

Constitutive and facultative crassulacean acid metabolism (CAM) in Cuban oregano, Coleus amboinicus (Lamiaceae)
Klaus Winter A C , Aurelio Virgo A , Milton Garcia A , Jorge Aranda A and Joseph A. M. Holtum A B
A C , Aurelio Virgo A , Milton Garcia A , Jorge Aranda A and Joseph A. M. Holtum A B
A Smithsonian Tropical Research Institute, PO Box 0843-03092, Balboa, Ancón, Republic of Panama.
B College of Science and Engineering, James Cook University, Townsville, Qld 4811, Australia.
C Corresponding author. Email: winterk@si.edu
Functional Plant Biology 48(7) 647-654 https://doi.org/10.1071/FP20127
Submitted: 1 May 2020 Accepted: 23 July 2020 Published: 14 September 2020
Journal Compilation © CSIRO 2021 Open Access CC BY-NC-ND
Abstract
Plants exhibiting the water-conserving crassulacean acid metabolism (CAM) photosynthetic pathway provide some of the most intriguing examples of photosynthetic diversity and plasticity. Here, a largely unnoticed facet of CAM-plant photosynthesis is highlighted: the co-occurrence of ontogenetically controlled constitutive and environmentally controlled facultative CAM in a species. Both forms of CAM are displayed in leaves of Coleus amboinicus Lour. (Lamiaceae), a semi-succulent perennial plant with oregano-like flavour that is native to southern and eastern Africa and naturalised elsewhere in the tropics. Under well-watered conditions, leaves assimilate CO2 predominantly by the C3 pathway. They also display low levels of CO2 uptake at night accompanied by small nocturnal increases in leaf tissue acidity. This indicates the presence of weakly expressed constitutive CAM. CAM expression is strongly enhanced in response to drought stress. The drought-enhanced component of CAM is reversible upon rewatering and thus considered to be facultative. In contrast to C. amboinicus, the thin-leaved closely related Coleus scutellarioides (L.) Benth. exhibits net CO2 fixation solely in the light via the C3 pathway, both under well-watered and drought conditions. However, low levels of nocturnal acidification detected in leaves and stems indicate that the CAM cycle is present. The highly speciose mint family, which contains few known CAM-exhibiting species and is composed predominantly of C3 species, appears to be an excellent group of plants for studying the evolutionary origins of CAM and for determining the position of facultative CAM along the C3–full CAM trajectory.
Additional keywords: CO2 fixation, CAM evolution, drought stress, leaf acidity, photosynthesis.
Introduction
Coleus amboinicus Lour. (Lamiaceae) (previously Plectranthus amboinicus (Lour.) Spreng.; Paton et al. 2019) is one of the most widely recognised members of the genus Coleus. It is a semi-succulent perennial herb native to southern and eastern Africa that is widely cultivated and naturalised elsewhere in the tropics and subtropics. Its leaves are highly aromatic, have many traditional medical uses (Lukhoba et al. 2006; Arumugam et al. 2016), and serve as a substitute for oregano (Origanum vulgare L.) to flavour food. Common names of C. amboinicus include Mexican mint and Cuban oregano. Plectranthinae is the only subtribe in the mint family in which species with the water-conserving CAM photosynthetic pathway of photosynthesis have been identified (Kluge and Ting 1978; Smith and Winter 1996). Conclusive photosynthetic pathway information for C. amboinicus is not available.
CAM photosynthesis is believed to occur in well over 5% of angiosperm species, and is also found in ferns, fern allies and gymnosperms (Smith and Winter 1996). In most CAM-exhibiting species, the expression of CAM is constitutive (or obligate), i.e. the CAM pathway is always present in mature photosynthetic tissues (Winter et al. 2015; Winter 2019). The degree to which plants engage in constitutive CAM relative to C3 photosynthesis is species-specific and modulated by environment. There is a small, yet steadily increasing, number of species in which CAM can be shown to be facultative: CAM is elicited in response to environmental stress, typically drought, and is downregulated once the stress ceases (Winter and Holtum 2007, 2014). Under well-watered conditions, facultative CAM plants acquire carbon mostly via C3 photosynthesis or, in some cases, C4 photosynthesis (Holtum et al. 2017; Winter and Holtum 2017).
Building on observations by Kluge and Ting (1978), who reported nocturnal acidification in Plectranthus prostratus Gürke (now Coleus prostratus (Gürke) A.J.Paton; Paton et al. 2019), Herppich and collaborators studied the occurrence of CAM in several species of what was then considered Plectranthus native to Yemen and southern Africa (Herppich 1996, 1997; Herppich and Herppich 1996; Herppich et al. 1998). Features of CAM were noted in four of seven species. CAM expression ranged from very low levels in a species with predominantly C3 photosynthetic CO2 fixation in the light, to relatively high levels in Plectranthus marrubioides (now Coleus cylindraceus (Hochst. ex Benth.) A.J.Paton; Paton et al. 2019) where nocturnal CO2 fixation contributed >50% to total daily carbon gain in well-watered plants.
