

Life-history traits and the mating system of an Australian water strider, Tenagogerris euphrosyne
Paul G. R. Colvin A , Mark A. Elgar A and Therésa M. Jones A BA Department of Zoology, The University of Melbourne, Parkville, Vic. 3010, Australia.
B Corresponding author. Email: theresa@unimelb.edu.au
Australian Journal of Zoology 54(2) 107-116 https://doi.org/10.1071/ZO05058
Submitted: 14 September 2005 Accepted: 15 March 2006 Published: 11 May 2006
Abstract
The hemipteran infraorder Gerromorpha (the semiaquatic bugs, or water striders) has been used extensively as a model taxon in evolutionary biology and ecology. Most studies have focused on North American and European species while there is a paucity of comparable data on the gerromorphs of Africa and Australia. In a series of laboratory experiments we explore patterns of ontogenetic development, foraging and mating behaviour in Tenagogerris euphrosyne (Kirkaldy), a medium-sized water strider distributed along the east coast of Australia. Our study revealed that T. euphrosyne nymphs passed through five instars before adulthood, but that mortality and rates of cannibalism were high. Foraging and mating trials revealed that the foraging success of adult males (but not females) was positively correlated with mating success. Observations indicated that T. euphrosyne exhibits a Type I water strider mating behaviour: mating followed a brief period of struggling by the female, after which males were able to ride on a female’s back for extended periods. Females gained direct fertility benefits from mating and were able to store sperm and lay fertilised eggs (albeit a reduced quantity) for several weeks even in the absence of males.
Introduction
Species belonging to the highly diverse hemipteran infraorder Gerromorpha (the semiaquatic bugs, or water striders) are adapted to life on the water surface, and have developed a wide range of morphologies and behaviours that make them evolutionarily fascinating. The two-dimensional environment that they inhabit also renders them an extremely tractable taxon for collection and study. It is perhaps unsurprising then that the phylogeny of gerromorphs is well resolved (Andersen 1982, 1997; Damgaard et al. 2005) and that water striders have been used as model organisms in a variety of contexts, including locomotary function (Bondarenko 1994; Denny 2004), ontogenetic development (Harada and Spence 2000), morphological polymorphism (Fairbairn and Preziosi 1996; Nummelin 1997), foraging behaviour (Blanckenhorn 1991; Ortigosa and Rowe 2002), mate attraction (Wilcox 1995) and, most recently, the evolution and maintenance of sexual conflict (Rowe et al. 1994; Arnqvist and Rowe 1995, 2002; Arnqvist 1997; Rowe and Arnqvist 2002). However, while water striders are almost ubiquitous in their distribution worldwide, most studies exploring variation in behavioural and morphological traits or life-history parameters have focussed on the temperate species of North America and Europe; to date there are an overwhelming lack of comparable data on the tropical species of Africa and Australia (Spence and Andersen 1994; Andersen and Weir 2004a).
Australia is home to 129 species of waterstriders, comprising 30 genera and six families (Andersen and Weir 2004a), and their phylogeny is well documented (Andersen and Weir 1997, 2004a, 2004b). In contrast, studies of the ecology, life-history traits or mating strategies of most of these species are sparse or, more usually, completely absent (Andersen and Weir 2004a). An important reason for addressing this regional disparity in research effort is that taxonomic evidence and limited field observations suggest that Australian species have exciting and novel evolutionary histories (Andersen and Weir 2004a). For example, a recent study of the Australian Zeus bug Phoreticovelia disparata, one of the smaller semiaquatic bugs (females ~3 mm; males ~1 mm), revealed sex-role-reversed nuptial feeding, a mating strategy that is not only unique in the animal kingdom, but also challenges a core paradigm in evolutionary theory (Arnqvist et al. 2003).
The genus Tenagogerris comprises three water strider species unique to Australia (T. femoratus, T. euphrosyne, T. pallidus). T. euphrosyne (Kirkaldy) is distributed along the east coast of Australia, inhabiting a variety of freshwater habitats such as lakes, billabongs and slow-flowing streams and rivers where it skates on the water surface, foraging mainly on dead and decaying insects (Andersen and Weir 2004a). In Victoria, adults are scarce between May and October (the colder winter months), suggesting that, in this region, individuals may enter reproductive diapause; in contrast, adults are abundant throughout the year in the warmer regions of north-eastern Australia (Andersen and Weir 2004a). Field-collected specimens suggest that T. euphrosyne has several morphological features typical of water striders. It is wing-dimorphic, although the frequency of the dispersing winged (macropterous) form is far lower than the non-dispersing non-winged (apterous) form. Males are typically smaller than females (body length of males = 6.8–8.9 mm; females = 8.4–10.0 mm) and are frequently observed riding on the backs of females. Although T. euphrosyne is the most common water strider species in south-east Australia, surprisingly little is known about its reproductive biology (Andersen and Weir 1997, 2004a). Anecdotal field observations suggest that males ride on the backs of females for unusually long periods following copulation (Andersen and Weir 1997, 2004a), a feature common in water striders (Arnqvist 1997); however, the occurrence and consequences of this mating behaviour have not been quantified. Moreover, estimates of life-history traits such as fecundity or fertilisation and the degree and causes of mortality are absent. Indeed, the mating system is not documented.
