

Apparent resistance to brodifacoum in Rattus rattus in a New Zealand site with no history of anticoagulant-based rodent control
Suman P. K. Sran A * , Brett G. Gartrell B , Penny Fisher C and Doug P. Armstrong A
A * , Brett G. Gartrell B , Penny Fisher C and Doug P. Armstrong A
A Wildlife Ecology Group, School of Agriculture and Environment, Massey University, PB 11222, Palmerston North, New Zealand.
B School of Veterinary Science, Massey University, PB 11222, Palmerston North, New Zealand.
C Research Associate, Landcare Research, Lincoln 7608, New Zealand.
Wildlife Research 50(1) 28-38 https://doi.org/10.1071/WR21064
Submitted: 7 July 2020 Accepted: 16 April 2022 Published: 7 July 2022
© 2023 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Context: Anticoagulants have been used in New Zealand for decades, but few data are available on the sustainability of these toxins for rodent control. It is important to regularly monitor for resistance in long-term brodifacoum-use areas and establish a database for future references.
Aims: This study aimed to estimate the effective dose (ED50) of brodifacoum for ship rats from an area of New Zealand with no history of brodifacoum use, in order to establish a blood-clotting response test for assessing resistance in rodent populations from other areas.
Methods: A ranging study was conducted whereby successive groups of ship rats were administered brodifacoum doses that were increased or decreased progressively, until an International Normalised Ratio (INR) of 3.6 was reached. Linear regression was used to model the relationship between dose and INR, and ED50 dose was estimated using the resulting model.
Results: None of the rats appeared susceptible to brodifacoum at previously reported LD50 exposures for this species. The ED50 of brodifacoum was estimated to be 2.88 mg/kg for males and 3.81 mg/kg for females. These values are 6–8 times greater than the previously published lethal dose values for ship rats in New Zealand.
Conclusions: Blood-clotting inhibition was detected in the rats only following high doses of brodifacoum, which may indicate resistance within the sampled population.
Implications: Relatively low susceptibility, or resistance, to brodifacoum in New Zealand ship rats may be mediated by spatial connections between areas with different histories and patterns of anticoagulant rodenticide use.
Keywords: anticoagulants, blood clotting response test, brodifacoum, effective dose, pest management, Rattus rattus, resistance, rodent control.
Introduction
Anticoagulant resistance in rodents was discovered soon after first-generation anticoagulant rodenticides (FGARs) were introduced, when warfarin treatment failed to control Norway rat (Rattus norvegicus) populations in Scotland (Boyle 1960). Subsequently, there were many reports throughout Europe of resistance to warfarin as well as other first-generation anticoagulants like coumatetralyl and diphacinone (Bentley 1968; Lund 1969; Greaves and Rennison 1973). With declining efficacy of FGARs against resistant rodent populations, more potent second-generation anticoagulants (SGARs) were marketed in the 1970s (Hadler and Shadbolt 1975). Bromadialone and diphanacoum were the first to be marketed and were initially found to be effective against rats resistant to FGARs (Hadler et al. 1975; Marsh 1977). However, resistance to these compounds was subsequently reported (Redfern and Gill 1978; Greaves and Cullen-Ayers 1988; Quy et al. 1995). Three other SGARs (brodifacoum, flocoumafen and difethialone) were introduced to the market as ‘resistance breakers’, being effective against rodent populations resistant to FGARs and other SGARs (Buckle et al. 2013). Although brodifacoum and flocoumafen have proven to be very effective in rodent control around the world, continuous long-term use of any anticoagulant rodenticide may lead to selection of resistance-conferring mutations by eliminating the susceptible individuals of the population. Many studies have confirmed resistance to anticoagulants throughout the world. For example, anticoagulant resistance has been widely documented in Europe (Rost et al. 2004; Pelz et al. 2005), the United States (Jackson and Ashton 1979), Indonesia (Andru et al. 2013), Japan (Ishizuka et al. 2007) and China (Liang 2005). Most of the documented literature on resistance to anticoagulants in rats concerns Norway rats (Li et al. 2004; Rost et al. 2009; Rymer 2017). Less work has been done on potential resistance in ship rats (black rats, Rattus rattus), because they are not a major pest species in countries where extensive research on anticoagulant resistance has been done (Berny et al. 2018). Resistance to FGARs like warfarin and the more potent SGARs, bromadialone and difenacoum, has been reported in ship rats in Denmark, France, USA, Germany, UK and Japan (Desidiri et al. 1978; Lund 1984; Myllymäki 1995; Ishizuka et al. 2007).
