

Animal welfare outcomes of professional vehicle-based shooting of peri-urban rusa deer in Australia
Jordan O. Hampton A B F * , Darryl I. MacKenzie C and David M. Forsyth D E
A B F * , Darryl I. MacKenzie C and David M. Forsyth D E
A Ecotone Wildlife, PO Box 76, Inverloch, Vic. 3096, Australia.
B Harry Butler Institute, Murdoch University, 90 South Street, WA 6150, Australia.
C Proteus, PO Box 7, Outram 9062, New Zealand.
D Vertebrate Pest Research Unit, NSW Department of Primary Industries, 1447 Forest Road, Orange, NSW 2800, Australia.
E School of Biological, Earth and Environmental Sciences, University of New South Wales, Sydney, NSW 2052, Australia.
F Present address: Faculty of Veterinary and Agricultural Sciences, University of Melbourne, Parkville, Vic. 3052, Australia.
Wildlife Research 50(9) 603-616 https://doi.org/10.1071/WR21131
Submitted: 6 September 2021 Accepted: 28 December 2021 Published: 5 May 2022
© 2023 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Context: Vehicle-based shooting has been widely used to kill deer, but the animal-welfare outcomes of this technique have not been evaluated in Australasia.
Aim: To assess the animal-welfare outcomes of peri-urban deer culling by quantifying the fates of deer seen and shot at, the duration of procedures, and the number and location of bullet wounds in deer.
Methods: We assessed vehicle-based night shooting of peri-urban rusa deer (Cervus timorensis) by professional contractors in eastern Australia. Shooters targeted the heads of deer using .223 Remington® rifles and 55 grain bullets. Independent veterinarians conducted ante-mortem (i.e. from the shooting vehicle) and post-mortem (i.e. inspecting the carcass) observations. The ante-mortem data were used to estimate the proportion of deer seen that were shot at, killed, wounded, and escaped. The influence of variables predicted to affect shooting outcomes was assessed. The numbers and locations of bullet wounds were recorded post-mortem.
Key results: Of the 269 deer seen in 21 nights, 48% were shot at and 85% of those shot at were killed by either one (87%), two (10%) or three (3%) shots. The frequency of non-fatal wounding (i.e. escaping wounded) was 3.5% for those shot at and hit, and the median time to insensibility for the deer that were shot multiple times was 289 s. There was variation among shooters in their ability to hit a deer, and also to do so with a killing shot. The number of bullet wounds per deer ranged from 1 to 3 (mean = 1.1), with 83% of shots striking the brain and 17% striking the anterior skull, neck and jaw.
Conclusions: The animal welfare outcomes we observed were comparable to those reported from other professional ground-based shooting programs for ungulates, but were poorer than those reported for professional ground-based shooting of peri-urban kangaroos.
Implications: Our results suggest that one way to improve the animal welfare outcomes of vehicle-based shooting of peri-urban deer is by improving shooter training. Assessment of shooter performance should be a routine part of ground-based shooting programs.
Keywords: Cervids, culling, invasive species, pest management, population control, ungulates, urban ecology, wildlife management.
Introduction
Wild deer can have a wide range of undesirable economic, social, environmental and human-health impacts (Côté et al. 2004; Conover 2011; Valente et al. 2020). In Australia and New Zealand, where deer are non-native and many populations are increasing in range and abundance (Moriarty 2004a; King and Forsyth 2021), the undesirable impacts of deer include altered native vegetation communities (Wardle et al. 2001; Burns et al. 2021), reduced profitability of agricultural enterprises (Lindeman and Forsyth 2008; Page et al. 2017; Cripps et al. 2019; Latham et al. 2020), vehicle collisions (Burgin et al. 2015; Davies et al. 2020), and pollution of drinking-water catchments (Bennett et al. 2015).
One approach to reducing the undesirable impacts of deer is to remove deer by lethal control. Ground-based shooting has been used to kill deer and other ungulate species in Australia and New Zealand for many decades (Caughley 1983; Moriarty 2004a; Bennett et al. 2015). Ground-based shooting is undertaken by either professionals (i.e. remunerated) or volunteers (unpaid ‘recreational hunters’ or ‘sporting shooters’). Shooting deer is sometimes contentious because of concerns about animal welfare outcomes (Rutberg 1997; Aebischer et al. 2014; DeNicola et al. 2019), concerns that tend to be exacerbated when shooting occurs in and around human settlements (Curtis et al. 1995; Moriarty 2004b; Burgin et al. 2015). However, the welfare outcomes of ground-based shooting of deer have not been evaluated in Australasia (Forsyth et al. 2017).
The most robust assessments of the animal welfare outcomes of lethal control methods such as shooting have five features (reviewed in Smith and Ryeng 2022). First, independent observers, rather than the shooters themselves, collect the data (so as to minimise potential biases; Schulz et al. 2013). Second, a sufficient number of shooting events are observed so that the frequency of adverse animal welfare events can be robustly estimated (Hampton et al. 2019a). Third, ante-mortem (before death) and post-mortem (after death) data are combined to quantify the following two parameters (Ryeng and Larsen 2021): (i) the duration of suffering from shooting prior to insensibility or death, and (ii) the frequency of immediate insensibility. Fourth, the frequency of non-fatal wounding, considered the worst animal welfare outcome for any shooting program because it causes protracted (but unmeasured) suffering, is quantified (Aebischer et al. 2014). Fifth, the numbers and anatomical locations of bullet wounds, related to the accuracy of shooting, are reported.
The approach described above, in which ante-mortem and post-mortem data are combined to assess shooting methods, was first developed for whaling practices in Scandinavia in the 1980s (Øen 2021). It has since been applied to vehicle-based shooting of impala (Aepyceros melampus) in southern Africa (Lewis et al. 1997) and to boat-based shooting of seals in Canada (Daoust and Caraguel 2012; Hampton et al. 2021a) and Norway (Ryeng and Larsen 2021). In Australia, the approach has been used to assess the welfare outcomes of aerial (helicopter-based) shooting of large ungulates (Hampton et al. 2014, 2017), and vehicle-based shooting of European rabbits (Oryctolagus cuniculus; Hampton et al. 2015, 2016a, 2020) and eastern grey kangaroos (Macropus giganteus; Hampton and Forsyth 2016).
Most quantitative assessments of animal welfare outcomes for ground-based shooting of deer have been conducted on red deer (Cervus elaphus) in the United Kingdom. Using independent observations of recreational shooters, Bradshaw and Bateson (2000) concluded that ground-based shooting killed 93% of deer within 2 min of being shot at and hit and that 2% escaped wounded. Cockram et al. (2011) used biochemical stress markers (e.g. plasma cortisol concentrations) and carcase characteristics to assess four red deer shooting methods. Two other studies used post-mortem data to describe the numbers and locations of bullet wounds (i.e. inference about ante-mortem outcomes could not be made; Urquhart and McKendrick 2003, 2006). Another indirect method applied to four deer species in the United Kingdom was to collect data from interviews with shooters (Aebischer et al. 2014). Other studies have provided information about particular outcomes of relevance to animal welfare, including ‘flight distance’ for deer and other large mammals, defined as the distance animals run between shooting and incapacitation for animals shot in the thorax (chest-shooting; Kanstrup et al. 2016; Stokke et al. 2018). None of these studies combined ante-mortem and post-mortem data collected by an independent observer to quantify key animal welfare outcomes.
Several variables could potentially affect the welfare outcomes of ground-based shooting programs. The precision of a firearm/optics/ammunition configuration is central to the ability of a shooter to strike a desired anatomical zone (Hampton et al. 2021a). Rifle calibre and ammunition characteristics (bullet mass and powder load) determine the amount of kinetic energy transferred to the target, which declines with an increasing shooting distance (Aebischer et al. 2014; Hampton et al. 2016a). Shooting distance has been shown to be an important determinant of welfare outcomes for ground-based shooting of deer (Aebischer et al. 2014) and other mammals, such as, for example, European rabbits (Hampton et al. 2015). The mass of the target animal is also likely to be important because, all other things being equal, heavier individuals are less likely to be killed as quickly as are smaller individuals (Stokke et al. 2018). Because many deer species are strongly sexually dimorphic, with adult males being larger than adult females (Weckerly 1998), welfare outcomes might differ between those age–sex classes. Welfare outcomes are also likely to vary among shooters because of consistent individual differences in target acquisition and shooting accuracy, even after accounting for shooting distance (Aebischer et al. 2014; Hampton et al. 2014, 2017). For example, Aebischer et al. (2014) reported that the probability of a shot hitting its target for red deer stalking in the United Kingdom was higher for shooters who practiced at least once a week and for those shooters with an advanced certificate or qualification in deer stalking.
