

Spatial ecology of invasive Bennett’s wallaby in South Island, New Zealand
A. David M. Latham A * , M. Cecilia Latham A and Bruce Warburton A
A * , M. Cecilia Latham A and Bruce Warburton A
A Manaaki Whenua – Landcare Research, PO Box 69040, Lincoln 7640, New Zealand.
Wildlife Research 50(12) 1109-1122 https://doi.org/10.1071/WR22127
Submitted: 14 July 2022 Accepted: 3 February 2023 Published: 2 March 2023
© 2023 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Context: Bennett’s wallaby (Notamacropus rufogriseus) was introduced into New Zealand in the late-1800s, and has subsequently become a significant pest to agriculture, silviculture, and native vegetation. Effective management of wallabies requires robust spatial and temporal ecological information, which can be used operationally to increase detection and kill rates.
Aims: To determine home range size and shape, resource (i.e. habitat and topography) selection, and daily movement patterns of three populations of Bennett’s wallaby in the South Island.
Methods: We attached GPS-collars to 38 adult wallabies (17 females, 21 males) between May 2018 and May 2019, and obtained usable location data from 30 individuals. We used kernel density estimators and resource selection functions to quantify seasonal home ranges and resource selection, respectively.
Key results: The mean seasonal home range sizes of males and females were similar (26.8 ha and 24.8 ha, respectively; combined range = 1.2–101.9 ha), although the largest home ranges belonged to large males. Resource selection was strongly influenced by distance to concealment cover, ridgelines and streams, i.e. wallabies selected to be closer to these features, though not necessarily at them. Wallabies selected areas close to concealment cover throughout 24-h periods, but most strongly during the day, which is when they bed in dense cover. Wallabies sampled outside of peak breeding moved most during dusk and dawn (median = ~50 m/h), whereas those sampled during peak breeding moved widely during day, dusk, and night (110–280 m/h), but not dawn (median = ~50 m/h).
Conclusions: The home range sizes of wallabies in New Zealand were highly variable, but on average were similar for males and females. Wallabies selected pasture close to areas with some cover, suggesting a trade-off between access to high-quality forage and concealment cover.
Implications: Our results provide robust data for modelling management strategies for wallabies, and contribute directly to operational planning by identifying habitats where wallabies are most likely to be found temporally. This information can be used to determine the appropriate survey methods and control tools to maximise detection rates and kill rates of wallabies, based on habitat.
Keywords: damage, eradication, home range ecology, invasive mammalian herbivore, Macropodidae, movement behaviour, Notamacropus rufogriseus, pest management, red-necked wallaby, resource selection, South Canterbury, sustained control.
Introduction
Introduced and native wild mammalian herbivores can have significant unwanted impacts on native vegetation (Jaksic 1998; Côté et al. 2004; Dolman and Wäber 2008) and primary production (Putman and Moore 1998; Hone 2007; Latham et al. 2020a). Some species of macropods (wallabies and kangaroos) are included in this suite of pest herbivores, especially where they are overabundant, which includes parts of their native range in Australia (Arnold et al. 1989; le Mar and McArthur 2001; Wiggins and Bowman 2011; Gordon et al. 2021) and countries to which they have been introduced (Long 2003; Latham and Warburton 2021). This has resulted in the need to manage pest macropods using lethal or non-lethal methods to mitigate the damage they cause (Warburton 1986; le Mar and McArthur 2001; Wiggins and Bowman 2011).
Two key strategies for managing introduced wallabies (i.e. Notamacropus spp., Petrogale penicillata, and Wallabia bicolor) are sustained control and eradication (Latham and Warburton 2021). To date, sustained control, especially using shooting and poisoning, has been the primary strategy used for managing pest wallabies (Warburton 1986; le Mar and McArthur 2001). However, large-scale eradication of vertebrate pests is increasingly being suggested as the way forward for pest management (Russell et al. 2015; Owens 2017). Despite this, eradication is challenging and broadscale eradication programmes have rarely been successful (Riney 1956; Howard 1958; Parkes et al. 2017). The benefit of eradication is that it permanently removes the unwanted impacts caused by the pest, but it makes sense as a policy only if it is achievable (Parkes and Murphy 2003; Parkes et al. 2017).
Although eradication programmes have previously been successfully developed with limited ecological knowledge of the target species (e.g. rats; Rattus spp.; Russell and Holmes 2015), basic ecological information is important, especially when attempted eradication fails for unknown reasons. For example, understanding the spatiotemporal population dynamics of the pest can optimise the timing and likelihood of success of an eradication attempt (Harper et al. 2015). Similarly, home range information can be used to quantify detection and trappability to guide the optimal spacing and placement of survey devices, traps, and bait stations (Efford 2004; Mackenzie et al. 2022). Spatial ecology can highlight habitats and geographical features that may permit reinvasion by the pest. Once eradication is believed to have been achieved, it must be confirmed to demonstrate that an inability to detect target animals means that none survived, rather than that survivors remain but were not detected (Morrison et al. 2007; Latham et al. 2021). Confirming eradication is also underpinned by spatial ecology, such as habitat use or selection, because this affects the probability that a pest will be detected (Latham et al. 2021).
Bennett’s wallaby (or red-necked wallaby; Notamacropus rufogriseus) is native to parts of mainland eastern Australia and Tasmania, and has been introduced into New Zealand and some European countries (Eldridge and Coulson 2015; Latham and Warburton 2021). Brought from Tasmania to Christchurch, New Zealand, in 1870 and liberated on the eastern Hunters Hills, near Waimate, in 1874 (Thomson 1922; Studholme 1954), Bennett’s wallabies have spread from their release site and occupied an area estimated at 5300 km2 as of 2015 (Latham et al. 2019). This area comprises what is termed the ‘containment area’ and is bounded by natural geographical features (such as coastline, large rivers, and mountain ranges) that prevent or minimise natural dispersal. However, dispersing wallabies have ‘escaped’ the containment area and established low-density populations around its periphery, and this has increased the geographical extent of this species to an estimated 14 000 km2 of central and eastern South Island (Latham et al. 2019). Bennett’s wallaby is considered a significant pest to agriculture, silviculture, and native vegetation in New Zealand (Latham and Warburton 2021), although empirical data to support this are sparse (Latham et al. 2020a). Nevertheless, if Bennett’s wallabies are permitted to establish and attain moderate- to high-density populations in newly invaded areas, the extent of the damage they cause to New Zealand’s production assets and native vegetation is likely to be greatly increased (Latham et al. 2020a).
To ameliorate the damage caused by Bennett’s wallaby and prevent or slow its spread, various agencies in New Zealand have been responsible for conducting sustained lethal control programmes against wallabies since the 1940s (Catt 1975; Warburton 1986). Initially this was done by the Department of Internal Affairs, with about 70 000 Bennett’s wallaby being killed by government cullers between 1947 and 1956. The South Canterbury Wallaby Board (SCWB) was especially effective at maintaining wallaby populations at low densities from 1971 until they were replaced by Environment Canterbury in November 1989 (although the SCWB was not officially disbanded until June 1992; Latham and Warburton 2021). At this time, government subsidies that had applied to wallaby control stopped, and affected landowners had to meet the full cost of control. Since 1989 wallaby numbers have steadily increased, and they have spread, through both natural dispersal and deliberate relocations by humans (Latham et al. 2019).
In response to the progressive spread of Bennett’s wallaby in the South Island, the New Zealand Ministry for Primary Industries was tasked with leading a national wallaby eradication programme (NWEP), with initial funding from 2020 to 2024. The preliminary aims of NWEP were to find and eradicate populations of wallabies outside the containment area and suppress their numbers within the containment area to mitigate damage and reduce dispersal pressure, which is believed to be density-dependent (Warburton and Latham 2020). The ultimate goal of the NWEP is the eradication of all wallabies from New Zealand.
