

Restoring dissolved organic carbon subsidies from floodplains to lowland river food webs: a role for environmental flows?
Darren S. Baldwin A B H , Matthew J. Colloff B , Simon M. Mitrovic C D , Nick R. Bond E G and Ben Wolfenden FA The Murray–Darling Freshwater Research Centre, LaTrobe University, PO Box 821, Wodonga, Vic. 3689, Australia.
B CSIRO Land and Water, GPO Box 1700, Canberra, ACT 2601, Australia.
C Applied Ecology Team, School of Life Sciences, University of Technology, Sydney, PO Box 123, Broadway, NSW 2007, Australia.
D New South Wales Department of Primary Industries: Water, Elizabeth Macarthur Agricultural Institute, Private Bag 4008, Narellan, NSW 2567, Australia.
E Australian Rivers Institute, Griffith University, 170 Kessels Road, Nathan, Qld 4111, Australia.
F Institute of Land Water and Society, Charles Sturt University, PO Box 789, Albury, NSW 2640, Australia.
G Present address: Murray–Darling Freshwater Research Centre, La Trobe University, PO Box 821, Wodonga, Vic. 3689, Australia.
H Corresponding author. Email: darren.baldwin@csiro.au
Marine and Freshwater Research 67(9) 1387-1399 https://doi.org/10.1071/MF15382
Submitted: 7 October 2015 Accepted: 28 January 2016 Published: 12 April 2016
Abstract
Environmental flows are managed events in river systems designed to enhance the ecological condition of aquatic ecosystems. Although not traditionally seen as important in lowland rivers, there is mounting evidence that terrestrial subsidies can be an important energy source in aquatic metazoan food webs. We argue that the apparent lack of importance of terrestrial subsidies to many lowland river food webs may reflect an artefact resulting from historical anthropogenic changes to lowland river–floodplain ecosystems, including the loss of lateral connectivity between rivers and their floodplains, changes in floodplain land use and carbon stores, and loss of sites of transformation within the main channel. The loss of floodplain subsidies to the main river channel can be partially redressed using environmental flows; however, this will require mimicking important aspects of natural high-flow events that have hitherto been overlooked when targeting environmental flows to a limited suite of biota. We suggest that key biotic targets for environmental flow releases may not be achievable unless river–floodplain subsidies are sufficiently restored. Environmental flows can go some way to addressing this shortfall, but only if floodplain subsidies to river channels are explicitly included in the design and management of environmental flows.
Additional keywords: bioenergetics modelling, C, levee, Murray–Darling Basin.
Introduction
One of the most striking impacts of river regulation globally is a reduction in the frequency with which rivers spill out onto the surrounding floodplain. In the case of flood-control dams, this reduction reflects the direct goals of river regulation, although in many other circumstances it is a side effect of storing or diverting water for human consumption. Recognising the loss of connectivity between river channels and their floodplains has resulted in environmental degradation; jurisdictions world-wide have begun implementing environmental flows (Arthington 2012). Environmental flows are managed releases of water designed to enhance the ecological condition of riverine ecosystems altered as a result of river regulation (Arthington 2012; see also http://water.worldbank.org/topics/environmental-services/environmental-flows, accessed 3 September 2015). Reinstating natural floods within an environmental-flow context is often contentious because of the risks posed to human assets on floodplains and because of the large volumes of water required; however, it is, nonetheless, often seen as an essential component of river restoration. However, the goals of environmental flows are often focussed on maintaining particular species or communities that are reliant on inundated floodplains, such as water birds and floodplain vegetation, often with a focus on flow events critical to the life history of the target biota. In the present paper, we argue that if environmental flows are being utilised, consideration should be given to their potential to restore subsidies of materials, specifically dissolved organic carbon (DOC), from a floodplain to lowland river food webs.
There are several means by which floodplain inundation may contribute to riverine food webs. First, large quantities of material (nutrients and carbon) may be leached from floodplain soils and vegetation, thereby supporting high rates of algal production and bacterial respiration. Second, this increase in basal resources can promote higher rates of secondary production by aquatic invertebrates (including the emergence of zooplankton from a propagule bank), which fuels aquatic food webs, that is, the so-called flood-pulse concept (Junk et al. 1989; Winemiller 2004). Furthermore, by recharging soil moisture and rejuvenating semi-permanent wetland habitat, floods contribute to greater terrestrial primary production during the inter-flood period, which further enhances resource availability during subsequent flood events by reciprocal provisioning (Baldwin et al. 2013).
The contribution by floods to riverine production can be enormous. For example, it was estimated that the inundation of the Barmah Forest (south-eastern Australia) in 2010, which flooded ~50 000 ha of forest, returned 7 tonnes (or 7 Mg) of zooplankton, 15 Mg of phytoplankton (assuming a 50 : 1 ratio between phytoplankton biomass and chlorophyll-a concentration) and 300 Mg of dissolved organic carbon to the Murray River (Nielsen et al. 2015). Similarly, a flood event on the Parana River in South America resulted in the mobilisation of an additional 4 × 106 Mg of carbon (mostly as DOC), approximately twice the normal carbon load for the river (Depetris and Kemp 1993).
However, whereas the role of the flood pulse and floodplain inundation in driving production in large lowland rivers seems inarguable, there remains much debate about the relative importance of terrestrial and aquatic carbon sources to riverine food webs (Jenerette and Lal 2005), especially, the importance of dissolved organic carbon. Empirical data certainly support the role of terrestrial detritus as important carbon sources for aquatic food-webs in specific places and at specific times (e.g. Reid et al. 2008; Hladyz et al. 2012); however, the circumstances, and relative contributions to sustaining food-web biomass, remain conjectural. The classical view is that pathways of bacterial uptake for terrestrial carbon are inefficient because of that carbon being recalcitrant (biologically inactive). Furthermore, only a small fraction of the bacterial production is actually incorporated into higher food web, whereas a large amount of the production stays within the ‘microbial loop’ (Thorp and Delong 2002); this occurs, in part, because of the potentially lower nutritional value of bacterial cells, which may, for example, lack essential polyunsaturated fatty acids necessary for metazoan growth (Saikia and Nandi 2010). From this basis, it is argued that algae represent a more important source of basal energy (Thorp and Delong 2002), although much of this algal production may also be occurring on the floodplain (Jardine et al. 2012). Better understanding of the relative importance of these different carbon sources has potentially important implications for restoring streams and rivers (Stanley et al. 2012), including efforts to restore river–floodplain connectivity with environmental flows.
The goal of the present paper is to review the evidence base for the role of DOC in aquatic food webs within the context of anthropogenically altered floodplain rivers. We focus on rivers of the southern Murray–Darling Basin in Australia as a case study, and pay particular attention to the ways in which environmental flows may alter carbon and food-web dynamics. We conclude with recommendations for future research needs to address some of the more critical current knowledge gaps.
The importance of DOC in contemporary riverine food webs
The importance of terrestrial DOC subsidies (sensu Polis et al. 1997) to large lowland river food webs is far from certain. In their synthesis paper, Thorp and Delong (2002) reviewed lowland river food webs from across the globe and found that the principal basal resource was almost always autochthonous carbon. Although they recognised that, at times, there is a large flux of carbon from floodplains to the channel of unconstrained rivers, they argued that this carbon was mostly recalcitrant, and the fraction that was readily available was mostly retained in the microbial loop. This hypothesis is partially supported by a more recent review of food-web studies in 31 large rivers (Roach 2013), which also found algal production to be the predominant basal resource in most lowland rivers. In all, Roach (2013) identified only five river systems where carbon from terrestrial (C3) plants was the dominant basal resource and an additional seven rivers where terrestrial carbon may have partially contributed to basal production. The overriding factor of whether or not terrestrial input into food webs was an important resource to maintain aquatic organisms appeared to be high flows or increased carbon loads, either through high concentrations of DOM in the river or from litter fall (Roach 2013).
Despite the predominance of studies reporting autochthonous sources as the dominant basal resource, several studies have found evidence to suggest that allochthonous DOC can be an important basal resource in inland aquatic ecosystems. In their seminal paper, Carpenter et al. (2005) showed that terrestrial carbon accounted for up to half the carbon (energy) flow to a variety of fish species inhabiting small lakes (0.9–2.5 ha). Similarly, Tanentzap et al. (2014) found that terrestrial sources supported 34–66% of fish biomass in lakes, with the percentage of fish biomass supported by terrestrial sources increasing as forest cover in the catchment increased. Reid et al. (2008) also found (using stable isotope analysis) terrestrial leaf litter to be a major basal food source supporting fish and predatory invertebrates.
Far from being recalcitrant, several studies have shown that DOC from floodplains is highly bioavailable (Baldwin 1999; Wallace et al. 2008; Baldwin et al. 2014). For example, in 2010–2011, following the ‘Millennium Drought’ (1997–2010), there was a moderate-sized flood along the Murray River (see Whitworth et al. 2012 for a detailed discussion of the flood). During the flood event, a large amount of dissolved organic carbon entered the river from its floodplain (Whitworth et al. 2012). Rather than carbon not being bioavailable, from data presented in Whitworth et al. (2012), Baldwin et al. (2014) estimated that up to 650 kg DOC river-km–1 day–1 was consumed along a 500-km stretch of the river over a 6-month period. By way of comparison, the average gross primary production along the same stretch of river during a period without overbank flows was estimated to be 5.7 kg C km–1 day–1 (Oliver and Merrick 2006).
