

Feral cat GPS tracking and simulation models to improve the conservation management of night parrots
Stephen A. Murphy A B * , Hugh McGregor A C , Nicholas P. Leseberg A B , James Watson A B and Alex S. Kutt C D
A C , Nicholas P. Leseberg A B , James Watson A B and Alex S. Kutt C D
A Threatened Species Recovery Hub, National Environmental Science Program, University of Queensland, St Lucia, Qld 4067, Australia.
B School of Earth and Environmental Science, University of Queensland, St Lucia, Qld 4067, Australia.
C School of Natural Sciences, University of Tasmania, Hobart, Tas. 7005, Australia.
D School of Ecosystem and Forest Science, The University of Melbourne, Parkville, Vic. 3010, Australia.
Wildlife Research 50(5) 325-334 https://doi.org/10.1071/WR21123
Submitted: 23 August 2021 Accepted: 21 June 2022 Published: 13 July 2022
© 2023 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Context: Night parrots (Pezoporus occidentalis) are one of Australia’s most endangered birds, and there is evidence suggesting feral cats (Felis catus) are a major cause of decline. However, because night parrots currently have a restricted distribution, little is known of the ecology of feral cats around their remaining populations. This limits the development of effective management strategies.
Aims: The aims of this study were to understand feral cat movement and habitat selection around night parrots, and to then estimate the effectiveness of possible management actions.
Methods: Research was conducted around the only confirmed night parrot population in eastern Australia. In 2019 and 2020, we obtained GPS data from nine feral cats, and used step selection functions to assess preferred habitats. Management options were then simulated based on cat movement data, including altering trap numbers and layout, and changing routes for night spotlight shooting (using existing roads, random walking or creating new roads in preferred habitats).
Key results: Feral cats preferred alluvial and riparian habitats and avoided rocky woodlands and roads. Simulated control efforts were more successful if traps are placed at ‘pinch points’ where drainage lines converged, and if new roads were created near to creek lines and alluvial habitats.
Conclusions: Feral cats move around the last known population of night parrots in eastern Australia, travelling through and using many shared habitats. Targeting creek lines and alluvial areas in cat control operations would improve effectiveness and potentially reduce predation impacts on night parrots.
Implications: Conservation of endangered birds like night parrots can be enhanced through understanding the ecology of threats such as feral cats to develop locally tailored control operations.
Keywords: adaptive management, applied ecology, endangered species, feral cats, invasive species, night parrots, simulation models, wildlife tracking.
Introduction
Feral predators such as the cat (Felis catus) are a significant threat to biodiversity, and can annually kill a remarkable number of birds (Woinarski et al. 2017a). Ground-nesting species are particularly at risk, and night parrots (Pezoporus occidentalis) have life history traits that may make them especially vulnerable (Woinarski et al. 2017b). First, they nest and forage on the ground (Jackett et al. 2017; Murphy et al. 2017a, 2017b), and at certain times need to drink at places frequented by cats (Kearney et al. 2016; this study). Night parrots also lack some of the adaptations typically associated with visual acuity in nocturnal birds (Iwaniuk et al. 2020). Second, there is direct evidence of predation such as historical reports of domestic cats bringing home numerous night parrot body parts in the 1800s (Ashby 1924), and more recent observations of cats frequenting night parrot nesting areas when nestlings become vocal just prior to fledging (N. Leseberg, unpubl. data). This suggests that management of feral cats where night parrots occur is a critical conservation action.
Feral cats are notoriously difficult to control, especially when at low densities. Lethal control is a typical approach, but there are potential unintended consequences that may counteract the management intent, such as increased immigration of feral cats (Doherty and Ritchie 2017). Approaches such as baiting can be ineffective in many circumstances (Fancourt et al. 2021), and may be undesirable if impacts on dingoes (Canis lupus dingo, which are thought to sometimes regulate cats, Kennedy et al. 2012) cannot be mitigated. Shooting and trapping are expensive exercises (Ruykys and Carter 2019), largely because they involve intensive input from experienced personnel and can normally be only undertaken in short sessions in remote settings. Recently developed poison delivery systems that exploit cat grooming behaviour show some promise (Moseby et al. 2020); however, they rely on installation along predictable pathways where target predators move, such as watercourses, which can frequently be inundated. For the foreseeable future, cat control must rely largely on refining existing control techniques, both direct and indirect, via habitat management (Ruykys and Carter 2019). This means developing integrated strategies that use multiple control methods, sensitive land management that improves the quality of the resources available for the potentially affected species, and most significantly, gaining a better understanding of cat movements and behaviour to target control programs more cost-effectively (Comer et al. 2020).
Recent surveys of night parrots in Queensland and Western Australia suggest remnant populations are small. In Queensland’s case, where most research and monitoring has taken place to date, the night parrot population size is likely to be around 20 individuals (N. Leseberg and S. Murphy, unpubl. data). Being small, these populations are especially vulnerable to cat predation, even if cats themselves occur at a low density, which appears to be the case in Queensland (Murphy et al. 2018; Kearney et al. 2022). Indeed, Moseby et al. (2015) and Greenwell et al. (2019) showed that even individual cats can impart a devastating impact on small populations of native animals. As such, cat control is one of the primary conservation land management actions for threatened species, despite the potential costs.