Despite the long-time pest status of Bennett’s wallaby, little is known about the spatial ecology of this species in New Zealand (Latham and Warburton 2021). Observations have shown they can move about 1–3 km in a night as they forage (McLeod 1986). However, home range, resource (habitat and geographical features) selection, and movement ecology have not been quantified in New Zealand, although they have been in Australia (Wiggins et al. 2010; Wiggins and Bowman 2011; Garnick et al. 2014, 2016; Garnick and Coulson 2021). We aimed to collect data on home range size and shape and daily movement patterns from three populations of Bennett’s wallaby within the containment area. We also quantified wallaby resource selection, with emphasis on habitats and geographical features that may affect temporal detection rates and/or kill rates of wallabies. For example, shooting is a common method for reducing wallaby numbers (Warburton 1986), but detecting and shooting wallabies is likely to be affected by the availability and time of use of concealment cover. Distance from open foraging habitats to concealment cover has been assessed in Australia, with some studies showing that wallabies foraged close to concealment cover (Johnson 1987; le Mar and McArthur 2005), whereas others showed that foraging also occurred in open habitat with no concealment cover nearby (Garnick et al. 2014). We tested the prediction that Bennett’s wallaby in New Zealand would select feeding areas that were close to dense vegetative cover that provides concealment, because this provides some degree of protection from being seen and shot.
Methods
Study areas
We assessed the spatial ecology of Bennett’s wallaby at three study sites, all high-country stations (farms) carrying sheep and beef cattle in South Canterbury (previously described in Latham et al. 2021). All study sites were located within the containment area delineated for Bennett’s wallaby by Environment Canterbury (the management agency responsible for biosecurity in this region) (Latham et al. 2019). The study sites were at least 35 km apart from each other.
The densities of wallabies varied considerably within the containment area but may have been as high as two or more wallabies per hectare in the most heavily infested areas (Latham and Warburton 2021). We chose study areas with estimated low to moderate densities of wallabies, using the Guilford Score, which is a visual assessment of faecal pellet abundance, tracks, and wallaby sightings (Latham and Warburton 2021). Assessments of our study areas placed them at 3 (with a range of 2–4) on the Guilford Score, indicating that wallabies occurred at low to moderate densities (Latham et al. 2021).
Glen Cary Station (170.42°E, 44.63°S), Waimate District, in the southern part of the containment area, was assessed in May 2018. This site has a predominantly easterly aspect, and was characterised by pasture at lower elevations (~450–600 m above sea level; a.s.l.), tall snow tussocks (Chionochloa spp.) at higher elevations (~600–900 m a.s.l.), and scrub dominated by matagouri (Discaria toumatou) in gullies. There was a small block of Pinus radiata in a gully at ~500–650 m a.s.l.
Blue Cliffs Station (170.88°E, 44.51°S), Waimate District, in the eastern central part of the containment area, was assessed from early August to mid-September 2018. This site is within a stream catchment that faces east, resulting in hill faces on the south side of the stream having a primarily northerly aspect and the north side of the stream having a primarily southerly aspect. The site was a mosaic of pasture and matagouri scrub (especially on north-facing slopes), tall snow tussocks at higher elevations (~600–700 m a.s.l.), New Zealand flax (Phormium tenax, especially on south-facing slopes), and patches of remnant native forest and scrub.
Grampians Station (170.60°E, 44.21°S), Mackenzie District, in the western central part of the containment area, was assessed from late February to early May 2019. This site is situated on the Mackenzie River, which flows in a northerly direction, so one set of hill faces have a primarily eastern aspect and the other, a primarily western aspect. This was the highest-elevation site (~700–1100 m a.s.l.), and was a mosaic of unimproved pasture, matagouri scrub, and tall snow tussocks and subalpine scrub at higher elevations (~900–1100 m a.s.l.).
The climate at all three sites was dominated by a dry (annual rainfall ~300–600 mm), temperate, continental climate. Summers are sunny and mild (mean high ~19–22°C; mean low ~7–10°C), and winters are cool (mean high ~5–12°C; mean low ~−3–1°C). Snow can accumulate at all sites in winter. A suite of other mammalian herbivores occurred at all sites, including domestic sheep (Ovis aries) and domestic cattle (Bos taurus), as well as common brushtail possums (Trichosurus vulpecula), European rabbits (Oryctolagus cuniculus), brown hares (Lepus europaeus), feral pigs (Sus scrofa), chamois (Rupicapra rupicapra), red deer (Cervus elaphus), and fallow deer (Dama dama). Hunting for sport and meat, and pest control, occurred at all sites, but was halted, insofar as was possible (i.e. some illegal hunting may have occurred), for about 3 months before we started and for the duration of our research at each site. This was done because effective pest control can reduce wallaby numbers to low levels and/or alter their behaviour, so we may have struggled to capture enough animals. Once collared, we did not want animals shot.
Wallaby capture and GPS collaring
We used two methods to capture Bennett’s wallabies (described in detail in Latham et al. (2020b)). At all study sites we used tunnel nets set on wallaby runs (i.e. well-used trails created by their large, soft-padded hindfeet) under farm fences to capture adult wallabies. At Glen Cary and the Grampians we also used a net-gun fired from an MD 520N helicopter (MD Helicopters Inc., Mesa, Arizona, USA). We used a combination of physical and chemical immobilisation for wallabies captured using tunnel nets, whereas all wallabies netted using a helicopter were physically restrained. Chemical immobilisation was done using a low-dose (1–4 mg kg−1) of tiletamine–zolazepam (Zoletil™, Jurox Inc., North Kansas City, Missouri, USA), as recommended by le Mar and McArthur (2000).
Once adult wallabies were immobilised, we fitted them with a customised GPS radio-collar (designed and built inhouse by Manaaki Whenua – Landcare Research staff; see Latham et al. 2021 for full details). We only collared wallabies without obvious physical injuries and that were not suffering from physical exhaustion (based on a subjective assessment by field personnel) (see Latham et al. 2020b). The weight of the custom collars was ~200 g. Adult Bennett’s wallabies weigh about 9–18 kg, but occasionally up to 25 kg (Latham and Warburton 2021). We only collared adult wallabies (≥9 kg), and therefore the weight of the collars was ≤2.2% of wallaby body weight (Latham et al. 2020b), well within the accepted international standard (Casper 2009; Latham et al. 2015).
We programmed the customised GPS collars to take two fix rates. First, collars attempted to take one fix per hour, every hour, for the battery life of the collar (~3 months). Second, collars collected a very high fix rate (one fix every 5 s) every morning (0600–1200 h) for the battery life of the collar. The high fix-rate schedule was needed for a study that quantified detection parameters for Bennett’s wallaby, but was used here to complement the hourly data (see Latham et al. 2021). The collars had a store-onboard function for location data; however, we also used two wireless sensor networks, DigiMesh® (Digi International, Hopkins, Minnesota, USA) and LoRa (LoRa Alliance®, Fremont, California, USA), for remotely communicating with the GPS collars and acquiring location data in near real time (Ekanayake and Hedley 2018).
All capture, handling, and collaring protocols used in our study were approved by the Manaaki Whenua – Landcare Research Animal Ethics Committee (approval no. 17/11/03).