More recently, Ramana and Chaitanya (2015) reported CAM-type nocturnal increases in tissue acidity in several species of Coleus including Coleus aromaticus, a synonym of Coleus amboinicus, and Coleus blumei, a synonym of Coleus scutellarioides (Paton et al. 2019). Well-watered plants of these species showed significant overnight accumulation of total organic acids, a result that would be consistent with obligate CAM. However, nocturnal increases in H+ were considerably larger than nocturnal increases in malic acid and citric acid combined, raising questions about the nature of the organic acid anion involved in CAM in these plants. Net CO2 exchange was not determined by these authors, nor did they study the effects of drought stress on photosynthetic pathway physiology.
Facultative CAM sensu stricto, i.e. the reversible induction or upregulation of CAM in response to soil water-deficit stress, has never been explicitly demonstrated in any member of the Lamiaceae, although a close inspection of published data for P. marrubioides, classified as an obligate CAM plant by Herppich et al. (1998), does reveal evidence of drought-enhanced nocturnal CO2 fixation. However, drought-enhanced nocturnal malate accumulation was not observed in P. marrubioides (fig. 4A, B in Herppich et al. 1998).
In the study presented here, the possible presence of CAM, in particular facultative CAM, was explored in leaves of C. amboinicus. Measurements of day-night CO2 exchange and of nocturnal changes in tissue acidity demonstrated that this species has the ability to display both strong facultative CAM and weakly expressed constitutive CAM. CO2 exchange responses of C. amboinicus to drought stress were compared with those of C. scutellarioides, a thin-leaved species that exhibits net CO2 fixation solely in the light. Our original hypothesis that C. scutellarioides is exclusively C3 was, however, proven incorrect during the course of the study.
Materials and methods
Growth of plants
Coleus amboinicus Lour. was grown from 5 cm stem cuttings in 1.6 L, 23 cm high TP49 tree pots (Stuewe & Sons) filled with Miracle-Gro potting mix (Miracle-Gro Lawn Products). Cuttings were obtained from plants purchased at Riba-Smith supermarket in Panama City, Republic of Panama. Plants were kept underneath a rain shelter and received ~30% of daily full solar radiation. They were 20–30 cm tall when used for gas-exchange and nocturnal acidification measurements. Coleus scutellarioides (L.) Benth. was cultivated from seeds of plants collected in a local garden in Panama City. Pot volume was 1.0 L. Soil and growing conditions were as for C. amboinicus.
Coleus amboinicus was also grown under natural sunlight and rainfall conditions in a raised garden box at the Smithsonian Tropical Research Institute’s Santa Cruz Experimental Research Facility in Gamboa, Republic of Panama, from October 2016 to April 2017 (Fig. 1). The dimensions of the raised garden box were 1.5 × 1.5 × 0.3 m. Its frame was made of 3-cm thick wood panels. Approximately 50 small plants that had been established from cuttings were inserted in forest top soil at the end of September 2016.

|
Titratable acidity
Leaf discs were excised with a cork borer at dusk and dawn (four 1.5 cm diameter disks per sample) and, after fresh mass (FM) determination, were frozen in liquid nitrogen. In one experiment with C. scutellarioides, stem samples (1.5–3.0 g FM) were also taken. Samples were freeze-dried (Freezone 4.5, Labconco) for dry mass determination, and boiled in 60 mL of 50% (v/v) ethanol for 10 min. The volume was brought back to 60 mL with water, and samples were boiled again for 10 min. After cooling to room temperature, samples were titrated with 5 mM KOH to pH 6.5.
Net CO2 exchange
In the experiments depicted in the ‘Results’, the major portion of mature, attached leaves was enclosed into a clamp-on PMK 10 gas-exchange cuvette (Walz GmbH) connected to a through-flow (open) gas-exchange system consisting of Walz components and a LI-6262 CO2 analyser (LI-COR Biosciences). Air containing 400 µmol mol–1 CO2 was delivered to the leaf cuvette at a flow rate of 1.26 L min–1. Illumination was by a SS-GU300-w LED light (Sunshine Systems). Leaf cuvette and plant were placed inside a temperature-regulated controlled-environment chamber (GC8-T, EGC).
Gas exchange was also measured on entire, attached leaves using a GWK 3M chamber (Walz) (data not shown). In total, leaves of five different plants of C. amboinicus and leaves of two plants of C. scutellarioides were studied. Results from replicate experiments were consistent. Representative gas-exchange experiments are depicted in the ‘Results’.