Here, we present a series of laboratory experiments that provide the first data on patterns of ontogenetic development and mating behaviour in T. euphrosyne. We examine specifically: (i) the impact of the foraging environment on egg–adult survival, including estimates of intermoult intervals and incidences of cannibalism; (ii) the rate of intraspecific competition for food and mates; (iii) the relationship between the operational sex ratio and male riding behaviour; and (iv) the effect of mating opportunity on female fecundity and fertilisation success.
Materials and Methods
We collected adults and nymphs of Tenagogerris euphrosyne from the Goulburn River, in northern Victoria, Australia (grid reference 36.24S, 145.22E) between October 2003 and January 2004. Individuals were transported to the laboratory in collection vials, containing moist paper towel, which were placed in a cooler with ice packs. They were maintained in the laboratory in plastic rearing containers (30 cm by 24 cm by 13 cm deep) with aerated water (2 cm depth). A polystyrene foam strip (10 cm by 8 cm by 1 cm) acted as a resting and oviposition substrate. Blowflies (Lucilia cuprina) and Drosophila were provided ad libitum and the containers were cleaned weekly. Individuals were sexed (following Andersen and Weir 1997), uniquely marked on their thorax using enamel paints (Humbrol Super Enamel) and maintained in single-sex cultures with no more than 15 individuals per container.
Experimental adults were weighed (to the nearest 0.00001 g) immediately before a trial. At the end of the experimental period, a digital photograph was taken of the body and forefemur using a Moticam 480 digital camera (Motic®) attached to a binocular microscope (×20 magnification). Body and forefemur lengths were then measured (to the nearest 0.01 mm) from these photographs using the Motic Images 2000 imaging software (Motic®). Lengths were highly repeatable (body length intraclass correlation: r = 0.98, F1,14 = 0.24, P = 0.63; forefemur length: r = 0.99, F1,14 = 2.30, P = 0.15). As body length and weight were correlated for both males and females (females: t = 4.95, n = 36, r = 0.63, P < 0.0001; males: t = 5.65, n = 37, r = 0.68, P < 0.0001) body length only was used in the analyses.
Effect of feeding regime on nymph development
The quantity and quality of available food for developing T. euphrosyne nymphs is likely to vary substantially both across and potentially within populations. To explore the effect of variation in food supply on egg to adult survival in T. euphrosyne, a total of 600 fertilised eggs from 50 field-caught females were distributed into 60 replicate batches of ten eggs each. Each egg batch contained eggs from at least two females, and the eggs from any individual female were distributed across replicates and treatments. To minimise variation in hatching date, all eggs contributing to a batch were laid within a 48-h period.
Each batch of eggs was housed in a plastic container under standard rearing conditions (see above) and emerging nymphs were subjected to one of four treatments, which varied the feeding regime. For each feeding regime, food was introduced into a container, left for 60 h and then any unconsumed food removed for a 24-h period. This procedure ensured that individuals experienced some period of starvation. The process was repeated until all individuals had either moulted to an adult or died. On any given feeding event, each surviving nymph within a tub was provided either: (i) one D. melanogaster (Single Drosophila); (ii) one blowfly, L. cuprina (Lucilia); (iii) one D. melanogaster and one L. cuprina (Mixed); or (iv) D. melanogaster equivalent in weight to an average blowfly, L. cuprina (0.017 ± 0.001 g) (Mass Drosophila). The two prey types varied in size and percentage chitin: average percentage chitin (expressed as a percentage of dry weight) of L. cuprina was 12.62 ± 0.03% (n = 5 flies), and of D. melanogaster was 54.14 ± 0.01% (n = 5 flies). Accordingly, it was assumed that the four feeding regimes varied in the quantity and/or quality of prey types available.
Each container was assessed every 2 days and the number of individuals that had moulted or died was recorded. We noted whether a nymph died during a moult and whether it had been cannibalised following death. This provides an approximate estimate of the degree of cannibalism; however, our design does not permit us to determine whether cannibalism was the cause of death. The development time, from the introduction of fertilised eggs to the emergence of nymphs, was calculated as the mean number of days to hatching for all eggs that hatched. The time taken between subsequent moults was calculated similarly, using the average date of the previous moult as the baseline start-date. The total development time from egg to adult was calculated as the sum of days between each of the five moults plus 5 days (the average age of visibly fertilised eggs used in the experiment). While these values provide a conservative measure of development times, it was not possible to obtain more accurate data by marking individual nymphs.
