholarGoogle Scholar | 23888067PubMed |
Cochard H, Petit G, Torres-Ruiz JM, Delzon S (2019) XIM4 meeting report, Sept. 25–27 2019, Padua (Italy): the hydraulic community has taken a step forward towards non-invasive measurements. Journal of Plant Hydraulics 6, e002
| XIM4 meeting report, Sept. 25–27 2019, Padua (Italy): the hydraulic community has taken a step forward towards non-invasive measurements.Crossref | GoogleScholarGoogle Scholar |
Corso D, Delzon S, Lamarque LJ, Cochard H, Torres-Ruiz JM, King A, Brodribb T (2020) Neither xylem collapse, cavitation, or changing leaf conductance drive stomatal closure in wheat. Plant, Cell & Environment 43, 854–865.
| Neither xylem collapse, cavitation, or changing leaf conductance drive stomatal closure in wheat.Crossref | GoogleScholarGoogle Scholar |
Dainese R, de CFL Lopes B, Tedeschi G, Lamarque LJ, Delzon S, Fourcaud T, Tarantino A (2022) Cross-validation of the high-capacity tensiometer and thermocouple psychrometer for continuous monitoring of xylem water potential in saplings. Journal of Experimental Botany 73, 400–412.
| Cross-validation of the high-capacity tensiometer and thermocouple psychrometer for continuous monitoring of xylem water potential in saplings.Crossref | GoogleScholarGoogle Scholar | 34505895PubMed |
Duursma RA, Choat B (2017) fitplc: an R package to fit hydraulic vulnerability curves. Journal of Plant Hydraulics 4, e002
| fitplc: an R package to fit hydraulic vulnerability curves.Crossref | GoogleScholarGoogle Scholar |
Emilio T, Lamarque LJ, Torres-Ruiz JM, King A, Charrier G, Burlett R, Conejero M, Rudall PJ, Baker WJ, Delzon S (2019) Embolism resistance in petioles and leaflets of palms. Annals of Botany 124, 1173–1183.
| Embolism resistance in petioles and leaflets of palms.Crossref | GoogleScholarGoogle Scholar |
Gauthey A, Peters JMR, Carins-Murphy MR, Rodriguez-Dominguez CM, Li X, Delzon S, King A, López R, Medlyn BE, Tissue DT, Brodribb TJ, Choat B (2020) Visual and hydraulic techniques produce similar estimates of cavitation resistance in woody species. New Phytologist 228, 884–897.
| Visual and hydraulic techniques produce similar estimates of cavitation resistance in woody species.Crossref | GoogleScholarGoogle Scholar | 32542732PubMed |
Guan X, Pereira L, McAdam SAM, Cao KF, Jansen S (2021) No gas source, no problem: proximity to pre-existing embolism and segmentation affect embolism spreading in angiosperm xylem by gas diffusion. Plant, Cell & Environment 44, 1329–1345.
| No gas source, no problem: proximity to pre-existing embolism and segmentation affect embolism spreading in angiosperm xylem by gas diffusion.Crossref | GoogleScholarGoogle Scholar |
Guan X, Werner J, Cao K-F, Pereira L, Kaack L, Mcadam SAM, Jansen S (2022) Stem and leaf xylem of angiosperm trees experiences minimal embolism in temperate forests during two consecutive summers with moderate drought. Plant Biology
| Stem and leaf xylem of angiosperm trees experiences minimal embolism in temperate forests during two consecutive summers with moderate drought.Crossref | GoogleScholarGoogle Scholar | 34990084PubMed |
Hammond WM, Yu K, Wilson LA, Will RE, Anderegg WRL, Adams HD (2019) Dead or dying? Quantifying the point of no return from hydraulic failure in drought-induced tree mortality. New Phytologist 223, 1834–1843.