Results
In well-watered plants of P. amboinicus, light-driven C3 photosynthesis was the principal carbon acquisition pathway (Fig. 2.), but the diel pattern of CO2 exchange deviated significantly from what one might expect for a typical C3 species. Net CO2 uptake exhibited a temporary dip during the light period. Nocturnal respiratory CO2 loss, rather than staying constant, gradually decreased over the course of the dark period, with CO2 exchange transitioning to very small rates of net CO2 uptake towards the end of the dark period. In response to drought stress, net CO2 fixation in the light dropped to essentially zero on Days 6 and 7 (except for a short peak immediately after the onset of the light period), whereas CO2 uptake in the dark was rapidly and markedly upregulated. After only 3 days of withholding irrigation, net CO2 uptake was observed throughout almost the entire dark period. Upon rewatering, CO2 fixation in the light swiftly recovered and nocturnal CO2 fixation gradually declined. On Day 11, 5 days after rewatering, the nocturnal CO2 balance was similar to that observed for the well-watered plant on Day 1. CO2 exchange was negative during the first two-thirds of the night but was positive at the end of the night.
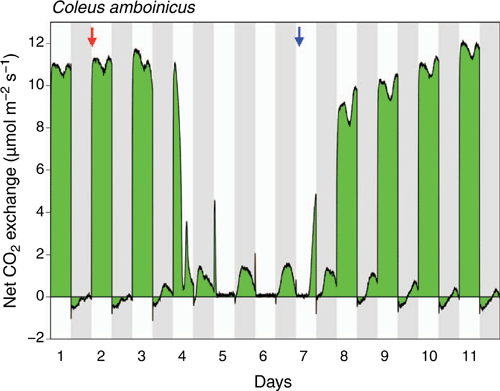
|
Consistent with leaf gas exchange, well-watered plants of C. amboinicus showed small but significant nocturnal increases in leaf-tissue acidity (Fig. 3, treatment A). Nocturnal H+ increase (ΔH+) rose 5- to 6-fold in response to drought stress (Fig. 3, treatments B and D). Upon rewatering, the component of ΔH+ elevated in response to drought stress reversed fully (treatment C) or almost fully (treatment E).
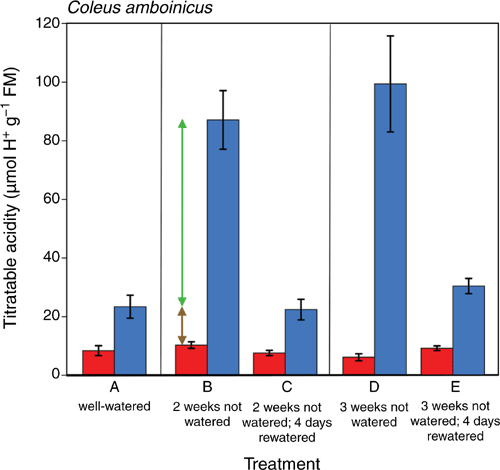
|

|
In well-watered plants, the basal, permanently expressed ΔH+ increased with leaf age, from 4 µmol g–1 FM in the youngest leaf (leaf 1) to 17 µmol g–1 FM in the oldest leaf (leaf 6) (Fig. 4). The drought stress-triggered increase of ΔH+ also responded to leaf age in that the ΔH+ levels gradually increased from leaves 1 to 3 and were greatest (up to 115 µmol g–1 FM) in fully expanded mature leaves (leaves 4 to 6).
The responses of plants grown outdoors in a raised garden box from November 2016 to May 2017 throughout a wet and a dry season corroborated those of potted plants (Fig. 5). Nocturnal acidification was present but low at the end of the wet season (November and December 2016). The ΔH+ levels, when expressed on the bases of both leaf fresh mass and area, increased markedly with the onset of the dry season in January and February, reaching peak values during March and April, before tailing off in April as the new wet season began.
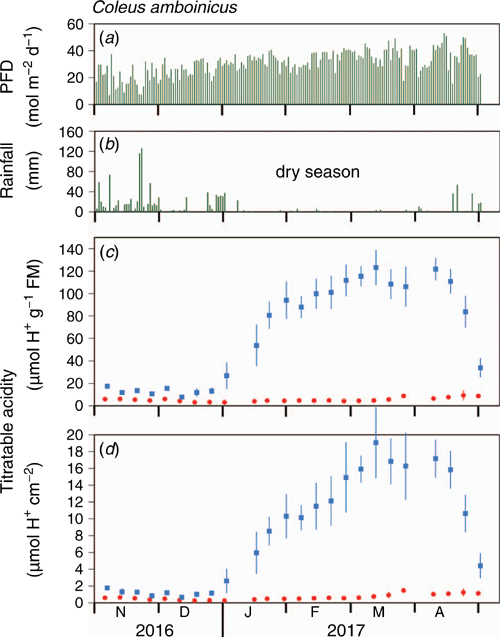
|
In contrast to C. amboinicus, the diel net CO2 exchange pattern of leaves of well-watered C. scutellarioides was outwardly consistent with a classic C3 photosynthetic phenotype. During the entire wet–dry–wet cycle shown in Fig. 6, net CO2 uptake was restricted to the 12-h light periods. In the initial well-watered state, rates of CO2 uptake were relatively constant at around 11 µmol m–2 s–1. Net CO2 loss during the dark period was also relatively constant although the rate decreased during the middle of the night, giving the overall nocturnal trace a bowed appearance. Exposure to drought stress led to a decline of CO2 gain during the light and a reduction of CO2 loss during the dark. The curved nature of the nocturnal trace of CO2 exchange became more pronounced. Upon rewatering, the diel pattern of CO2 exchange reverted to the pattern observed for the well-watered plant on Day 1.