These studies are complemented by more direct evidence that DOC from floodplain sources is not constrained to the microbial loop. Both laboratory (Baldwin et al. 2014) and field studies (Cook et al. 2015) have shown that DOC can be rapidly assimilated into riverine biofilms, from where it can enter the metazoan food web through grazing by aquatic invertebrates. Similarly, it has been shown that within 8 days, zooplankton density approximately tripled in mesocosms containing water from a lowland river that had been dosed with DOC leached from leaf litter, compared with non-dosed controls (Mitrovic et al. 2014). Further, the addition of glucose (which has more resemblance to autochthonous carbon sources), a less complex source of carbon than DOC, led to the development of different zooplankton communities and fewer large species (copepods), suggesting that allochthonous DOC may play an important role in determining the structure and diversity of elements of aquatic food webs. In another study, Zeug and Winemiller (2008), using stable isotope analysis, showed that riparian C3 plants were the predominant basal resource for almost all consumers in the main channel of the Brazos River in Texas (USA) and most consumers in adjacent oxbow lakes.
So, the question arises as to why we do not see more evidence that allochthonous carbon is important in the functioning of lowland rivers? Zeug and Winemiller (2008) contended that one of the reasons for the discrepancy is that many of the studies that have shown that autochthonous, rather than allochthonous, carbon sources are more important in riverine function are undertaken during periods when there was little or no connectivity between a river and its floodplain (e.g. Thorp et al. 1998; Bunn et al. 2003; Delong and Thorp 2006; see also Oliver and Merrick 2006). Conversely, they suggested that studies that have included different flow regimes (including high flows) have shown that terrestrial carbon can be an important basal resource (Huryn et al. 2001; Wantzen et al. 2002; Zeug and Winemiller 2008; Hladyz et al. 2012; Cook et al. 2015). Another (complementary) explanation is the significant anthropogenic changes to river–floodplain ecosystems world-wide, particularly in temperate regions (Bayley 1995).
Effects of anthropogenic changes on the carbon dynamics of lowland rivers and floodplains
River regulation and the loss of lateral flows from the floodplain to the main stem of the river
The flow regimes of many lowland rivers are regulated with headwater dams that capture high flows that would otherwise inundate lowland floodplain wetlands. There are an estimated 40 000 large dams (with walls higher than 15 m) world-wide (World Commission on Dams 2000), and over half of the largest river systems in the world are heavily modified by dams and storages (Nilsson et al. 2005). These dams and storages have had a profound effect on the connections between lowland rivers and their floodplains, affecting the extent, frequency, duration and timing of floodplain inundation (Kingsford 2000; Vörösmarty et al. 2010). In addition, so as to protect agricultural land and communities from inundation, many rivers have been disconnected from their floodplains by levee banks and other flood-control structures. For example, over 80% of the floodplain of the Lower Mississippi River (~10 × 106 ha) is subjected to greatly reduced frequency of inundation by levees (Kesel 2003), although failure of levees during major flood events has been high (Tobin 1995).
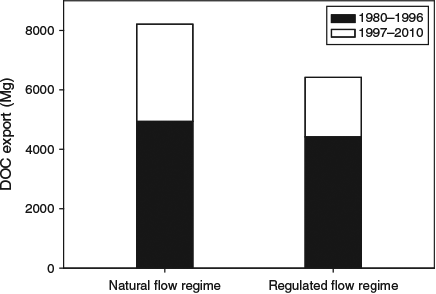
The loss of lateral connectivity between rivers and their floodplains has altered not just water movement, but also the flux of materials and energy (nutrients and carbon) and biota (plant propagules and animals) from and to the river (Bond et al. 2014). Consequently, the export of DOC from floodplain ecosystems to the main river channel is likely to have been substantially reduced. As an example, we have modelled carbon export from a moderately sized floodplain forest (Koondrook–Perricoota Forest, south-eastern Australia) under natural and regulated flow regimes (see Supplementary material for this paper). During periods when the flow regime is close to the long-term average, there would not have been a substantial difference in the amount of DOC exported from the forest under natural or regulated flow regimes (4940 Mg compared with 4420 Mg for the period 1980–1996; Fig. 1). However, during an extended period of drought (1997–2010), 3270 Mg of DOC would have been exported from the floodplain under natural flow conditions, whereas only 2000 Mg would have been exported under regulated flow conditions. The main difference was that, even in the face of severe drought condition, there would have been several significant flood events in the forest in the absence of river regulation; however, because of regulation, these floods were captured by upstream water storages (see Supplementary material).
|
Westhorpe and Mitrovic (2012) also modelled DOC export from the lower floodplain Namoi River, on the basis of scenarios with flow regulation and simulated pre-development conditions. They found that in wet years with floods, almost 23 000 Mg of DOC could have been exported under pre-regulation flow regimes, whereas under current flow regimes, only 14 775 Mg of DOC was exported. In years with no major floods, much lower DOC export occurred, with, on average, 1500 Mg exported under no-regulation and 741 Mg under a regulation scenario.
In the highly regulated Murrumbidgee River catchment, in eastern Australia, low rates of ecosystem respiration, and a high contribution of phytoplankton to gross primary production, have also been attributed to the reduction in terrestrial carbon inputs from river regulation (Vink et al. 2005). Given that overbank floods are one of the main pathways for allochthonous carbon to enter the river channel, it is not surprising that studies in such systems fail to find that allochthonous DOC is an important basal resource for food webs in rivers with river regulation and flood-mitigation infrastructure.
Changes in land use
Floodplains are highly productive compared with surrounding dryland ecosystems, because of sediment and nutrient deposition from previous floods (e.g. Tockner et al. 1999) and the high availability of surface water and alluvial groundwater. So as to take advantage of this productive land, floodplain forests, woodlands and shrublands have been extensively cleared and replaced with irrigated and dryland crops and pastures. In floodplain forests and woodlands, much more of the carbon that is fixed on the floodplain remains in situ (as standing biomass, large woody debris, litter and soil carbon), than in cropping and pasture land, until it is mobilised following inundation. Under crops or pastures, a large proportion of the carbon fixed on the floodplain is exported for human or stock consumption. Furthermore, although it is possible for DOC to be leached from crops and pastures if inundated (Whitworth et al. 2012), the likelihood of flooding of these systems is often significantly reduced by flood-protection levees (see above). Hence, shifts to agricultural and grazing systems decrease the amount of carbon stored on floodplains. Furthermore, loss of woody vegetation adjacent to the river channel reduces the amount of carbon that is available to enter the river through direct litter fall and tree fall (e.g. Mac Nally et al. 2002).
The clearing of native vegetation and the establishment of agriculture and grazing on floodplains have tended, historically, to precede river regulation, and provide some of the rationale for it. These justifications include the protection of property and crops from inundation, the supply of water for the development of irrigation districts and the amelioration of the effects of drought (e.g. Worster 1985; Powell 1993).
Loss of sites of transformation
Even if DOC could enter the river channel, loss of sites of transformation have most probably reduced the efficiency with which the DOC could be assimilated into high-order consumers. For DOC to enter the metazoan food web, it first needs to be assimilated into bacteria, either those in the water column or those associated with biofilms. Carbon has been shown to be limiting to bacterial growth in some Australian lowland rivers (Westhorpe et al. 2010; Mitrovic et al. 2014). Transfer of carbon into the metazoan food web is arguably more efficient through biofilms than through planktonic pathways (see Baldwin et al. 2014). However, hard substrates, particularly large woody debris, which would have been sites of transformation for converting dissolved organic carbon into biofilm carbon, have been removed from many lowland rivers to facilitate navigation as well as for flood mitigation (e.g. Gippel 1995).
It has been hypothesised that one reason that DOC is not identified as an important basal resource for lowland river food webs is because of the inefficiencies associated with the transfer of carbon through planktonic bacteria rather than biofilms (Baldwin et al. 2014). However, despite inefficiencies with allochthonous DOC incorporation into the planktonic food web (often involving one or more steps in the food chain compared to phytoplankton carbon routes; Jansson et al. 2000), the quantity and concentration of DOC available after overbank inundation events can be much higher than those of DOC available via autochthonous sources (Westhorpe and Mitrovic 2012). This can allow zooplankton and fish the potential to develop much greater biomass (Mitrovic et al. 2014). There is also evidence that bioavailability may, in some instances, increase with inundation (Hitchcock and Mitrovic 2015).
Biofilms on large woody debris also have the potential to reduce the carbon-uptake length of a river reach (Baldwin et al. 2014). The carbon-uptake length is the length of river necessary to reduce the concentration of a constituent, in this case DOC, by a certain proportion. The more biofilm in a given reach, the shorter the uptake length, and the more likely that DOC exported from the floodplain will remain in the immediate vicinity of the return flows. In the absence of large woody debris and their associated biofilm, DOC would be lost from the river reach, by being transported downstream from the immediate area before being taken up by biota.
Case study: historical changes in land use and effects of river regulation on carbon dynamics in the southern Murray–Darling Basin
The following section details some gross changes to carbon dynamics associated with clearing, land-use changes and river regulation in lowland floodplains of the southern Murray–Darling Basin following European colonisation in the 1830s and 1840s (Fig. 2). Reviews of the effects of river regulation in eastern Australia have tended to downplay changes in carbon dynamics and their implications for food webs and productivity (Walker and Thoms 1993; Kingsford 2000) and have focussed instead on changes in flow regimes and their effects on biotic communities and on flow restoration and protection of rivers (Arthington and Pusey 2003). However, in a review of the dynamics of carbon in Australian floodplain rivers, Robertson et al. (1999, p. 825) hypothesised that ‘…for those rivers where floodplains have been alienated from their river channels by altered flow regimes, and for which there have been changes to the quality of in-channel organic matter … there has been a shift in the balance such that longitudinal fluxes of organic matter and in-channel algal production dominate carbon pools. This is because allochthonous inputs from floodplains and riparian habitats have been greatly reduced.’