| Dead or dying? Quantifying the point of no return from hydraulic failure in drought-induced tree mortality.Crossref | GoogleScholarGoogle Scholar | 31087656PubMed |
Harrison Day BL, Carins-Murphy MR, Brodribb TJ (2022) Reproductive water supply is prioritized during drought in tomato. Plant, Cell & Environment 45, 69–79.
| Reproductive water supply is prioritized during drought in tomato.Crossref | GoogleScholarGoogle Scholar |
Hietz P, Rosner S, Sorz J, Mayr S (2008) Comparison of methods to quantify loss of hydraulic conductivity in Norway spruce. Annals of Forest Science 65, 502
| Comparison of methods to quantify loss of hydraulic conductivity in Norway spruce.Crossref | GoogleScholarGoogle Scholar |
Hochberg U, Windt CW, Ponomarenko A, Zhang Y-J, Gersony J, Rockwell FE, Holbrook NM (2017) Stomatal closure, basal leaf embolism, and shedding protect the hydraulic integrity of grape stems. Plant Physiology 174, 764–775.
| Stomatal closure, basal leaf embolism, and shedding protect the hydraulic integrity of grape stems.Crossref | GoogleScholarGoogle Scholar | 28351909PubMed |
Hochberg U, Ponomarenko A, Zhang YJ, Rockwell FE, Holbrook NM (2019) Visualizing embolism propagation in gas-injected leaves. Plant Physiology 180, 874–881.
| Visualizing embolism propagation in gas-injected leaves.Crossref | GoogleScholarGoogle Scholar | 30842264PubMed |
Holbrook NM, Ahrens ET, Burns MJ, Zwieniecki MA (2001) In vivo observation of cavitation and embolism repair using magnetic resonance imaging. Plant Physiology 126, 27–31.
| In vivo observation of cavitation and embolism repair using magnetic resonance imaging.Crossref | GoogleScholarGoogle Scholar | 11351066PubMed |
Jacob V, Choat B, Churchill AC, Zhang H, Barton CVM, Krishnananthaselvan A, Post AK, Power SA, Medlyn BE, Tissue DT (2022) High safety margins to drought-induced hydraulic failure found in five pasture grasses. Plant Cell and Environment 45, 1631–1646.
| High safety margins to drought-induced hydraulic failure found in five pasture grasses.Crossref | GoogleScholarGoogle Scholar |
Jansen S, Schuldt B, Choat B (2015) Current controversies and challenges in applying plant hydraulic techniques. New Phytologist 205, 961–964.
| Current controversies and challenges in applying plant hydraulic techniques.Crossref | GoogleScholarGoogle Scholar | 25580652PubMed |
Johnson KM, Jordan GJ, Brodribb TJ (2018) Wheat leaves embolized by water stress do not recover function upon rewatering. Plant, Cell & Environment 41, 2704–2714.
| Wheat leaves embolized by water stress do not recover function upon rewatering.Crossref | GoogleScholarGoogle Scholar |
Johnson KM, Brodersen CR, Carins-Murphy MR, Choat B, Brodribb TJ (2020) Xylem embolism spreads by single-conduit events in three dry forest angiosperm stems. Plant Physiology 184, 212–222.
| Xylem embolism spreads by single-conduit events in three dry forest angiosperm stems.Crossref | GoogleScholarGoogle Scholar | 32581116PubMed |
Johnson KM, Lucani C, Brodribb TJ (2022) In vivo monitoring of drought-induced embolism in Callitris rhomboidea trees reveals wide variation in branchlet vulnerability and high resistance to tissue death. New Phytologist 233, 207–218.
| In vivo monitoring of drought-induced embolism in Callitris rhomboidea trees reveals wide variation in branchlet vulnerability and high resistance to tissue death.Crossref | GoogleScholarGoogle Scholar | 34625973PubMed |
Kaack L, Weber M, Isasa E, Karimi Z, Li S, Pereira L, Trabi CL, Zhang Y, Schenk HJ, Schuldt B, Schmidt V, Jansen S (2021) Pore constrictions in intervessel pit membranes provide a mechanistic explanation for xylem embolism resistance in angiosperms. New Phytologist 230, 1829–1843.