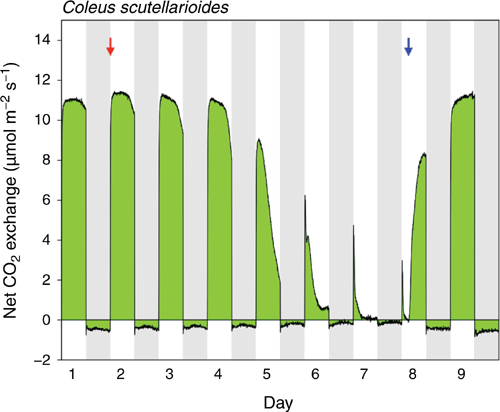
|
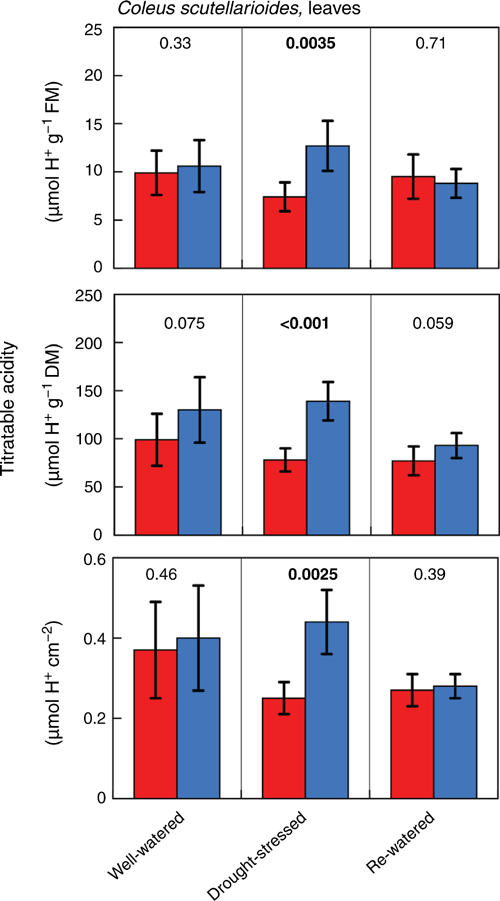
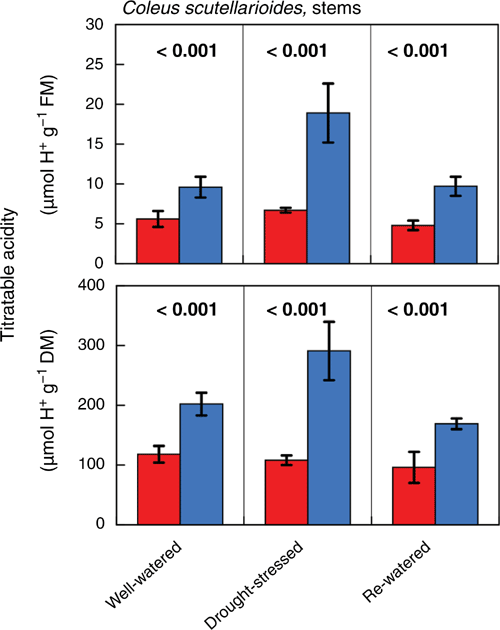
The subtle difference in the curved shape of the nocturnal gas-exchange observed for C. scutellarioides from a conventional level C3-type trace can be an indicator of the presence of low-level CAM. This possibility was confirmed. Leaves of C. scutellarioides exhibited small nocturnal increases in titratable acidity in water-stressed plants but not in well-watered plants. The differences were discernible irrespective of whether acidification was expressed on a fresh mass, dry mass or leaf area basis (Fig. 7). Stem tissues of C. scutellarioides also acidified at night. The pattern of acidification in stems differed from leaves in that acidification was present in the stems of well-watered, drought-stressed and rewatered plants (Fig. 8). The level of stem acidification increased when plants were drought-stressed and decreased to levels present in well-watered plants when droughted plants were rewatered.