Feral cat control needs to navigate a pathway through effort, cost-effectiveness, innovation, and perverse outcomes (Joseph et al. 2009), especially when the risk of extinction for the species to be protected is very real (Murphy et al. 2018). Therefore, planning careful strategies, rather than simply instigating control for its own sake, is exceptionally important. In this study, we had two objectives. First, we aimed to understand the movement patterns and habitat selection of feral cats in a landscape in which night parrots occur. Specifically, we investigated via GPS tracking, cat movement patterns, transit routes and habitat preferences. In addition, we used a subset of GPS tracking data to simulate the effectiveness of different control scenarios and therefore to predict potential effectiveness of various cat control techniques (i.e. shooting, trapping, and baiting, or some combination of each).
Materials and methods
Study site
This study was undertaken on Pullen Pullen Special Wildlife Reserve and neighbouring Mount Windsor and Brighton Downs cattle stations in south-western Queensland (23.3°S 141.6°E) (Fig. 1). The area is hot and arid, with approximate mean maximum temperatures of 39°C in January and 23°C in July. Annual rainfall is highly unpredictable, with dry periods punctuated by occasional wet years. Median annual rainfall at Brighton Downs is 240 mm, and 65% of the rainfall occurs between December and March (Bureau of Meteorology 2017). The vegetation is typical of the Channel Country: largely treeless, and comprising tussock and hummock grasslands, gibber rises, floodplains with claypans and sparse annual plant cover, very open woodland escarpments, and ephemeral drainage lines fringed by Acacia species, and Coolabah (Eucalyptus coolabah). Habitats are described in detail in Murphy et al. (2017b).

|
Trapping and handling
Cats were captured using a combination of soft-jawed foothold traps (size 1.5 Oneida Victor Soft Catch, Ohio, USA, and size 1.5 Bridger, Minnesota, USA) or cages (30 cm × 30 cm × 60 cm) baited with cat urine and/or faeces. Visual lures, consisting of feathers and/or foil, were installed near traps to attract attention. The traps were checked every morning. Caught cats were removed from traps and restrained inside canvas holding bags by tying together their forelegs and hind legs (separately) with plastic cable ties. This avoided the need for sedatives and ensured cats were released in an alert state. GPS trackers were fitted while the cats were restrained and sex, weight, coat colour and basic morphometrics recorded before release at capture site. Handling typically took less than 10 min.
Tracking
Two types of GPS devices were used, and both allowed data retrieval via satellite. First, we used SPOT Trace™ units (Garmin, Olathe, Kansas, USA), which were glued using two-part epoxy to a leather or synthetic harness. The harnesses had neck and chest straps, which were glued and bolted together for fitting and were designed so that the unit sat between the shoulder blades facing skywards. The units weighed approximately 100 g, including the AAA batteries and harness material. The main advantage of the SPOT™ units was their low cost, at roughly 10% of the second type of unit we used, which were Iridium Litetrack-130s (Lotek, Havelock North, New Zealand). These were fitted to the cat using a single neck collar and weighed approximately 130 g. Iridium units recorded data at 1-h intervals and SPOT trace units recorded data at 5-min intervals. However, for this study we used 1-h interval data from SPOT trace units to ensure comparability with the Iridium data, and that all movement data were consistent.
Habitat selection
All simulations, spatial manipulations and modelling were undertaken in R ver. 4.1.2 (R Core Team 2020). To measure feral cat habitat selection, we applied step selection functions (Thurfjell et al. 2014) with the R library amt (Signer et al. 2019). This was used instead of home-range wide resource selection functions because at least two of the cats captured did not appear to be restricted to a core home range. We limited our analysis to steps of between 1 and 3 h apart. For each step, five random steps were generated based on the probability distributions of turn angles and distance between fixes for each individual cat (gamma and von mises distributions respectively). Models included habitat variables based on Queensland’s state-wide regional ecosystem mapping (Queensland Herbarium 2021). To reduce the dimensionality of our analyses, we used local knowledge to aggregate these descriptions from 26 down to nine habitat types. For example, ‘Sparse herbland, open water or bare areas on flood plain lakes and interdune clay pans and lakes’ and ‘Variable sparse to open herbland on frequently flooded alluvial plains’ were aggregated to ‘Sparse grasslands’. The full list of the habitat types used in our analyses is presented in Table 1, and the aggregations from the regional ecosystem mapping is shown in Appendix 1 (see Supplementary material). We included vegetation with an interaction term with the GPS location time of day (i.e. night, being 1800−0600 hours, or day), and whether locations were within 25 m of a road. Then, multiple conditional logistic regression models with different habitat variables were compared within an Information Theory framework (Burnham and Anderson 1998) using Akaike Information Criterion values corrected for small sample size (AICc). A null model was also included. In each conditional logistic model, steps were considered strata and each individual cat considered as a cluster. In the models that considered vegetation as categorical, the gibber habitats (those with scant vegetation) were considered as the intercept, against which the odds of selection of other variables were measured.