Intraspecific competition for food and mates
Intraspecific competition for food and mates is likely to be intense in any species, and variation in an individual’s ability in these two behaviours is likely to have far-reaching consequences for lifetime fitness. For male water striders, the ability to secure food items or acquire females relies on a similar grasping action of the forelegs. In contrast, female water striders utilise their forelegs for grasping prey, but do not employ them during the mating process. As a consequence, foraging ability and mate acquisition may be correlated for male (see Blanckenhorn et al. 1998), but not female, water striders.
To assess the potential skew in foraging and mating success across T. euphrosyne adults and to explore the relationship between foraging and mating potential for both sexes, we assigned 100 field-caught adults into 20 same-sex groups consisting of either five males (n = 10 groups) or five females (n = 10 groups). Each group was maintained under standard rearing conditions (see above) and fed with ad libitum blowflies twice weekly. We assessed the foraging ability and potential reproductive success of individuals within a group by allowing them to compete for access to food and then mates.
Foraging trials
The foraging activity of water striders was enhanced by removing all food from each container before an experimental trial. For each trial, a single dead blowfly was placed in the container and we recorded the identity and time taken for the first individual to grasp the prey with its forelegs and/or extend its proboscis towards the piece of food. The prey was replaced if no individual touched it within 10 min, and the trial was restarted. The trial was abandoned and the data discarded if the prey had to be replaced more than three times (males = 1 trial; females = 2 trials).
Following a successful feeding event, the other individuals in the container were given access to prey by placing an additional four blowflies to the container. Individuals were given 1 h to feed, after which time the food was removed. The feeding trial was then repeated 4 h later, thereby allowing individuals in each group to compete for prey on two occasions over a 5-h period. The experimental procedure was repeated for another four days, yielding a total of 10 competitive foraging trials for each group. The data from these trials was used to rank the competitive ability of each individual on a five-point scale according to the number of times it competed successfully for the prey item. Tied individuals were given the mid-rank between their summed successive ranks.
Mate competition trials
We repeated the competition trials (with the same group compositions) 5–10 days later, only this time individuals competed for access to mates rather than food. For each trial, a single individual of the opposite sex was introduced to a container and left for 5 min or until it paired successfully. A pairing was deemed successful if the male sustained a grip on the female that extended beyond the initial precopulatory struggle. Successfully paired individuals were separated to prevent copulation. A new trial commenced 30 min later, and the process repeated until up to 10 pairings were observed per group.
We recorded the identity of the pairing individual and the time taken for the initiation of pairing, and the duration of the initial struggle. The data from these trials were used to rank the competitive ability of each individual on a five-point scale according to the number of times it paired successfully. Tied individuals were given the mid-rank between their summed successive ranks. Data from trials involving individuals that died between the two experimental treatments were discarded.
For foraging and mate competition trials, the four observed frequency distributions (male foraging; male mating; female foraging; female mating) generated by the data were compared to average expected Poisson error distributions. Each Poisson distribution was calculated using the mean number of food items or mates obtained by individuals in each of the 10 trials as the Poisson mean.
Relationship between operational sex ratio and riding behaviour
The distribution of riding events across male and female water striders is commonly influenced by the population operational sex ratio (Clark 1988; Arnqvist 1992; Sih and Krupa 1995). To explore the effect of variation in the operational sex ratio on riding behaviour in T. euphrosyne, a total of 90 males and 90 females reared under standard conditions (see above) from field-caught nymphs were assigned to groups of six individuals and were then subject to one of three treatments that varied the group sex ratio. There were 10 replicates each of male-biased groups (four males and two females), of female-biased groups (two males and four females), and of parity groups (three males and three females). These water striders had been reared in mixed-sex aggregations for 2–3 weeks, during which time females had commenced egg laying and were thus sexually mature.
Each group was maintained in a standard plastic rearing container (see above) and was provided with ad libitum frozen Drosophila, and a 1-cm-thick strip of polystyrene foam (10 × 8 cm). Individuals were observed for 10 min following introduction, and all riding, struggling and aggressive behaviour noted. Groups were subsequently observed every 1–2 days until at least one pair had formed and riding commenced. After this time a container was observed every 3 h between 0900 and 1500 hours, every 1–2 days, for a total of 9 days. The time to initiate pairing and the number and identity of all paired individuals were noted.
Influence of mating opportunity on fecundity and fertilisation success
To explore the consequences of potential mating opportunity on female fecundity and fertilisation success, field-caught females (n = 42) were selected haphazardly from the stock population and placed in an individual container (diameter = 125 mm, height = 120 mm) with ~2-cm-deep water, a foam strip (10 cm × 8 cm × 1 cm), and fed an ad libitum diet of blowflies and Drosophila. The foam strip was replaced every 3–4 days, and the total number of fertilised and unfertilised eggs laid on it was noted. Fertilised eggs have eyespots at 3–5 days old, while unfertilised eggs remain opaque.
All females were subject to the same three-week pre-experimental period, after which time they were allocated randomly to one of three treatments that varied the exposure of females to males: (i) a male was placed in the female’s container and then removed after 3 days (0-males); (ii) a male was placed in the female’s container, where he remained for the next 3 weeks (1-male); and (iii) a male was placed in the female’s container for 1 week, after which he was replaced by a new male each week for 3 weeks (3-males).