|
|
Discussion
Although C3 photosynthesis is the principal carbon acquisition pathway in C. amboinicus, CAM photosynthesis, with both constitutive and facultative features, is clearly present in this species. Whereas leaves always show a background level of weakly expressed constitutive CAM, they also have the ability to strongly upregulate CAM in a facultative manner in response to drought stress. Compared with well-watered plants, nocturnal accumulation of H+ was 5- to 10-fold greater in mature leaves of drought-stressed plants. In the fully CAM-induced state, nocturnal CO2 assimilation reached 1.5 µmol m–2 s–1, which is a relatively high rate in comparison to many other facultative CAM species, equivalent to approximately 15% of rates of C3-photosynthetic CO2 assimilation of unstressed C. amboinicus in the light. Thus, C. amboinicus joins the subset of CAM plants that have relatively high facultative CAM-expression, such as Clusia pratensis and Mesembryanthemum crystallinum (Winter 1973; Winter and Holtum 2014). In many other facultative CAM plants, nocturnal carbon gain in the fully-induced CAM state is typically less than 5% of the diurnal C3 (or C4) photosynthetic carbon gain (Winter 2019); species in the Basellaceae, Portulacaceae and Talinaceae belong to this category (Winter and Holtum 2014, 2017; Holtum et al. 2018).
It is well established that in species with constitutive CAM, CAM activity increases as photosynthetic tissues mature (Jones 1975; Winter 2019). Consistent with these previous observations, background levels of weakly expressed CAM in C. amboinicus increased as leaves aged. In the same way, the magnitude of the facultative induction of CAM-type nocturnal acidification increased with leaf age, indicating that facultative CAM, although by definition a strictly environmentally-triggered phenomenon, is still subject to similar ontogenetic constraints as constitutive CAM (Winter 1973; Borland et al. 1998).
As more species with facultative CAM are discovered, it is possible that, like C. amboinicus, many of these will display the trait in the form of drought-enhanced CAM upregulation on top of pre-existing weakly-expressed constitutive CAM, rather than starting with a relatively clean non-CAM phenotype, as demonstrated for M. crystallinum, some Calandrinia spp. and Portulaca spp. (Winter and Holtum 2007, 2011, 2014). In fact, the extensively studied facultative CAM species Clusia minor has long been known to exhibit low-level CAM even when well-watered (Borland et al. 1998; Lüttge 2006). Future species comparisons will show whether there is correlation between the degree of CAM expression in the fully drought-induced state and the absence or presence of weakly expressed CAM in the well-watered state.
The adaptive significance of CAM in C. amboinicus in its natural habitats is currently unclear. To learn more about the ecology of CAM in C. amboinicus, in situ monitoring of seasonal changes of C3 and CAM activities, growth, and reproduction is required. In Africa, these plants are found at low altitudes in woodland or coastal bush, on rocky slopes, and loamy or sandy flats (Datiles and Acevedo-Rodríguez 2014). In Panama and elsewhere, plants are typically cultivated under semi-shaded conditions. In the present study, drought-stressed plants maintained at 30% of natural sunlight accumulated similar amounts of H+ overnight as drought-stressed plants outdoors under full sunlight conditions (Figs 3, 4, 5), consistent with a preference of C. amboinicus for partial shade.
Facultative CAM species operating in either the C3 (C4 in the case of Portulaca) or CAM mode are excellent study systems for identifying the molecular and biochemical underpinnings of the CAM pathway (Winter and Holtum 2014; Brilhaus et al. 2016; Ferrari et al. 2020). Facultative CAM species can also significantly contribute to our understanding of CAM evolution, as facultative CAM may be either an evolutionary ‘end point’ or an intermediate state along the evolutionary trajectory from the ancestral C3 pathway to full, strongly expressed CAM. Furthermore, species like C. amboinicus raise the interesting question of whether facultative CAM is an addition to weakly expressed permanent CAM or vice versa. Either way, for elucidating how species transitioned from C3 to CAM, obtaining a clean C3 control species within a particular lineage for comparative purposes can be a major obstacle, especially when the lineage under investigation is rich in CAM species (e.g. Heyduk et al. 2018, 2019). Agavoideae, Cactaceae and Orchidaceae are examples of such lineages. In the Lamiaceae, the sixth largest angiosperm family with over 7000 species, most species are C3 plants and thus far CAM is only known in the genus Coleus as all species with CAM previously described as Plectranthus can now be considered Coleus. The preponderance of the C3 pathway should greatly facilitate the identification of a suitable, closely related C3 control species in the Lamiaceae, making it an attractive family for future CAM evolution studies, especially since it is well described taxonomically and phylogenetically (Paton et al. 2004, 2018, 2019; Suddee et al. 2004; Li et al. 2016).