|
Trapping simulations
We examined the effect of trap number, deployment duration and site (random vs non-random) on the probability of trap set encounter given the GPS movements of cats. We treated cage traps, foot-hold traps and Felixer (Thylation, Adelaide, Australia) grooming traps together. Cats in largely treeless arid areas are known to move mostly along drainage lines (Moseby et al. 2009), and this is the case in the study area, as identified by camera trapping data (S. Murphy, H. McGregor and A. Kutt, unpubl. data). As such, our simulations only placed sets along drainage lines, placed both randomly and manually. For random set placement, a large sample of drainage lines (~550 km) was first manually mapped within the convex hulls (the smallest convex polygons that contain all of the points) for each cat based on high resolution satellite imagery (30-cm and 60-cm resolution; ESRI 2019). From 1 to 50 traps were then placed randomly along these mapped lines (for each cat) using the st.sample function in R (Pebesma et al. 2018). Random trap set simulations were run 100 times. For manual set placement, one author with experience setting cat traps (SM) chose five sites within the convex hull of each cat. This was done ‘blind’, i.e. without referring to the GPS tracking data. The sites were placed at the point where multiple parallel braided drainage channels coalesced to drainage ‘pinch points’ because these were suspected to be places experiencing a high level of cat activity.
To calculate cat movements and how these interacted with the simulated trap placements, we created a habitat utilisation surface for each cat on each day with 30-m resolution, using the function brownian.bridge.dyn (Kranstauber et al. 2012). Utilisation probabilities for each day were scaled from 0 to 1, and we used these values as a proxy for trap encounter probability. Daily encounter probabilities were extracted for each trap using the function raster::extract (Hijmans 2019), with the maximum probability value returned with a 30-m buffer around each trap. Probability values greater than 0.01 were accepted as instances where cats were likely to have encountered traps. Despite being quite low, visual inspection of rasters showed that this value captured the most likely trajectories between GPS fixes. The primary variable of interest was the time it took a cat to first encounter a trap under each scenario, which incorporated 1– 50 randomly placed traps, and five manually placed traps.
Spotlighting simulations
To examine the effectiveness of spotlighting and shooting, and how this could be improved as a management tool, we examined the number of cat detections over a series of simulated spotlighting ‘patrols’, based on the cat GPS tracking data from four cats, which overlapped in time and space. We used the location, time, and frequency of patrols, as the simulation variables, because these are the factors that land managers have most control over and are the costliest.
For location, we compared detection rates for six different classes of patrol track: (1) fixed patrol along an existing 52.6 km station road, at approximately 13 km/h (‘car along existing’); (2) fixed patrol along a 53.0 km hypothetical road (also at 13 km/h), aligned parallel to watercourses (‘car along new’, see below for justification); (3) cross country driving patrol starting from a random point along the existing track (‘car from existing’); (4) cross country driving patrol starting from a random point along the new track (‘car from new’); (5) cross country walking patrol, at 2–3 km/h, starting from a random point along the existing track (‘walk from existing’); and (6) cross country walking, also at 2–3 km/h, patrol starting from a random point along the new track (‘walk from new’). A hypothetical new track parallel to watercourses was needed because existing vehicle tracks in this landscape tend to be non-randomly placed perpendicular to watercourses and at higher elevations.
The random tracks were generated using the function rTrack (Pebesma et al. 2021), and were each constrained by the number of points (2800) and time step arguments (5 s), which generated tracks for the required 4-h patrol time. The sd0 argument in the function was set to 0.99 and 0.3 to produce realistic driving and walking tracks (respectively) in terms of shape and length, although the length varied due to the random way the tracks were created (~23–28 km for random drive and ~6–9 km for walking).
With respect to patrol time, we ran the simulations over 2 × 4-h blocks starting at 0800 and 0000 hours. This ensured simulations included most hours typically covered by a spotlighting patrol. Rather than select dates randomly for patrols, we ran simulations for every night for which overlapping spatio-temporal data existed for four cats (n = 114 nights). Simulations were run for existing and new fixed patrols twice per night (starting at 0800 and 0000 hours) on every night. Every cross country patrol started at a new random location along existing and new roads, and then proceeded over random trajectories for each of the 2 × 4-h blocks (also starting at 0800 and 0000 hours).
The habitat utilisation rasters generated for the trapping simulations do not have high precision time values, which are critical for the temporally dynamic shooting simulations. As such, for the shooting simulations, for each date we converted the relevant cat and patrol spatial data to a series of time-stamped trajectories (i.e. lines between temporally adjacent points). Pairwise distances between temporally coincident trajectories (within 60-s windows) were calculated using the function avedistTrack (Pebesma et al. 2021). Pairwise distances less than 300 m were kept; 300 m was chosen based on the estimated typical maximum range that cat eye shine can be detected by spotlight in our open study area. Only one distance (the shortest) was kept for sequential runs of detections (≤1 min) for each cat.
We modelled the effect of patrol class and hour of the night (both treated as categorical predictors) on whether or not cats were detected in each hour of the patrols (binomial response) using multiple logistic regression with binomial error distribution, using the R function stats::glm. We compared models that included each predictor separately and their interactions. The effect of patrol frequency was estimated based on logistic regression coefficients of the model with the lowest AICc value. The response variable was converted to cat detections per hour for plotting.