| Pore constrictions in intervessel pit membranes provide a mechanistic explanation for xylem embolism resistance in angiosperms.Crossref | GoogleScholarGoogle Scholar | 33595117PubMed |
Kim W (2013) Mechanics of xylem sap drinking. Biomedical Engineering Letters 3, 144–148.
| Mechanics of xylem sap drinking.Crossref | GoogleScholarGoogle Scholar |
Klepsch M, Zhang Y, Kotowska MM, Lamarque LJ, Nolf M, Schuldt B, Torres-Ruiz JM, Qin DW, Choat B, Delzon S, Scoffoni C, Cao KF, Jansen S (2018) Is xylem of angiosperm leaves less resistant to embolism than branches? Insights from microCT, hydraulics, and anatomy. Journal of Experimental Botany 69, 5611–5623.
| Is xylem of angiosperm leaves less resistant to embolism than branches? Insights from microCT, hydraulics, and anatomy.Crossref | GoogleScholarGoogle Scholar | 30184113PubMed |
Lamarque LJ, Corso D, Torres-ruiz JM, Badel E, Brodribb TJ, Burlett R, et al. (2018) An inconvenient truth about xylem resistance to embolism in the model species for refilling Laurus nobilis L. Annals of Forest Science 75, 88
| An inconvenient truth about xylem resistance to embolism in the model species for refilling Laurus nobilis L.Crossref | GoogleScholarGoogle Scholar |
Larter M, Brodribb TJ, Pfautsch S, Burlett R, Cochard H, Delzon S (2015) Extreme aridity pushes trees to their physical limits. Plant Physiology 168, 804–807.
| Extreme aridity pushes trees to their physical limits.Crossref | GoogleScholarGoogle Scholar | 26034263PubMed |
Levionnois S, Ziegler C, Jansen S, Calvet E, Coste S, Stahl C, Salmon C, Delzon S, Guichard C, Heuret P (2020) Vulnerability and hydraulic segmentations at the stem-leaf transition: coordination across Neotropical trees. New Phytologist 228, 512–524.
| Vulnerability and hydraulic segmentations at the stem-leaf transition: coordination across Neotropical trees.Crossref | GoogleScholarGoogle Scholar | 32496575PubMed |
Li X, Delzon S, Torres-Ruiz J, Badel E, Burlett R, Cochard H, Jansen S, King A, Lamarque LJ, Lenoir N, St-Paul NM, Choat B (2020a) Lack of vulnerability segmentation in four angiosperm tree species: evidence from direct X-ray microtomography observation. Annals of Forest Science 77, 37
| Lack of vulnerability segmentation in four angiosperm tree species: evidence from direct X-ray microtomography observation.Crossref | GoogleScholarGoogle Scholar |
Li X, Smith R, Choat B, Tissue DT (2020b) Drought resistance of cotton (Gossypium hirsutum) is promoted by early stomatal closure and leaf shedding. Functional Plant Biology 47, 91–98.
| Drought resistance of cotton (Gossypium hirsutum) is promoted by early stomatal closure and leaf shedding.Crossref | GoogleScholarGoogle Scholar | 31825787PubMed |
Losso A, Bär A, Dämon B, Dullin C, Ganthaler A, Petruzzellis F, Savi T, Tromba G, Nardini A, Mayr S, Beikircher B (2019) Insights from in vivo micro-CT analysis: testing the hydraulic vulnerability segmentation in Acer pseudoplatanus and Fagus sylvatica seedlings. New Phytologist 221, 1831–1842.
| Insights from in vivo micro-CT analysis: testing the hydraulic vulnerability segmentation in Acer pseudoplatanus and Fagus sylvatica seedlings.Crossref | GoogleScholarGoogle Scholar | 30347122PubMed |
Lucani CJ, Brodribb TJ, Jordan GJ, Mitchell PJ (2019) Juvenile and adult leaves of heteroblastic Eucalyptus globulus vary in xylem vulnerability. Trees 33, 1167–1178.