Seasonal home range analysis
We used GPS location data obtained from GPS-collared wallabies to describe seasonal home range size and shape (for our study, we use ‘home ranges’ hereafter to refer to seasonal home ranges). Home range estimates are sensitive to sample size and estimation method (Boyle et al. 2008). Therefore, we excluded individual wallabies for which we had fewer than 40 GPS locations (Girard et al. 2002). We used kernel density estimators (KDEs; Worton 1989) to estimate wallaby home range size and shape using the adehabitatHR package (Calenge 2006) in R (R Development Core Team 2022). We used the KDEs to create a utilisation distribution (UD) for each monitored wallaby and then estimated home range area based on the 95% UD. Two key parameter choices need to be made to estimate the UD: the overall shape of the distribution of the animal’s relocations (the density kernel) and the bandwidth, which calculates the size of the distribution. We used a bivariate normal density kernel to represent the UD (Worton 1989). Because KDEs can differ based on the algorithm used to estimate the bandwidth, we tested three alternative methods: (1) the reference bandwidth; (2) the least-square cross-validation bandwidth; and (3) the plug-in bandwidth. We visually inspected the results of the different bandwidth estimation methods and retained home range estimates obtained using the reference bandwidth approach, because we considered these represented cohesive home ranges rather than areas of greatest use within a home range (Worton 1989).
Landscape covariates for resource selection analyses
Explanatory variables included in our resource selection analyses were derived using ArcGIS Pro 2.9.3 (ESRI (Environmental Systems Research Institute) 2022), and included land-cover type, topography, and ‘distance to’ covariates. Earlier studies have shown that more useful insights into resource selection can be gained from considering habitat attributes at multiple discrete scales (Boyce 2006; Leblond et al. 2011). Therefore, we collected landscape covariates at two spatial scales (circular buffers of 50- and 100-m radii) and used a multi-model approach to determine which scale better described wallaby resource selection (Leblond et al. 2011). These scales were assumed to correspond to two hierarchical movement decisions (as defined by Leblond et al. (2011)). The smallest spatial scale was chosen to account for GPS location error, as well as to assess wallaby perception for habitats within their immediate vicinity, which are often homogeneous relative to larger spatial scales. The larger spatial scale was based on the mean distance moved per hour by all collared wallabies. This spatial scale was chosen to assess if wallaby resource selection is better explained by their perception of the more heterogeneous environment beyond their immediate surrounds (Leblond et al. 2011). Within each buffer (i.e. 50 m and 100 m), we derived land-cover types from the Land Cover Database (LCDB ver. 5.0; www.lcdb.scinfo.org.nz), which had been ground-truthed by field personnel (ADML, unpubl. data). We classified land-cover types into ‘grey’ scrub (especially matagouri), tall tussock, low-productivity grassland, improved pasture, exotic forest (P. radiata, which was only present at Glen Cary), and remnant native broadleaved hardwood forest (only present at Blue Cliffs).
We converted land-cover types to a 25-m resolution raster, and included these in all analyses as the proportion of each land-cover type within the two discrete buffers around each GPS location. Topographic variables were derived from a 25-m resolution digital elevation model (DEM) of the study area, and included aspect (four categories, i.e. cardinal directions, using south as the reference category), elevation (m), slope (°), and terrain ruggedness (standard deviation of elevation). Within the two different-sized buffers around each GPS location, we recorded the most common aspect, mean elevation, mean slope, and terrain ruggedness. ‘Distance to-’ variables comprised distance to nearest ridgeline, stream, and concealment cover (i.e. from pasture or short tussock to one of the following: any forested cover type, scrub, or tall tussock). A negative coefficient for a distance to- covariate should be interpreted as selection for areas closer to that feature. For example, a strong negative coefficient for distance to stream would indicate that the study species selects areas closer to streams, whereas a positive coefficient shows the opposite.
Resource selection
We evaluated resource selection within the scale of their home range, i.e. third-order selection (sensu Johnson 1980), using resource selection functions (RSFs; Manly et al. 2002). An RSF is a widely used statistical analysis for assessing habitat selection (i.e. use in proportion to availability as opposed to habitat use, which is not corrected to account for the availability of each habitat). It can also accommodate non-habitat covariates (if available), such as distribution of prey species for a predator (Latham et al. 2013), as well as complex interaction terms for important covariates (Manly et al. 2002). Briefly, an RSF assesses the landscape characteristics at used locations (in our case, GPS locations), and compares these characteristics to those obtained from available locations (i.e. randomly selected locations the animal could have used within its home range). This is similar to a used (‘1’)–unused (‘0’) statistical design, but in this instance we only know used locations (based on GPS), and unused locations are instead replaced by available locations (Manly et al. 2002).
Given potential differences in topography and habitat, and the time of year wallabies were collared in each study area, we conducted a separate RSF analysis for each study site. We defined the area available to each wallaby (i.e. the domain of availability; Manly et al. 2002) as their home range, estimated using 100% minimum convex polygons (MCP) using the ‘adehabitatHR’ package (Calenge 2006) in R (R Development Core Team 2022). We used 100% MCPs, as opposed to 95% MCPs or some other method of home range estimation, to define the domain of availability because they provide a more complete description of the area an individual could choose to use within its range. Within each 100% MCP, we randomly drew five available locations per used location, which equated to a mean of 154 (range: 4–870) available locations per hectare of 100% MCP, to describe resource availability for each animal.
Prior to analysis we conducted data screening, following Bjørneraas et al. (2010), to identify erroneous locations that were beyond the possible range of wallaby movements or that produced abnormal movement spikes. This analysis identified 0.02%, 0.16% and 0.98% of locations as erroneous in Glen Cary, Blue Cliffs and Grampians, respectively. We also rarefied high fix rate location data to 15-min intervals to decrease autocorrelation between successive locations (Fieberg et al. 2010). We fitted resource selection functions using mixed-effects logistic regression (Johnson et al. 2006) using the package lme4 (Bates et al. 2015) in R (R Development Core Team 2022). We included a random intercept for each individual wallaby to account for repeated observations obtained from the same animal, as well as an unbalanced sampling design (Gillies et al. 2006).
We followed three steps to build our RSF models for each study site. First, we assessed multi-collinearity between pairs of covariates, using Pearson correlation coefficients (rp). For those pairs with rp ≥ |0.7|, we retained the most biologically important variable of the pair for further analyses, as recommended by Montgomery and Peck (1992) and based on wallaby ecology in New Zealand (Latham and Warburton 2021). Second, we identified the best scale to contextualise landscape variables in wallaby resource selection modelling by comparing models containing uncorrelated variables collected within 50 m vs 100 m buffers, using the Akaike Information Criterion (AIC; Burnham and Anderson 2002). Third, we added interaction terms between distance to concealment cover, ridge, and stream (each analysed separately) and time of day (t, recorded in decimal hours), to assess diel (24 h) variation in the strength of selection for these landscape features. Following Forester et al. (2009), we assessed diel patterns by including interaction terms with four harmonics of time of day, calculated as s1 = sin(2πt/24), s2 = sin(4πt/24), c1 = cos(2πt/24), and c2 = cos(4πt/24).
For each study site, we considered explanatory variables with 95% confidence intervals not overlapping zero to have a significant influence on resource selection by Bennett’s wallaby. We assessed the predictive performance of the RSF models using k-folds cross-validation (Boyce et al. 2002). An RSF model with a Spearman rank correlation (rs) >0.8 is considered to have performed well (Boyce et al. 2002).
Movement patterns
We described the daily movement patterns of wallabies by comparing the distribution of step-lengths (m/h) across four time periods: dawn, day, dusk, and night. We defined dawn and dusk as 1 h before and 1 h after sunrise and sunset, respectively. We determined the timing of sunrise and sunset by using those recorded for the nearby city of Timaru, South Island, for the months of the year when research was done in each study area (https://www.timeanddate.com/sun/new-zealand/timaru). We defined day as those hours that fell between dawn and dusk, and night as those hours between dusk and dawn.