Results
Trapping and data acquisition
Eleven cats were caught (10 in footholds and one in a cage) between 5 August 2019 and 8 July 2020 over 1335 trap nights. Two SPOT units failed to produce any useful data and lasted less than 1 or 2 days, which could have been caused by unit failure or a disruption to power.
Habitat selection
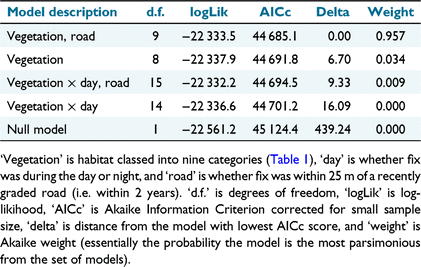
We obtained usable data from nine individual cats (three females and six males), with 8313 fixes 1 h or more apart. After considering only steps between 1 and 3 h apart, 6893 steps were included in the analysis. For the step selection models, the most parsimonious (i.e. delta AICc <4) model of cat habitat selection included both vegetation classes and proximity to roads (Table 2). Within the top model (Table 3, Fig. 2), spinifex and sparsely vegetated habitats were not significantly selected over gibber habitats, escarpments and flats with scattered vegetation were selected more significantly, and alluvial and riparian habitats were substantially and significantly selected. Riparian habitats were selected almost three-fold over gibber habitats. Areas within 25 m of roads were avoided.
|

|

|
Trapping simulations
Simulations involving 1–50 randomly placed traps for six cats over 623 tracking days, repeated 100 times, generated >1.9 million values for trap encounter probabilities greater than the 0.01 probability threshold used to define trap encounters. Qualitatively, the time it took cats to encounter traps reduced rapidly with an increasing number of traps up until about 10 traps, after which it tailed off slowly (Fig. 3). Manual placement of traps at drainage pinch points approximately halved the time it took cats to encounter traps (7.5 ± 6.3 days cf. 14.8 ± 12.6 days respectively for five traps at pinch points and five traps randomly placed along drainages).

|
Spotlighting simulations
Spotlighting was simulated for 114 nights over the six track classes (5472 spotlighting hours), and these generated 255 detections (as defined by cats being <300 m within a 60 s window of the observer, Fig. 4). The model with the lowest AICc value was one that included non-interacting terms for hour and track class. Detections were significantly lower in the hour between 0100 and 0200 hours (coefficient estimate = −0.86, Z = −2.78, P = 0.005). Walking from the existing track resulted insignificantly fewerdetections (coefficient estimate = −0.85, Z = −2.46, P = 0.014). Significantly more cats were detected by driving along the new track (coefficient estimate = 1.12, Z = 5.96, P < 0.0001). All other track classes had no effect on cat detection.

|
Discussion
Two potential limitations of our approach should be initially highlighted. First, that our sample size of nine tracked cats could be too small to provide reliable inferences for the whole cat population in and around Pullen Pullen where night parrots occur. Second, it may be inferred from our approach that future management needs to track cats prior to any control session, rather than just reviewing the local landscape and habitat patterns. In the first instance, although small, we argue that our sample was representative of the population; we tracked three females (2.6–3 kg) and six males (3.2–4.5 kg), and these were all trapped using scent as opposed to food, which can result in a biased sample (Gorney et al. 1999). Additionally, the GPS data obtained from the nine cats werevery detailed, covering multiple seasons, and they revealed patterns of habitat use consistent with camera trap data from the same study area (authors S. Murphy, H. McGregor and A. Kutt, unpubl. data). Similarly, our results are also consistent with other studies from the arid zone (Moseby et al. 2009; Doherty et al. 2015). As such, we believe that inferences about cat movements for our study area are sound. On the second point, we feel that future management within the region involving similar habitats could confidently draw useful inferences from our results. We also recognise that cat control in different environments (e.g. savanna woodland) would benefit from local cat tracking information to refine management, and any long term management strategy should review their approach regularly, as new data are obtained.
We found feral cats to be wide ranging and to use most habitats in the area around the Queensland population of night parrots; they were also most likely to select for riparian and other productive (e.g. floodplain) habitats. Most concerning, but not surprisingly, feral cats were also recorded frequently moving through habitats associated with night parrots i.e. dense spinifex and gibber plains. Only rocky woodlands and roads were avoided by cats, the latter being a significant result because tracks and roads are often the locations where cat monitoring and control is focussed (Wysong et al. 2020). This suggests that in this arid environment, trapping and monitoring for feral cats is best targeted in alluvial areas.
Previous studies have suggested that in the arid zone, feral cats often use locations where there is a high density of vegetation cover (Moseby et al. 2009). Feral cats are also known to shelter in trees in this region, especially in summer and when there is heightened abundance caused by resource pulses (Pettigrew 1993). In addition, native predators such as dingoes frequently use roads and tracks (Vernes et al. 2001), and there might be a degree of habitat separation through meso-predator suppression that affects feral cat behaviour and distribution. Understanding how predators use the landscape is a critical means of more targeted and effective management and control (Bengsen et al. 2012).