|
Initially, land-use changes involved the establishment of extensive sheep- and cattle-grazing runs within existing communities of native floodplain vegetation and the disruption or cessation of Aboriginal burning regimes, but without radical changes to the treescape. Whereas these changes had implications for carbon dynamics, for example, via the altered composition and distribution of riparian vegetation communities and their regenerative capacity (Robertson and Rowling 2000; Price et al. 2010), the details and scale of these effects are poorly understood. By way of contrast, widespread tree clearing followed the 1850s Victorian gold rushes and the expansion of intensive settlement from the 1860s (Barr and Carey 1992), as well as the felling of riparian river red gum forests for timber (Colloff 2014). Walker et al. (1993) estimated there are 12–15 billion fewer trees in the Murray–Darling Basin now than before European settlement, being a loss of ~65%.
Riparian forests and woodlands had provided large inputs of carbon to rivers from leaf litter and from coarse woody debris that accumulated on the floodplain and within river channels. Removal of timber from the Murray channel to improve navigability (‘de-snagging’) between 1865 and 1925 resulted in loss of an estimated 1500 snags river-km–1 (Phillips 1972; Treadwell et al. 1999), being equivalent to a surface area of 195 000 m2 river-km–1 for potential colonisation by biofilms (Baldwin et al. 2014). Clearing of fallen timber from floodplain forests for firewood, possibly by up to 85% of loads pre-European settlement (Mac Nally et al. 2011), further reduced biofilm potential (cf. below).
So as to protect crops and pastures from inundation, an extensive series of levee banks was constructed along the central Murray from the 1890s (Dexter 1978), isolating the river from its floodplain and reducing the extent and frequency of floods and the transfer of allochthonous carbon to the channel. Establishment of irrigation districts along the Murray and Murrumbidgee valleys from the late 1880s heralded the construction of major dams and storages and the development of water resources from the 1900s (Powell 1989), with basin-wide irrigation diversions intensifying from ~4000 GL in 1955 to over 10 000 GL in 1985 (Colloff et al. 2015) and 12 major storages completed between 1928 and 1979 (Fig. 2b). The resultant reductions in the volume, frequency and duration of overbank flows caused major contractions in the flood regimes of wetlands in the southern basin (Sims et al. 2012). Consequently, considerably less organic matter now accumulates on smaller areas of floodplain than before European colonisation and there are fewer, shorter periods for generation of DOC during flood events and its transfer to river channels (e.g. see Supplementary material).
In addition to DOC derived from woody vegetation, emergent macrophytes provide an important source of carbon in low-lying wetlands and floodplains adjacent to rivers (Vivian et al. 2014). This above-ground biomass can be considerable (in the order of tens of tonnes per hectare, e.g. Colloff et al. 2014), and DOC leached from this material is available for export to rivers during flood events (e.g. Whitworth et al. 2012). Grassy wetlands and reed beds have declined greatly in extent since river regulation in the southern Murray–Darling Basin (Colloff 2014; Colloff et al. 2014), indicating major reductions in macrophyte-derived DOC sources.
Characteristics of rivers and floodplains that determine the relative importance of DOC as a basal resource for riverine food webs
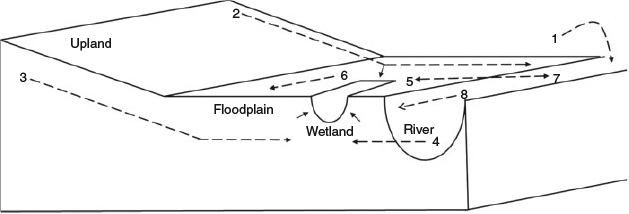
There are multiple pathways for the movement of DOC throughout river–floodplain systems (Fig. 3), including the following:
-
Litter- and tree fall from riparian vegetation directly into the river channel and associated benches.
-
Overland flow from the upland and floodplain during rain events, including flows along channels or into wetlands.
-
Subsurface leaching of DOC through the soil profile or groundwater from the floodplain to the river channel, which may be intercepted by wetlands.
-
Subsurface flows of water from the river channel to the floodplain.
-
Lateral movement of DOC from the river to the floodplain during overbank flood events.
-
Movement of DOC across and along the floodplain during the flood event (including flood runners), which will leach DOC from litter and soil but also entrain DOC in floodplain wetlands.
-
Floodwater returns from the floodplain back to the river channel.
-
Longitudinal movement of DOC along the river channel.
|
The relative importance of each pathway will depend on the physical characteristics of the river and its floodplain, and will vary in time and space (see Supplementary material for a more detailed discussion on how each pathway affects DOC).
It is conceptually possible to identify those characteristics of lowland river–floodplain systems in which DOC is (or was) an important basal resource. First, the load of readily bioavailable DOC that enters (or entered) the river from the floodplain must make a significant contribution to the overall energy budget of the river; for example, the annualised DOC load should be at least of the same order of magnitude as the amount of carbon fixed in the river channel. This means that river reaches that have (or had) large areas of floodplain that are (were) regularly inundated would be more likely to have food webs supported by allochthonous DOC. Similarly, floodplains that are dominated by vegetation types that are a good source of bioavailable DOC are more likely to have food webs supported by allochthonous DOC than are those with vegetation that produces poorly available DOC (e.g. Wallace et al. 2008) or, indeed, those without any appropriate vegetation. There is some evidence that threshold bioavailable DOC concentrations may need to be met for effective transfer of energy through the food web (Hitchcock et al. 2015). Lower concentrations of DOC may result in losses through respiration and trophic transfer efficiency across steps in the microbial food chain. Higher DOC concentrations over longer periods could provide enough energy to subsidise the food web (Karlsson et al. 2007). This finding underscores the importance of the DOC supply from floodplains in achieving such threshold concentrations and the benefits from inundating areas with substantial natural floodplain vegetation and DOC sources rather than those with reduced quantity and quality of DOC. The targeting of these areas with environmental flows at appropriate intervals to ensure that adequate DOC resource is available, or to be timed to meet enhanced leaf fall or leaf senescence, may be a tool to achieve this.
When considering whether, historically, DOC may have been important to riverine food webs before human modification, the extent of the channel networks on the floodplain should be taken into consideration, especially the surface area of the total channel network compared with the size of the receiving channel. Floodplains that are highly channelised (including anabranches) are more likely to contribute DOC to support riverine food webs (Hladyz et al. 2011; McGinness and Arthur 2011). These channels serve to move water to and from the floodplain and can be filled either from overland flow from the catchment (Pathway 2) or from overbank flows. Because they are low points in the landscape, they tend to fill first, and, in small overbank flows, may be the only part of the floodplain that fills. Because they are the first to fill, they are also the part of the floodplain that is flooded most often (i.e. both during small, medium and large events) and, hence, unlike other parts of the floodplain, may be flooded multiple times in the same year. Because they are low points in the landscape, litter, mostly from fringing vegetation, accumulates in the dry channel or any remnant pools (Hladyz et al. 2011; McGinness and Arthur 2011).
The food web in a small river connected to a floodplain with an extensive network of channels would more likely be supported by DOC than would a very large river channel with a floodplain with few, if any, channels. Other things to take into consideration include how often the channels would have flooded on an annual basis, the nature of the natural floodplain vegetation, and the mechanisms of litter fall and accumulation. Litter accumulation in channels would be more likely to occur in floodplains dominated by trees rather than grasses. In floodplains dominated by deciduous trees, because litter fall occurs only once in a year, DOC export from the floodplain channels to the main river will mostly occur during the first inundation event following leaf fall. Therefore, DOC export from the floodplain will at best occur only once a year. In such a case, the importance of DOC export from the floodplain to the riverine food web will depend on the ability of the riverine biota to capture and store the DOC, which in turn will depend on the sites of transformation in the river system (see above). In floodplains dominated by evergreens such as Eucalyptus species, even though leaf fall may occur predominantly in one season, it occurs throughout the year (e.g. Glazebrook and Robertson 1999). For floodplain eucalypts such as the river red gum, litter fall is a normal physiological response to drought stress and major declines (≥90%) and subsequent recovery in leaf area index are common (Doody et al. 2015). Multiple flooding of river channels will lead to multiple pulses of DOC back to the river channel, although terrestrial ageing of fallen leaves means that the timing of inundation will play a role in the magnitude, and possibly lability, of DOC released (Watkins et al. 2010).
River reaches with large amounts of large woody debris (and, hence, a proliferation of biofilm) should be more efficient at trapping allochthonous DOC (Baldwin et al. 2014). Therefore, it is more likely that allochthonous DOC would be an important basal resource in heavily snagged reaches than reaches without snags.
Finally, rivers with multiple connections to their floodplains are more likely to be seen as using DOC as a basal resource along their entire lengths than are rivers with only a few connections. As DOC moves downstream, it undergoes a series of biotic and abiotic reactions that serve to reduce its concentration (Fig. 3, Pathway 8). Furthermore, because the most readily bioavailable components of DOC are consumed first, the further away from the source of the DOC, the more likely the remaining DOC will be more recalcitrant. Therefore, both the quantity and quality of DOC will diminish with an increasing distance from its source, and therefore the less likely allochthonous DOC will be an important basal resource supporting riverine food webs.