Results
In total, we fitted GPS collars to 38 adult wallabies (17 females, 21 males). By study site, we collared eight wallabies (one female, seven males) at Glen Cary, 15 (10 females, five males) at Blue Cliffs, and 15 (six females, nine males) at the Grampians (Latham et al. 2021). Thirty wallabies yielded enough GPS data for subsequent analyses. Of the eight wallabies that had no usable data, three were from Glen Cary (including two early mortalities and one collar failure), three were from Blue Cliffs (one early mortality and two collar failures), and two were from the Grampians (two collar failures) (Latham et al. 2020b). We retrieved 465 655 locations from Glen Cary, 931 438 locations from Blue Cliffs, and 905 815 locations from the Grampians. The percentage of missed fix attempts was 5.5% in Glen Cary, 13.2% in Blue Cliffs, and 6.0% in Grampians.
Seasonal home ranges
Seasonal home range sizes and shapes varied widely among study areas and individual animals (Table 1, Fig. 1). Home ranges were, on average, larger at the Grampians than at the other two study sites, although two wallabies from Glen Cary and five from Blue Cliffs had reasonably large, elongated home ranges. Home range shape was circular for most animals at the Grampians. The largest home ranges at all study sites belonged to males. However, the mean home range sizes of males (26.8 ha; s.e. = 5.88) and females (24.8 ha; s.e. = 5.94) were comparable. Because of missing weight records for nine wallabies (all from Glen Cary and Grampians), we only correlated home range size with wallaby weight for Blue Cliffs. Although variable, home range size at Blue Cliffs was positively correlated with wallaby weight (r = 0.583, P = 0.046).
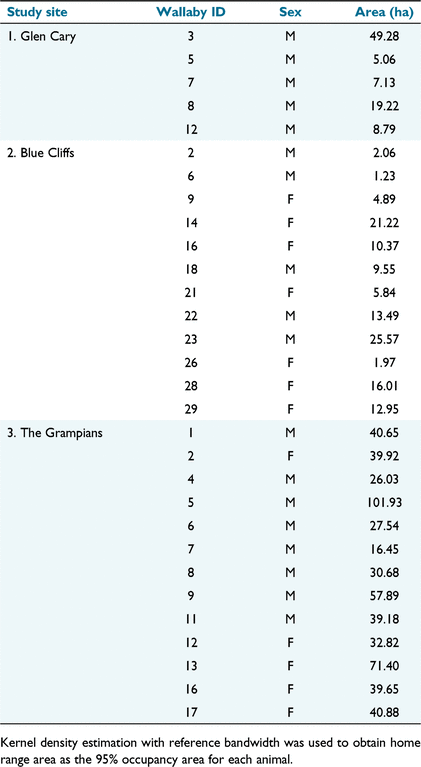
|

|
We did not record any permanent shifts in the core areas of use for any wallabies at any of the study sites, albeit based on a relatively short monitoring period (<1–2 months per wallaby). In Glen Cary, collared wallabies were monitored for 10–19 days in autumn (May) 2018. One wallaby (no. 3) at Glen Cary made daily forays (~1.5 km in a straight-line distance) from low-productivity grassland at higher elevations, through scrub, to paddocks with improved pasture, presumably to feed during the hours of darkness (Fig. 1a). Wallaby no. 8 used the heads of three separate gullies, which were separated by >1 km. Collared wallabies at Blue Cliffs were monitored for 8–41 days in spring (August and September) 2018. During this period, several wallabies made daily long-distance forays (>1 km in a straight-line distance) between forest, tussock, and low-productivity grasslands (e.g. wallabies 14, 22, 23, and 28; Fig. 1b). Collared wallabies at the Grampians were monitored for 27–55 days in autumn (March and April) 2019. During this period, most made numerous short-distance movements between low-productivity grassland, tussock, and scrub, all habitat types that were present within most wallaby home ranges (Fig. 1c). Straight-line, long-distance forays were less obvious at Grampians (with the exception of wallabies 5 and 13).
Resource selection
We found that the scale at which landscape covariates influenced resource selection by Bennett’s wallaby varied among sites. The buffer size that best explained the influence of landscape variables was 100 m at Glen Cary and Blue Cliffs, but 50 m at the Grampians.
Resource selection was strongly influenced by distance to concealment cover, ridgelines, and streams, but selection for these features varied with time of day (Fig. 2). At Glen Cary, wallabies selected areas closer to cover most strongly during daylight, whereas selection for areas closer to ridgelines was strongest at night. Wallaby selection for areas closer to streams was similar throughout the day and night, albeit with a small peak of selection about midday. At Blue Cliffs, wallabies also selected areas closer to concealment cover most strongly during daylight. Unlike Glen Cary, wallabies at Blue Cliffs selected areas closer to ridgelines during the day and avoided areas closer to streams. At the Grampians, wallabies selected areas closer to concealment cover most strongly at sunrise and sunset. They also selected areas close to ridgelines and streams, but there was no noticeable pattern across a 24 h period.

|
At Glen Cary, wallabies selected northern and eastern aspects rather than southern aspects (Table 2). On average, wallabies at Blue Cliffs selected southern aspects, whereas wallabies at the Grampians selected western aspects. At Glen Cary and Blue Cliffs, wallabies selected areas with steeper slopes, whereas at the Grampians they preferred areas with gentler slopes. Wallabies at the Grampians preferred areas at lower elevations, whereas at the other two sites elevation was not a significant predictor of wallaby habitat selection.

|
Although distance to concealment cover was important at all study sites, there were no clear patterns in selection for habitat (vegetation types), and there was variability between sites (Table 2). Wallabies at Glen Cary selected for areas with a higher proportion of tall tussock, whereas these areas were avoided at Blue Cliffs and Grampians. On average, wallabies at Glen Cary and Grampians avoided areas with a high proportion of improved pasture and/or low-productivity grassland. Finally, wallabies generally avoided areas with a high proportion of scrub (which should not be interpreted as an avoidance of scrub per se), although this effect was not statistically significant for wallabies at the Grampians.
Based on k-folds cross-validation, our RSF models performed reasonably well for Glen Cary rs = 0.75 (P = 0.02), and very well for Blue Cliffs rs = 0.98 (P < 0.01) and the Grampians rs = 0.99 (P < 0.01).
Movement patterns
Wallabies at Glen Cary and Blue Cliffs moved longer distances during dusk and dawn than they did during the day or night (Fig. 3). At the Grampians, the longest movements were observed during the day and night (means of 290 and 232 m/h, respectively), whereas the shortest distances were observed at dawn (mean = 62 m/h). Although wallabies at the Grampians tended not to make straight-line long-distance movements compared with the other two sites (see ‘Seasonal home ranges’ above), they did, on average, move far larger distances per hour than did wallabies at the other two sites (Fig. 3). This was particularly obvious (and variable) at the Grampians during day, dusk, and night (Fig. 3), and comprised movements that tended to be tortuous and that alternated between long and short distances moved within their largely circular home ranges.

|
Discussion
Our study represents the most comprehensive assessment of the spatial ecology and movement behaviour of invasive Bennett’s wallaby in New Zealand to date. We assessed the spatial ecology of three populations of wallabies within the containment area and obtained a large amount of location data from 30 wallabies (18 males and 12 females), from May 2018 to May 2019. This sample size permitted us to make strong inferences about wallaby spatial ecology, although we acknowledge the temporal deployment of GPS collars was a limitation of this study (i.e. we did not have location data from all months of the year, or from multiple years). Moreover, we did not assess habitat-induced biases in GPS locations, which can influence analytical results (Frair et al. 2010; Latham et al. 2015). Given there was no closed-canopy forest at two study sites, and wallabies showed strong selection for streams in steep gullies (where satellite coverage is often poor), we do not believe that habitat-induced biases greatly influenced our results.