Statistics
Data were transformed where necessary to achieve normality and then analysed using the model-fitting platform in JMP 4.0.2 (Academic 1989–2000). When normality could not be achieved through transformation, we used the equivalent non-parametric test. Significant differences between groups were compared using Student’s t-tests with Bonferonni correction (Rice 1989). Data are presented as means ± standard error, unless otherwise stated.
Results
Effect of feeding regime on nymph development
Survival
Almost 50% (291 of 574) of the dead nymphs showed some degree of cannibalism; 50 nymphs died during moult and 233 died from undetermined causes (i.e. they displayed no sign of moulting or cannibalism). Cannibalism was equally likely to be observed in the presence and absence of food (F3,112 = 2.62, P = 0.11), but varied across the four treatment groups (mean number of nymphs cannibalised per container in the Single Drosophila treatment = 3.17 ± 0.26; Blowfly = 2.77 ± 0.43; Mass Drosophila = 0.15 ± 0.24; Mixed = 2.67 ± 0.31; F3,112 = 5.07, P = 0.003). Post hoc analyses revealed that nymphs reared on a diet of Mass Drosophila were less likely to be observed cannibalised than nymphs reared on a Blowfly or Single Drosophila diet (all P < 0.05). All other comparisons were non-significant.
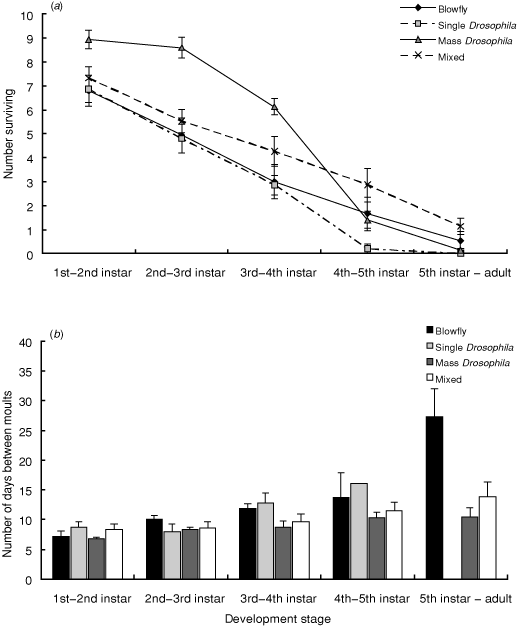
The number of surviving adults varied significantly across the treatment groups (χ32 = 11.92, P = 0.008) (Fig. 1a). Post hoc tests revealed that nymphs reared on the Mixed treatment were more likely to moult into adults than individuals reared on either Single Drosophila (P < 0.01) or Mass Drosophila treatments (P < 0.05). All other comparisons were non-significant.
|
The number of surviving individuals declined progressively with each successive moult (repeated-measures ANOVA: F3,56 = 15.18, P < 0.0001) (Fig. 1a) and varied across the four feeding regimes (F3,56 = 5.37, P = 0.003) (Fig. 1a). There was also a significant interaction between feeding regime and moult number (F12,56 = 4.51, P < 0.0001) (Fig. 1a).
The average total time between eggs to adult varied across the three treatment groups that successfully produced adults (F2,9 = 7.79, P = 0.01). Post hoc analyses revealed that individuals reared on a diet of blowflies alone took longer to mature as adults than individuals reared on Mass Drosophila or a Mixed diet (both P < 0.05).
Time between moults
Across the four treatment groups, the average time between moults (Fig. 1b) was comparable from the egg to 1st instar (F3,56 = 1.39, P = 0.25), 1st to 2nd instar (F3,53 = 1.09, P = 0.36), 2nd to 3rd instar (F3,50 = 2.10, P = 0.11) and 3rd to 4th instar (F3,25 = 0.62, P = 0.61), but varied between the 4th instar and final moult to adult (F2,9 = 5.54, P = 0.03). Post hoc analyses revealed that individuals reared on a diet of blowflies alone took longer to moult from 5th instar nymphs to adults than individuals reared on Mass Drosophila or a Mixed diet (both P < 0.05).
Among the 12 containers that successfully yielded adults, the time between successive moults tended to increase from the 1st to final moult (repeated-measures ANOVA: F2,9 = 3.28, P = 0.09). There was also significant variation across the three treatment groups (F2,9 = 7.79, P = 0.01), but the interaction between treatment group and moult was non-significant (F2,9 = 0.76, P = 0.65).
Intraspecific competition for food and mates
Males: foraging trials
A total of 39 (78%) males gained access to at least one piece of food. Across all trials, the observed distribution of food items obtained by males was not significantly different from an expected random distribution (χ52 = 4.42, n = 50, P = 0.49).
A male’s ranked foraging success was positively correlated with the length of his forefemur (F1,24 = 4.46, β (s.e.) = 2.61 (1.24), P = 0.05), but was unrelated to his body length (F1,24 = 0.93, P = 0.34). The distribution of male foraging ranks was comparable across all 10 trials (F8,31 = 0.33, P = 0.95).