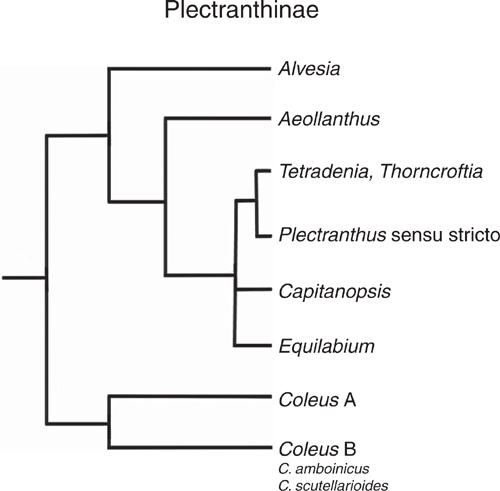
Over the last 30 years, Coleus was merged into Plectranthus by most authors, but a recent phylogenetic study of the monophyletic subtribe Plectranthinae (Paton et al. 2018, 2019; building upon Paton et al. 2004) recommends generic recognition of Coleus and distinguishes two sister groups: (i) the Plectranthus clade that contains Plectranthus sensu stricto and the genera Alvesia, Aeollanthus, Capitanopsis, Equilabium, Tetradenia and Thorncroftia, and (ii) the Coleus clade that consists of two clades, one of which diversified from ~14.4 million years ago and contains C. amboinicus and C. scutellarioides, two species in which the magnitude of CAM-expression greatly differs (Fig. 9).
|
Studies of two closely related Erycina species (Orchidaceae), one exhibiting CAM and one ostensibly exhibiting C3, indicated similar expression patterns of key CAM pathway genes suggesting that early steps of the transitioning to CAM occurred in the ancestor of both species (Heyduk et al. 2019). In fact, during the early phase of this study we considered using C. scutellarioides as a C3 control species until we discovered that the 24 h gas exchange deviated slightly from a strict C3-type pattern and that a small degree of nocturnal acidification was measurable, especially in stems. This experience highlights that informed choices of species for studies of CAM evolution require not just robust phylogenies but also detailed physiological measurements on CAM presence and absence.
Conflicts of interest
The authors declare no conflicts of interest.
Acknowledgements
This research was supported by the Smithsonian Tropical Research Institute and by Australian Research Council grant DP160100098. Carmen Galdames identified Coleus scutellarioides.
References
Arumugam G, Kumara Swamy M, Sinniah UR (2016) Plectranthus amboinicus (Lour.) Spreng: botanical, phytochemical, pharmacological and nutritional significance. Molecules 21, 369| Plectranthus amboinicus (Lour.) Spreng: botanical, phytochemical, pharmacological and nutritional significance.Crossref | GoogleScholarGoogle Scholar | 27043511PubMed |
Borland AM, Técsi LI, Leegood RC, Walker RP (1998) Inducibility of crassulacean acid metabolism (CAM) in Clusia species; physiological/biochemical characterization and intercellular localization of carboxylation and decarboxylation processes in three species which exhibit different degrees of CAM. Planta 205, 342–351.
| Inducibility of crassulacean acid metabolism (CAM) in Clusia species; physiological/biochemical characterization and intercellular localization of carboxylation and decarboxylation processes in three species which exhibit different degrees of CAM.Crossref | GoogleScholarGoogle Scholar |
Brilhaus D, Bräutigam A, Mettler-Altmann T, Winter K, Weber A (2016) Reversible burst of transcriptional changes during induction of crassulacean acid metabolism (CAM) in Talinum triangulare. Plant Physiology 170, 102–122.
| Reversible burst of transcriptional changes during induction of crassulacean acid metabolism (CAM) in Talinum triangulare.Crossref | GoogleScholarGoogle Scholar | 26530316PubMed |
Datiles MJ, Acevedo-Rodríguez P (2014) Plectranthus amboinicus (Indian borage). Available at www.cabi.org/isc/datasheet/119834 [Verified 21 November 2018]
Ferrari RC, Bittencourt PP, Rodrigues MA, Moreno-Villena JJ, Alves FRR, Gastaldi VD, Boxall SF, Dever LV, Demarco D, Andrade SCS, Edwards EJ, Hartwell J, Freschi L (2020) C4 and crassulacean acid metabolism within a single leaf: deciphering key components behind a rare photosynthetic adaptation. New Phytologist 225, 1699–1714.
| C4 and crassulacean acid metabolism within a single leaf: deciphering key components behind a rare photosynthetic adaptation.Crossref | GoogleScholarGoogle Scholar | 31610019PubMed |
Herppich WB (1996) Photosynthesis but not CAM responded flexibly to changes in irradiance in Plectranthus marrubioides (Lamiaceae). Photosynthetica 34, 1–12.
| Photosynthesis but not CAM responded flexibly to changes in irradiance in Plectranthus marrubioides (Lamiaceae).Crossref | GoogleScholarGoogle Scholar |
Herppich WB (1997) Stomatal responses to changes in air humidity are not necessarily linked to nocturnal CO2 uptake in the CAM plant Plectranthus marrubioides Benth. (Lamiaceae). Plant, Cell & Environment 20, 393–399.