Prior to this work, the home range ecology of Bennett’s wallaby was unknown in New Zealand, although it had been assessed in Australia, where they are indigenous. For example, based on 95% fixed kernels with a least-squares cross-validation smoothing parameter, le Mar et al. (2003) found that the mean size of home ranges of male and female Bennett’s wallaby in forestry in north-west Tasmania was 71.9 ha (±11.3 s.d.) and 33.1 ha (±14.7 s.d.), respectively, compared with our means of 26.8 ha for males and 24.8 ha for females. Although the mean for females is not too dissimilar, the mean for males from Tasmania is substantially larger than reported for this study, with only two males from the Grampians having home ranges of the same or larger size (71.4 ha and 101.9 ha; Table 2). This may be because le Mar et al. (2003) had radio-collars deployed for a longer period (11 months) than we did in the current study, or it might be related to differences in habitat (agricultural in our study vs forestry in north-west Tasmania) and therefore food productivity, or the population dynamics between the two geographical areas may have affected male home range ecology differently. In agricultural northeastern Tasmania, home range sizes (estimated using the 95% local convex-hull method) were 10.3 ± 1.9 ha and 11.6 ± 1.7 ha before and after lethal control, respectively (Wiggins et al. 2010). The differences in home range areas between the two Tasmanian studies were attributed to differences in habitat quality (agricultural vs forestry) and food productivity between the two areas, and to the type of hardware used to acquire location data (GPS in Wiggins et al. (2010) vs VHF le Mar et al. (2003)). Given we did not deploy GPS collars for a complete year, our home range estimates are seasonal estimates and may therefore be smaller than annual home ranges.
Most home ranges in our study were approximately circular, especially at the Grampians. However, many individuals (primarily males; see Fig. 1) had home ranges that were elliptical, notably at Blue Cliffs, but also wallaby 3 at Glen Cary. The difference in home range shape may be related to topography (e.g. wallaby home ranges extending along a gully), or it may have been related to food availability, especially paddocks containing crops or improved pasture. As non-territorial animals (Johnson 1989), Bennett’s wallaby may show high home range plasticity, incorporating nearby crops and improved pasture into their home ranges (or foraging ranges) as those become available, thereby creating elliptical home ranges.
Home range shape is more than an interesting ecological observation: it has direct implications for quantifying the detectability or trappability of an animal (Latham 2019). For example, many simulation models that predict the removal and surveillance efforts needed to achieve and confidently declare pest eradication assume that animals are removed or detected according to a half-normal detection/capture probability function (Efford 2004). This function assumes that the probability of detection or capture is maximal at the centre of the home range, and that this probability decays following a sigmoidal curve, with the shape of the curve determined by the parameter sigma (σ). Sigma assumes that the area an animal occupies 95% of the time is a circle with a radius of 2.45 σ. If some animals have home ranges that are irregular, the probability of use, and of detection by a device, might not be maximal at the animal’s home range centre, or equivalent in all directions at a given distance, as is assumed by the half-normal function (Efford 2004). Alternative specifications for the detection function have been suggested to accommodate non-circular home ranges (e.g. uniform detection probability for devices located at distances from home range centre smaller than σ and zero detection probability otherwise; Efford 2004), or directly modelling the detection probability as a function of kernel probability density (Ball et al. 2005). In our study, home range shape at Blue Cliffs arguably violates the assumption of circular home ranges (despite having been previously used under the assumption that they are circular: Latham et al. 2021). We recommend that the effect of elliptical home ranges on detectability or trappability analyses that assume half-normal detection functions be assessed.
We found support for our prediction that distance to concealment cover was an important variable influencing resource selection by wallabies. Concealment cover has previously been reported as important for resting and denning sites for Bennett’s wallaby in New Zealand (McLeod 1986) and Australia (Johnson 1987; le Mar and McArthur 2005), especially during daytime (Latham and Warburton 2021). McLeod (1986) reported that scrub, tall tussock, and native flax provide important cover for males, whereas females often use more secluded places in dense undergrowth (e.g. scrub or forest). Our results showed that wallabies strongly selected areas closer to concealment cover at all times, but especially during daylight (although this pattern was present at the Grampians, it was less strong; Fig. 2). This distance to variable, by definition, assesses wallaby selection for the ecotone boundary between cover (tall tussock, scrub, or forest) and open habitats where wallabies acquire much of their food (improved and unimproved pasture and short tussock) (Latham and Warburton 2021). It therefore encapsulates two critical components of wallaby ecology – food and cover from the perceived risk of being hunted. Distance to concealment cover and ridgelines were the only variables wallabies consistently selected at all study sites, i.e. wallabies, on average, selected to be closer to cover and ridgelines than further away from these features. Given that wallabies at all sites selected to be close to cover, the avoidance of areas with a high proportion of scrub and tall tussock by wallabies at Blue Cliffs appears unusual. However, this should be interpreted as selection for pasture close to areas with a low proportion of cover (tall tussock, scrub, or forest). In other words, there was a trade-off between wallabies wanting to be near food sources, but also close to adequate concealment cover. Denser vegetation cover may also provide thermoregulatory benefits for wallabies (Garnick et al. 2014). Wallabies in this study tended to avoid areas with a high proportion of scrub and tall tussock, suggesting they were not seeking a thermoregulatory benefit from the cold (especially relevant to Blue Cliffs, which was assessed in winter). Nevertheless, a low proportion of scrub, tall tussock, or forest could provide respite from the heat of the day. Selection or avoidance for all other habitat and geographical covariates was site dependent. Although some other covariates were important at a site level (e.g. selection for tall tussock at Glen Cary), distance to concealment cover and ridgelines appears to be consistently important for wallabies within the containment area in South Canterbury.
We found that wallabies generally moved about 50 m/h at most study sites and at most times of the day. Day, dusk and night at the Grampians were the exceptions, when wallabies travelled 110–280 m/h (Fig. 3). Given that wallabies usually rest in cover during daytime (Latham and Warburton 2021), we cannot explain the particularly large hourly movements during daytime at the Grampians. It may simply be an artefact of our partition of a 24-h period, with wallabies at the Grampians moving from feeding areas to cover during daylight, as opposed to dawn and dusk, which appears to be when wallabies at Glen Cary and Blue Cliffs were most active. Nevertheless, this does not explain the much larger distances moved at the Grampians relative to the other two sites. We speculate that the larger home ranges and distances moved at the Grampians may be related to the dearth of improved pasture at this site and the need to move greater distances to find forage and browse to meet their energy requirements. Another possible factor is that research at the Grampians spanned the peak breeding season (February and March; Catt 1977), whereas it did not at the other two sites. If the breeding season did influence movement behaviour of wallabies at the Grampians, the results in Fig. 3 suggest that breeding-related movements (i.e. to find mates, or for males, to defend mates and chase competitors away; Catt 1977) were common at all times of the day except dawn, when feeding or grooming (i.e. activities that involve less longitudinal movement) may have been more prevalent.
Management implications
Our results add to our understanding of the natural history and ecology of Bennett’s wallaby in New Zealand, and provide invaluable data for modelling management strategies for this invasive species. For example, the ecological data we have presented here are important for parameterising models assessing detectability or trappability, habitat suitability, movement behaviour, invasion pathways, geographical spread, proof of eradication, and so on. Having home range data for a range of individuals, especially those with small home ranges, provides conservative values of σ, which should be used in proof-of-eradication modelling to avoid overestimating the probability that eradication has been achieved. A cautionary note: although our data may be useful for models that predict invasion or geographical spread, our data were from resident wallabies, not dispersers. To better understand wallaby ecology at the invasion front, suitable methodology must be used on wallaby populations that border the containment area. Ideally this would include a better understanding of the dispersal ecology of wallabies, although the study of animal dispersal is challenging (Nathan 2001).