Males: mate competition trials
A total of 30 (60%) males paired with a female at least once. In 41% (26 of 63) of successful pairings, a brief precopulatory struggle was observed. The probability of a male engaging in a precopulatory struggle was unrelated to his rank mating success (χ52 = 4.67, P = 0.46). When they occurred, the duration of a precopulatory struggle was similar for all males regardless of rank (mean duration of struggle = 13.42 ± 0.11 s, range = 5–39 s, F5,43 = 0.49, P = 0.77) and was comparable across all 10 trials (F9,43 = 0.70, P = 0.70).
The observed distribution of successful riding attempts across males did not vary from an expected random distribution (χ52 = 4.90, n = 50, P = 0.42).
A male’s ranked mating success was unrelated to either body length (F1,24 = 2.07, P = 0.16), or forefemur length (F1,24 = 2.98, P = 0.10) and was comparable across all trials (F8,24 = 0.72, P = 0.68).
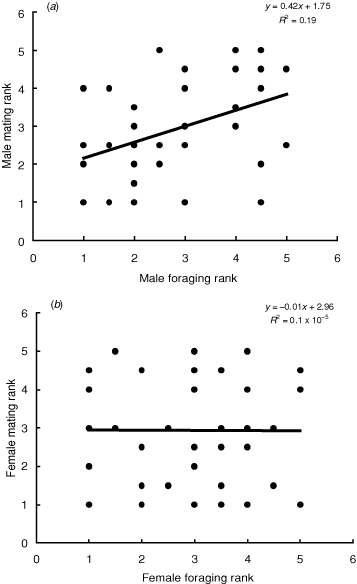
A male’s ranked mating success was positively correlated with his rank foraging success (F1,48 = 11.14, β (s.e.) = 0.42 (0.13), P = 0.002) (Fig. 2a).
|
Females: foraging trials
A total of 44 (88%) females gained access to at least one piece of food. Across all trials, the observed distribution of food items obtained by females was not significantly different from an expected random distribution (χ52 = 1.64, n = 50, P = 0.90).
A female’s ranked foraging success was negatively related to her body length (F1,19 = 8.11, β (s.e.) = –2.23 (0.78), P = 0.01), but unrelated to the length of her forefemur (F1,19 = 1.74, P = 0.20). There was no significant variation in the distribution of female rank mating success across the nine groups of females (F8,19 = 1.22, P = 0.34).
Females: mate competition trials
A total of 31 (76%) females formed a pair at least once. Most successful pairings (57 of 69) involved a precopulatory struggle. However, nine (29%) females who were successful in at least one mate competition trial did not engage in a precopulatory struggle.
The probability of a female engaging in a precopulatory struggle was unrelated to her rank mating success (χ52 = 5.18, P = 0.39). When they occurred, the duration of a precopulatory struggle was similar for all females regardless of rank (mean duration of struggle = 11.52 ± 0.60 s, range = 2–26 s, F4,43 = 1.20, P = 0.33) and was comparable across all 10 trials (F9,39 = 1.31, P = 0.26).
The observed distribution of females with a riding male did not vary from an expected random distribution (χ52 = 4.65, n = 50, P = 0.46).
A female’s ranked mating success was unrelated to either body length (F1,19 = 0.15, P = 0.70), or forefemur length (F1,19 = 2.06, P = 0.17) and was comparable across all trials (F8,19 = 0.44, P = 0.88).
A female’s ranked foraging success was not related to her rank mating success (t = 0.05, r = 0.14, P = 0.96) (Fig. 2b).
Relationship between operational sex ratio and riding behaviour
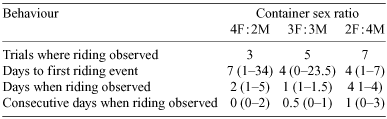
In the larger containers containing six individuals, males were observed riding on the backs of females in 50% (15 of 30) of trials. On average, only one or two riding pairs were observed per container over the 4-week period and the total number of consecutive days riding was relatively short (Table 1). The probability that riding occurred was comparable across the three treatments (χ22 = 3.02, P = 0.22).
|
Our observations suggest that mating behaviour can be divided into three phases. In the first ‘precopulatory phase’, males and females were observed in close proximity to one another, but females moved away from approaching males. In the second ‘riding and copulation phase’, males paired with females and commenced riding. Riding was initiated by the male and typically involved him attempting to grasp the female’s abdomen with his forelimbs. A brief struggle was observed in approximately half of the riding attempts, but females subsequently permitted males to ride without further struggling behaviour. Males rode on the backs of females for extended periods during this phase, but were not observed riding for the entire period. Finally, in the third ‘oviposition phase’, males ceased riding and females commenced egg-laying. During the separation phase, males were never observed riding, but their approach to females often resembled that during the precopulation phase, making it difficult to distinguish between the precopulation and oviposition phases.