Using the information on movement and habitat use obtained from the GPS collared feral cats, we simulated how trapping could be improved. The relative cost and effectiveness of feral cat management is an important consideration for land managers (Nutter et al. 2004; Lohr and Algar 2020). The results from trapping simulations demonstrated that trap encounter probability was very variable, even for large numbers of traps. For the random placements (although still constrained to drainage lines), some simulation iterations did not produce their first encounter for long periods, even when large numbers of traps were involved. For example, one simulation using 50 traps took 17 days to yield the first trap encounter for any of the six cats. In the case of the night parrot reserve, where this study took place, the property does not have permanent staff on-site and, as such, most visits by management are rarely longer than 10–12 days; this illustrates the mismatch and potential ineffectiveness of trapping and the importance for evidence-based control approaches (Doherty and Ritchie 2017). By comparison, manual trap placement on drainage lines at pinch points, where the naturally braided channels converged into more narrow pathways, generated encounters more quickly (for example, mean time to first encounter for five random traps was 14.8 ± 12.6 days compared with 7.5 ± 6.3 days for five manually placed traps). We recommend that managers use high-resolution satellite imagery to help guide trap placement at such locations prior to field operations.
Similarly, with the shooting simulations, the use of existing property tracks were not the optimal pathways for targeting feral cat control. Track based surveys for feral predators are generally considered the most efficient and standard manner to find and estimate abundance in remote arid landscapes owing to the ease of transit and the large distances that can be covered (Edwards et al. 2000). However, in our study, using knowledge of the spatial and temporal movement of feral cats at the night parrot reserve, the use of existing tracks with vehicle patrols resulted in a low number of detections. In contrast, a hypothetical new vehicle track positioned to run parallel to main drainage lines (i.e. using the combined knowledge of cat habitat preferences and shooting simulations) in the study area resulted in a threefold increase in cat detection rate. This suggests one option for future feral cat management is either to undertake patrols off track and in parallel to these drainage lines, or to grade new tracks in locations that maximise feral cat encounters.
From a management perspective, the relative benefits, or potential costs, of the creation of new tracks, how these tracks are then maintained, and potential unforeseen outcomes like increased weed incursion or increased dingo use of this infrastructure potentially affecting cat behaviour, need to be considered (Oppel et al. 2014). In addition, if cats are avoiding roads due to vehicles and shooting frequency, the benefit of a new track could decline. As these roads are theoretically cat management tracks only, the frequency of use over time could be moderated to quell any effect of sensitising cats to their use. Another alternative is to undertake more random car or walking patrols on cross-country tracks (meaning without a formed or graded track) targeting riparian locations; but both car and walking patrols yielded considerably fewer mean number of detections than new tracks along known cat movement routes.
This study has demonstrated that feral cats are found throughout the region occupied by night parrots and use similar habitats, despite a preference for riparian zones. This separation in preferred habitat (i.e. cats in treed drainage lines) provides some comfort with respect to a reduced probability of regular interactions and predation. Feral cats in this region are known to undergo sporadic weather and prey mediated population booms and congregate in the shelter of riparian vegetation (Pettigrew 1993; Palmer 1999). Feral cats also rest at night in riparian trees (S. Murphy, H. McGregor and A. Kutt, unpubl. data). These findings are also useful for inferring the cat predation risk to other threatened species that occur on the reserve and the likelihood of encounter, given the knowledge of feral cat habitat use and the distribution of other species (Kearney et al. 2021; Kutt et al. 2021).
The adaptation of management in response to new knowledge is a critical component of feral predator control (Lohr and Algar 2020) and adaptive management is the foundation underpinning the activities of many not-for-profit conservation organisations (Carr et al. 2017). In this case, we have demonstrated that more effective management can be potentially achieved by explicitly linking knowledge of the behaviour of a species to more targeted on-ground action. We note some further applied research that would enhance the understanding of effective feral cat control, such as an understanding of the proficiency of a shooter and the types of optics used in scopes (i.e. infra-red or thermal), the type of trap arrays, lures and baits used, and examination of long term weather patterns preceding the control, including the distribution of prey in the landscape (Reynolds and Tapper 1996). In the present study, we simply assumed that a cat is recorded, trapped or removed when detected, even though there is a wide range of factors that influence the success of lethal control. This study presents a clear example of how to manage invasive predators using a combination of ecological knowledge and scenario-based adaptive management (Doherty and Ritchie 2017) more effectively.
Supplementary material
Supplementary material is available online.
Data availability
The data that support this study cannot be publicly shared due to concerns of human disturbance to core night parrot habitat and may be shared upon reasonable request to the corresponding author if appropriate.
Conflicts of interest
All authors declare no conflicts of interest.
Declaration of funding
This research received support from the Australian Government’s National Environmental Science Program through the Threatened Species Recovery Hub. Bush Heritage Australia provided in-kind support.
Acknowledgements
We are grateful to the Britton family (Brighton Downs), and Alistair MacDonald and Mick Cupitt (Mount Windsor) who provided logistical support. Mehdi Moradi provided advice about the use of the key functions in the R trajectories package. We also thank the editor, associate editor and two anonymous reviewers who improved the manuscript. This work was carried out under Queensland Department of Agriculture and Fisheries Animal Ethics Approval CA 2019/01/1250 and Queensland Biosecurity Permit PRID000510. We acknowledge the Maiawali people as the Traditional Owners of the area we conducted this work. We recognise and respect the enduring relationship that they have with their lands and waters, and we pay our respects to Elders past, present and emerging.