In addition to advantages for modelling, there is also the potential for direct operational gains by using the information from our home range, resource selection, and movement analyses to improve the effectiveness and efficacy of control tools, such as optimising the spatial deployment of aerially delivered toxic baits (as has been done for possums (Nugent et al. 2012) and rabbits (Latham et al. 2016)), and of bait stations (as has been done for ship rats (Rattus rattus; Mackenzie et al. 2022)). This is particularly relevant to the aerial application of 1080 baits, when baits are strip (trickle) sown and need to be placed in areas that have the highest use by the target species, a requirement that may be needed for Bennett’s wallaby. Having robust information about home range sizes will also be useful for optimising the spatial layout of bait stations for the control of Bennett’s wallaby, as well as for wallaby monitoring using camera traps or other methods.
Supplementary material
Supplementary material is available online.
Data availability
GPS data for all study animals can be found in: Latham ADM, Warburton B (2023) GPS location data for Bennett’s wallaby in South Island, New Zealand [Dataset]. Manaaki Whenua – Landcare Research – Datastore. doi: https://doi.org/10.7931/h3d1-ej64.
Conflicts of interest
The authors declare they have no conflicts of interest.
Declaration of funding
Funding was provided by the New Zealand Ministry for Primary Industries’ (MPI) Sustainable Farming Fund (Project No. 405254) and the MPI national wallaby eradication programme (NWEP), Environment Canterbury, Otago Regional Council, and Manaaki Whenua – Landcare Research (MWLR) Strategic Science Investment Funding.
Acknowledgements
We are grateful for the support of the Waitaki Wallaby Liaison Group, especially Mike Paterson, for supporting this research. We thank landowners/managers John Abelen (Glen Cary), Tom Bell (Blue Cliffs), and Guy King (Grampians) for allowing us into their properties. The electronic expertise (building customised GPS collars) of Jagath Ekanayake, MWLR, also made the project possible. We thank Travis Ashcroft, MPI, and Brent Glentworth and Graham Sullivan, Environment Canterbury (ECan), for project advice and logistical support; Morgan Coleman, Grant Morriss, Oscar Pollard, and Ivor Yockney, MWLR, for field assistance; Corrie Tegelaars for ground-netting wallabies; and Craig McMillan, Heliventures New Zealand, and Tony McNutt, Mainland Vector Contracting, for aerial capture of wallabies. Finally, we thank Mandy Barron (MWLR), Chris Jones (MWLR), Ray Prebble (MWLR), Brent Glentworth (ECan), Travis Ashcroft (MPI), the NWEP research advisory group, and two anonymous reviewers and an associate editor for their input into earlier versions of this manuscript.
References
Arnold, GW, Steven, DE, and Weeldenburg, JR (1989). The use of surrounding farmland by western grey kangaroos living in a remnant of Wandoo woodland and their impact on crop production. Wildlife Research 16, 85–93.| The use of surrounding farmland by western grey kangaroos living in a remnant of Wandoo woodland and their impact on crop production.Crossref | GoogleScholarGoogle Scholar |
Ball, SJ, Ramsey, D, Nugent, G, Warburton, B, and Efford, M (2005). A method for estimating wildlife detection probabilities in relation to home-range use: insights from a field study on the common brushtail possum (Trichosurus vulpecula). Wildlife Research 32, 217–227.
| A method for estimating wildlife detection probabilities in relation to home-range use: insights from a field study on the common brushtail possum (Trichosurus vulpecula).Crossref | GoogleScholarGoogle Scholar |
Bates, D, Mächler, M, Bolker, B, and Walker, S (2015). Fitting linear mixed-effects models using lme4. Journal of Statistical Software 67, 1–48.
| Fitting linear mixed-effects models using lme4.Crossref | GoogleScholarGoogle Scholar |
Bjørneraas, K, Van Moorter, B, Rolandsen, CM, and Herfindal, I (2010). Screening global positioning system location data for errors using animal movement characteristics. The Journal of Wildlife Management 74, 1361–1366.
| Screening global positioning system location data for errors using animal movement characteristics.Crossref | GoogleScholarGoogle Scholar |
Boyce, MS (2006). Scale for resource selection functions. Diversity and Distributions 12, 269–276.
| Scale for resource selection functions.Crossref | GoogleScholarGoogle Scholar |
Boyce, MS, Vernier, PR, Nielsen, SE, and Schmiegelow, FKA (2002). Evaluating resource selection functions. Ecological Modelling 157, 281–300.
| Evaluating resource selection functions.Crossref | GoogleScholarGoogle Scholar |
Boyle, SA, Lourenco, WC, da Silva, LR, and Smith, AT (2008). Home range estimates vary with sample size and methods. Folia Primatologica 80, 33–42.
| Home range estimates vary with sample size and methods.Crossref | GoogleScholarGoogle Scholar |
Burnham KP, Anderson DR (2002) ‘Model selection and multimodel inference: a practical information-theoretic approach.’ 2nd edn. (Springer: New York, NY, USA)
Calenge, C (2006). The package “adehabitat” for the R software: a tool for the analysis of space and habitat use by animals. Ecological Modelling 197, 516–519.
| The package “adehabitat” for the R software: a tool for the analysis of space and habitat use by animals.Crossref | GoogleScholarGoogle Scholar |
Casper, RM (2009). Guidelines for the instrumentation of wild birds and mammals. Animal Behaviour 78, 1477–1483.
| Guidelines for the instrumentation of wild birds and mammals.Crossref | GoogleScholarGoogle Scholar |
Catt DC (1975) Growth, reproduction and mortality in Bennett’s wallaby (Macropus rufogriseus fruticus) in South Canterbury, New Zealand. MSc Thesis, University of Canterbury, Christchurch, New Zealand.
Catt, DC (1977). The breeding biology of Bennett’s wallaby (Macropus rufogriseus fruticus) in South Canterbury, New Zealand. New Zealand Journal of Zoology 4, 401–411.
| The breeding biology of Bennett’s wallaby (Macropus rufogriseus fruticus) in South Canterbury, New Zealand.Crossref | GoogleScholarGoogle Scholar |
Côté, SD, Rooney, TP, Tremblay, J-P, Dussault, C, and Waller, DM (2004). Ecological impacts of deer overabundance. Annual Review of Ecology, Evolution, and Systematics 35, 113–147.
| Ecological impacts of deer overabundance.Crossref | GoogleScholarGoogle Scholar |
Dolman, PM, and Wäber, K (2008). Ecosystem and competition impacts of introduced deer. Wildlife Research 35, 202–214.
| Ecosystem and competition impacts of introduced deer.Crossref | GoogleScholarGoogle Scholar |
Efford, M (2004). Density estimation in live-trapping studies. Oikos 106, 598–610.
| Density estimation in live-trapping studies.Crossref | GoogleScholarGoogle Scholar |
Ekanayake, JC, and Hedley, CB (2018). Advances in information provision from wireless sensor networks for irrigated crops. Wireless Sensor Network 10, 71–92.
| Advances in information provision from wireless sensor networks for irrigated crops.Crossref | GoogleScholarGoogle Scholar |
Eldridge MDB, Coulson GM (2015) Family Macropodidae (kangaroos and wallabies). In ‘Handbook of mammals of the world. Vol. 5’. (Eds DE Wilson, RA Mittermeier) pp. 630–735. (Lynx Edicions: Barcelona, Spain)
ESRI (Environmental Systems Research Institute) (2022) ‘ArcGIS: release 2.9.3.’ (Environmental Systems Research Institute (ESRI): Redlands, CA, USA)
Fieberg, J, Matthiopoulos, J, Hebblewhite, M, Boyce, MS, and Frair, JL (2010). Correlation and studies of habitat selection: problem, red herring or opportunity? Philosophical Transactions of the Royal Society B: Biological Sciences 365, 2233–2244.
| Correlation and studies of habitat selection: problem, red herring or opportunity?Crossref | GoogleScholarGoogle Scholar |
Forester, JD, Im, HK, and Rathouz, PJ (2009). Accounting for animal movement in estimation of resource selection functions: sampling and data analysis. Ecology 90, 3554–3565.