| Juvenile and adult leaves of heteroblastic Eucalyptus globulus vary in xylem vulnerability.Crossref | GoogleScholarGoogle Scholar |
Maherali H, Pockman WT, Jackson RB (2004) Adaptive variation in the vulnerability of woody plants to xylem cavitation. Ecology 85, 2184–2199.
| Adaptive variation in the vulnerability of woody plants to xylem cavitation.Crossref | GoogleScholarGoogle Scholar |
Martin-StPaul NK, Longepierre D, Huc R, Delzon S, Burlett R, Joffre R, Rambal S, Cochard H (2014) How reliable are methods to assess xylem vulnerability to cavitation? The issue of ‘open vessel’ artifact in oaks. Tree Physiology 34, 894–905.
| How reliable are methods to assess xylem vulnerability to cavitation? The issue of ‘open vessel’ artifact in oaks.Crossref | GoogleScholarGoogle Scholar | 25074860PubMed |
Moshelion M, Halperin O, Wallach R, Oren R, Way DA (2015) Role of aquaporins in determining transpiration and photosynthesis in water-stressed plants: crop water-use efficiency, growth and yield. Plant, Cell & Environment 38, 1785–1793.
| Role of aquaporins in determining transpiration and photosynthesis in water-stressed plants: crop water-use efficiency, growth and yield.Crossref | GoogleScholarGoogle Scholar |
Pereira L, Bittencourt PRL, Oliveira RS, Junior MBM, Barros FV, Ribeiro RV, Mazzafera P (2016) Plant pneumatics: stem air flow is related to embolism – new perspectives on methods in plant hydraulics. New Phytologist 211, 357–370.
| Plant pneumatics: stem air flow is related to embolism – new perspectives on methods in plant hydraulics.Crossref | GoogleScholarGoogle Scholar | 26918522PubMed |
Peters JMR, Gauthey A, Lopez R, Carins-Murphy MR, Brodribb TJ, Choat B (2020) Non-invasive imaging reveals convergence in root and stem vulnerability to cavitation across five tree species. Journal of Experimental Botany 71, 6623–6637.
| Non-invasive imaging reveals convergence in root and stem vulnerability to cavitation across five tree species.Crossref | GoogleScholarGoogle Scholar | 32822502PubMed |
Petruzzellis F, Tomasella M, Miotto A, Natale S, Trifilò P, Nardini A (2020) A leaf selfie: using a smartphone to quantify leaf vulnerability to hydraulic dysfunction. Plants 9, 234
| A leaf selfie: using a smartphone to quantify leaf vulnerability to hydraulic dysfunction.Crossref | GoogleScholarGoogle Scholar |
Pockman WT, Sperry JS, O’Leary JW (1995) Sustained and significant negative water pressure in xylem. Nature 378, 715–716.
| Sustained and significant negative water pressure in xylem.Crossref | GoogleScholarGoogle Scholar |
Pratt RB, Castro V, Fickle JC, Jacobsen AL (2020) Embolism resistance of different aged stems of a California oak species (Quercus douglasii): optical and microCT methods differ from the benchtop-dehydration standard. Tree Physiology 40, 5–18.
| Embolism resistance of different aged stems of a California oak species (Quercus douglasii): optical and microCT methods differ from the benchtop-dehydration standard.Crossref | GoogleScholarGoogle Scholar | 31553460PubMed |
Raven JA (2002) Selection pressures on stomatal evolution. New Phytologist 153, 371–386.
| Selection pressures on stomatal evolution.Crossref | GoogleScholarGoogle Scholar | 33863217PubMed |
Ritchie GA, Hinckley TM (1971) Evidence for error in pressure-bomb estimates of stem xylem potentials. Ecology 52, 534–536.