| Stomatal responses to changes in air humidity are not necessarily linked to nocturnal CO2 uptake in the CAM plant Plectranthus marrubioides Benth. (Lamiaceae).Crossref | GoogleScholarGoogle Scholar |
Herppich WB, Herppich M (1996) Ecophysiological investigations on plants of the genus Plectranthus (fam. Lamiaceae) native to Yemen and southern Africa. Flora 191, 401–408.
| Ecophysiological investigations on plants of the genus Plectranthus (fam. Lamiaceae) native to Yemen and southern Africa.Crossref | GoogleScholarGoogle Scholar |
Herppich WB, Herppich M, Von Willert DJ (1998) Ecophysiological investigations on plants of the genus Plectranthus (Lamiaceae). Influence of environment and leaf age on CAM, gas exchange and leaf water relations in Plectranthus marrubioides Benth. Flora 193, 99–109.
| Ecophysiological investigations on plants of the genus Plectranthus (Lamiaceae). Influence of environment and leaf age on CAM, gas exchange and leaf water relations in Plectranthus marrubioides Benth.Crossref | GoogleScholarGoogle Scholar |
Heyduk K, Ray JN, Ayyampalayam S, Leebens-Mack J (2018) Shifts in gene expression profiles are associated with weak and strong crassulacean acid metabolism. American Journal of Botany 105, 587–601.
| Shifts in gene expression profiles are associated with weak and strong crassulacean acid metabolism.Crossref | GoogleScholarGoogle Scholar | 29746718PubMed |
Heyduk K, Hwang M, Albert VA, Silvera K, Lan T, Farr KM, Chang T-H, Chan M-T, Winter K, Leebens-Mack J (2019) Altered gene regulatory networks are associated with the transition from C3 to crassulacean acid metabolism in Erycina (Oncidiinae: Orchidaceae). Frontiers in Plant Science 9, 2000
| Altered gene regulatory networks are associated with the transition from C3 to crassulacean acid metabolism in Erycina (Oncidiinae: Orchidaceae).Crossref | GoogleScholarGoogle Scholar | 30745906PubMed |
Holtum JAM, Hancock LP, Edwards EJ, Winter K (2017) Optional use of CAM photosynthesis in two C4 species, Portulaca cyclophylla and Portulaca digyna. Journal of Plant Physiology 214, 91–96.
| Optional use of CAM photosynthesis in two C4 species, Portulaca cyclophylla and Portulaca digyna.Crossref | GoogleScholarGoogle Scholar | 28511087PubMed |
Holtum JAM, Hancock LP, Edwards EJ, Winter K (2018) Crassulacean acid metabolism in the Basellaceae (Caryophyllales). Plant Biology 20, 409–414.
| Crassulacean acid metabolism in the Basellaceae (Caryophyllales).Crossref | GoogleScholarGoogle Scholar | 29369469PubMed |
Jones MB (1975) The effect of leaf age on leaf resistance and CO2 exchange of the CAM plant Bryophyllum fedtschenkoi. Planta 123, 91–96.
| The effect of leaf age on leaf resistance and CO2 exchange of the CAM plant Bryophyllum fedtschenkoi.Crossref | GoogleScholarGoogle Scholar | 24436027PubMed |
Kluge M, Ting IP (1978) ‘Crassulacean acid metabolism.’ (Springer-Verlag: Berlin, Germany)
Li B, Cantino PD, Olmstead RG, Bramley GLC, Xiang C-L, Ma Z-H, Tan Y-H, Zhang D-X (2016) A large-scale chloroplast phylogeny of the Lamiaceae sheds new light on its subfamilial classification. Scientific Reports 6, 34343
| A large-scale chloroplast phylogeny of the Lamiaceae sheds new light on its subfamilial classification.Crossref | GoogleScholarGoogle Scholar | 27748362PubMed |
Lukhoba CW, Simmonds MSJ, Paton AJ (2006) Plectranthus: a review of ethnobotanical uses. Journal of Ethnopharmacology 103, 1–24.
| Plectranthus: a review of ethnobotanical uses.Crossref | GoogleScholarGoogle Scholar | 16289602PubMed |
Lüttge U (2006) Photosynthetic flexibility and ecophysiological plasticity: questions and lessons from Clusia, the only CAM tree, in the neotropics. New Phytologist 171, 7–25.
| Photosynthetic flexibility and ecophysiological plasticity: questions and lessons from Clusia, the only CAM tree, in the neotropics.Crossref | GoogleScholarGoogle Scholar | 16771979PubMed |
Paton AJ, Springate D, Siuddee S, Otieno D, Grayer RJ, Harley MM, Willis F, Simmonds MSJ, Powell MP, Savolainen V (2004) Phylogeny and evolution of basils and allies (Ocimeae, Labiatae) based on three plastid DNA regions. Molecular Phylogenetics and Evolution 31, 277–299.