One aspect of the welfare outcomes of ground-based shooting that has not, to our knowledge, been previously assessed is the number of animals shot from the group. A procedural document for ground-based shooting of deer (Sharp 2013) proposed that, when possible, all deer in a group should be killed before another group is targeted, and that the smallest groups should be targeted first by shooters. It is not known how feasible this advice is for deer. Following the discharge of a rifle, other deer in the group are likely to flee (Reimoser 2012). Hence, as deer group size increases, it is expected that the proportion of deer that escape will increase. Such a relationship is described by the functional response (Hone 1990, 1994), the form of which depends on (i) a threshold group size at which the total number of shots fired at deer in the group is saturated, and (ii) the effect of declining deer group size on the total number of shots fired at deer in the group. Evaluating this relationship for ground-based shooting of deer requires the number of deer in the group being shot at to be recorded, along with the shots fired at deer in the group (i.e. these are ante-mortem observations).
Here, we assess the animal welfare outcomes of vehicle-based shooting of peri-urban deer in eastern Australia. Our objective was to quantify (1) the fates of deer seen and shot at, (2) the duration of key procedures, and (3) the numbers and locations of bullet wounds in shot deer. We compare and contrast these results with similar studies conducted on ungulates and macropods. Finally, we make recommendations as to how the animal welfare outcomes of ground-based deer shooting programs could be improved.
Materials and methods
Study area and species
Our study was conducted in the Illawarra region (~80 km south of Sydney), New South Wales, Australia. The rusa deer (Cervus timorensis) population in this region is descended from animals released in Royal National Park in 1906 (Moriarty 2004c; Keith and Pellow 2005; Moriarty 2009). Rusa deer are the third-largest deer species present in Australia, with adult females and males weighing approximately 70 and 140 kg respectively (Moriarty 2004c). Native to tropical Southeast Asia, rusa deer can breed year-round; in our study area, most births occur in March–April (Bentley 1995). Young become independent at 3–4 months (Tuckwell 2003; New Zealand Government 2018). Rusa deer typically rest in thick cover during the day, emerging at or after dusk to feed on grasses in open areas (Moriarty 2004c). The only strong social bond in deer is that between the mother and her dependent offspring (Albon et al. 1992; Rosenberry et al. 2009), but temporary groups of up to 15 rusa deer can form in preferred habitats (see below).
A management program was established in the Illawarra region in 2011 to minimise the impacts of rusa deer on road and rail accidents, the damage to residential gardens and fences, competition with livestock through competition for forage, impacts on native plant species, and illegal hunting (Dawson 2017). In the Wollongong and Lake Illawarra region, there were 107 motor vehicle accidents involving deer during 2005–2017, of which 90 were rated as serious and resulted in 28 injuries and one death (Invasive Species Council 2018). Professional shooters were contracted to shoot deer in urban, peri-urban and rural areas. Land types include suburban streets, golf courses, ‘hobby’ farms and water catchments. Some areas are heavily infested with the introduced plant Lantana camara (Gooden et al. 2009), which rusa deer use as refuges.
Shooting procedure
Several procedural documents (i.e. standard operating procedures, codes of practice and manuals; Hampton et al. 2016b) guide the ground-based shooting of deer in Australia. There is a national model standard operating procedure for ground-based shooting of deer (Sharp 2013), but a unique methodology is often developed for a particular management program (Brown 2017). Stipulations within standard operating procedures govern several variables of potential relevance to animal welfare outcomes, including permitted shooting seasons, rifle calibre and bullet weight, and shooter training and proficiency.
For the program studied here, shooting was conducted during the hours of darkness (typically beginning 1 h after sunset and ending ~6 h later) during 2018 and 2019. Dry and calm (minimal wind) nights were selected for shooting, and shooting did not occur on weekends, public holidays or school holidays (Dawson 2017). The shooting teams consisted of three people (excluding the independent veterinarian observer) in dual-cab four-wheel drive utility vehicles. One team member drove the vehicle (up to 10 km hour−1) and two team members stood in the tray (truck bed); one was the shooter, and the other controlled the white-light spotlight (100 W, 240 mm diameter). Four shooters employed by professional pest control companies conducted the shooting. All shooters used bolt-action .223 Remington® rifles, fitted with telescopic scopes, and fired 55 grain lead-based soft-point bullets, identical to that typically used in professional kangaroo culling (Hampton and Forsyth 2016). Suppressors were used to reduce shooting noise (Williams et al. 2018). The shooters conducted their own precision testing (Hampton et al. 2021a) off-site prior to arriving at the study area (i.e. it was not observed by the independent veterinarian). As for kangaroo shooting (Commonwealth of Australia 2008), the only anatomical target was the cranium (‘head shooting’; Urquhart and McKendrick 2006). Dead deer were removed from the site by the shooting team.
Observations
We adapted the methods developed for observation of vehicle-based shooting of European rabbits (Hampton et al. 2015, 2016a, 2020) to enable ante-mortem and post-mortem data to be collected by independent veterinarians. Briefly, veterinarians experienced in collecting animal welfare data from wildlife shooting programs accompanied deer culling teams and sat in the front passenger seat of the shooting vehicles. Observers were independent in that they were not employees of the managing agency or the contracted culling companies. Three independent veterinarian observers collected data, but only one in each vehicle. The number of deer seen and shot at were recorded. Group size was defined as the number of animals initially seen together before a shot was fired at any group member, as used by Hampton and Forsyth (2016) for peri-urban kangaroo shooting. The number of shots fired at each deer was recorded.
One time-to-event parameter was quantified for each deer shot, namely, time to insensibility (‘unconsciousness’; TTI, seconds), which was the duration between initial shooting and insensibility (Hampton et al. 2021b). Insensibility was defined as the moment at which the animal ceased voluntary movements, including the absence of respiratory movements, with sudden relaxation of the body, as used previously in wildlife shooting studies (Lewis et al. 1997; Smith and Ryeng 2022). There is debate about the definition of insensibility in wildlife studies (Hampton and Forsyth 2016). In particular, recognising insensibility relies on distinguishing conscious or voluntary movements (e.g. mobile animals, lifting of the head in recumbent animals) from unconscious or involuntary movements (e.g. muscular twitches or kicking; Smith and Ryeng 2022). Autonomic movements that do not reflect sensibility typically occur in animals rendered insensible via cranial trauma, as has been observed in livestock studies (Claudia Terlouw et al. 2015). Observational studies of livestock slaughter and lethal wildlife control methods such as kill-traps typically use palpation to assess physiological responses (e.g. palpebral (blinking) reflexes; Littin et al. 2002). In contrast, studies of ballistic methods used to kill wildlife (Hampton et al. 2021a) rely on remote observation, and hence slightly different criteria for assessing insensibility have been applied (Knudsen 2005). There is more subjectivity in the application of remote methods of identifying insensibility and, hence, some studies have used terms such as ‘apparent time to death’ (Hampton et al. 2015) for this metric.
We then quantified the proportion of deer for which time to insensibility was zero. This parameter has been quantified in the same way in several similar studies but has sometimes been termed ‘instantaneously killed’ (Lewis et al. 1997), ‘instant immobilisation’ (Parker et al. 2006), ‘instant death’ (Hampton et al. 2014), ‘instant incapacitation’ (McTee et al. 2017) or ‘immediate collapse’ (Ryeng and Larsen 2021), with all terms having identical meaning. We reported the probability of being rendered immediately insensible, which is equivalent to ‘instantaneous death rate’ (Ryeng and Larsen 2021).
Shooting distance was estimated in one of two ways, namely, either through the use of a laser range finder (Pulsar Digisight N960®, Yukon Advanced Optics Worldwide, Vilnius, Lithuania) or by measuring the linear distance between a GPS waypoint saved where the shooting vehicle stopped, and a GPS waypoint saved where the targeted deer had been standing (Stokke et al. 2018).
Sample size
The desired sample size for our observations was guided by published statistical guidelines for animal welfare studies (Hampton et al. 2019a); we sought minimum ante-mortem and post-mortem sample sizes of 113, assuming a frequency of animal welfare outcomes of interest of ~5% (see Hampton and Forsyth 2016).
Statistical analyses
All statistical analyses were conducted using the software R version 3.6.6 software (R Core Team 2020, R: a language and environment for statistical computing; R Foundation for Statistical Computing, Vienna, Austria)).