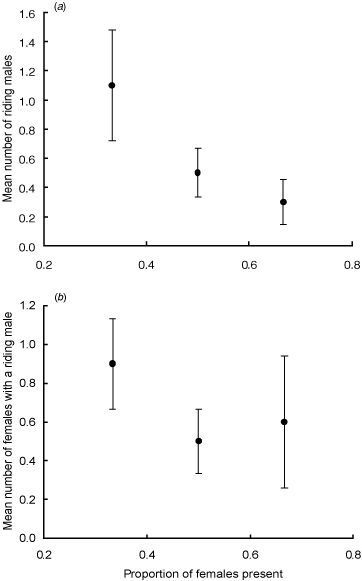
Riding pairs were more likely to occur as the proportion of males in a container increased (Nominal logistic regression: χ12 = 3.02, P = 0.08) (Table 1). The average number of males observed riding within a container was negatively correlated with the proportion of females present (rs = -0.38, P = 0.04) (Fig. 3a). In contrast, there was no relationship between the number of females being ridden and the container sex-ratio (rs = –0.26, P = 0.17) (Fig. 3b). The time taken before the first pairing event was comparable across all sex ratios (rs = 0.20, P = 0.48) (Table 1) and the observed total number of days riding was unrelated to container sex ratio (rs = –0.15, P = 0.60) (Table 1).
|
Influence of mating opportunity on fecundity and fertilisation success
In total, 37 of 42 females laid eggs during the three-week pre-experimental period, and 31 of the 42 females laid eggs during the experimental period. There was no significant effect of experimental period or exposure to males (treatment group) on the probability of a female laying eggs (Nominal logistic model; effect of experimental period: χ12 = 2.69, P = 0.10; treatment group: χ22 = 0.83, P = 0.66). The number of eggs laid tended to be related to female weight (t = 1.74, n = 37, r = 0.22, P = 0.09).
The total number of eggs laid was comparable for the three treatment groups (F2,38 = 1.75, P = 0.19). On average, females laid fewer eggs during the pre-experimental period than the experimental period (mean total number of pre-experimental eggs laid = 20.9 ± 2.77; experimental eggs = 32.12 ± 4.91; t = 2.14, P = 0.02).
The number of eggs laid varied over the 6-week period (repeated-measures ANOVA: F5,34 = 5.49, P = 0.0008) (Fig. 4a). The number of eggs declined over the pre-experimental period, increased following introduction of a male at Week 4 and then declined once more over the experimental period. This overall pattern was consistent across the three treatment groups (F2,38 = 0.68, P = 0.51), but there was a significant interaction between time and treatment (F10,68 = 2.19, P = 0.03). Post hoc analyses revealed that, in Week 6, females in the 0-males treatment laid significantly fewer eggs than females in the 1-male (P = 0.02) or (P = 0.05) 3-males treatment. All other comparisons were non-significant.
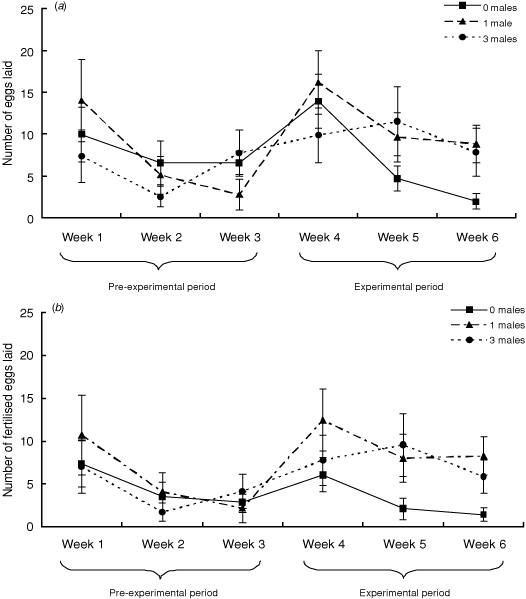
|
The total number of fertilised eggs increased between the pre-experimental and experimental periods (mean number of fertilised pre-experimental eggs laid = 14.0 ± 2.45; experimental eggs = 23.4 ± 4.33; t = 1.94, P = 0.04) and was comparable across the three treatment groups (F2,38 = 2.30, P = 0.11).
A repeated-measures ANOVA revealed significant variation in fertilisation success over the pre-experimental period (F5,34 = 2.56, P = 0.04) (Fig. 4b). Patterns of fertilisation were comparable across the three treatment groups (F2,38 = 1.98, P = 0.15). The interaction between time and treatment was not significant (F10,68 = 1.39, P = 0.20).
The average proportion of fertilised eggs laid by females during the pre-experimental period did not differ between treatments (0-males = 54%, 1-male = 57%, 3-males = 65%; χ22 = 2.23, P = 0.14). In contrast, after the experimental treatment, there was significant variation across the treatment groups (0-males = 39%, 1-male = 82%, 3-males = 78%; χ22 = 11.36, P = 0.0008). Post hoc analyses revealed that, on average, females in the 0-males treatment laid a lower proportion of fertilised eggs than females in the 1-male (P = 0.003) or 3-males (P = 0.006) treatments. There was no difference between females in the 1-male and 3-males treatment (P = 0.83).