Environmental flows and carbon subsidies to lowland rivers
Environmental flows have the capacity to influence mobilisation and the quality of DOC mobilised (Westhorpe and Mitrovic 2012). There is a range of environmental flow types that have been either implemented or suggested for rivers (Acreman et al. 2014). Some environmental flows have directly considered DOC mobilisation in the process of development such as the New South Wales Integrated Monitoring of Environmental Flows Program in Australia, which had a carbon hypothesis for particular environmental flow types (Chessman 2003). Even where allochthonous DOC is not considered, some of the different types of environmental flows may still act to influence DOC concentrations and loads. Some of the common types of environmental flows and their likely effect on DOC concentration, load and food webs are given below.
High-flow protection rules
These environmental flows generally protect high-flow events from extraction for irrigation by limiting the amount of water that can be extracted. For instance, these rules may set flow thresholds above which irrigation water cannot be extracted or restrict the amount of water permitted to be extracted during high-flow events (such as tributary flows or dam spills). The access will depend on licence types and these flow rules have been applied in several large catchments such as the Namoi, Gwydir and Hunter. By engaging floodplain habitat, riparian zones, low-lying river channels and in-channel features, these types of flows will generally increase DOC concentrations and mobilise carbon. Westhorpe and Mitrovic (2012) found that DOC concentrations increased from ~5–7 mg L–1 under low-flow scenarios to a mean of 20.4 mg L–1 during floods, with concentrations sometimes over 40 mg L–1. By using a linear relationship between DOC concentration and flow, they estimated that in years with high-flow or flood events, the DOC loads increased ~40% (or by 6000 Mg) with environmental flows, which was closer to predicted natural loads. Years with low flows did not lead to greatly changed loads for the different scenarios because high-flow protection environmental flows were not activated, showing the importance of protecting the less frequent high-flow events that break out of river channels into flood runners and inundate floodplains. These findings are similar to those of other studies that have shown that high-flow events of small duration account for a disproportionately high amount of the DOC load delivered to rivers (Dalzell et al. 2007; Hitchcock and Mitrovic 2015).
First-flush protection rules
These environmental flow rules are aimed at protecting the first part of a flow event that may carry greater DOC and nutrient load. For instance first-flush rules may protect the first part of a flow event from extraction such as in the Hunter River, Australia, where the first 12 h are protected to improve river and estuarine productivity (Hitchcock et al. 2010). They tend to be activated when flows reach a certain level or a river height and it is assumed that a flow pulse will be carried down the river. The first flush can be shown to carry higher concentrations of DOC and nutrients as it wets the river channel and low-lying benches. Westhorpe and Mitrovic (2012) showed that, in some instances, the highest DOC concentrations were associated with the early part of the flood event. However, the loads were much greater during the peaks of flow when DOC was generally higher. These rules have been used quite widely in some jurisdictions and have been applied to, or suggested for, rivers and estuaries such as the Hunter, Bega, Wyong and Brunswick Rivers in Australia.
Water purchasing for environmental benefit
Water can be purchased on open markets for use as an environmental flow and has been done so for several rivers including the Murray, Murrumbidgee, Macquarie and Gwydir Rivers. Without diversion infrastructure, environmental water deliveries need to exceed discharges required to fill adjacent wetlands for successful floodplain inundation, which can be expensive in terms of both water and delivery costs. However, if the purchased water can wet the floodplain (benches or billabongs) and waters return to the river, then this may be an effective method. In most cases, it will be necessary to coordinate deliveries of held water entitlements, other planned environmental water, tributary inflows and other water deliveries to achieve peak volumes for a sufficient duration to fill adjacent wetlands. Because water accounting, types of entitlements, and delivery arrangements vary widely among jurisdictions, planning coordinated deliveries requires significant pre-organisation. Unlike water delivered for irrigated agriculture, water delivered for environmental purposes under standard water entitlements is typically not protected and can thus be made available to meet other consumptive demand. The consequence of this lack of protection is that environmental water delivered to wetlands or river channels can be extracted before lateral or longitudinal mobilisation of DOC can be achieved (Banks and Docker 2014).
Environmental dam releases or environmental contingency allowances
Another common mechanism for delivering environmental flows is releases from dams for specified purposes. Some rivers that have these environmental flows include the Hunter, Gwydir, Severn and Lachlan rivers. These are often discretionary rules that might be contingent on an environmental need such as dispersing an algal bloom (Mitrovic et al. 2011), wetland replenishment, or fish- or bird-breeding events (Kingsford and Auld 2005). Unlike water entitlements held under general water-access licences, environmental water that is accrued specifically for the purpose of environmental flows is often protected from extraction, and is more readily delivered to facilitate longitudinal and lateral connectivity. DOC from dam releases tends to be low in concentration and autochthonous in nature (Hadwen et al. 2010; Rohlfs et al. 2015), and so will have little benefit from an allochthonous DOC food-web perspective. However, there is the potential for these flows to be piggy-backed onto downstream unregulated tributary events that may help increase river height to reach floodplain inundation levels, particularly if the commence-to-fill levels are known for bench, billabong or floodplain inundation. In this case, the environmental flow would be very useful for increasing delivery of allochthonous DOC to rivers.
Transparency and translucency rules (T & T rules)
These environmental flow rules apply to some dams, where the rules seek to both protect low flows and some aspects of the higher flows, while maintaining a higher level of variability in rivers and immediately pass a proportion of reservoir inflows downstream. The transparency rules pass all water through the dam below a given level (e.g. the 80th percentile), whereas the translucency rule releases a proportion of the flow into the dam (e.g. 20%) above the translucency level, to maintain variability and some flow pulses in the system (Growns and Reinfelds 2014). These environmental flow rules better mimic the historical flow regimes that biota are adapted to, albeit at a lower level. If the 20% translucency is applied, 20% of large flow events should be released downstream. There is some potential that these flow rules may be of some benefit in terms of allochthonous DOC delivery if large inflows are recorded, if a larger translucency percentage is utilised, or if releases piggy-back on to other unregulated tributary flows to enable bench and floodplain wetting. However, T&T flows often lack the protection of other environmental contingency flows and so their use in longitudinal DOC transport is limited. Some dams that currently use these rules are Burrinjuck Dam (Murrumbidgee River), Copeton Dam (Gwydir River) and Burrendong Dam (Macquarie River).
End-of-system flow rules
These environmental flow rules generally protect water at the bottom of a catchment by maintaining a minimum flow, which may be based on historic seasonal flows. These rules try to ensure that a minimum target flow is reached, to potentially improve water quality and reduce algal blooms (Mitrovic et al. 2008) or provide water for estuaries to stop upstream salt-wedge incursion (Lester et al. 2011). As these rules protect the lower end of the hydrograph, they are unlikely to be very useful for the mobilisation of large amounts of allochthonous DOC, although they may play an important role in providing downstream nutrient and carbon energy-resource supplies for these productive ecosystems (for example, estuaries, bays and the coastal zone).
Implicit in the preceding discussion is the assumption that overbank environmental flows are allowed to return to the river channel. For example, under natural flow conditions, many large floodplain lakes and wetlands would have been filled by overbank (sheet) flows, allowing lateral connectivity between the river and floodplain. Currently, it is common for environmental flows to be used to inundate these floodplain water bodies, specifically to enhance the condition of associated biota (Bond et al. 2014). Infrastructure (including pumps, levees and channels) is used to efficiently direct environmental water allocations into these water bodies. However, although it is common to provide a sufficient volume of water to inundate the water body, often there is not enough water to ensure any return flow (see Bond et al. 2014 for examples). Alternatively, there may be a reluctance to allow return flow from the floodplain because of the perceived risk of poor water-quality outcomes in the receiving water (see discussion in the following section).
Synthesis
The apparent dominance of autochthonous carbon in the food webs of floodplain–river ecosystems is likely to stem from the widespread flow regulation in studied rivers and from the episodic nature of floodplain carbon deliveries. However, there is mounting evidence that terrestrially derived carbon plays a fundamental role in the pattern and process of floodplain–river ecosystems. As the delivery of environmental flows (including the construction and operation of new floodplain infrastructure such as weirs, levees and regulators) grows to become one of the key tools used in the rehabilitation of regulated rivers, there is a pressing need for empirical evidence to support achievable outcomes for impaired riverine communities. Planning environmental flows to support DOC provisioning in regulated floodplain–river ecosystems is currently limited by several important factors, including the following:
-
a continuing focus on the apparent needs of salient biota in river ecosystems rather than the ecosystem processes that support those biota,
-
poor knowledge of the thresholds (concentrations or loads of DOC and co-limiting nutrients) needed to support ecosystem functioning,
-
a lack of understanding of the DOC concentrations and timing to which riverine communities are naturally adapted,
-
a lack of understanding of the consequences to riverine communities for reducing or totally removing episodic heterotrophy and moving towards systems that, for the most part, are reliant on in-channel autotrophic production (in particular planktonic autotrophic production),
-
severed longitudinal and lateral linkages, many of which are unlikely to be restored because of current socio-economic constraints,
-
the inability to deliver environmental flows to support DOC mobilisation at ecologically relevant scales, partially because of legislative or social constraints on the flooding of private land, and
-
a reluctance to manage for overbank flows, or if overbank flows do occur, a reluctance to allow flood water to return to the main channel, because of the risk of adverse water-quality outcomes, including the development of hypoxic blackwater plumes.