Relationship between shots fired and deer group size
The relationship between number of shots fired and deer group size was evaluated using five models, three of which assumed non-linear ‘saturating’ relationships and two of which assumed linear relationships (Table 1). These types of model have been used to evaluate the ‘functional response’ between ungulates killed per unit effort and ungulate density for helicopter-based shooting of feral pigs (Sus scrofa; Hone 1990; Choquenot et al. 1999) and deer (Bengsen et al. 2022), but our objective was to assess how shots fired changed with deer group size because this relationship is of interest from an animal welfare perspective (Sharp 2013). All five models were fitted using the

|
Shooting outcomes
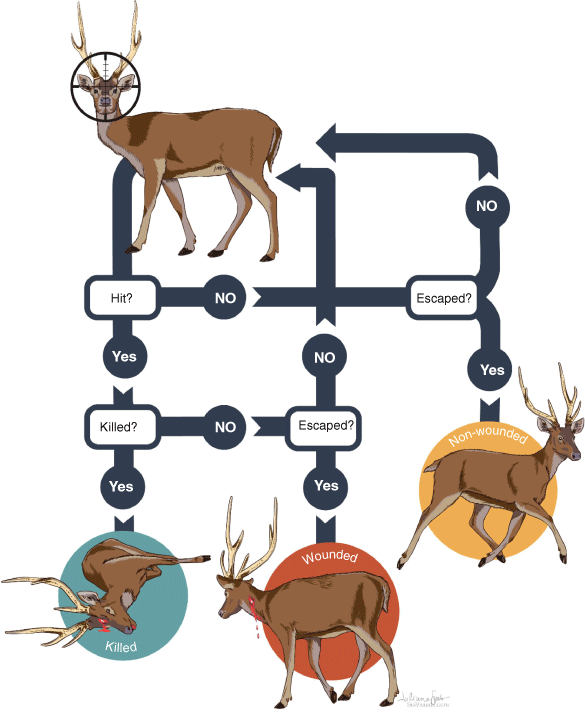
Shooting data were analysed using the same methods as used by Hampton et al. (2015), with the outcomes conceived as the results of a series of binary processes. The following three main outcomes can arise from any event in which a deer is shot at: the deer can be killed, escape wounded or escape non-wounded (Fig. 1). Each process has an associated probability: the probability of a shooter hitting a deer (pH), the probability the shot kills a deer that was hit (pK), and the probability of a deer escaping without being hit (pE). The probability of any observed outcome was expressed as the product of the respective probabilities for the relevant sequence. For example, if a shooter killed a deer with their first shot (K), then the probability for that outcome was expressed as:
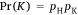
If a shooter’s first shot wounded a deer, their second shot missed, and the deer escaped before a third shot (i.e. the sequence WME), then the probability was expressed as:
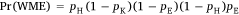
That is, the first shot hit but did not kill the deer (with probability pH (1 − pK)), the deer did not escape but the second shot missed ((1 − pE)(1 − pH)), and then the animal escaped wounded (pE). It was assumed that the outcomes for each deer were independent.
|
Influence of predictor variables
Each of the component probabilities (pH, pK and pE) was modelled as a function of predictor variables (hereinafter ‘predictors’), allowing the relative importance of different predictors of interest, and the effect of these predictors, to be estimated. A technique similar to logistic regression was used, in which probabilities were transformed on to the logit (or log-odds) scale for regression analysis. Incorporation of predictors into the modelling allows a probability to vary according to the value of associated predictors and estimated effect sizes. For example,
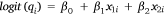
where qi is a probability for Shot i, x1i and x2i are the values of two predictors associated with Shot i (e.g. number of previous shots, or shooting distance) and β0, β1 and β2 are effect sizes (i.e. model parameters) to be estimated.
We included four predictors in this analysis (Table 2). Each combination of predictors defines a model that represents a different set of hypotheses about what factors affect the respective probability. Therefore, models that were identified as being ‘better’ indicate which set of hypotheses had greater support from the available data. We again used AIC to compare the fit of the models and perform model selection. AIC model weights (w) were calculated as a measure of support for each model and were summed across models as a measure of overall support for the importance of a predictor variable that was common to that set of models. Summed model weights (sx; for predictor variable x) were interpreted in terms of evidence ratios (ER; Burnham and Anderson 2002; Anderson 2007), as follows:
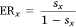

|
A two-stage approach was used to identify which combination of predictors was the most important. First, a separate model selection process was conducted for each component probability to avoid fitting a very large number of models to the data (when fitting all possible combinations of predictors for all combinations of probabilities). Predictors that had evidence ratios of >1 were considered in a second model selection process that involved fitting models with all combinations of the identified predictors for each probability, for all combinations of probabilities. Note that in the first round of model selection, a general structure was assumed for the two non-focal probabilities (e.g. for pE and pK if focusing on pH). The general structure involved all predictors that were being considered for that particular probability to allow variation in the non-focal probabilities should it be required. Model-averaging was again used to combine the results from each model included in the second round of model selection. Inference was made from these model-averaged estimates.
Time to insensibility
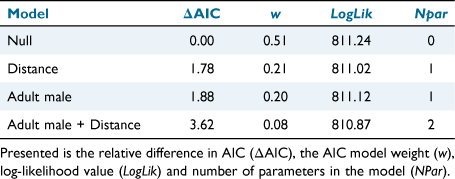
For those deer that were killed, we estimated the probability of being rendered immediately insensible. Cox’s proportional hazards models (Cox 1972) were used to evaluate whether time to insensibility was different for adult males (having larger mass) from that for other age–sex classes, and also whether there was a relationship with shooting distance. We expected that adult males would have a lower probability of being killed, and, conversely, a higher probability of being wounded, than for all other age–sex classes (Hampton et al. 2014). The following four models were fit to the data and compared using AIC: (1) null; (2) distance; (3) age–sex class, and; (4) age–sex class and distance. The proportional hazard quantifies the multiplicative effect on the mortality rate; hence, hazard values of <1 indicate lower mortality (higher survival) and values of >1 indicate higher mortality (lower survival).
Wound numbers and locations
For each deer that was shot and killed, the carcass was examined and its sex and age class (adult or juvenile, the latter including fawns and yearlings), and the number and location of bullet wound tracts were recorded. For logistical and safety reasons, the interval between shooting and inspection varied from 1 to 33 min. We assigned bullet wound locations on the basis of the peri-cranial anatomical zones (brain, neck, jaw, anterior skull) displaying the most damage, as was assigned for peri-urban kangaroos (Hampton and Forsyth 2016). Anatomical landmarks for the location of the brain were as per convention (Shearer and Ramirez 2012; Sharp 2013). Our definition of the anterior skull consisted of the orbit (eye sockets) and maxilla (upper jaw and snout). See DeNicola et al. (2019) for radiographic examination of ballistic trauma of cranial structures in head and neck shooting of deer. The probabilities of bullet wound tracts affecting these four anatomical structures were estimated. ‘Head shots’ (Urquhart and McKendrick 2006) were deemed to have occurred when bullets penetrated the brain (calvarium of the skull; Daoust and Caraguel 2012).
Results
Data scope
In total, 269 rusa deer were seen in 110 groups during the 21 nights of our study, with 148 shots being fired at 130 of those deer. The mean (±s.d.) shooting distance was 83 ± 49 m (range: 20–284 m).
Relationship between shots fired and deer group size
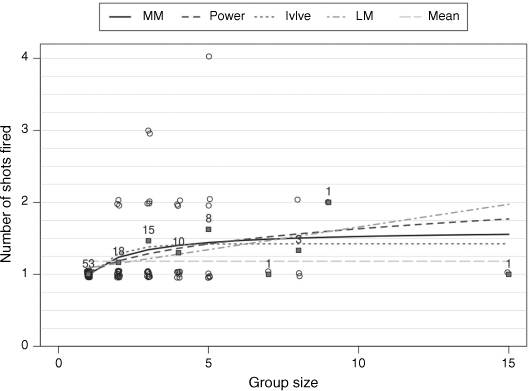
Deer group sizes ranged from 1 to 15, with one being the modal group size encountered by shooters (Table 3). On the basis of AIC, the Ivlev and Michaelis–Menten models were clearly better than were the other models (Table 3). These two models had similar support and showed similar non-linear relationships between the number of shots fired and deer group size (Fig. 2). The estimated mean (±s.e.) number of shots fired at a group size of one for the top-ranked Ivlev models was 1.00 ± 0.06, increasing rapidly to an asymptotic value of 1.56 ± 0.11. That is, even for rusa deer groups much larger than one, shooters fired an average of 1.6 shots at the group.

|
|
Shooting outcomes
Of the 269 deer seen, the probability that any one of them would be shot at was 0.48 (95% CI: 0.42–0.54). Of the deer shot at, 114 were hit (pH = 0.88; 0.81–0.93), and 16 escaped without being hit (pE = 0.12; 0.07–0.19). Of the deer that were hit, 110 were killed (pK = 0.97; 0.91–0.99). Of the deer that were killed, 96 (0.87; 0. 80–0.93) were rendered immediately insensible. The remaining deer had various outcomes, including 16 deer escaped without apparently being hit, 14 were killed after first being wounded, and four escaped wounded (Table 4). Of the 110 deer that were killed, only one carcass was not located, but this deer was assumed to have been killed because it was observed to be rendered immediately insensible with what appeared to be a head shot and then rolled down a steep hill into thick vegetation where the shooting team could not safely venture.