Discussion
Our laboratory experiments provide the first insights into factors potentially influencing the development of nymphs, adult life-history traits and the mating behaviour of the apterous form of the Australian water strider, T. euphrosyne.
Fecundity and fertilisation
Following their initial isolation from males, each apterous T. euphrosyne female laid ~20 eggs over a 3-week period. The number of eggs laid and the number of fertilised eggs produced declined progressively during this time. In the absence of males, fertilisation success was reduced but complete sperm depletion was not observed: many females still laid fertilised eggs (albeit a lower number) even after 3 weeks of isolation from males. This finding supports many previous studies suggesting that, even in the absence of males, mated water strider females can continue to fertilise eggs successfully for several weeks (Kaitala 1987; Arnqvist 1988; Rubenstein 1989). In T. euphrosyne, sperm depletion is unlikely to be a strong selective pressure for apterous females because, in natural populations, they are most often found in large, mixed-sex aggregations (T. M. Jones and P. G. R. Colvin, personal observations). In contrast, the ability to store sperm for extended periods may be significant for the dispersing macropterous form as these females may have limited access to further males. From a male perspective, successful sperm storage by females enhances the potential for sperm competition to play a significant role in determining male fertilisation success (Simmons 2001). However, whether second male sperm precedence, which is typical for several other water striders (Arnqvist 1988, 1997; Rubenstein 1989), is present in this species, remains to be determined.
Following exposure to a male, the number of eggs laid and fertilised increased significantly for all females, suggesting that, even though not observed, mating occurred. However, the rate of fecundity (~1 egg per day) is almost an order of magnitude lower than that reported for other similar-sized water striders such as Gerris lateralis (Arnqvist and Danielsson 1999) or G. thoracicus (Kaitala 1991). The age of our experimental females was unknown and it is conceivable that age-related declines in female fecundity may, in part, account for this difference. The impact of additional extrinsic factors such as variation in food availability (Kaitala 1991) or climate on levels of fecundity also requires further assessment. The pattern of fecundity and fertilisation success observed in the three treatments following the post-mating period suggests that the degree of male access is an important factor in female reproductive success for T. euphrosyne. Females provided with a male for an initial 3-day period only suffered a steady decline in fecundity and fertilisation over the subsequent 3-week period. In contrast, females provided with either a single male or multiple males for the duration of the experiment were able to replenish depleting sperm stocks and maintained high levels of fecundity and fertilisation success. Moreover, the similarity in fecundity and fertility of the two groups of females provided with constant access to males suggests that their mating rates may be comparable and thus T. euphrosyne females are likely to remate with a previous mate.
Finally, the data indicate that repeated mating provides T. euphrosyne females with direct fertilisation benefits as is common among insect species (Arnqvist and Nilsson 2000). Further work is required to establish whether they also gain indirect benefits from mating such as increased offspring survival or fecundity (Jennions and Petrie 2000), or whether there are associated costs with mating, as is the case for other water striders (Rowe 1994).
Nymph survival, nutrition and cannibalism
In the laboratory, T. euphrosyne passed through five moults to reach the adult stage, a common feature of water strider species (Andersen 1982; Andersen and Weir 2004a). All surviving adults were apterous, which is unsurprising because only a small proportion of macropterous individuals have been observed in this species (Andersen and Weir 2004a), and it is unlikely that the laboratory conditions were sufficient to promote wing development (Spence and Andersen 1994).
Survival of nymphs was extremely low in our laboratory cultures: 95% died before reaching the adult stage. Such low levels of postembryonic survival have been observed previously in gerrids. In Gerris lacustris, only 10–20% of nymphs survive to the adult stage, with the degree of mortality varying with the season (Zimmermann et al. 1982). Similarly, field observations on Aquarius remigis found 70–90% winter mortality (Blanckenhorn 1994). Notably, these studies were conducted on field populations and it is perhaps surprising that such high levels of mortality should be observed in the laboratory. Food limitation, which directly affects juvenile survival and population growth (Spence and Andersen 1994), is the most parsimonious explanation for the low levels of survival we observed. Indeed, our data show that nymphal mortality varies with food availability: only three treatment groups produced any mature adults – single Drosophila were insufficient as a food supply for the production of adults. This may have arisen because the indigestible chitin content of D. melanogaster is over three times that of blowflies (12% v. 54%); however, it is also likely that because Drosophila are considerably smaller than blowflies, nymphs reared under these conditions were severely food limited. While not measured in this study, it is also conceivable that variation in food availability or individual foraging success may also promote a future trade-off in adult survival and fecundity (Kaitala 1987, 1991; Blanckenhorn 1990).