Some of these barriers are easily addressed. Questions of targets of environmental flows (charismatic biota compared with underlying ecosystem processes) are simply human constructs and, hence, are easily changed. However, more information needs to be gathered on the importance of ecosystem processes (including floodplain subsidies to lowland rivers) in supporting socially and culturally important organisms before there will be an appetite to change natural resource goals for environmental flows.
Risk of generating hypoxic blackwater also should not be a barrier limiting DOC subsidies from floodplains to riverine food webs. Computer models now exist that can be used to predict the likelihood of generating hypoxia following floodplain inundation (Howitt et al. 2007; Whitworth and Baldwin 2016). Although these models have been developed specifically for river red gum-dominated floodplains in south-eastern Australia, they can readily be modified to account for other vegetation types (Whitworth and Baldwin 2016). Using these models for scenario testing shows that the risk of generating hypoxic blackwater can be mitigated by appropriate timing of floodplain inundation, managing retention times, ensuring that water is not allowed to stand on the floodplain for too long, and having a sufficiently large dilution flow in the main channel to mitigate hypoxia if it does occur. Similar approaches can also be undertaken to assess and mitigate other water-quality risks associated with inundation floodplains, such as mobilisation of salt or seeding of blue-green algal blooms (e.g. see Baldwin 2014).
Some constraints are much more difficult to overcome. Because of extensive human colonisation of floodplain ecosystems, there is a large social and cultural barrier to the restoration of natural inundation patterns to large areas of floodplain ecosystems. Although inevitable on floodplains, natural overbank flows are now characterised as natural disasters rather than natural events. Therefore, deliberate inundation of privately held floodplain would undoubtedly result in significant public backlash. Although in no way advocating the inundation of privately owned floodplain without the tacit approval of the relevant landholder, nevertheless, the loss of the subsidy should be taken into account in natural resource management. For example, whole-of-system bioenergetics modelling can potentially be undertaken to determine how much actual energy (for example, from DOC, zooplankton and phytoplankton) and potential energy (through increased in-channel phytoplankton, epiphyte and macrophyte growth from floodplain subsidies of limiting nutrients) would be supplied from unconstrained floodplain inundation compared with other flow scenarios including limited environmental flows and no overbank flows. Such a modelling exercise would allow estimates of the constraints that limiting floodplain inundation has on riverine production, especially on the production of top-order consumers, including fish. This could be undertaken in conjunction with studies looking at the economic benefits of floods, particularly in arid and semi-arid landscapes. For example, from an agricultural perspective, floods can be a much more efficient way of recharging the soil-moisture profile and groundwater than is rainfall (e.g. Baldwin et al. 2013), resulting in a significant increase in post-flood primary production on the floodplain compared with rainfall responses (Baldwin et al. 2013); other examples have been canvassed by Opperman et al. (2009). Inundation of floodplains can also result in substantial potential benefits for the supply of ecosystem services (Zedler and Kercher 2005; Opperman et al. 2010). With appropriate incentives and governance arrangements, the benefits of these services can be realised by affected farmers. Furthermore, under increasing future water scarcity as a result of climate change, environmental flows represent a means for realising the benefits of adaptation services; the ecosystem processes and services that help people adapt to environmental change (Lavorel et al. 2015). Ultimately, public acceptance of changing flooding regimes to promote environmental and socio-economic benefits will require a commitment of relevant government and non-government agencies to increasing community understanding of the benefits of floods.
Although it is not possible to return river systems to a natural state, the loss of floodplain subsidies to rivers has most likely strongly influenced contemporary lowland river ecology. Restoring processes such as energy flows that sustain river–floodplain ecosystems, while continuing to provide water for irrigation and human infrastructure on floodplains, represents a major challenge (Opperman et al. 2009; Roseman and DeBruyne 2015). Nonetheless, many of the valued goods and services humans derive from floodplain rivers, depend on the health of those systems. Environmental flows have become a major restoration tool, but their benefits may depend on the explicit inclusion of floodplain subsidies as a restoration goal.
Acknowledgements
We thank Dr Rick Stoffels (CSIRO Land and Water) and two anonymous referees for critically reviewing an earlier version of this manuscript.
References
Acreman, M. C., Arthington, A. H., Colloff, M. J., Couch, C., Crossman, N. D., Dyer, F., Overton, I. C., Pollino, C. A., Stewardson, M. J., and Young, W. J. (2014). Environmental flows for natural, hybrid and novel riverine ecosystems in a changing world. Frontiers in Ecology and the Environment 12, 466–473.| Environmental flows for natural, hybrid and novel riverine ecosystems in a changing world.Crossref | GoogleScholarGoogle Scholar |
Arthington, A. H. (2012). ‘Environmental Flows: Saving Rivers in the Third Millennium.’ (University of California Press: Berkeley, CA.)
Arthington, A. H., and Pusey, B. J. (2003). Flow restoration and protection in Australian rivers. River Research and Applications 19, 377–395.
| Flow restoration and protection in Australian rivers.Crossref | GoogleScholarGoogle Scholar |
Baldwin, D. S. (1999). DOM and dissolved P leached from fresh and ‘terrestrially’-aged river red gum leaves: implications for assessing river–floodplain interactions. Freshwater Biology 41, 675–685.
| DOM and dissolved P leached from fresh and ‘terrestrially’-aged river red gum leaves: implications for assessing river–floodplain interactions.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1MXlslKjtLg%3D&md5=a382f10d3958aaa30d5e9c93750d3d8dCAS |
Baldwin, D. S. (2014). Evaluating the River Murray system aggregated risks from environmental watering actions at TLM Icon Sites. MDFRC Publication 30/2014. Available at http://www.mdba.gov.au/kid/files/2560%20-%20Evaluating%20the%20River%20Murray%20system%20aggregated%20risks%20from%20environmental%20watering%20actions%20at%20TLM%20Icon%20Sites.pdf [Verified 30 November 2015].
Baldwin, D. S., Rees, G. N., Wilson, J. S., Colloff, M. J., Whitworth, K. L., Pitman, T. L., and Wallace, T. A. (2013). Provisioning of bioavailable carbon between wet and dry periods in a semi-arid floodplain. Oecologia 172, 539–550.
| Provisioning of bioavailable carbon between wet and dry periods in a semi-arid floodplain.Crossref | GoogleScholarGoogle Scholar | 23124331PubMed |
Baldwin, D. S., Whitworth, K. L., and Hockley, C. L. (2014). Uptake of dissolved organic carbon by biofilms provides insights into the potential impact of loss of large woody debris on the functioning of lowland rivers. Freshwater Biology 59, 692–702.
| Uptake of dissolved organic carbon by biofilms provides insights into the potential impact of loss of large woody debris on the functioning of lowland rivers.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC2cXktV2hsr4%3D&md5=a3a610119b79de858bb36a2b328529b3CAS |
Banks, S. A., and Docker, B. B. (2014). Delivering environmental flows in the Murray–Darling Basin (Australia): legal and governance aspects. Hydrological Sciences Journal 59, 688–699.
| Delivering environmental flows in the Murray–Darling Basin (Australia): legal and governance aspects.Crossref | GoogleScholarGoogle Scholar |
Barr, N. C., and Carey, J. (1992). ‘Greening a Brown Land: the Australian Search for Sustainable Land Use.’ (Macmillan: Melbourne.)
Bayley, P. B. (1995). Understanding large river: floodplain ecosystems. Bioscience 45, 153–158.
| Understanding large river: floodplain ecosystems.Crossref | GoogleScholarGoogle Scholar |
Bond, N., Costelloe, J., King, A., Warfe, D., Reich, P., and Balcombe, S. (2014). Ecological risks and opportunities from engineered artificial flooding as a means of achieving environmental flow objectives. Frontiers in Ecology and the Environment 12, 386–394.
| Ecological risks and opportunities from engineered artificial flooding as a means of achieving environmental flow objectives.Crossref | GoogleScholarGoogle Scholar |
Bunn, S. E., Davies, P. M., and Winning, M. (2003). Sources of organic carbon supporting the food web of an arid zone floodplain river. Freshwater Biology 48, 619–635.
| Sources of organic carbon supporting the food web of an arid zone floodplain river.Crossref | GoogleScholarGoogle Scholar |
Carpenter, S. R., Cole, J. J., Pace, M. L., Van De Bogert, M., Bade, D. L., Bastviken, D., Gille, C. M., Hodgson, J. R., Kitchell, J. F., and Kritzberg, E. S. (2005). Ecosystem subsidies: Terrestrial support of aquatic food webs from C-13 addition to contrasting lakes. Ecology 86, 2737–2750.
| Ecosystem subsidies: Terrestrial support of aquatic food webs from C-13 addition to contrasting lakes.Crossref | GoogleScholarGoogle Scholar |
Chessman, B. C. (Ed.) (2003). Integrated monitoring of environmental flows (IMEF); State summary report 1998–2000. Department of Infrastructure, Planning and Natural Resources, Sydney.
Colloff, M. J. (2014). ‘Flooded Forest and Desert Creek: Ecology and History of the River Red Gum.’ (CSIRO Publishing: Melbourne.)