|
Influence of predictor variables
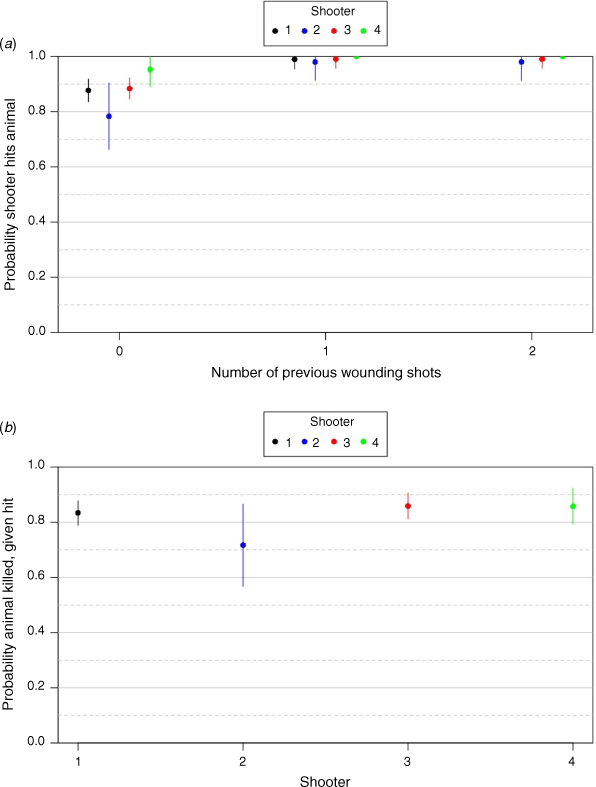
The first stage of model selection evaluating the influence of four predictor variables is summarised in Supplementary Tables S1–S6. The second stage of model selection indicated that the probability of hitting a deer (pH) varied with shooter ID and the number of previous wounding shots (Table 5, Fig. 3). There was variation among shooters in their ability to hit a deer (Fig. 3a), and also to do so with a killing shot (Fig. 3b). The probability of hitting a deer increased as the number of previous wounding shots increased. pH was close to 1 for all four shooters, except for if the deer had not been previously wounded (which includes when the first shot is taken). In that case, Shooter 2 was estimated to have the lowest mean (±s.e.) pH (0.78 ± 0.12), Shooters 1 and 3 had similar probabilities (0.88 ± 0.04), and Shooter 4 had the highest probability (0.95 ± 0.06; Fig. 3a). The probability of killing a deer that was hit was estimated to be lowest for Shooter 2 (0.72 ± 0.15), and similar for the other three shooters (approximately 0.85 ± 0.05). All deer that were missed by the first shot escaped; hence, pE was constrained to be 1 for this group of animals. Shooting distance was not an important predictor of the probability of hitting a deer.

|
|
Time to insensibility
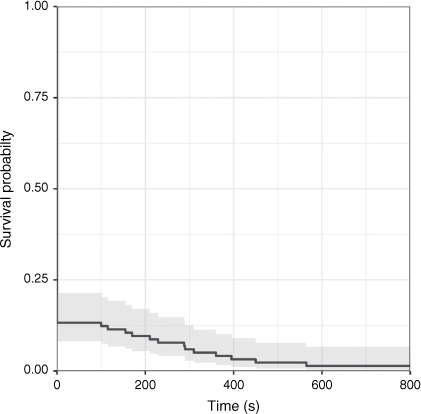
For the 14 deer that were killed but not rendered immediately insensible, the median TTI was 289 s (range: 100–840 s; Fig. 4). Our analysis indicated that the most-supported model did not include any predictors (wi = 0.51; Table 6). Distance and adult males as predictors explained little additional variation in the data, but the respective models had non-negligible AIC model weights because they had similar numbers of parameters to the best-supported model. The estimated effect sizes are in the direction we predicted (i.e. TTI increases with an increasing distance, and is higher for adult males than for other age–sex classes), although they are close to zero and have large standard errors.
|
|
Wound numbers and locations
The mean (±s.d.) duration from shooting to carcass inspection was 8 ± 7 min (range = 1–33 min). Of the 109 carcasses that were inspected, 31 animals were adult males, 45 were adult females, 17 were juvenile males and 16 were juvenile females. Bullet-wound tract counts showed that 95 deer were shot once, 11 deer were shot twice and three deer were shot three times, with the mean (±s.d.) number of times an animal was shot being 1.16 ± 0.43. Of 126 bullet-wound tracts examined, 83% (95% CI: 75–89%) of wound tracts indicated that the brain had been hit, 7% the anterior skull (3–13%), 5% the neck (2–11%), and 5% (1–10%) the jaw. No deer exhibited wounds to other parts of the body (i.e. thorax, abdomen or limbs). The mean (±s.d.) number of shots required to kill adult male rusa deer (1.23 ± 0.50) was only slightly higher than for other age–sex classes (i.e. adult female and all juveniles pooled; 1.14 ± 0.42).
Discussion
Many hundreds of thousands of deer have been shot at by ground-based shooters in Australasia (Caughley 1983; Harris 2002; Bengsen et al. 2020), but this is the first study to quantify the welfare outcomes of this technique. We found that professional vehicle-based shooters operating in a peri-urban environment shot at approximately half of the rusa deer that they encountered. However, regardless of the observed group size, only one deer was usually shot at. Of the deer that were killed, 13% required one or more subsequent shots. Hence, time to insensibility was usually zero. Of the deer that were shot at and hit, non-fatal wounding occurred in 3.5% of cases. The probability of hitting a deer increased as the number of previous wounding shots increased, presumably owing to increasing incapacitation of the deer. Animal welfare outcomes varied among shooters. Our post-mortem observations indicated that all deer had been shot in the head or neck, as was stipulated in the program plan.
Non-fatal wounding is the least desirable animal welfare outcome (Aebischer et al. 2014). Several factors could have contributed to four deer escaping wounded in this study. First, the practice of head shooting might have increased the risk of wounding because fatal haemorrhage is less likely than with chest-shooting (Stokke et al. 2018) if anatomical structures surrounding the brain (neck, jaw, anterior skull) are struck. In our study, 17% of deer that were killed and found were initially struck in these peri-cranial structures, and 74% of these deer required subsequent shots to the brain. For this reason, for deer shot in other contexts (e.g. aerial shooting), shooters are more likely to shoot at the thorax to incapacitate a deer rather than attempting a head shot (Sharp 2012; NSW National Parks and Wildlife Service 2020; Hampton et al. 2021b). Second, the flight responses of rusa deer made achieving a second shot for wounded deer difficult. Third, habitat features, particularly the dense Lantana thickets (Gooden et al. 2009), made wounded but mobile deer difficult for the shooting team to detect. Similar findings have been reported from the aerial shooting of chital deer (Axis axis) in Queensland, whereby wounded deer have escaped follow-up shooting by sheltering in thickets of rubber vine (Cryptostegia grandiflora; Hampton et al. 2021b).
Our data suggested that adult male rusa deer were slightly more difficult to render immediately insensible than were other age–sex classes. This was expected because of the larger size of male rusa deer. Stokke et al. (2018) demonstrated the importance of animal mass in determining flight distance for mammals shot in the thorax, with larger animals being less quickly incapacitated. Similar effects relating to male animals being more likely to have adverse welfare outcomes have been shown for aerial shooting of other sexually dimorphic ungulates, e.g. feral camels (Camelus dromedarius; Hampton et al. 2014).
The importance of the identity of shooters for animal welfare outcomes was also unsurprising. Shooters must anticipate and react to animal movement, place their shots accurately, and track and locate wounded animals (Hampton et al. 2021a). There is likely to be individual variation in these skills. Several other studies have demonstrated the importance of shooter identity for ground-based shooting of deer (Langenau 1986; Aebischer et al. 2014). This effect has also been shown for aerial shooting of feral camels (Hampton et al. 2014) and feral horses (Equus caballus; Hampton et al. 2017) in Australia. That there is an effect of shooter identity suggests that animal welfare outcomes in vehicle-based deer culling programs might be improved by screening or auditing shooters (Hampton et al. 2021a), and by providing training so that minimum welfare outcome standards are achieved (DeNicola et al. 2019).