Anticoagulants have been used in New Zealand for decades, including for the extensive control of ship rats, which are a major invasive predator in New Zealand forests. However, very little research has been done on anticoagulant resistance in ship rats or other rodents in New Zealand. The New Zealand Department of Conservation has used brodifacoum extensively for island rodent eradications, but stopped using it for mainland control operations in 2002 due to concerns about its persistence in the environment (Eason et al. 2002). However, it continues to be used extensively on the mainland by regional councils, community groups and private landowners. Recently, pest control operations in New Zealand have come under scrutiny for their animal welfare impacts (Beausoleil et al. 2016), and anticoagulant poisons were highlighted as being the least favourable option from this perspective. In order to evaluate the future role of anticoagulant poisons in pest control in New Zealand, it is necessary to screen for potential anticoagulant resistance in ship rat populations in areas with a history of long-term exposure to anticoagulants.
Anticoagulant resistance was suspected in parts of Auckland and on Raoul Island in 1978 but no investigation took place. However, rats were eradicated successfully in Raoul Island using brodifacoum by aerial application in 2002 (Wodzicki 1978). The first evidence of heritable genetic resistance was established in New Zealand in 2017, with identification of a Tyr25Phe mutation in eight out of 482 ship rats screened (Goulois et al. 2016). Although this mutation is associated with resistance to all FGARs and some SGARs (Goulois et al. 2016), demonstration of phenotypic changes is required to conclusively document anticoagulant resistance in New Zealand rodent populations. Novel mutations affecting anticoagulant resistance like Ala14Val and Ala26Val have also been identified in ship rats in New Zealand (Cowan et al. 2017), but they need to be assessed using lethal feeding period (LFP) tests or blood-clotting response (BCR) tests to assess the degree of resistance it confers towards each anticoagulant compound used in New Zealand. If the mutations confer little practical resistance, anticoagulants can still be effective as a control strategy. Similarly, if these mutations cause resistance to FGARs only, SGARs may still be an option for effective control. The information obtained from LFP and BCR tests is therefore crucial for making practical decisions about control of resistant ship rat populations.
The standard BCR test (Prescott et al. 2007) for Norway rats and house mice (Mus musculus) involves comparing blood coagulation times before and after administration of a sublethal dose of anticoagulant. Animals that show considerably increased blood-clotting times after administration of a dose of anticoagulant are considered susceptible (Erica Gill et al. 1993). This test does not rely on poisoning mortality of animals, as is the case for LFP tests, and is therefore considered more humane, has fewer ethical constraints, and can be performed in 24 h. BCR tests are very sensitive and can detect even small differences in the susceptibility of rodent species to specific anticoagulant rodenticides (Prescott and Buckle 2000).
The sublethal dose administered to animals to test for blood-clotting responses is called an effective dose (ED50). For example, an ED50 dose will not kill the animal, but will produce a critical measurable change in blood-clotting time in 50% of individuals (Prescott et al. 2007). Once the ED50 has been calculated for a species in the absence of anticoagulant exposure, it can be tested whether the ED50 is significantly higher in populations with a history of exposure, giving evidence of resistance. The ED50 value may or may not differ significantly between males and females (Prescott et al. 2007).
The initial aim of our study was to determine the effective dose (ED50) of brodifacoum for a New Zealand population of ship rats with no known exposure to brodifacoum. This would then provide a baseline that could be used in BCR tests to test for resistance in ship rats in parts of New Zealand with long-term brodifacoum use. Because there are no laboratory-strain ship rats available in New Zealand, we captured wild ship rats from an area with no documented history of anticoagulant use and conducted a ranging experiment to determine the brodifacoum dose that produced a blood-clotting response in 50% or males and females.
Material and methods
Study site
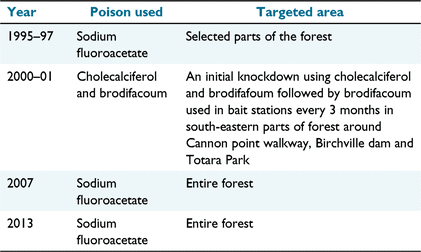
We captured wild ship rats from the north-west region (40°58′34.66″S, 175°1′35.45″E) of Akatarawa forest (Fig. 1) in New Zealand, a site where there was no prior anticoagulant history and the pest control history was well documented (Table 1). Akatarawa Forest is in the steep hill country of the Akatarawa Ranges, covering 15 500 ha, and is managed by Greater Wellington Regional Council (GWRC). Trapping was carried out in the north-west region of Akatarawa because there was temporary use of anticoagulants in the south-east region (18 km away) in 2000–01 (Table 1). Brodifacoum has also been used in the neighbouring private lands by GWRC.