Colloff, M. J., Ward, K. A., and Roberts, J. (2014). Ecology and conservation of grassy wetlands dominated by spiny mud grass Pseudoraphis spinescens in the southern Murray–Darling Basin, Australia. Aquatic Conservation: Marine and Freshwater Ecosystems 24, 238–255.
| Ecology and conservation of grassy wetlands dominated by spiny mud grass Pseudoraphis spinescens in the southern Murray–Darling Basin, Australia.Crossref | GoogleScholarGoogle Scholar |
Colloff, M. J., Caley, P., Saintilan, N., Pollino, C. A., and Crossman, N. C. (2015). Long-term ecological trends of flow-dependent ecosystems in a major river basin. Marine and Freshwater Research 66, 957–969.
| Long-term ecological trends of flow-dependent ecosystems in a major river basin.Crossref | GoogleScholarGoogle Scholar |
Cook, R., Gawne, B., Pertie, R., Baldwin, D. S., Rees, G. N., Nielsen, D. L., and Ning, N. S. P. (2015). River metabolism and carbon dynamics in response to flooding in a lowland river. Marine and Freshwater Research 66, 919–927.
| River metabolism and carbon dynamics in response to flooding in a lowland river.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC2MXhsFKiu7vP&md5=d7c6f8e2819fe7b6e525cd1e1309e9e0CAS |
CSIRO (2008). Water availability in the Murray–Darling Basin. A report to the Australian Government from the CSIRO Murray–Darling Basin Sustainable Yields Project. (CSIRO: Canberra.) Available at https://publications.csiro.au/rpr/download?pid=legacy:530&dsid=DS1 [Verified 22 February 2016].
Dalzell, B. J., Filley, T. R., and Harbor, J. M. (2007). The role of hydrology in annual organic carbon loads and terrestrial organic matter export from a midwestern agricultural watershed. Geochimica et Cosmochimica Acta 71, 1448–1462.
| The role of hydrology in annual organic carbon loads and terrestrial organic matter export from a midwestern agricultural watershed.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXis1Gjsrc%3D&md5=5fc67ab54482cb500362c9de622cec09CAS |
Delong, M. D., and Thorp, J. H. (2006). Significance of instream autotrophs in trophic dynamics of the Upper Mississippi River. Oecologia 147, 76–85.
| Significance of instream autotrophs in trophic dynamics of the Upper Mississippi River.Crossref | GoogleScholarGoogle Scholar | 16170563PubMed |
Depetris, P. J., and Kemp, S. (1993). Carbon dynamics and sources in the Parana River. Limnology and Oceanography 38, 382–395.
| Carbon dynamics and sources in the Parana River.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK3sXmtV2qsrs%3D&md5=f444be474a3637a313a879d58ef19cc6CAS |
Dexter, B. D. (1978). Silviculture of the river red gum forests of the central Murray floodplain. Proceedings of the Royal Society of Victoria 90, 175–191.
Doody, T. M., Colloff, M. J., Davies, M., Koul, V., Benyon, R. G., and Nagler, P. L. (2015). Quantifying water requirements of riparian river red gum (Eucalyptus camaldulensis) in the Murray–Darling Basin, Australia: implications for environmental water allocations. Ecohydrology 8, 1471–1487.
| Quantifying water requirements of riparian river red gum (Eucalyptus camaldulensis) in the Murray–Darling Basin, Australia: implications for environmental water allocations.Crossref | GoogleScholarGoogle Scholar |
Gippel, C. J. (1995). Environmental hydraulics of large woody debris in streams and rivers. Journal of Environmental Engineering 121, 388–395.
| Environmental hydraulics of large woody debris in streams and rivers.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK2MXltFGqtb4%3D&md5=2299cb804ad3d6f9b2e0ff66a7e92c57CAS |
Glazebrook, H. S., and Robertson, A. I. (1999). The effect of flooding and flood timing on leaf litter breakdown rates and nutrient dynamics in a river red gum (Eucalyptus camaldulensis) forest. Australian Journal of Ecology 24, 625–635.
| The effect of flooding and flood timing on leaf litter breakdown rates and nutrient dynamics in a river red gum (Eucalyptus camaldulensis) forest.Crossref | GoogleScholarGoogle Scholar |
Growns, I., and Reinfelds, I. (2014). Environmental flow management using transparency and translucency rules. Marine and Freshwater Research 65, 667–673.
| Environmental flow management using transparency and translucency rules.Crossref | GoogleScholarGoogle Scholar |
Hadwen, W. L., Fellows, C. S., Westhorpe, D. P., Rees, G., Mitrovic, S. M., Taylor, B., Baldwin, D. S., Silvester, E., and Croome, R. (2010). Longitudinal trends in river functioning: patterns of nutrient and carbon processing in three Australian rivers. River Research and Applications 26, 1129–1152.
| Longitudinal trends in river functioning: patterns of nutrient and carbon processing in three Australian rivers.Crossref | GoogleScholarGoogle Scholar |
Hitchcock, J. N., and Mitrovic, S. M. (2015). After the flood: changing dissolved organic carbon bioavailability and bacterial growth following inflows to estuaries. Biogeochemistry 124, 219–233.
| After the flood: changing dissolved organic carbon bioavailability and bacterial growth following inflows to estuaries.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC2MXlsV2msr8%3D&md5=c12f75aa1e19c0267f91ee0b744c16d1CAS |
Hitchcock, J., Mitrovic, S. M., Kobayashi, Y., and Westhorpe, D. P. (2010). Responses of estuarine bacterioplankton, phytoplankton and zooplankton to in-situ dissolved organic carbon and inorganic nutrient additions. Estuaries and Coasts 33, 78–91.
| Responses of estuarine bacterioplankton, phytoplankton and zooplankton to in-situ dissolved organic carbon and inorganic nutrient additions.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXotVOisw%3D%3D&md5=f775e4b6b9a0c8f812319368c7989de8CAS |
Hitchcock, J. H., Mitrovic, S. M., Hadwen, W., Roelke, D., and Rohlfs, A. (2015). Terrestrial dissolved organic carbon subsidizes estuarine zooplankton: an in-situ mesocosm study. Limnology and Oceanography 61, 254–267.
| Terrestrial dissolved organic carbon subsidizes estuarine zooplankton: an in-situ mesocosm study.Crossref | GoogleScholarGoogle Scholar |
Hladyz, S., Watkins, S., Whitworth, K., and Baldwin, D. S. (2011). Flows and hypoxic blackwater events in managed ephemeral streams. Journal of Hydrology 401, 117–125.
| Flows and hypoxic blackwater events in managed ephemeral streams.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXktF2hs7Y%3D&md5=60dfb3e3e563636007dd4b037855f3f9CAS |
Hladyz, S., Nielsen, D. L., Suter, P. J., and Krull, E. S. (2012). Temporal variations in organic carbon utilization by consumers in a lowland river. River Research and Applications 28, 513–528.
| Temporal variations in organic carbon utilization by consumers in a lowland river.Crossref | GoogleScholarGoogle Scholar |
Howitt, J., Baldwin, D. S., Rees, G. N., and Williams, J. (2007). Modelling blackwater: predicting water quality during flooding of lowland river forests. Ecological Modelling 203, 229–242.
| Modelling blackwater: predicting water quality during flooding of lowland river forests.Crossref | GoogleScholarGoogle Scholar |
Huryn, A. D., Riley, R. H., Young, R. G., Arbuckle, C. J., Peacock, K., and Lyons, G. (2001). temporal shift in the contribution of organic matter to consumer production in a grassland river. Freshwater Biology 46, 213–226.
| temporal shift in the contribution of organic matter to consumer production in a grassland river.Crossref | GoogleScholarGoogle Scholar |
Jansson, M., Bergström, A., Blomqvist, P., and Drakare, S. (2000). Allochthonous organic carbon and phytoplankton/bacterioplankton production relationships in lakes. Ecology 81, 3250–3255.
| Allochthonous organic carbon and phytoplankton/bacterioplankton production relationships in lakes.Crossref | GoogleScholarGoogle Scholar |
Jardine, T. D., Pusey, B. J., Hamilton, S. K., Pettit, N. E., Davies, P. M., Douglas, M. M., Sinnamon, V., Halliday, I. A., and Bunn, S. E. (2012). Fish mediate high food web connectivity in the lower reaches of a tropical floodplain river. Oecologia 168, 829–838.
| Fish mediate high food web connectivity in the lower reaches of a tropical floodplain river.Crossref | GoogleScholarGoogle Scholar | 21983712PubMed |
Jenerette, G. D., and Lal, R. (2005). Hydrologic sources of carbon cycling uncertainty throughout the terrestrial–aquatic continuum. Global Change Biology 11, 1873–1882.
| Hydrologic sources of carbon cycling uncertainty throughout the terrestrial–aquatic continuum.Crossref | GoogleScholarGoogle Scholar |
Junk, W. J., Bayley, P. B., and Sparks, R. E. (1989). The flood pulse concept in river-floodplain systems. Canadian Special Publication of Fisheries and Aquatic Sciences 106, 110–127.