References
Ashby, E (1924). Notes on extinct or rare Australian birds, with suggestions as to some of the causes of their disappearance. Emu – Austral Ornithology 23, 178–183.| Notes on extinct or rare Australian birds, with suggestions as to some of the causes of their disappearance.Crossref | GoogleScholarGoogle Scholar |
Bengsen, AJ, Butler, JA, and Masters, P (2012). Applying home-range and landscape-use data to design effective feral-cat control programs. Wildlife Research 39, 258–265.
| Applying home-range and landscape-use data to design effective feral-cat control programs.Crossref | GoogleScholarGoogle Scholar |
Bureau of Meteorology (2017) Climate data online; Brighton Downs, Station Number 37007. Available at http://www.bom.gov.au/jsp/ncc/cdio/weatherData/av?p_display_type=dailyZippedDataFile&p_stn_num=37007&p_c=-273918624&p_nccObsCode=136&p_startYear=2021 [Accessed 12 February 2021]
Burnham KP, Anderson DR (1998) ‘Model Selection and Multimodel Inference: A Practical Information–theoretic Approach,’ 2nd edn. (Springer Science & Business Media, Inc.: New York. NY, USA)
Carr, B, Fitzsimons, J, Holland, N, Berkinshaw, T, Bradby, K, Cowell, S, Deegan, P, Koch, P, Looker, M, Varcoe, T, Walsh, P, and Weisenberger, F (2017). Capitalising on conservation knowledge: using conservation action planning, healthy country planning and the open standards in Australia. Ecological Management & Restoration 18, 176–189.
| Capitalising on conservation knowledge: using conservation action planning, healthy country planning and the open standards in Australia.Crossref | GoogleScholarGoogle Scholar |
Comer, S, Clausen, L, Cowen, S, Pinder, J, Thomas, A, Burbidge, AH, Tiller, C, Algar, D, and Speldewinde, P (2020). Integrating feral cat (Felis catus) control into landscape-scale introduced predator management to improve conservation prospects for threatened fauna: a case study from the south coast of Western Australia. Wildlife Research 47, 762–778.
| Integrating feral cat (Felis catus) control into landscape-scale introduced predator management to improve conservation prospects for threatened fauna: a case study from the south coast of Western Australia.Crossref | GoogleScholarGoogle Scholar |
Doherty, TS, and Ritchie, EG (2017). Stop jumping the gun: a call for evidence-based invasive predator management. Conservation Letters 10, 15–22.
| Stop jumping the gun: a call for evidence-based invasive predator management.Crossref | GoogleScholarGoogle Scholar |
Doherty, TS, Bengsen, AJ, and Davis, RA (2015). A critical review of habitat use by feral cats and key directions for future research and management. Wildlife Research 41, 435–446.
| A critical review of habitat use by feral cats and key directions for future research and management.Crossref | GoogleScholarGoogle Scholar |
Edwards, GP, de Preu, ND, Shakeshaft, BJ, and Crealy, IV (2000). An evaluation of two methods of assessing feral cat and dingo abundance in central Australia. Wildlife Research 27, 143–149.
| An evaluation of two methods of assessing feral cat and dingo abundance in central Australia.Crossref | GoogleScholarGoogle Scholar |
ESRI (2019) ‘ArcGIS 10.7.1.’ (Environmental Systesm Research Institute, Inc: Redlands, CA, USA)
Fancourt, BA, Augusteyn, J, Cremasco, P, Nolan, B, Richards, S, Speed, J, Wilson, C, and Gentle, MN (2021). Measuring, evaluating and improving the effectiveness of invasive predator control programs: feral cat baiting as a case study. Journal of Environmental Management 280, 111691.
| Measuring, evaluating and improving the effectiveness of invasive predator control programs: feral cat baiting as a case study.Crossref | GoogleScholarGoogle Scholar | 33272660PubMed |
Gorney, E, Clark, WS, and Yom-Tov, Y (1999). A test of the condition-bias hypothesis yields different results for two species of sparrowhawks (Accipiter). The Wilson Bulletin 111, 181–187.
Greenwell, CN, Calver, MC, and Loneragan, NR (2019). Cat gets its tern: a case study of predation on a threatened coastal seabird. Animals 9, 445.
| Cat gets its tern: a case study of predation on a threatened coastal seabird.Crossref | GoogleScholarGoogle Scholar |
Hijmans RJ (2019) ‘raster: Geographic Data Analysis and Modeling. R package version 3.0-2.’ Available at https://CRAN.R-project.org/package=raster
Iwaniuk, AN, Keirnan, AR, Janetzki, H, Mardon, K, Murphy, S, Leseberg, NP, and Weisbecker, V (2020). The endocast of the night parrot (Pezoporus occidentalis) reveals insights into its sensory ecology and the evolution of nocturnality in birds. Scientific Reports 10, 9258.