| Accounting for animal movement in estimation of resource selection functions: sampling and data analysis.Crossref | GoogleScholarGoogle Scholar |
Frair, JL, Fieberg, J, Hebblewhite, M, Cagnacci, F, DeCesare, NJ, and Pedrotti, L (2010). Resolving issues of imprecise and habitat-biased locations in ecological analyses using GPS telemetry data. Philosophical Transactions of the Royal Society B: Biological Sciences 365, 2187–2200.
| Resolving issues of imprecise and habitat-biased locations in ecological analyses using GPS telemetry data.Crossref | GoogleScholarGoogle Scholar |
Garnick, S, and Coulson, G (2021). Population persistence, breeding seasonality and sexual dimorphism in the red-necked wallaby. Australian Mammalogy 43, 290–299.
| Population persistence, breeding seasonality and sexual dimorphism in the red-necked wallaby.Crossref | GoogleScholarGoogle Scholar |
Garnick, S, Di Stefano, J, Elgar, MA, and Coulson, G (2014). Inter- and intraspecific effects of body size on habitat use among sexually-dimorphic macropodids. Oikos 123, 984–992.
| Inter- and intraspecific effects of body size on habitat use among sexually-dimorphic macropodids.Crossref | GoogleScholarGoogle Scholar |
Garnick, S, Di Stefano, J, Elgar, MA, and Coulson, G (2016). Ecological specialisation in habitat selection within a macropodid herbivore guild. Oecologia 180, 823–832.
| Ecological specialisation in habitat selection within a macropodid herbivore guild.Crossref | GoogleScholarGoogle Scholar |
Gillies, CS, Hebblewhite, M, Nielsen, SE, Krawchuk, MA, Aldridge, CL, Frair, JL, Saher, DJ, Stevens, CE, and Jerde, CL (2006). Application of random effects to the study of resource selection by animals. Journal of Animal Ecology 75, 887–898.
| Application of random effects to the study of resource selection by animals.Crossref | GoogleScholarGoogle Scholar |
Girard, I, Ouellet, J-P, Courtois, R, Dussault, C, and Breton, L (2002). Effects of sampling effort based on GPS telemetry on home-range size estimations. Journal of Wildlife Management 66, 1290–1300.
| Effects of sampling effort based on GPS telemetry on home-range size estimations.Crossref | GoogleScholarGoogle Scholar |
Gordon, I, Snape, M, Fletcher, D, Howland, B, Coulson, G, Festa-Bianchet, M, Caley, P, McIntyre, S, Pople, T, Wimpenny, C, Baines, G, and Alcock, D (2021). Herbivore management for biodiversity conservation: a case study of kangaroos in the Australian Capital Territory (ACT). Ecological Management & Restoration 22, 124–137.
| Herbivore management for biodiversity conservation: a case study of kangaroos in the Australian Capital Territory (ACT).Crossref | GoogleScholarGoogle Scholar |
Harper, GA, van Dinther, M, Russell, JC, and Bunbury, N (2015). The response of black rats (Rattus rattus) to evergreen and seasonally arid habitats: informing eradication planning on a tropical island. Biological Conservation 185, 66–74.
| The response of black rats (Rattus rattus) to evergreen and seasonally arid habitats: informing eradication planning on a tropical island.Crossref | GoogleScholarGoogle Scholar |
Hone J (2007) ‘Wildlife damage control.’ (CSIRO Publishing: Melbourne, Vic., Australia)
Howard WE (1958) ‘The rabbit problem in New Zealand.’ (Department of Scientific and Industrial Research Information Series 16: Wellington, New Zealand)
Jaksic, FM (1998). Vertebrate invaders and their ecological impacts in Chile. Biodiversity and Conservation 7, 1427–1445.
| Vertebrate invaders and their ecological impacts in Chile.Crossref | GoogleScholarGoogle Scholar |
Johnson, DH (1980). The comparison of usage and availability measurements for evaluating resource preference. Ecology 61, 65–71.
| The comparison of usage and availability measurements for evaluating resource preference.Crossref | GoogleScholarGoogle Scholar |
Johnson, CN (1987). Macropod studies at Wallaby Creek. 4. Home range and movements of the red-necked wallaby. Australian Wildlife Research 14, 125–132.
| Macropod studies at Wallaby Creek. 4. Home range and movements of the red-necked wallaby.Crossref | GoogleScholarGoogle Scholar |
Johnson, CN (1989). Grouping and the structure of association in the red-necked wallaby. Journal of Mammalogy 70, 18–26.
| Grouping and the structure of association in the red-necked wallaby.Crossref | GoogleScholarGoogle Scholar |
Johnson, CJ, Nielsen, SE, Merrill, EH, McDonald, TL, and Boyce, MS (2006). Resource selection functions based on use–availability data: theoretical motivation and evaluation methods. Journal of Wildlife Management 70, 347–357.
| Resource selection functions based on use–availability data: theoretical motivation and evaluation methods.Crossref | GoogleScholarGoogle Scholar |
Latham ADM (2019) A prioritisation roadmap for the detectability and home range parameters, g0 and sigma, for New Zealand’s invasive mammalian predators. Manaaki Whenua – Landcare Research Contract Report LC3647 for Taranaki Taku Tūranga, Lincoln, New Zealand.
Latham ADM, Warburton B (2021) Notamacropus eugenii, N. r. rufogriseus, N. parma, N. dorsalis, Petrogale penicillata, Wallabia bicolor. Family Macropodidae. In ‘The handbook of New Zealand mammals’. 3rd edn. (Eds CM King, DM Forsyth) pp. 1–26. (CSIRO Publishing: Melbourne, Vic., Australia)
Latham, ADM, Latham, MC, Knopff, KH, Hebblewhite, M, and Boutin, S (2013). Wolves, white-tailed deer, and beaver: implications of seasonal prey switching for woodland caribou declines. Ecography 36, 1276–1290.
| Wolves, white-tailed deer, and beaver: implications of seasonal prey switching for woodland caribou declines.Crossref | GoogleScholarGoogle Scholar |
Latham, ADM, Latham, MC, Anderson, DP, Cruz, J, Herries, D, and Hebblewhite, M (2015). The GPS craze: six questions to address before deciding to deploy GPS technology on wildlife. New Zealand Journal of Ecology 39, 143–152.
Latham, ADM, Latham, MC, Nugent, G, Smith, J, and Warburton, B (2016). Refining operational practice for controlling introduced European rabbits on agricultural lands in New Zealand. PLoS ONE 11, .
| Refining operational practice for controlling introduced European rabbits on agricultural lands in New Zealand.Crossref | GoogleScholarGoogle Scholar |
Latham, ADM, Latham, MC, and Warburton, B (2019). Current and predicted future distributions of wallabies in mainland New Zealand. New Zealand Journal of Zoology 46, 31–47.
| Current and predicted future distributions of wallabies in mainland New Zealand.Crossref | GoogleScholarGoogle Scholar |
Latham, ADM, Latham, MC, Norbury, GL, Forsyth, DM, and Warburton, B (2020a). A review of the damage caused by invasive wild mammalian herbivores to primary production in New Zealand. New Zealand Journal of Zoology 47, 20–52.
| A review of the damage caused by invasive wild mammalian herbivores to primary production in New Zealand.Crossref | GoogleScholarGoogle Scholar |
Latham, ADM, Davidson, B, Warburton, B, Yockney, I, and Hampton, JO (2020b). Efficacy and animal welfare impacts of novel capture methods for two species of invasive wild mammals in New Zealand. Animals 10, .