The animal welfare outcomes reported here can be compared and contrasted with those reported for other ground-based shooting methods. Of most similarity to our study is the vehicle-based shooting of eastern grey kangaroos in a peri-urban environment (Hampton and Forsyth 2016). The frequency of adverse animal welfare events, including animals missed and non-fatally wounded, was higher for rusa deer in our study than for kangaroos. Median time to insensibility for rusa deer not rendered immediately insensible (289 s, n = 14) was much longer than for kangaroos (12 s, n = 3) and was likely to be related to the thick vegetation and steep terrain typically encountered in this study when compared with open and flat habitats typical of kangaroo shooting in the ACT (Hampton and Forsyth 2016). The frequency of immediate insensibility for deer hit (78%) was lower than that reported for vehicle-based shooting of kangaroos (98%; Hampton et al. 2019b) but was comparable to that for other professional shooting methods such as boat-based shooting of harp seals (Pagophilus groenlandicus; 84%; Ryeng and Larsen 2021), and exceeded that of other non-professional shooting methods such as vehicle-based shooting of European rabbits (60%; Hampton et al. 2015). Interspecific differences in the anatomy and bone density of the skulls of deer and kangaroos are also likely to have influenced shooting outcomes.
Ground-based and helicopter-based shooting are fundamentally different techniques, but the results derived here can be compared with those from aerial shooting of deer in eastern Australia (Hampton et al. 2021b). Unlike vehicle-based shooting, aerial shooting involves a pursuit phase, often involves repeat shooting, utilises much shorter shooting distances and typically uses the thorax rather than the head as the anatomical aim point. Accordingly, the frequency of immediate insensibility for deer hit in this study (78%) was substantially higher than for aerial shooting (<20%). Non-fatal wounding also occurred in aerial shooting (Hampton et al. 2021b), but the methods used to estimate this parameter were different from those applied in this study, precluding direct comparison.
Rusa deer seem to be more difficult to kill than are other mammals commonly subject to ground-based shooting in Australasia (e.g. macropods). The well developed flight response of rusa deer was reflected by the observation that no deer that were first missed were subsequently killed. That is, shooters only got one shot at a deer before it fled such that there was no opportunity for a subsequent shot. Groups of rusa deer also proved hard to kill; even for larger groups of deer, shooters managed to fire only an average of 1.6 shots at a group. This result suggests that it is infeasible to kill all deer in a group before targeting another group, as proposed by Sharp (2013). However, it may make sense to target small groups first when multiple groups are encountered (Sharp 2013). Indeed, the modal rusa deer group size in our study was one. Our findings about the difficulty of killing a deer that was missed, or other members of a group that was shot at, are consistent with rusa deer developing behavioural responses to ground-based shooting (such as increasing vigilance rates and staying close to escape cover) that have been documented in other deer species (e.g. fallow deer Dama dama; Pecorella et al. 2016). Our results are in stark contrast to those for eastern grey kangaroos, a species for which it is common for vehicle-based professional shooters to kill multiple, or all, animals from a social group (Hampton and Forsyth 2016). Understanding the functional response of ground-based shooters to increasing deer group size is important because functional responses determine relative effort required to kill a deer and, in turn, influence the costs associated with this deer control method (Van Deelen and Etter 2003; Bengsen et al. 2022).
The rifle and ammunition could have influenced the animal welfare outcomes we observed. The ammunition (55 grain bullets loaded in .223® Remington cartridges) delivered lower muzzle kinetic-energy levels (~1700 J) than those typically associated with deer shooting, for example, a legally stipulated minimum of 2373 J for red deer in Scotland (Urquhart and McKendrick 2006). The calibre of the rifle (.223) was smaller than that recommended by the national deer code of practice, which states ‘the minimum calibre is .243 with 100 grain projectile’ (Sharp 2013). The same prescription is made by the voluntary Model Code of Practice for the Welfare of Animals: Feral Livestock Animals: Destruction or Capture, Handling and Marketing (Standing Committee on Agriculture, Animal Health Committee 2002), which guides helicopter-based shooting of deer in some Australian jurisdictions (Hampton et al. 2021b). However, .223 calibre rifles and even lighter bullets (50 grain) have been used for vehicle-based professional shooting of deer using the head (and neck) as targets for Philippine deer (Rusa marianna), which are of a similar mass to that of rusa deer, in Guam (DeNicola et al. 2019). The shooting equipment used in the program we evaluated was chosen for community safety reasons (Dawson 2017). Lighter bullets delivering less kinetic energy carry several advantages for public safety and convenience. First, they are less likely to create pass-through shots whereby a bullet or its fragments may create an exit wound and continue to travel after striking a deer (DeNicola et al. 2019). Second, they are less likely to injure a person, domestic animal or damage property in the event of a missed shot. Third, they typically create less noise, a source of public nuisance in peri-urban shooting, which can also be reduced through the use of sound suppressors (Williams et al. 2018).
White-light illumination was used in the rusa deer shooting program, but thermal and night-vision rifle scopes are commonly used in ground-based deer shooting programs in Australia, for example, for sambar deer (Cervus unicolor) in Victoria (Brown 2017; Watter et al. 2020), and for peri-urban kangaroos in the Australian Capital Territory (Hampton and Forsyth 2016). The use of thermal or night vision might improve animal welfare outcomes through: (a) increasing the probability of detecting deer, including wounded deer; (b) reducing shooting distances because shooters can approach closer without being detected than with white-light (although our study did not show a strong effect of shooting distance on animal welfare outcomes); and (c) reducing flight responses from conspecifics (Hanson et al. 2019; Logan et al. 2019). The relatively weak influence of shooting distance on outcomes was somewhat surprising, given the strength of this variable in other published studies, for example, in Hampton et al. (2015).
Specialised techniques are often required for shooting in peri-urban areas to be safe, effective and socially acceptable (DeNicola et al. 2000). Managing deer in close proximity to high-density human settlements is a relatively recent challenge in Australia (Burgin et al. 2015), whereas methods used to manage peri-urban deer internationally (especially in the USA) have been refined over many decades (Kilpatrick et al. 1997; Curtis 2020). Methods used in the USA for shooting peri-urban deer that are not yet widely used in Australia include the use of suppressors (DeNicola and Williams 2008), bait to attract deer to shooting sites that are safer from a community safety perspective but more lethal for deer (Kilpatrick and Stober 2002), alternative anatomical aim points (e.g. neck shooting; DeNicola et al. 2019) and alternative bullet construction (e.g. copper bullets; McCann et al. 2016). These methods could improve the animal welfare outcomes of ground-based shooting programs in Australia. We encourage a quantitative approach to the evaluation of these methods.
Conclusions
Vehicle-based shooting of peri-urban rusa deer produced animal welfare outcomes comparable to those of professional ground-based shooting programs for ungulates, but the outcomes were poorer than those reported for professional ground-based shooting of kangaroos. The poorer outcomes for rusa deer than for kangaroos were likely to be related to differences in terrain, shooting distance and species susceptibility, especially their flight behaviours. Our study suggests that one way to improve the animal welfare outcomes of vehicle-based shooting of peri-urban deer is by improving shooter training. Assessment of shooter performance should be a routine part of ground-based shooting programs.
Supplementary material
Supplementary material is available online.
Data availability
The data that support this study cannot be publicly shared due to privacy reasons and may be shared upon reasonable request to the corresponding author if appropriate.
Conflicts of interest
David Forsyth is a guest Associate Editor of Wildlife Research but was blinded from the peer-review process for this paper.
Declaration of funding
This study was funded by the Centre for Invasive Species Solutions projects ‘Cost-effective management of wild deer’ (PO1-L-001) and ‘Management of wild dogs and deer in peri-urban landscapes: strategies for safe communities’ (PO1-L-003).
Acknowledgements
This was an observational study of operational pest animal management activities and did not require Animal Ethics Committee (AEC) approval. Brendan Cowled and Edwina Leslie assisted with data collection. Comments by Andrew Bengsen, Sebastien Comte, Anthony DeNicola and an anonymous reviewer improved the manuscript.
References
Aebischer, NJ, Wheatley, CJ, and Rose, HR (2014). Factors associated with shooting accuracy and wounding rate of four managed wild deer species in the UK, based on anonymous field records from deer stalkers. PLoS One 9, e109698.| Factors associated with shooting accuracy and wounding rate of four managed wild deer species in the UK, based on anonymous field records from deer stalkers.Crossref | GoogleScholarGoogle Scholar | 25334012PubMed |
Albon, SD, Staines, HJ, Guinness, FE, and Clutton-Brock, TH (1992). Density-dependent changes in the spacing behaviour of female kin in red deer. Journal of Animal Ecology 61, 131–137.
| Density-dependent changes in the spacing behaviour of female kin in red deer.Crossref | GoogleScholarGoogle Scholar |
Anderson DR (2007) ‘Model based inference in the life sciences: a primer on evidence.’ (Springer: New York City, NY, USA)
Bengsen, AJ, Forsyth, DM, Harris, S, Latham, ADM, McLeod, SR, and Pople, A (2020). A systematic review of ground-based shooting to control overabundant mammal populations. Wildlife Research 47, 197–207.
| A systematic review of ground-based shooting to control overabundant mammal populations.Crossref | GoogleScholarGoogle Scholar |
Bengsen, AJ, Forsyth, DM, Pople, T, Brennan, M, Amos, M, Leeson, M, Cox, ET, Gray, B, Orgill, O, Hampton, JO, Crittle, T, and Haebich, K (2022). Effectiveness and costs of helicopter‐based shooting of deer. Wildlife Research , .