| The endocast of the night parrot (Pezoporus occidentalis) reveals insights into its sensory ecology and the evolution of nocturnality in birds.Crossref | GoogleScholarGoogle Scholar | 32518353PubMed |
Jackett, N, Greatwich, B, Swann, G, and Boyle, A (2017). A nesting record and vocalisations of the night parrot Pezoporus occidentalis from the East Murchison, Western Australia. Australian Field Ornithology 34, 144–150.
| A nesting record and vocalisations of the night parrot Pezoporus occidentalis from the East Murchison, Western Australia.Crossref | GoogleScholarGoogle Scholar |
Joseph, LN, Maloney, RF, and Possingham, HP (2009). Optimal allocation of resources among threatened species: a project prioritization protocol. Conservation Biology 23, 328–338.
| Optimal allocation of resources among threatened species: a project prioritization protocol.Crossref | GoogleScholarGoogle Scholar | 19183202PubMed |
Kearney, MR, Porter, WP, and Murphy, SA (2016). An estimate of the water budget for the endangered night parrot of Australia under recent and future climates. Climate Change Responses 3, 14.
| An estimate of the water budget for the endangered night parrot of Australia under recent and future climates.Crossref | GoogleScholarGoogle Scholar |
Kearney, SG, Kern, PL, and Kutt, AS (2021). A baseline terrestrial vertebrate fauna survey of Pullen Pullen; a significant conservation reserve in south-west Queensland. Australian Zoologist 41, 231–240.
| A baseline terrestrial vertebrate fauna survey of Pullen Pullen; a significant conservation reserve in south-west Queensland.Crossref | GoogleScholarGoogle Scholar |
Kearney, SG, Kern, PL, Murphy, SA, Janetzki, H, and Kutt, AS (2022). The diet of dingoes, feral cats and eastern barn owl on Pullen Pullen Reserve, southwest Queensland. Australian Mammalogy 44, 176–181.
| The diet of dingoes, feral cats and eastern barn owl on Pullen Pullen Reserve, southwest Queensland.Crossref | GoogleScholarGoogle Scholar |
Kennedy, M, Phillips, BL, Legge, S, Murphy, SA, and Faulkner, RA (2012). Do dingoes suppress the activity of feral cats in northern Australia? Austral Ecology 37, 134–139.
| Do dingoes suppress the activity of feral cats in northern Australia?Crossref | GoogleScholarGoogle Scholar |
Kranstauber, B, Kays, R, LaPoint, SD, Wikelski, M, and Safi, K (2012). A dynamic Brownian bridge movement model to estimate utilization distributions for heterogeneous animal movement. Journal of Animal Ecology 81, 738–746.
| A dynamic Brownian bridge movement model to estimate utilization distributions for heterogeneous animal movement.Crossref | GoogleScholarGoogle Scholar | 22348740PubMed |
Kutt, AS, Kearney, SG, and Kern, PL (2021). More than just night parrots: a baseline bird survey of Pullen Pullen Reserve, south-western Queensland. Australian Field Ornithology 38, 1–12.
| More than just night parrots: a baseline bird survey of Pullen Pullen Reserve, south-western Queensland.Crossref | GoogleScholarGoogle Scholar |
Lohr, CA, and Algar, D (2020). Managing feral cats through an adaptive framework in an arid landscape. Science of The Total Environment 720, 137631.
| Managing feral cats through an adaptive framework in an arid landscape.Crossref | GoogleScholarGoogle Scholar | 32325591PubMed |
Moseby, KE, Stott, J, and Crisp, H (2009). Movement patterns of feral predators in an arid environment – implications for control through poison baiting. Wildlife Research 36, 422–435.
| Movement patterns of feral predators in an arid environment – implications for control through poison baiting.Crossref | GoogleScholarGoogle Scholar |
Moseby, KE, Peacock, DE, and Read, JL (2015). Catastrophic cat predation: a call for predator profiling in wildlife protection programs. Biological Conservation 191, 331–340.
| Catastrophic cat predation: a call for predator profiling in wildlife protection programs.Crossref | GoogleScholarGoogle Scholar |
Moseby, KE, McGregor, H, and Read, JL (2020). Effectiveness of the Felixer grooming trap for the control of feral cats: a field trial in arid South Australia. Wildlife Research 47, 599–609.
| Effectiveness of the Felixer grooming trap for the control of feral cats: a field trial in arid South Australia.Crossref | GoogleScholarGoogle Scholar |
Murphy, SA, Austin, JJ, Murphy, RK, Silcock, J, Joseph, L, Garnett, ST, Leseberg, NP, Watson, JEM, and Burbidge, AH (2017a). Observations on breeding night parrots (Pezoporus occidentalis) in western Queensland. Emu – Austral Ornithology 117, 107–113.
| Observations on breeding night parrots (Pezoporus occidentalis) in western Queensland.Crossref | GoogleScholarGoogle Scholar |
Murphy, SA, Silcock, J, Murphy, R, Reid, J, and Austin, JJ (2017b). Movements and habitat use of the night parrot Pezoporus occidentalis in south-western Queensland. Austral Ecology 42, 858–868.
| Movements and habitat use of the night parrot Pezoporus occidentalis in south-western Queensland.Crossref | GoogleScholarGoogle Scholar |
Murphy, SA, Paltridge, R, Silcock, J, Murphy, R, Kutt, AS, and Read, J (2018). Understanding and managing the threats to night parrots in south-western Queensland. The Emu – Austral Ornithology 118, 135–145.