Karlsson, J., Lymer, D., Vrede, K., and Jansson, M. (2007). Differences in efficiency of carbon transfer from dissolved organic carbon to two zooplankton groups: an enclosure experiment in an oligotrophic lake. Aquatic Sciences 69, 108–114.
| Differences in efficiency of carbon transfer from dissolved organic carbon to two zooplankton groups: an enclosure experiment in an oligotrophic lake.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXlsVCgu78%3D&md5=1af1736354407f3a9fe4efbba54bb435CAS |
Kesel, R. H. (2003). Human modifications to the sediment regime of the Lower Mississippi River flood plain. Geomorphology 56, 325–334.
| Human modifications to the sediment regime of the Lower Mississippi River flood plain.Crossref | GoogleScholarGoogle Scholar |
Kingsford, R. T. (2000). Ecological impacts of dams, water diversions and river management on floodplain wetlands in Australia. Austral Ecology 25, 109–127.
| Ecological impacts of dams, water diversions and river management on floodplain wetlands in Australia.Crossref | GoogleScholarGoogle Scholar |
Kingsford, R. T., and Auld, K. M. (2005). Waterbird breeding and environmental flow management in the Macquarie Marshes, arid Australia. River Research and Applications 21, 187–200.
| Waterbird breeding and environmental flow management in the Macquarie Marshes, arid Australia.Crossref | GoogleScholarGoogle Scholar |
Lavorel, S., Colloff, M. J., McIntyre, S., Doherty, M. D., Murphy, H. T., Metcalfe, D. J., Dunlop, M., Williams, R. J., Wise, R. M., and Williams, K. J. (2015). Ecological mechanisms underpinning climate adaptation services. Global Change Biology 21, 12–31.
| Ecological mechanisms underpinning climate adaptation services.Crossref | GoogleScholarGoogle Scholar | 25131443PubMed |
Lester, R. E., Webster, I. T., Fairweather, P. G., and Young, W. J. (2011). Linking water-resource models to ecosystem-response models to guide water-resource planning; an example from the Murray–Darling Basin, Australia. Marine and Freshwater Research 62, 279–289.
| Linking water-resource models to ecosystem-response models to guide water-resource planning; an example from the Murray–Darling Basin, Australia.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXjsVKktr8%3D&md5=c2bdc438cc1ffd9e51f03618ec65b81fCAS |
Mac Nally, R., Parkinson, A., Horrocks, G., and Young, M. (2002). Current loads of coarse woody debris on southeastern Australian floodplains: evaluation of change and implications for restoration. Restoration Ecology 10, 627–635.
| Current loads of coarse woody debris on southeastern Australian floodplains: evaluation of change and implications for restoration.Crossref | GoogleScholarGoogle Scholar |
Mac Nally, R., Cunningham, S. C., Baker, P. J., Horner, G. J., and Thompson, J. R. (2011). Dynamics of Murray–Darling floodplain forests under multiple stressors: the past, present, and future of an Australian icon. Water Resources Research 47, W00G05.
| Dynamics of Murray–Darling floodplain forests under multiple stressors: the past, present, and future of an Australian icon.Crossref | GoogleScholarGoogle Scholar |
McGinness, H. M., and Arthur, A. D. (2011). Carbon dynamics during flood events in a lowland river: the importance of anabranches. Freshwater Biology 56, 1593–1605.
| Carbon dynamics during flood events in a lowland river: the importance of anabranches.Crossref | GoogleScholarGoogle Scholar |
Mitrovic, S. M., Chessman, B. C., Davie, A., Ryan, N., and Avery, E. (2008). Development of riverine diatom blooms of Cyclotella and Nitzschia and estimating effective suppression flows. Hydrobiologia 596, 173–185.
| Development of riverine diatom blooms of Cyclotella and Nitzschia and estimating effective suppression flows.Crossref | GoogleScholarGoogle Scholar |
Mitrovic, S. M., Hardwick, L., and Dorani, F. (2011). Use of flow management to mitigate cyanobacterial blooms in the Lower Darling River. Journal of Plankton Research 33, 229–241.
| Use of flow management to mitigate cyanobacterial blooms in the Lower Darling River.Crossref | GoogleScholarGoogle Scholar |
Mitrovic, S. M., Westhorpe, D. P., Kobayashi, T., Baldwin, D. S., Ryan, D., and Hitchcock, J. N. (2014). Short-term changes in zooplankton density and community structure in response to different sources of dissolved organic carbon in an unconstrained lowland river: evidence for food web support. Journal of Plankton Research 36, 1488–1500.
| Short-term changes in zooplankton density and community structure in response to different sources of dissolved organic carbon in an unconstrained lowland river: evidence for food web support.Crossref | GoogleScholarGoogle Scholar |
Nielsen, D. L., Cook, R. A., Ning, N., Gawne, B., and Petrie, R. (2015). Carbon and nutrient subsidies to a lowland river following floodplain inundation. Marine and Freshwater Research 67, 1302–1312.
| Carbon and nutrient subsidies to a lowland river following floodplain inundation.Crossref | GoogleScholarGoogle Scholar |
Nilsson, C., Reidy, C. A., Dynesius, M., and Revenga, C. (2005). Fragmentation and flow regulation of the world’s large river systems. Science 308, 405–408.
| Fragmentation and flow regulation of the world’s large river systems.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXjtFOnt7g%3D&md5=f1a8af3a1903c9a3cce664f73a08a44dCAS | 15831757PubMed |
Oliver, R. L., and Merrick, C. J. (2006). Partitioning of river metabolism identifies phytoplankton as a major contributor in the regulated Murray River (Australia). Freshwater Biology 51, 1131–1148.
| Partitioning of river metabolism identifies phytoplankton as a major contributor in the regulated Murray River (Australia).Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XmvVymsrg%3D&md5=20700108772759d9e31af657c75838a7CAS |
Opperman, J. J., Galloway, G. E., Fargione, J., Mount, J. F., Richter, B. D., and Secchi, S. (2009). Sustainable floodplains through large-scale reconnection to rivers. Science 326, 1487–1488.
| Sustainable floodplains through large-scale reconnection to rivers.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXhsFyit77F&md5=0250752f0f5c7e99b16fc78d905861e6CAS | 20007887PubMed |
Opperman, J. J., Luster, R., McKenney, B. A., Roberts, M., and Meadows, A. W. (2010). Ecologically functional floodplains: connectivity, flow regime, and scale. Journal of the American Water Resources Association 46, 211–226.
| Ecologically functional floodplains: connectivity, flow regime, and scale.Crossref | GoogleScholarGoogle Scholar |
Phillips, P. (1972). ‘River Boat Days on the Murray, Darling and Murrumbidgee.’ (Landsdowne Press: Melbourne.)
Polis, G. A., Anderson, W. B., and Holt, R. D. (1997). Toward an integration of landscape ecology and food web ecology: the dynamics of spatially subsidized food webs. Annual Review of Ecology and Systematics 28, 289–316.
| Toward an integration of landscape ecology and food web ecology: the dynamics of spatially subsidized food webs.Crossref | GoogleScholarGoogle Scholar |
Powell, J. M. (1989). ‘Watering the Garden State: Land and Community in Victoria 1834–1988.’ (Allen and Unwin: Sydney.)
Powell, J. M. (1993). ‘The Emergence of Bioregionalism in the Murray–Darling Basin.’ (Murray–Darling Basin Commission: Canberra.)
Price, J. N., Wong, N. K., and Morgan, J. W. (2010). Recovery of understorey vegetation after release from a long history of sheep grazing in a herb-rich woodland. Austral Ecology 35, 505–514.
| Recovery of understorey vegetation after release from a long history of sheep grazing in a herb-rich woodland.Crossref | GoogleScholarGoogle Scholar |
Reid, D. J., Quinn, G. P., Lake, P. S., and Reich, P. (2008). Terrestrial detritus supports the food webs in lowland intermittent streams of south‐eastern Australia: a stable isotope study. Freshwater Biology 53, 2036–2050.
| Terrestrial detritus supports the food webs in lowland intermittent streams of south‐eastern Australia: a stable isotope study.Crossref | GoogleScholarGoogle Scholar |
Roach, K. A. (2013). Environmental factors affecting incorporation of terrestrial material into large river food webs. Freshwater Science 32, 283–298.
| Environmental factors affecting incorporation of terrestrial material into large river food webs.Crossref | GoogleScholarGoogle Scholar |
Robertson, A. I., and Rowling, R. W. (2000). Effects of livestock on riparian zone vegetation in an Australian dryland river. Regulated Rivers: Research and Management 16, 527–541.
| Effects of livestock on riparian zone vegetation in an Australian dryland river.Crossref | GoogleScholarGoogle Scholar |
Robertson, A. I., Bunn, S. E., Boon, P. I., and Walker, K. F. (1999). Sources, sinks and transformations of organic carbon in Australian floodplain rivers. Marine and Freshwater Research 50, 813–829.
| Sources, sinks and transformations of organic carbon in Australian floodplain rivers.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3cXks1ymug%3D%3D&md5=2a8a03d8287ce412f748d360e345d1f6CAS |
Rohlfs, A., Mitrovic, S. M., Williams, S., and Coleman, D. (2015). Can tributary in-flows improve DOC regime recovery in a snowmelt river regulated by a large reservoir? Marine and Freshwater Research 67, 1338–1345.
| Can tributary in-flows improve DOC regime recovery in a snowmelt river regulated by a large reservoir?Crossref | GoogleScholarGoogle Scholar |
Roseman, E. F., and DeBruyne, R. L. (2015). The renaissance of ecosystem integrity in North American large rivers. Restoration Ecology 23, 43–45.
| The renaissance of ecosystem integrity in North American large rivers.Crossref | GoogleScholarGoogle Scholar |
Saikia, S., and Nandi, S. (2010). C and P in aquatic food chain: a review on C : P stoichiometry and PUFA regulation. Knowledge & Management of Aquatic Ecosystems 398, 03.