Bennett, A, Haydon, S, Stevens, M, and Coulson, G (2015). Culling reduces fecal pellet deposition by introduced sambar (Rusa unicolor) in a protected water catchment: Culling Reduces Fecal Deposition by Sambar. Wildlife Society Bulletin 39, 268–275.
| Culling reduces fecal pellet deposition by introduced sambar (Rusa unicolor) in a protected water catchment: Culling Reduces Fecal Deposition by Sambar.Crossref | GoogleScholarGoogle Scholar |
Bentley A (1995) Fallow deer. In ‘The Mammals of Australia’. (Ed. R Strahan) pp. 732–733. (Australian Museum/Reed New Holland: Sydney, NSW, Australia)
Bradshaw, EL, and Bateson, P (2000). Welfare implications of culling red deer (Cervus elaphus). Animal Welfare 9, 3–24.
Brown D (2017) Tools for managing wild deer: ground shooting in the Alpine National Park Deer Control Trial. In ‘National Wild Deer Management Workshop Proceedings’. (Eds DM Forsyth, T Pople, B Page, T Moriarty, D Ramsey, J Parkes, A Wiebkin, C Lane) pp. 24–26. (Invasive Animals Cooperative Research Centre: Canberra, ACT, Australia)
Burgin, S, Mattila, M, McPhee, D, and Hundloe, T (2015). Feral deer in the suburbs: an emerging issue for Australia? Human Dimensions of Wildlife 20, 65–80.
| Feral deer in the suburbs: an emerging issue for Australia?Crossref | GoogleScholarGoogle Scholar |
Burnham KP, Anderson DR (2002) ‘Model selection and multi-model inference.’ (Springer: New York City, NY, USA)
Burns, H, Gibbons, P, and Claridge, A (2021). Quantifying variations in browsing pressure caused by feral deer for a range of threatened ecological communities and plant growth forms. Austral Ecology 46, 1156–1169.
| Quantifying variations in browsing pressure caused by feral deer for a range of threatened ecological communities and plant growth forms.Crossref | GoogleScholarGoogle Scholar |
Caughley G (1983) ‘The deer wars: the story of deer in New Zealand.’ (Heinemann: Auckland, New Zealand)
Choquenot, D, Hone, J, and Saunders, G (1999). Using aspects of predator–prey theory to evaluate helicopter shooting for feral pig control. Wildlife Research 26, 251–261.
| Using aspects of predator–prey theory to evaluate helicopter shooting for feral pig control.Crossref | GoogleScholarGoogle Scholar |
Claudia Terlouw, EM, Bourguet, C, Deiss, V, and Mallet, C (2015). Origins of movements following stunning and during bleeding in cattle. Meat Science 110, 135–144.
| Origins of movements following stunning and during bleeding in cattle.Crossref | GoogleScholarGoogle Scholar |
Cockram, M, Shaw, D, Milne, E, Bryce, R, McClean, C, and Daniels, M (2011). Comparison of effects of different methods of culling red deer (Cervus elaphus) by shooting on behaviour and post mortem measurements of blood chemistry, muscle glycogen and carcase characteristics. Animal Welfare 20, 211–224.
Commonwealth of Australia (2008) ‘National code of practice for the humane shooting of kangaroos and wallabies for non-commercial purposes.’ (Department of Environment and Heritage: Canberra, ACT, Australia)
Conover MR (2011) Impacts of deer on society. In ‘Biology and Management of White-Tailed Deer’. (Ed. DG Hewitt) pp. 399–408. (CRC Press: Boca Raton, FL, USA)
Côté, SD, Rooney, TP, Tremblay, J-P, Dussault, C, and Waller, DM (2004). Ecological impacts of deer overabundance. Annual Review of Ecology, Evolution, and Systematics 35, 113–147.
| Ecological impacts of deer overabundance.Crossref | GoogleScholarGoogle Scholar |
Cox, DR (1972). Regression models and life‐tables. Journal of the Royal Statistical Society: Series B 34, 187–202.
Cripps, JK, Pacioni, C, Scroggie, MP, Woolnough, AP, and Ramsey, DSL (2019). Introduced deer and their potential role in disease transmission to livestock in Australia. Mammal Review 49, 60–77.
| Introduced deer and their potential role in disease transmission to livestock in Australia.Crossref | GoogleScholarGoogle Scholar |
Curtis, PD (2020). After decades of suburban deer research and management in the eastern United States: where do we go from here? Human-Wildlife Interactions 14, 111–128.
Curtis PD, Stout R, Myers L (1995) Citizen task force strategies for suburban deer management: the Rochester experience. In ‘Urban Deer: a Manageable Resource’. (Ed. JB McAninch) pp. 143–149. (The Wildlife Society: St Louis, MO, USA)
Daoust, PY, and Caraguel, C (2012). The Canadian harp seal hunt: observations on the effectiveness of procedures to avoid poor animal welfare outcomes. Animal Welfare 21, 445–455.
| The Canadian harp seal hunt: observations on the effectiveness of procedures to avoid poor animal welfare outcomes.Crossref | GoogleScholarGoogle Scholar |
Davies, C, Wright, W, Hogan, F, and Visintin, C (2020). Predicting deer–vehicle collision risk across Victoria, Australia. Australian Mammalogy 42, 293–301.
| Predicting deer–vehicle collision risk across Victoria, Australia.Crossref | GoogleScholarGoogle Scholar |
Dawson M (2017) Tools for managing wild deer: ground shooting in the Northern Illawarra Wild Deer Management Program. In ‘2016 National Wild Deer Management Workshop Proceedings’. (Eds DM Forsyth, T Pople, B Page, A Moriarty, D Ramsey, J Parkes, A Wiebkin, C Lane) pp. 22–23. (Invasive Animals Cooperative Research Centre: Canberra, ACT, Australia)
DeNicola, AJ, and Williams, SC (2008). Sharpshooting suburban white-tailed deer reduces deer–vehicle collisions. Human-Wildlife Conflicts 2, 28–33.
DeNicola AJ, VerCauteren KC, Curtis PD, Hyngstrom SE (2000) ‘Managing white-tailed deer in suburban environments.’ (Cornell Cooperative Extension: Ithaca, NY, USA)
DeNicola, AJ, Miller, DS, DeNicola, VL, Meyer, RE, and Gambino, JM (2019). Assessment of humaneness using gunshot targeting the brain and cervical spine for cervid depopulation under field conditions. PLoS One 14, e0213200.
| Assessment of humaneness using gunshot targeting the brain and cervical spine for cervid depopulation under field conditions.Crossref | GoogleScholarGoogle Scholar | 30818392PubMed |
Forsyth DM, Pople T, Page B, Moriarty A, Ramsey D, Parkes J, Wiebkin A, Lane C (Eds) (2017) ‘2016 National Wild Deer Management Workshop Proceedings', 17–18 November 2016, Adelaide, Australia. (Invasive Animals Cooperative Research Centre: Canberra, ACT, Australia)
Gooden, B, French, K, Turner, PJ, and Downey, PO (2009). Impact threshold for an alien plant invader, Lantana camara L., on native plant communities. Biological Conservation 142, 2631–2641.
| Impact threshold for an alien plant invader, Lantana camara L., on native plant communities.Crossref | GoogleScholarGoogle Scholar |
Hampton, JO, and Forsyth, DM (2016). An assessment of animal welfare for the culling of peri-urban kangaroos. Wildlife Research 43, 261–266.
| An assessment of animal welfare for the culling of peri-urban kangaroos.Crossref | GoogleScholarGoogle Scholar |
Hampton, JO, Cowled, BD, Perry, AL, Miller, CJ, Jones, B, and Hart, Q (2014). Quantitative analysis of animal-welfare outcomes in helicopter shooting: a case study with feral dromedary camels (Camelus dromedaries). Wildlife Research 41, 127–135.