| Understanding and managing the threats to night parrots in south-western Queensland.Crossref | GoogleScholarGoogle Scholar |
Nutter, FB, Stoskopf, MK, and Levine, JF (2004). Time and financial costs of programs for live trapping feral cats. Journal of the American Veterinary Medical Association 225, 1403–1405.
| Time and financial costs of programs for live trapping feral cats.Crossref | GoogleScholarGoogle Scholar | 15552316PubMed |
Oppel, S, Burns, F, Vickery, J, George, K, Ellick, G, Leo, D, and Hillman, JC (2014). Habitat-specific effectiveness of feral cat control for the conservation of an endemic ground-nesting bird species. Journal of Applied Ecology 51, 1246–1254.
| Habitat-specific effectiveness of feral cat control for the conservation of an endemic ground-nesting bird species.Crossref | GoogleScholarGoogle Scholar |
Palmer R (1999) The ecology of feral cats in the mid-reaches of the Diamantina River in far western Queensland. Final report to the invasive species section of the biodiversity group, Environment Australia. (University of Queensland: St Lucia)
Pebesma E, Bivand R, Racine E, Sumner M, Cook I, Keitt T (2018) ‘sf: Simple Features for R,’ R package version 0.6-1. Available at https://cran.r-project.org/web/packages/sf/index.html
Pebesma E, Klus B, Graeler B, Gorte N, Moradi M (2021) ‘trajectories: Classes and Methods for Trajectory Data,’ version 0.2-3. Available at https://cran.r-project.org/web/packages/trajectories/index.html
Pettigrew J (1993) A burst of feral cats in the Diamantina – a lesson for the management of pest species? In ‘Cat Management Workshop Proceedings – 1993’. (Eds G Siepen, C Owens) pp. 25–32. (Queensland Department of Environment and Heritage: Brisbane, Qld, Australia)
Queensland Herbarium (2021) Regional Ecosystem Description Database (REDD). Version 12.1 (December 2021) (Queensland Department of Environment and Science: Brisbane)
R Core Team (2020) ‘R Version 4.1.2: a language and environment for statistical computing.’ (R Foundation for Statistical Computing 3.0.1: Vienna, Austria)
Reynolds, JC, and Tapper, SC (1996). Control of mammalian predators in game management and conservation. Mammal Review 26, 127–155.
| Control of mammalian predators in game management and conservation.Crossref | GoogleScholarGoogle Scholar |
Ruykys, L, and Carter, A (2019). Removal and eradication of introduced species in a fenced reserve: quantifying effort, costs and results. Ecological Management & Restoration 20, 239–249.
| Removal and eradication of introduced species in a fenced reserve: quantifying effort, costs and results.Crossref | GoogleScholarGoogle Scholar |
Signer, J, Fieberg, J, and Avgar, T (2019). Animal movement tools (amt): R package for managing tracking data and conducting habitat selection analyses. Ecology and Evolution 9, 880–890.
| Animal movement tools (amt): R package for managing tracking data and conducting habitat selection analyses.Crossref | GoogleScholarGoogle Scholar | 30766677PubMed |
Thurfjell, H, Ciuti, S, and Boyce, MS (2014). Applications of step-selection functions in ecology and conservation. Movement Ecology 2, 4.
| Applications of step-selection functions in ecology and conservation.Crossref | GoogleScholarGoogle Scholar | 25520815PubMed |
Vernes, K, Dennis, A, and Winter, J (2001). Mammalian diet and broad hunting strategy of the dingo (Canis familiaris dingo) in the wet tropical rain forests of northeastern Australia. Biotropica 33, 339–345.
| Mammalian diet and broad hunting strategy of the dingo (Canis familiaris dingo) in the wet tropical rain forests of northeastern Australia.Crossref | GoogleScholarGoogle Scholar |
Woinarski, JCZ, Murphy, BP, Legge, SM, Garnett, ST, Lawes, MJ, Comer, S, Dickman, CR, Doherty, TS, Edwards, G, Nankivell, A, Paton, D, Palmer, R, and Woolley, LA (2017a). How many birds are killed by cats in Australia? Biological Conservation 214, 76–87.
| How many birds are killed by cats in Australia?Crossref | GoogleScholarGoogle Scholar |
Woinarski, JCZ, Woolley, LA, Garnett, ST, Legge, SM, Murphy, BP, Lawes, MJ, Comer, S, Dickman, CR, Doherty, TS, Edwards, G, Nankivill, A, Palmer, R, and Paton, D (2017b). Compilation and traits of Australian bird species killed by cats. Biological Conservation 216, 1–9.
| Compilation and traits of Australian bird species killed by cats.Crossref | GoogleScholarGoogle Scholar |
Wysong, ML, Iacona, GD, Valentine, LE, Morris, K, and Ritchie, EG (2020). On the right track: placement of camera traps on roads improves detection of predators and shows non-target impacts of feral cat baiting. Wildlife Research 47, 557–569.
| On the right track: placement of camera traps on roads improves detection of predators and shows non-target impacts of feral cat baiting.Crossref | GoogleScholarGoogle Scholar |