| Efficacy and animal welfare impacts of novel capture methods for two species of invasive wild mammals in New Zealand.Crossref | GoogleScholarGoogle Scholar |
Latham, ADM, Warburton, B, Latham, MC, Anderson, DP, Howard, SW, and Binny, RN (2021). Detection probabilities and surveillance sensitivities for managing an invasive mammalian herbivore. Ecosphere 12, .
| Detection probabilities and surveillance sensitivities for managing an invasive mammalian herbivore.Crossref | GoogleScholarGoogle Scholar |
Leblond, M, Frair, J, Fortin, D, Dussault, C, Ouellet, J-P, and Courtois, R (2011). Assessing the influence of resource covariates at multiple spatial scales: an application to forest-dwelling caribou faced with intensive human activity. Landscape Ecology 26, 1433–1446.
| Assessing the influence of resource covariates at multiple spatial scales: an application to forest-dwelling caribou faced with intensive human activity.Crossref | GoogleScholarGoogle Scholar |
le Mar, K, and McArthur, C (2000). Relocating radio-collared targeted marsupials after a 1080-poisoning operation. Tasforests 12, 155–160.
le Mar, K, and McArthur, C (2001). Changes in marsupial herbivore densities in relation to a forestry 1080-poisoning operation. Australian Forestry 64, 175–180.
| Changes in marsupial herbivore densities in relation to a forestry 1080-poisoning operation.Crossref | GoogleScholarGoogle Scholar |
le Mar, K, and McArthur, C (2005). Comparison of habitat selection by two sympatric macropods, Thylogale billardierii and Macropus rufogriseus rufogriseus, in a patchy eucalypt–forestry environment. Austral Ecology 30, 674–683.
| Comparison of habitat selection by two sympatric macropods, Thylogale billardierii and Macropus rufogriseus rufogriseus, in a patchy eucalypt–forestry environment.Crossref | GoogleScholarGoogle Scholar |
le Mar, K, McArthur, C, and Statham, M (2003). Home ranges of sympatric red-necked wallabies, red-bellied pademelons and common brushtail possums in a temperate eucalypt forestry environment. Australian Mammalogy 25, 183–191.
| Home ranges of sympatric red-necked wallabies, red-bellied pademelons and common brushtail possums in a temperate eucalypt forestry environment.Crossref | GoogleScholarGoogle Scholar |
Long JL (2003) ‘Introduced mammals of the world.’ (CSIRO Publishing: Melbourne, Vic., Australia)
Mackenzie, HR, Latham, MC, Anderson, DP, Hartley, S, Norbury, GL, and Latham, ADM (2022). Detection parameters for managing invasive rats in urban environments. Scientific Reports 12, .
| Detection parameters for managing invasive rats in urban environments.Crossref | GoogleScholarGoogle Scholar |
Manly BFJ, McDonald LL, Thomas DL, McDonald TL, Erickson WP (2002) ‘Resource selection by animals: statistical design and analysis for field studies.’ 2nd edn. (Kluwer: Boston, MA, USA)
McLeod S (1986) The feeding and behavioural interaction of Bennett’s wallaby (Macropus rufogriseus rufogriseus) with domestic sheep (Ovis aries) in South Canterbury, New Zealand. MSc Thesis, University of Canterbury, Christchurch, New Zealand.
Montgomery DC, Peck EA (1992) ‘Introduction to linear regression analysis.’ (Wiley: New York, NY, USA)
Morrison, SA, Macdonald, N, Walker, K, Lozier, L, and Shaw, MR (2007). Facing the dilemma at eradication’s end: uncertainty of absence and the Lazarus effect. Frontiers in Ecology and the Environment 5, 271–276.
| Facing the dilemma at eradication’s end: uncertainty of absence and the Lazarus effect.Crossref | GoogleScholarGoogle Scholar |
Nathan, R (2001). The challenges of studying dispersal. Trends in Ecology & Evolution 16, 481–483.
| The challenges of studying dispersal.Crossref | GoogleScholarGoogle Scholar |
Nugent, G, Warburton, B, Thomson, C, Cross, ML, and Coleman, MC (2012). Bait aggregation to reduce cost and toxin use in aerial 1080 baiting of small mammal pests in New Zealand. Pest Management Science 68, 1374–1379.
| Bait aggregation to reduce cost and toxin use in aerial 1080 baiting of small mammal pests in New Zealand.Crossref | GoogleScholarGoogle Scholar |
Owens, B (2017). Behind New Zealand’s wild plan to purge all pests. Nature 541, 148–150.
| Behind New Zealand’s wild plan to purge all pests.Crossref | GoogleScholarGoogle Scholar |
Parkes, J, and Murphy, E (2003). Management of introduced mammals in New Zealand. New Zealand Journal of Zoology 30, 335–359.
| Management of introduced mammals in New Zealand.Crossref | GoogleScholarGoogle Scholar |
Parkes, JP, Nugent, G, Forsyth, DM, Byrom, AE, Pech, RP, Warburton, B, and Choquenot, D (2017). Past, present and two potential futures for managing New Zealand’s mammalian pests. New Zealand Journal of Ecology 41, 151–161.
| Past, present and two potential futures for managing New Zealand’s mammalian pests.Crossref | GoogleScholarGoogle Scholar |
Putman, RJ, and Moore, NP (1998). Impact of deer in lowland Britain on agriculture, forestry and conservation habitats. Mammal Review 28, 141–164.
| Impact of deer in lowland Britain on agriculture, forestry and conservation habitats.Crossref | GoogleScholarGoogle Scholar |
R Development Core Team (2022) ‘R: a language and environment for statistical computing, version 4.1.3.’ (R Foundation for Statistical Computing: Vienna, Austria) Available at http://www.R-project.org
Riney, T (1956). Comparison of occurrence of introduced animals with critical conservation areas to determine priorities for control. New Zealand Journal of Science and Technology 38B, 1–18.
Russell, JC, and Holmes, ND (2015). Tropical island conservation: rat eradication for species recovery. Biological Conservation 185, 1–7.
| Tropical island conservation: rat eradication for species recovery.Crossref | GoogleScholarGoogle Scholar |
Russell, JC, Innes, JG, Brown, PH, and Byrom, AE (2015). Predator-free New Zealand: conservation country. BioScience 65, 520–525.
| Predator-free New Zealand: conservation country.Crossref | GoogleScholarGoogle Scholar |
Studholme EC (1954) ‘Te Waimate: early station life in New Zealand.’ (AH and AW Reed: Dunedin, New Zealand)
Thomson GM (1922) ‘The naturalisation of animals & plants in New Zealand.’ (Cambridge University Press: Cambridge, UK)
Warburton, B (1986). Wallabies in New Zealand: history, current status, research, and management needs. Forest Research Institute Bulletin 114, 1–29.
Warburton B, Latham ADM (2020) National wallaby research plan June 2020. Manaaki Whenua – Landcare Research Contract Report LC3777 for the New Zealand Ministry for Primary Industries, Lincoln, New Zealand.
Wiggins, NL, and Bowman, DMJS (2011). Macropod habitat use and response to management interventions in an agricultural–forest mosaic in north-eastern Tasmania as inferred by scat surveys. Wildlife Research 38, 103–113.
| Macropod habitat use and response to management interventions in an agricultural–forest mosaic in north-eastern Tasmania as inferred by scat surveys.Crossref | GoogleScholarGoogle Scholar |
Wiggins, NL, Williamson, GJ, McCallum, HI, McMahon, CR, and Bowman, DMJS (2010). Shifts in macropod home ranges in response to wildlife management interventions. Wildlife Research 37, 379–391.
| Shifts in macropod home ranges in response to wildlife management interventions.Crossref | GoogleScholarGoogle Scholar |
Worton, BJ (1989). Kernel methods for estimating the utilization distribution in home-range studies. Ecology 70, 164–168.
| Kernel methods for estimating the utilization distribution in home-range studies.Crossref | GoogleScholarGoogle Scholar |