| Quantitative analysis of animal-welfare outcomes in helicopter shooting: a case study with feral dromedary camels (Camelus dromedaries).Crossref | GoogleScholarGoogle Scholar |
Hampton, JO, Forsyth, DM, Mackenzie, DI, and Stuart, IG (2015). A simple quantitative method for assessing animal welfare outcomes in terrestrial wildlife shooting: the European rabbit as a case study. Animal Welfare 24, 307–317.
| A simple quantitative method for assessing animal welfare outcomes in terrestrial wildlife shooting: the European rabbit as a case study.Crossref | GoogleScholarGoogle Scholar |
Hampton, JO, Adams, PJ, Forsyth, DM, Cowled, BD, Stuart, IG, Hyndman, TH, and Collins, T (2016a). Improving animal welfare in wildlife shooting: the importance of projectile energy. Wildlife Society Bulletin 40, 678–686.
| Improving animal welfare in wildlife shooting: the importance of projectile energy.Crossref | GoogleScholarGoogle Scholar |
Hampton, JO, Hyndman, TH, Laurence, M, Perry, AL, Adams, P, and Collins, T (2016b). Animal welfare and the use of procedural documents: limitations and refinement. Wildlife Research 43, 599–603.
| Animal welfare and the use of procedural documents: limitations and refinement.Crossref | GoogleScholarGoogle Scholar |
Hampton, JO, Edwards, GP, Cowled, BD, Forsyth, DM, Hyndman, TH, Perry, AL, Miller, CJ, Adams, PJ, and Collins, T (2017). Assessment of animal welfare for helicopter shooting of feral horses. Wildlife Research 44, 97–105.
| Assessment of animal welfare for helicopter shooting of feral horses.Crossref | GoogleScholarGoogle Scholar |
Hampton, JO, MacKenzie, DI, and Forsyth, DM (2019a). How many to sample? Statistical guidelines for monitoring animal welfare outcomes. PLoS One 14, e0211417.
| How many to sample? Statistical guidelines for monitoring animal welfare outcomes.Crossref | GoogleScholarGoogle Scholar | 30699193PubMed |
Hampton, JO, Warburton, B, and Sandøe, P (2019b). Compassionate versus consequentialist conservation. Conservation Biology 33, 751–759.
| Compassionate versus consequentialist conservation.Crossref | GoogleScholarGoogle Scholar | 30411399PubMed |
Hampton, JO, DeNicola, AJ, and Forsyth, DM (2020). Assessment of Lead‐Free .22 LR Bullets for Shooting European Rabbits. Wildlife Society Bulletin 44, 760–765.
| Assessment of Lead‐Free .22 LR Bullets for Shooting European Rabbits.Crossref | GoogleScholarGoogle Scholar |
Hampton, JO, Arnemo, JM, Barnsley, R, Cattet, M, Daoust, P-Y, DeNicola, AJ, Eccles, G, Fletcher, D, Hinds, LA, Hunt, R, Portas, T, Stokke, S, Warburton, B, and Wimpenny, C (2021a). Animal welfare testing for shooting and darting free-ranging wildlife: a review and recommendations. Wildlife Research 48, 577–589.
| Animal welfare testing for shooting and darting free-ranging wildlife: a review and recommendations.Crossref | GoogleScholarGoogle Scholar |
Hampton, JO, Bengsen, AJ, Pople, A, Brennan, M, Leeson, M, and Forsyth, DM (2021b). Animal welfare outcomes of helicopter-based shooting of deer in Australia. Wildlife Research , .
| Animal welfare outcomes of helicopter-based shooting of deer in Australia.Crossref | GoogleScholarGoogle Scholar |
Hanson C, Hall T, DeNicola A, Silander S, Keitt B, Campbell K, Veitch C, Clout M, Martin A, Russell J (2019) Rhesus macaque eradication to restore the ecological integrity of Desecheo National Wildlife Refuge, Puerto Rico. In ‘Island Invasives: Scaling Up to Meet the Challenge. Proceedings of the International Conference on Island Invasives 2017’. (Eds CR Veitch, MN Clout, AR Martin, JC Russell, CJ West) pp. 249–255. (IUCN: Gland, Switzerland)
Harris LH (2002) ‘The Deer Menace: a History of Government Pest Control Operations, 1930–1987.’ (L. H. Harris: Wellington, New Zealand)
Hone, J (1990). Predator prey theory and feral pig control, with emphasis on evaluation of shooting from a helicopter. Wildlife Research 17, 123–130.
| Predator prey theory and feral pig control, with emphasis on evaluation of shooting from a helicopter.Crossref | GoogleScholarGoogle Scholar |
Hone J (1994) ‘Analysis of Vertebrate Pest Control.’ (Cambridge University Press: Cambridge, UK)
Invasive Species Council (2018) ‘Feral deer putting NSW drivers at risk.’ (Invasive Species Council: Melbourne, Vic., Australia)
Kanstrup, N, Balsby, TJS, and Thomas, VG (2016). Efficacy of non-lead rifle ammunition for hunting in Denmark. European Journal of Wildlife Research 62, 333–340.
| Efficacy of non-lead rifle ammunition for hunting in Denmark.Crossref | GoogleScholarGoogle Scholar |
Keith, D, and Pellow, B (2005). Effects of Javan rusa deer (Cervus timorensis) on native plant species in the Jibbon–Bundeena area, Royal National Park, New South Wales. Linnean Society of New South Wales 126, 99–110.
Kilpatrick, HJ, and Stober, WA (2002). Effects of temporary bait sites on movements of suburban white-tailed deer. Wildlife Society Bulletin 30, 760–766.
Kilpatrick, HJ, Spohr, SM, and DeNicola, AJ (1997). Darting urban deer: techniques and technology. Wildlife Society Bulletin 25, 542–546.
King CM, Forsyth DM (2021) ‘The Handbook of New Zealand Mammals.’ (CSIRO Publishing: Melbourne, Vic., Australia; and Otago University Press: Dunedin, New Zealand)
Knudsen, SK (2005). A review of the criteria used to assess insensibility and death in hunted whales compared to other species. The Veterinary Journal 169, 42–59.
| A review of the criteria used to assess insensibility and death in hunted whales compared to other species.Crossref | GoogleScholarGoogle Scholar | 15683763PubMed |
Langenau Jr, EE (1986). Factors associated with hunter retrieval of deer hit by arrows and shotgun slugs. Leisure Sciences 8, 417–438.
| Factors associated with hunter retrieval of deer hit by arrows and shotgun slugs.Crossref | GoogleScholarGoogle Scholar |
Latham, ADM, Latham, MC, Norbury, GL, Forsyth, DM, and Warburton, B (2020). A review of the damage caused by invasive wild mammalian herbivores to primary production in New Zealand. New Zealand Journal of Zoology 47, 20–52.
| A review of the damage caused by invasive wild mammalian herbivores to primary production in New Zealand.Crossref | GoogleScholarGoogle Scholar |
Lewis, A, Pinchin, A, and Kestin, S (1997). Welfare implications of the night shooting of wild impala (Aepyceros melampus). Animal Welfare 6, 123–131.
Lindeman MJ, Forsyth DM (2008) Agricultural impacts of wild deer in Victoria. Arthur Rylah Institute for Environmental Research Technical Report Series No. 182. Department of Sustainability and Environment, Melbourne, Vic., Australia.
Littin, KE, O'Connor, CE, Gregory, NG, Mellor, DJ, and Eason, CT (2002). Behaviour, coagulopathy and pathology of brushtail possums (Trichosurus vulpecula) poisoned with brodifacoum. Wildlife Research 29, 259–267.
| Behaviour, coagulopathy and pathology of brushtail possums (Trichosurus vulpecula) poisoned with brodifacoum.Crossref | GoogleScholarGoogle Scholar |
Logan, TW, Ashton-Butt, A, and Ward, AI (2019). Improving daytime detection of deer for surveillance and management. European Journal of Wildlife Research 65, 83.
| Improving daytime detection of deer for surveillance and management.Crossref | GoogleScholarGoogle Scholar |
McCann, BE, Whitworth, W, and Newman, RA (2016). Efficacy of non-lead ammunition for culling elk at Theodore Roosevelt National Park. Human–Wildlife Interactions 10, 268–282.
McTee, M, Young, M, Umansky, A, and Ramsey, P (2017). Better bullets to shoot small mammals without poisoning scavengers. Wildlife Society Bulletin 41, 736–742.
| Better bullets to shoot small mammals without poisoning scavengers.Crossref | GoogleScholarGoogle Scholar |
Moriarty, A (2004a). The liberation, distribution, abundance and management of wild deer in Australia. Wildlife Research 31, 291–299.
| The liberation, distribution, abundance and management of wild deer in Australia.Crossref | GoogleScholarGoogle Scholar |
Moriarty AJ (2004b) Wild deer herds in Australia’s urban fringe: issues, management and politics. In ‘Urban Wildlife: More Than Meets the Eye’. (Eds D Lunney, S Burgin) pp. 179–185. (Royal Zoological Society of New South Wales: Sydney, NSW, Australia)
Moriarty AJ (2004c) Ecology and environmental impact of Javan rusa deer (Cervus timorensis russa) in the Royal National Park. PhD Thesis, University of Western Sydney, Sydney, NSW, Australia.