| C and P in aquatic food chain: a review on C : P stoichiometry and PUFA regulation.Crossref | GoogleScholarGoogle Scholar |
Sims, N. C., Chariton, A., Jin, H., and Colloff, M. J. (2012). A classification of floodplain ecosystems in the Murray–Darling Basin based on changes in flows following water resource development. Wetlands 32, 239–248.
| A classification of floodplain ecosystems in the Murray–Darling Basin based on changes in flows following water resource development.Crossref | GoogleScholarGoogle Scholar |
Stanley, E. H., Powers, S. M., Lottig, N. R., Buffam, I., and Crawford, J. T. (2012). Contemporary changes in dissolved organic carbon (DOC) in human-dominated rivers: is there a role for DOC management? Freshwater Biology 57, 26–42.
| Contemporary changes in dissolved organic carbon (DOC) in human-dominated rivers: is there a role for DOC management?Crossref | GoogleScholarGoogle Scholar |
Tanentzap, A. J., Szkokan-Emilson, E. J., Kielstra, B. W., Arts, M. T., Yan, N. D., and Gunn, J. M. (2014). Forests fuel fish growth in freshwater deltas. Nature Communications 5, 4077.
| Forests fuel fish growth in freshwater deltas.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC2cXhvF2mu7%2FF&md5=c65553c118134b6ebd181510dc04af47CAS | 24915965PubMed |
Thorp, J. H., and Delong, M. D. (2002). Dominance of autochthonous autotrophic carbon in food webs of heterotrophic rivers. Oikos 96, 543–550.
| Dominance of autochthonous autotrophic carbon in food webs of heterotrophic rivers.Crossref | GoogleScholarGoogle Scholar |
Thorp, J. H., Delong, M. D., Greenwood, K. S., and Casper, A. F. (1998). Isotopic analysis of three food web theories in constricted and floodplain regions of a large river. Oecologia 117, 551–563.
| Isotopic analysis of three food web theories in constricted and floodplain regions of a large river.Crossref | GoogleScholarGoogle Scholar |
Tobin, G. A. (1995). The levee love affair: a stormy relationship? Journal of the American Water Resources Association 31, 359–367.
| The levee love affair: a stormy relationship?Crossref | GoogleScholarGoogle Scholar |
Tockner, K., Pennetzdorfer, D., Reiner, N., Schiemer, F., and Ward, J. V. (1999). Hydrological connectivity, and the exchange of organic matter and nutrients in a dynamic river–floodplain system (Danube, Austria). Freshwater Biology 41, 521–535.
| Hydrological connectivity, and the exchange of organic matter and nutrients in a dynamic river–floodplain system (Danube, Austria).Crossref | GoogleScholarGoogle Scholar |
Treadwell, S., Koehn, J., and Bunn, S. (1999) Large woody debris and other aquatic habitat. In ‘Riparian Land Management Technical Guidelines Volume One: Principles of Sound Management’. (Eds S. Lovett and P. Price.) pp. 79–96. (Land and Water Resources Research and Development Corporation: Canberra.)
Vink, S., Bormans, M., Ford, P. W., and Grigg, N. J. (2005). Quantifying ecosystem metabolism in the middle reaches of Murrumbidgee River during irrigation flow releases. Marine and Freshwater Research 56, 227–241.
| Quantifying ecosystem metabolism in the middle reaches of Murrumbidgee River during irrigation flow releases.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXjtlWrsrc%3D&md5=5957bb6f1b07f2c680c3a98aecd5209dCAS |
Vivian, L., Godfree, R. C., Colloff, M. J., Mayence, C. E., and Marshall, D. J. (2014). Wetland plant growth under contrasting water regimes associated with river regulation and drought: implications for environmental water management. Plant Ecology 215, 997–1011.
| Wetland plant growth under contrasting water regimes associated with river regulation and drought: implications for environmental water management.Crossref | GoogleScholarGoogle Scholar |
Vörösmarty, C. J., McIntyre, P. B., Gessner, M. O., Dudgeon, D., Prusevitch, A., Green, S., Glidden, S., Bunn, S. E., Sullivan, C. A., Reidy Liermann, C., and Davies, P. M. (2010). Global threats to human water security and river biodiversity. Nature 467, 555–561.
| Global threats to human water security and river biodiversity.Crossref | GoogleScholarGoogle Scholar | 20882010PubMed |
Walker, K. F., and Thoms, M. C. (1993). Environmental effects of flow regulation on the Lower River Murray, Australia. Regulated Rivers: Research and Management 8, 103–119.
| Environmental effects of flow regulation on the Lower River Murray, Australia.Crossref | GoogleScholarGoogle Scholar |
Walker, J., Bullen, F., and Williams, B. G. (1993). Ecohydrological changes in the Murray–Darling Basin. I. The number of trees cleared over two centuries. Journal of Applied Ecology 30, 265–273.
| Ecohydrological changes in the Murray–Darling Basin. I. The number of trees cleared over two centuries.Crossref | GoogleScholarGoogle Scholar |
Wallace, T. A., Ganff, G. G., and Brookes, J. D. (2008). A comparison of phosphorus and DOC leachates from different type of leaf litter in an urban environment. Freshwater Biology 53, 1902–1913.
| A comparison of phosphorus and DOC leachates from different type of leaf litter in an urban environment.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXhtFKisLnF&md5=829478cfd2abcacaf88a89667a1cec8aCAS |
Wantzen, K. M., de Arruda Machado, F., Voss, M., Boriss, H., and Junk, W. J. (2002). Seasonal isotopic shifts in fish of the Pantanal wetland, Brazil. Aquatic Sciences 64, 239–251.
| Seasonal isotopic shifts in fish of the Pantanal wetland, Brazil.Crossref | GoogleScholarGoogle Scholar |
Watkins, S. C., Quinn, G. P., and Gawne, B. (2010). Changes in organic-matter dynamics and physiochemistry, associated with riparian vegetation loss and river regulation in floodplain wetlands of the Murray River, Australia. Marine and Freshwater Research 61, 1207–1217.
| Changes in organic-matter dynamics and physiochemistry, associated with riparian vegetation loss and river regulation in floodplain wetlands of the Murray River, Australia.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXht1yjsrjO&md5=0da00ec4d8b611037146e8e47c06854fCAS |
Westhorpe, D. P., and Mitrovic, S. M. (2012). Dissolved organic carbon mobilisation in relation to variable discharges in a highly regulated lowland river. Marine and Freshwater Research 63, 1218–1230.
| Dissolved organic carbon mobilisation in relation to variable discharges in a highly regulated lowland river.Crossref | GoogleScholarGoogle Scholar |
Westhorpe, D. P., Mitrovic, S. M., and Kobayashi, Y. (2010). Limitation of lowland riverine bacterioplankton by dissolved organic carbon and inorganic nutrients. Hydrobiologia 652, 101–117.
| Limitation of lowland riverine bacterioplankton by dissolved organic carbon and inorganic nutrients.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXovV2jsbo%3D&md5=3324c65fb6567eb3ad97ea40b46eb147CAS |
Whitworth, K. L., and Baldwin, D. S. (2016). Improving our capacity to manage hypoxic blackwater events in lowland rivers: the blackwater risk assessment tool. Ecological Modelling 320, 292–298.
| Improving our capacity to manage hypoxic blackwater events in lowland rivers: the blackwater risk assessment tool.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC2MXhslyntr%2FM&md5=11a2a8d026f4040d9830e9bfd7e9768dCAS |
Whitworth, K. L., Baldwin, D. S., and Kerr, J. (2012). Drought, floods and water quality: drivers of a severe hypoxic blackwater event in a major river system (the southern Murray–Darling Basin, Australia). Journal of Hydrology 450–451, 190–198.
| Drought, floods and water quality: drivers of a severe hypoxic blackwater event in a major river system (the southern Murray–Darling Basin, Australia).Crossref | GoogleScholarGoogle Scholar |
Winemiller, K. O. (2004) Floodplain river food webs: generalizations and implications for fisheries management. In ‘Proceedings of the Second International Symposium on the Management of Large Rivers for Fisheries: Sustaining Livelihoods and Biodiversity in the New Millennium, Vol. 1’. (Eds R. L. Welcomme and T. Petr.) pp. 285–309. (Food and Agriculture Organization of the United Nations and the Mekong River Commission: Bangkok.)
World Commission on Dams (2000) Dams and Development: a New Framework for Decision-making. Earthscan, London. Available at http://hqweb.unep.org/dams/WCD/report/WCD_DAMS%20report.pdf [Verified 22 February 2012.]
Worster, D. (1985) ‘Rivers of Empire: Water, Aridity and the Growth of the American West.’ (Pantheon: New York.)
Zedler, J. B., and Kercher, S. (2005). Wetland resources: status, trends, ecosystem services, and restorability. Annual Review of Environment and Resources 30, 39–74.
| Wetland resources: status, trends, ecosystem services, and restorability.Crossref | GoogleScholarGoogle Scholar |
Zeug, S. C., and Winemiller, K. O. (2008). Evidence supporting the importance of terrestrial carbon in a large-river food web. Ecology 89, 1733–1743.
| Evidence supporting the importance of terrestrial carbon in a large-river food web.Crossref | GoogleScholarGoogle Scholar | 18589537PubMed |