Our data highlight some possible causes of mortality in T. euphrosyne. Over 25% of developing nymphs died during moulting, and over 50% of dead nymphs displayed signs of cannibalism. To determine whether these nymphs died as a direct result of cannibalism or were cannibalised following death requires more frequent observations than the three per week carried out in our study; however, high rates of cannibalism are commonly reported in water striders and these have serious implications for survival and future life-history traits (Spence and Andersen 1994). For example, in Gerris pingreensis, conspecific cannibalism can lead to higher rates of nymph survival and females maturing at a heavier weight (Spence and Cárcamo 1991). Varying hunger levels and habitat complexity may also influence levels of cannibalism in seminatural populations (Spence 1986). Food availability is likely to have influenced the rate of cannibalism among T. euphrosyne nymphs, but additional field-based studies are necessary to assess the relative importance of other extrinsic factors, such as predation, which are a major cause of mortality for water strider species (Spence and Cárcamo 1991; Spence and Andersen 1994), particularly as these may also have consequences for adult mating behaviour (Sih and Krupa 1995; Moses and Sih 1998).
Mating behaviour
Typically, water strider mating behaviour is divided into two distinct types (Arnqvist 1997). Type I matings are categorised by limited male courtship, but intense pre- and postmating struggling behaviours. In type II matings, males typically court females using ‘ripple’ signals on the water surface and pairing behaviour is categorised by no, or less intense, struggling. T. euphrosyne appears to exhibit Type I mating behaviour, which follows a three-phase cycle of precopulation, riding and ovipositing. The male typically approached a female, who either moved away or permitted him to commence riding by grasping her with his forelimbs. Struggles were relatively short and rarely ended in the termination of a riding attempt. Females remained acquiescent for several hours following a struggle, while the male rode on her back. This behaviour contrasts starkly with that of many other water striders, where 50–80% of struggles result in a male being dislodged (for a review see Arnqvist 1997). While we did not observe mating, we assume it occurred because sperm-limited females given access to males increased their egg-laying rate and fertilisation success.
We found a positive correlation between the numbers of males observed riding and the proportion of available females. In contrast, the probability of a female pairing with a male was comparable regardless of the operational sex ratio. The small single-sex competitive aggregations also revealed that male success in riding behaviour was correlated with his foraging success, a pattern observed previously in the water strider Aquarius remigis (Blanckenhorn et al. 1998). This correlation most probably arises because males use their forelimbs to both capture prey and secure themselves on the backs of females, an interpretation supported by the positive correlation between male foraging success and forelimb length. No such correlations were observed for females. An important point raised by Blanckenhorn et al. (1998) is that foraging and mating cannot occur simultaneously under natural conditions. Thus, the relative importance of these two activities and the potential impact of intrasexual dominance relationships remains unknown.
In conclusion, our study provides the first observations of development, foraging behaviour and the mating system of one of the most common water strider species in south-eastern Australia. Further work is necessary to assess the consequences of the behaviours described and, in particular, examine their relative importance in a field context.
Acknowledgments
We thank Bojun Bjorkman-Chiswell, Peter Colvin and Jeanette Colvin for assistance in the laboratory and field and two anonymous referees for their comments on a previous version of this manuscript. TMJ was funded by the Australian Research Council (grant DP0209680).
Andersen, N. M. (1982). The semiaquatic bugs (Hemiptera, Gerromorpha). Phylogeny, adaptations, biogeography, and classification. Entomonograph 3, 1–455.
Andersen, N. M. , and Weir, T. A. (2004b). Mesoveliidae, Hebridae, and Hydrometridae of Australia (Hemiptera: Heteroptera: Gerromorpha), with a reanalysis of the phylogeny of semiaquatic bugs. Invertebrate Systematics 18, 467–522.
| Crossref | GoogleScholarGoogle Scholar |
Arnqvist, G. , and Danielsson, I. (1999). Postmating sexual selection: the effects of male body size and recovery period on paternity and egg production rate in a water strider. Behavioral Ecology 10, 358–365.
| Crossref | GoogleScholarGoogle Scholar |
Spence, J. R. (1986). Relative importance of mortality factors in field populations of the water strider Gerris buenoi Kirkaldy (Heteroptera: Gerridae). Oecologia 70, 68–76.
| Crossref | GoogleScholarGoogle Scholar |

Spence, J. R. , and Andersen, N. M. (1994). Biology of water striders – interactions between systematics and ecology. Annual Review of Entomology 39, 101–128.
| Crossref | GoogleScholarGoogle Scholar |
Spence, J. R. , and Cárcamo, H. A. (1991). Effects of cannibalism and intra-guild predation on pondskaters (Gerridae). Oikos 62, 333–341.
Wilcox, R. S. (1995). Ripple communication in aquatic and semiaquatic insects. Ecoscience 2, 109–115.
Zimmermann, M. , Hauser, R. , and Husler, J. (1982). Estimation of stage-specific densities and survival rates of the immature stages in a population of Gerris lacustris (Hemiptera, Gerridae). Revue Suisse de Zoologie 89, 941–955.