| Evidence for error in pressure-bomb estimates of stem xylem potentials.Crossref | GoogleScholarGoogle Scholar |
Rodriguez-Dominguez CM, Carins Murphy MR, Lucani C, Brodribb TJ (2018) Mapping xylem failure in disparate organs of whole plants reveals extreme resistance in olive roots. New Phytologist 218, 1025–1035.
| Mapping xylem failure in disparate organs of whole plants reveals extreme resistance in olive roots.Crossref | GoogleScholarGoogle Scholar | 29528498PubMed |
Sack L, Holbrook NM (2006) Leaf Hydraulics. Annual Review of Plant Biology 57, 361–381.
| Leaf Hydraulics.Crossref | GoogleScholarGoogle Scholar | 16669766PubMed |
Sade N, Shatil-Cohen A, Attia Z, Maurel C, Boursiac Y, Kelly G, Granot D, Yaaran A, Lerner S, Moshelion M (2014) The role of plasma membrane aquaporins in regulating the bundle sheath-mesophyll continuum and leaf hydraulics. Plant Physiology 166, 1609–1620.
| The role of plasma membrane aquaporins in regulating the bundle sheath-mesophyll continuum and leaf hydraulics.Crossref | GoogleScholarGoogle Scholar | 25266632PubMed |
Savi T, Miotto A, Petruzzellis F, Losso A, Pacilè S, Tromba G, Mayr S, Nardini A (2017) Drought-induced embolism in stems of sunflower: a comparison of in vivo micro-CT observations and destructive hydraulic measurements. Plant Physiology and Biochemistry 120, 24–29.
| Drought-induced embolism in stems of sunflower: a comparison of in vivo micro-CT observations and destructive hydraulic measurements.Crossref | GoogleScholarGoogle Scholar | 28968592PubMed |
Scoffoni C, Vuong C, Diep S, Cochard H, Sack L (2014) Leaf shrinkage with dehydration: coordination with hydraulic vulnerability and drought tolerance. Plant Physiology 164, 1772–1788.
| Leaf shrinkage with dehydration: coordination with hydraulic vulnerability and drought tolerance.Crossref | GoogleScholarGoogle Scholar | 24306532PubMed |
Scoffoni C, Albuquerque C, Brodersen CR, Townes SV, John GP, Bartlett MK, Buckley TN, McElrone AJ, Sack L (2017) Outside-xylem vulnerability, not xylem embolism, controls leaf hydraulic decline during dehydration. Plant Physiology 173, 1197–1210.
| Outside-xylem vulnerability, not xylem embolism, controls leaf hydraulic decline during dehydration.Crossref | GoogleScholarGoogle Scholar | 28049739PubMed |
Sergent AS, Varela SA, Barigah TS, Badel E, Cochard H, Dalla-Salda G, Delzon S, Fernández ME, Guillemot J, Gyenge J, Lamarque LJ, Martinez-Meier A, Rozenberg P, Torres-Ruiz JM, Martin-StPaul NK (2020) A comparison of five methods to assess embolism resistance in trees. Forest Ecology and Management 468, 118175
| A comparison of five methods to assess embolism resistance in trees.Crossref | GoogleScholarGoogle Scholar |
Skelton RP, Brodribb TJ, Choat B (2017a) Casting light on xylem vulnerability in an herbaceous species reveals a lack of segmentation. New Phytologist 214, 561–569.
| Casting light on xylem vulnerability in an herbaceous species reveals a lack of segmentation.Crossref | GoogleScholarGoogle Scholar | 28124474PubMed |
Skelton RP, Brodribb TJ, McAdam SAM, Mitchell PJ (2017b) Gas exchange recovery following natural drought is rapid unless limited by loss of leaf hydraulic conductance: evidence from an evergreen woodland. New Phytologist 215, 1399–1412.