Moriarty AJ (2009) Science based management of wild deer in Australia: a case study-rusa deer in the Royal National Park. In ‘Workshop Proceedings: What are the Issues for the Management of Wild Deer in Australia?’. (Ed. SR McLeod) pp. 112–118. (Invasive Animals Cooperative Research Centre: Canberra, ACT, Australia)
New Zealand Government (2018) ‘Deer code of welfare.’ (New Zealand Government: Wellington, New Zealand)
NSW National Parks and Wildlife Service (2020) ‘Feral Animal Aerial Shooting Team (FAAST) training.’ (NSW National Parks and Wildlife Service: Sydney, NSW, Australia)
Øen, EO (2021). Animal welfare in the conduct of whaling: a review of the research and developments to improve animal welfare in the minke whale hunt in Norway 1981–2005. Senri Ethnological Studies 104, 287–318.
Page B, Pelton G, Austin J, Peters K, Forsyth DM, Wiebkin A, Freeman E, Howlett B, Williams L, Williams G, Waycott M, Woolnough A (2017) Agricultural and economic impacts of wild deer. In ‘2016 National Wild Deer Management Workshop Proceedings’. (Eds DM Forsyth, T Pople, B Page, A Moriarty, D Ramsey, J Parkes, A Wiebkin, C Lane) pp. 10–13. (Invasive Animals Cooperative Research Centre: Canberra, ACT, Australia)
Parker, H, Rosell, F, and Danielsen, J (2006). Efficacy of cartridge type and projectile design in the harvest of beaver. Wildlife Society Bulletin 34, 127–130.
| Efficacy of cartridge type and projectile design in the harvest of beaver.Crossref | GoogleScholarGoogle Scholar |
Pecorella, I, Ferretti, F, Sforzi, A, and Macchi, E (2016). Effects of culling on vigilance behaviour and endogenous stress response of female fallow deer. Wildlife Research 43, 189–196.
| Effects of culling on vigilance behaviour and endogenous stress response of female fallow deer.Crossref | GoogleScholarGoogle Scholar |
Reimoser S (2012) Influence of anthropogenic disturbances on activity, behavior and heart rate of roe deer (Capreolus capreolus) and red deer (Cervus elaphus), in context of their daily and yearly patterns. In ‘Deer: Habitat, Behavior and Conservation’. (Eds AA Cahler, JP Marsten) pp. 1–96. (Nova Science Publishers: Hauppauge, NY, USA)
Rosenberry, CS, Long, ES, Hassel-Finnegan, HM, Buonaccorsi, VP, Diefenbach, DR, and Wallingford, BD (2009). Lack of mother-offspring relationships in white‐tailed deer capture groups. The Journal of Wildlife Management 73, 357–361.
| Lack of mother-offspring relationships in white‐tailed deer capture groups.Crossref | GoogleScholarGoogle Scholar |
Rutberg AT (1997) The science of deer management: an animal welfare perspective. In ‘The Science of Overabundance: Deer Ecology and Population Management’. (Eds WJ McShea, HB Underwood, JH Rappole) pp. 37–54. (Smithsonian Institution Press: Washington, DC, USA)
Ryeng, K, and Larsen, S (2021). The relative effectiveness of two expanding bullet designs in young harp seals (Pagophilus groenlandicus): a randomised controlled field study in the Norwegian harp seal hunt. Animal Welfare 30, 155–167.
| The relative effectiveness of two expanding bullet designs in young harp seals (Pagophilus groenlandicus): a randomised controlled field study in the Norwegian harp seal hunt.Crossref | GoogleScholarGoogle Scholar |
Schulz, JH, Bonnot, TW, Millspaugh, JJ, and Mong, TW (2013). Harvest and crippling rates of mourning doves in Missouri. Wildlife Society Bulletin 37, 287–292.
| Harvest and crippling rates of mourning doves in Missouri.Crossref | GoogleScholarGoogle Scholar |
Sharp T (2012) ‘Standard Operating Procedure GOA002: Aerial Shooting of Feral Goats.’ (Centre for Invasive Species Solutions: Canberra, ACT, Australia)
Sharp T (2013) ‘Standard Operating Procedure DEE001: Ground Shooting of Feral Deer.’ (Centre for Invasive Species Solutions: Canberra, ACT, Australia)
Shearer JK, Ramirez A (2012) ‘Procedures for Humane Euthanasia. Humane Euthanasia of Sick, Injured and/or Debilitated Livestock.’ (Iowa State University: Ames, IA, USA)
Smith, SDG, and Ryeng, KA (2022). Developments in the quantitative assessment of welfare outcomes in hunted mammals. NAMMCO Scientific Publications , .
Standing Committee on Agriculture, Animal Health Committee (2002) ‘Model Code of Practice for the Welfare of Animals: Feral Livestock Animals: Destruction or Capture, Handling and Marketing: SCARM Report 34.’ (CSIRO Publishing: Melbourne, Vic., Australia)
Stokke, S, Arnemo, JM, Brainerd, S, Söderberg, A, Kraabøl, M, and Ytrehus, B (2018). Defining animal welfare standards in hunting: body mass determines thresholds for incapacitation time and flight distance. Scientific Reports 8, 13786.
| Defining animal welfare standards in hunting: body mass determines thresholds for incapacitation time and flight distance.Crossref | GoogleScholarGoogle Scholar | 30214004PubMed |
Tuckwell CD (2003) ‘The Deer Farming Handbook.’ (Rural Industries Research & Development Corporation: Canberra, ACT, Australia)
Urquhart, KA, and McKendrick, IJ (2003). Survey of permanent wound tracts in the carcases of culled wild red deer in Scotland. Veterinary Record 152, 497–501.
| Survey of permanent wound tracts in the carcases of culled wild red deer in Scotland.Crossref | GoogleScholarGoogle Scholar |
Urquhart, KA, and McKendrick, IJ (2006). Prevalence of ‘head shooting’ and the characteristics of the wounds in culled wild Scottish red deer. Veterinary Record 159, 75–79.
| Prevalence of ‘head shooting’ and the characteristics of the wounds in culled wild Scottish red deer.Crossref | GoogleScholarGoogle Scholar |
Valente, AM, Acevedo, P, Figueiredo, AM, Fonseca, C, and Torres, RT (2020). Overabundant wild ungulate populations in Europe: management with consideration of socio‐ecological consequences. Mammal Review 50, 353–366.
| Overabundant wild ungulate populations in Europe: management with consideration of socio‐ecological consequences.Crossref | GoogleScholarGoogle Scholar |
Van Deelen, T, and Etter, D (2003). Effort and the functional response of deer hunters. Human Dimensions of Wildlife 8, 97–108.
| Effort and the functional response of deer hunters.Crossref | GoogleScholarGoogle Scholar |
Wardle, DA, Barker, GM, Yeates, GW, Bonner, KI, and Ghani, A (2001). Introduced browsing mammals in New Zealand natural forests: aboveground and belowground consequences. Ecological Monographs 71, 587–614.
| Introduced browsing mammals in New Zealand natural forests: aboveground and belowground consequences.Crossref | GoogleScholarGoogle Scholar |
Watter, K, Thomas, E, White, N, Finch, N, and Murray, PJ (2020). Reproductive seasonality and rate of increase of wild sambar deer (Rusa unicolor) in a new environment, Victoria, Australia. Animal Reproduction Science 223, 106630.
| Reproductive seasonality and rate of increase of wild sambar deer (Rusa unicolor) in a new environment, Victoria, Australia.Crossref | GoogleScholarGoogle Scholar | 33166829PubMed |
Weckerly, FW (1998). Sexual-size dimorphism: influence of mass and mating systems in the most dimorphic mammals. Journal of Mammalogy 79, 33–52.
| Sexual-size dimorphism: influence of mass and mating systems in the most dimorphic mammals.Crossref | GoogleScholarGoogle Scholar |
Williams, W, McSorley, A, Hunt, R, and Eccles, G (2018). Minimising noise disturbance during ground shooting of pest animals through the use of a muzzle blast suppressor/silencer. Ecological Management & Restoration 19, 172–175.
| Minimising noise disturbance during ground shooting of pest animals through the use of a muzzle blast suppressor/silencer.Crossref | GoogleScholarGoogle Scholar |