| Phylogeny and evolution of basils and allies (Ocimeae, Labiatae) based on three plastid DNA regions.Crossref | GoogleScholarGoogle Scholar | 15019625PubMed |
Paton A, Mwanyambo M, Culham A (2018) Phylogenetic study of Plectranthus, Coleus and allies (Lamiaceae): taxonomy, distribution and medicinal use. Botanical Journal of the Linnean Society 188, 355–376.
| Phylogenetic study of Plectranthus, Coleus and allies (Lamiaceae): taxonomy, distribution and medicinal use.Crossref | GoogleScholarGoogle Scholar |
Paton AJ, Mwanyambo M, Govaerts RHA, Smitha K, Suddee S, Phillipson PB, Wilson TC, Forster PI, Culham A (2019) Nomenclatural changes in Coleus and Plectranthus (Lamiaceae): a tale of more than two genera. PhytoKeys 129, 1–158.
| Nomenclatural changes in Coleus and Plectranthus (Lamiaceae): a tale of more than two genera.Crossref | GoogleScholarGoogle Scholar | 31523157PubMed |
Ramana GV, Chaitanya KV (2015) Variations in δ13C rates and crassulacean acid metabolism in six Coleus species. British Journal of Applied Science and Technology 6, 295–303.
| Variations in δ13C rates and crassulacean acid metabolism in six Coleus species.Crossref | GoogleScholarGoogle Scholar |
Smith JAC, Winter K (1996) Taxonomic distribution of crassulacean acid metabolism. In ‘Crassulacean acid metabolism’. (Eds K Winter, JAC Smith) pp. 427–436. (Springer-Verlag: Berlin, Germany)
Suddee S, Paton AJ, Parnell JAN (2004) A taxonomic revision of tribe Ocimeae Dumort. (Lamiaceae) in continental South East Asia II. Plectranthinae. Kew Bulletin 59, 379–414.
| A taxonomic revision of tribe Ocimeae Dumort. (Lamiaceae) in continental South East Asia II. Plectranthinae.Crossref | GoogleScholarGoogle Scholar |
Winter K (1973) CO2-Fixierungsreaktionen bei der Salzpflanze Mesembryanthemum crystallinum unter variierten Außenbedingungen. Planta 114, 75–85.
| CO2-Fixierungsreaktionen bei der Salzpflanze Mesembryanthemum crystallinum unter variierten Außenbedingungen.Crossref | GoogleScholarGoogle Scholar | 24458666PubMed |
Winter K (2019) Ecophysiology of constitutive and facultative CAM photosynthesis. Journal of Experimental Botany 70, 6495–6508.
| Ecophysiology of constitutive and facultative CAM photosynthesis.Crossref | GoogleScholarGoogle Scholar | 30810162PubMed |
Winter K, Holtum JAM (2007) Environment or development? Lifetime net CO2 exchange and control of the expression of crassulacean acid metabolism in Mesembryanthemum crystallinum. Plant Physiology 143, 98–107.
| Environment or development? Lifetime net CO2 exchange and control of the expression of crassulacean acid metabolism in Mesembryanthemum crystallinum.Crossref | GoogleScholarGoogle Scholar | 17056756PubMed |
Winter K, Holtum JAM (2011) Induction and reversal of CAM photosynthesis in Calandrinia polyandra Benth: effects of soil moisture and nutrients. Functional Plant Biology 38, 576–582.
| Induction and reversal of CAM photosynthesis in Calandrinia polyandra Benth: effects of soil moisture and nutrients.Crossref | GoogleScholarGoogle Scholar | 32480910PubMed |
Winter K, Holtum JAM (2014) Facultative crassulacean acid metabolism (CAM) plants: powerful tools for unravelling the functional elements of CAM photosynthesis. Journal of Experimental Botany 65, 3425–3441.
| Facultative crassulacean acid metabolism (CAM) plants: powerful tools for unravelling the functional elements of CAM photosynthesis.Crossref | GoogleScholarGoogle Scholar | 24642847PubMed |
Winter K, Holtum JAM (2017) Facultative crassulacean acid metabolism (CAM) in four small C3 and C4 leaf-succulents. Australian Journal of Botany 65, 103–108.
| Facultative crassulacean acid metabolism (CAM) in four small C3 and C4 leaf-succulents.Crossref | GoogleScholarGoogle Scholar |
Winter K, Holtum JAM, Smith JAC (2015) Crassulacean acid metabolism: a continuous or discrete trait? New Phytologist 208, 73–78.
| Crassulacean acid metabolism: a continuous or discrete trait?Crossref | GoogleScholarGoogle Scholar | 25975197PubMed |