| Gas exchange recovery following natural drought is rapid unless limited by loss of leaf hydraulic conductance: evidence from an evergreen woodland.Crossref | GoogleScholarGoogle Scholar | 28620915PubMed |
Skelton RP, Dawson TE, Thompson SE, Shen Y, Weitz AP, Ackerly D (2018) Low vulnerability to xylem embolism in leaves and stems of North American oaks. Plant Physiology 177, 1066–1077.
| Low vulnerability to xylem embolism in leaves and stems of North American oaks.Crossref | GoogleScholarGoogle Scholar | 29789436PubMed |
Skelton RP, Anderegg LDL, Diaz J, Kling MM, Papper P, Lamarque LJ, Delzon S, Dawson TE, Ackerly DD (2021) Evolutionary relationships between drought-related traits and climate shape large hydraulic safety margins in western North American oaks. Proceedings of the National Academy of Sciences of the United States of America 118, e2008987118
| Evolutionary relationships between drought-related traits and climate shape large hydraulic safety margins in western North American oaks.Crossref | GoogleScholarGoogle Scholar | 33649205PubMed |
Smith-Martin CM, Skelton RP, Johnson KM, Lucani C, Brodribb TJ (2020) Lack of vulnerability segmentation among woody species in a diverse dry sclerophyll woodland community. Functional Ecology 34, 777–787.
| Lack of vulnerability segmentation among woody species in a diverse dry sclerophyll woodland community.Crossref | GoogleScholarGoogle Scholar |
Sperry JS, Tyree MT (1988) Mechanism of water stress-induced xylem embolism. Plant Physiology 88, 581–587.
| Mechanism of water stress-induced xylem embolism.Crossref | GoogleScholarGoogle Scholar | 16666352PubMed |
Sperry JS, Donnelly JR, Tyree MT (1988) A method for measuring hydraulic conductivity and embolism in xylem. Plant, Cell & Environment 11, 35–40.
| A method for measuring hydraulic conductivity and embolism in xylem.Crossref | GoogleScholarGoogle Scholar |
Stiller V (2009) Soil salinity and drought alter wood density and vulnerability to xylem cavitation of baldcypress (Taxodium distichum (L.) Rich.) seedlings. Environmental and Experimental Botany 67, 164–171.
| Soil salinity and drought alter wood density and vulnerability to xylem cavitation of baldcypress (Taxodium distichum (L.) Rich.) seedlings.Crossref | GoogleScholarGoogle Scholar |
Stout DL, Sala A (2003) Xylem vulnerability to cavitation in Pseudotsuga menziesii and Pinus ponderosa from contrasting habitats. Tree Physiology 23, 43–50.
| Xylem vulnerability to cavitation in Pseudotsuga menziesii and Pinus ponderosa from contrasting habitats.Crossref | GoogleScholarGoogle Scholar |
Suissa JS, Friedman WE (2021) From cells to stems: the effects of primary vascular construction on drought-induced embolism in fern rhizomes. New Phytologist 232, 2238–2253.
| From cells to stems: the effects of primary vascular construction on drought-induced embolism in fern rhizomes.Crossref | GoogleScholarGoogle Scholar | 34273190PubMed |
Torres-Ruiz JM, Cochard H, Mayr S, Beikircher B, Diaz-Espejo A, Rodriguez-Dominguez CM, Badel E, Fernández JE (2014) Vulnerability to cavitation in Olea europaea current-year shoots: further evidence of an open-vessel artifact associated with centrifuge and air-injection techniques. Physiologia Plantarum 152, 465–474.
| Vulnerability to cavitation in Olea europaea current-year shoots: further evidence of an open-vessel artifact associated with centrifuge and air-injection techniques.Crossref | GoogleScholarGoogle Scholar | 24611594PubMed |
Torres-Ruiz JM, Cochard H, Mencuccini M, Delzon S, Badel E (2016) Direct observation and modelling of embolism spread between xylem conduits: a case study in Scots pine. Plant, Cell & Environment 39, 2774–2785.
| Direct observation and modelling of embolism spread between xylem conduits: a case study in Scots pine.Crossref | GoogleScholarGoogle Scholar |
Torres-Ruiz JM, Cochard H, Choat B, Jansen S, López R, Tomášková I, Padilla-Díaz CM, Badel E, Burlett R, King A, Lenoir N, Martin-StPaul NK, Delzon S (2017) Xylem resistance to embolism: presenting a simple diagnostic test for the open vessel artefact. New Phytologist 215, 489–499.
| Xylem resistance to embolism: presenting a simple diagnostic test for the open vessel artefact.Crossref | GoogleScholarGoogle Scholar | 28467616PubMed |
Tyree MT, Ewers FW (1991) The hydraulic architecture of trees and other woody plants. New Phytologist 119, 345–360.
| The hydraulic architecture of trees and other woody plants.Crossref | GoogleScholarGoogle Scholar |
Tyree MT, Sperry JS (1989a) Characterization and propagation of acoustic emission signals in woody plants: towards an improved acoustic emission counter. Plant, Cell & Environment 12, 371–382.
| Characterization and propagation of acoustic emission signals in woody plants: towards an improved acoustic emission counter.Crossref | GoogleScholarGoogle Scholar |
Tyree MT, Sperry JS (1989b) Vulnerability of xylem to cavitation and embolism. Annual Review of Plant Physiology and Plant Molecular Biology 40, 19–36.
| Vulnerability of xylem to cavitation and embolism.Crossref | GoogleScholarGoogle Scholar |
Venturas MD, Pratt RB, Jacobsen AL, Castro V, Fickle JC, Hacke UG (2019) Direct comparison of four methods to construct xylem vulnerability curves: differences among techniques are linked to vessel network characteristics. Plant, Cell & Environment 42, 2422–2436.
| Direct comparison of four methods to construct xylem vulnerability curves: differences among techniques are linked to vessel network characteristics.Crossref | GoogleScholarGoogle Scholar |
Wagner Y, Feng F, Yakir D, Klein T, Hochberg U (2022) In situ, direct observation of seasonal embolism dynamics in Aleppo pine trees growing on the dry edge of their distribution.
| In situ, direct observation of seasonal embolism dynamics in Aleppo pine trees growing on the dry edge of their distribution.Crossref | GoogleScholarGoogle Scholar | 35514143PubMed |
Xiong D, Nadal M (2020) Linking water relations and hydraulics with photosynthesis. The Plant Journal 101, 800–815.
| Linking water relations and hydraulics with photosynthesis.Crossref | GoogleScholarGoogle Scholar | 31677190PubMed |
Yao GQ, Nie ZF, Turner NC, Li FM, Gao TP, Fang XW, Scoffoni C (2021) Combined high leaf hydraulic safety and efficiency provides drought tolerance in Caragana species adapted to low mean annual precipitation. New Phytologist 229, 230–244.
| Combined high leaf hydraulic safety and efficiency provides drought tolerance in Caragana species adapted to low mean annual precipitation.Crossref | GoogleScholarGoogle Scholar | 32749703PubMed |
Zhang F-P, Brodribb TJ (2017) Are flowers vulnerable to xylem cavitation during drought? Proceedings of the Royal Society B: Biological Sciences 284, 20162642
| Are flowers vulnerable to xylem cavitation during drought?Crossref | GoogleScholarGoogle Scholar | 28469026PubMed |
Zhang YJ, Rockwell FE, Wheeler JK, Holbrook NM (2014) Reversible deformation of transfusion tracheids in Taxus baccata is associated with a reversible decrease in leaf hydraulic conductance. Plant Physiology 165, 1557–1565.
| Reversible deformation of transfusion tracheids in Taxus baccata is associated with a reversible decrease in leaf hydraulic conductance.Crossref | GoogleScholarGoogle Scholar | 24948828PubMed |