|
|
Trapping and maintenance
Forty single-catch iron-mesh rat traps were set up close to Maungatotukutuku access point. To avoid catching rats from surrounding areas, no traps were placed within 1 km of the forest boundary. The rat population was at low density because of ongoing management using the non-anticoagulant toxicant sodium fluoroacetate (1080), hence the 12 nights of trapping effort (40 traps set per night) it took to catch the 36 rats used in the experiment (Table 2). Traps were placed 100 m apart and baited with cheese and peanut butter. Traps were checked every morning, and captured animals were transported in their traps to Massey University, Palmerston North within 24 h. On arrival at Massey University, each animal was transferred from its trap to an individual 30 cm × 30 cm × 25 cm rat cage, and was acclimatised for 15 days before the commencement of the experiment.

|
All rats captured were confirmed to be ship rats based on their morphology (Cunningham and Moors 1996). Only mature and healthy rats were used for the experiment. Any pregnant females and immature rats caught were euthanased by cervical dislocation while under general anaesthesia (isoflurane). Cages were provided with sawdust for bedding and shredded newspaper for nesting material, and food and water were provided ad libitum. Rats were provided with rat cereal feed pellets (Diet 86, manufactured at the feed mill at Massey University). The laboratory was maintained at 20 ± 2°C on a 12-h light/dark cycle.
Vitamin K is an important component of the blood-clotting system in mammals. Deficiency of vitamin K can lead to prolonged blood-clotting time. Similarly, excess of vitamin K can act as antidote to brodifacoum (Prescott et al. 2007). The level of vitamin K in ship rats captured from wild is unknown, and therefore it is important to provide rats with the appropriate amount of vitamin K for a sufficient acclimatisation period. The diet provided contained the minimum amount of vitamin K required by a rat (1 mg/kg per diet; Fu et al. 2007) to avoid vitamin K deficiency, and any initial excesses of vitamin K in their bodies was expected to have been utilised during the 15-day acclimatisation period.
Brodifacoum dose formulation
Successive doses of brodifacoum were formed by dissolving the required amount of brodifacoum in polyethylene glycol 200 (PEG200) in a beaker. The brodifacoum and polyethylene glycol (PEG) 200 were obtained from Sigma Aldrich Company Ltd, Auckland, New Zealand. The solution was shaken and heated to 60°C in a laboratory incubator overnight to aid dissolution of brodifacoum and achieve the required concentrations. The beaker was covered with aluminium foil to avoid light throughout the procedure. This methodology is standard for preparing brodifacoum formulations, which have been shown to be thermally stable below 150°C with no decomposition or transformation (https://circabc.europa.eu/sd/a/d88b968d-e5b8–4b4e-84bb-79a8b41acc4b/2010%2001%2017%20Final%20combined%20AR%20Brodifacoum_rev%20_2_.pdf).
Experimental design
The experiment involved a ranging study whereby ship rats of each sex were divided into groups of three, including a control group and multiple treatment groups. Rather than deciding the treatment groups a priori, the doses for each successive treatment were adjusted up or down until the effective dose was determined with sufficient precision (see statistical analysis). Such procedures reduce the number of animals used (Bruce 1985).
Published LD50 values of brodifacoum for ship rats include 0.46 mg/kg (O’Connor and Booth 2001) to 0.77 mg/kg (Mathur and Prakash 1981). The ED is a sublethal dose that produces a critical measurable response to an administered dose of anticoagulant. Therefore, it is always less than lethal dose (LD50) of brodifacoum, i.e. the dose that kills 50% individuals in a susceptible population. Based on the highest published LD50 dose of brodifacoum for ship rats 0.77 mg/kg, the first treatment groups of each sex were administered a dose of 0.72 mg/kg brodifacoum dissolved in PEG, and doses for each successive group were then expected to be decreased progressively until an international normalised ratio (INR) of 3.6 was reached.
The INR in this context is derived by calculating the ratio of the post-treatment clotting time to the pre-treatment clotting time, which is then multiplied by the International Sensitivity Index (ISI). A ship rat is considered to be a responder (susceptible) to a given dose of anticoagulant when it has an INR ≥ 3.6 (3.5–3.8) (Garg and Singla 2015). Every reagent or equipment manufactured to calculate blood-clotting time or prothrombin time is assigned an ISI value. ISI is indicative of a particular reagent or equipment compared with international reference tissue factor. An ISI ranging between 0.9 and 1.4 indicates a highly sensitive reagent. A CoaguChek® XS Plus analyser (Roche Diagnostics International Ltd) was used to determine blood-clotting times of animals before and after treatment, with an allocated ISI value of 1.
Pre-treatment
After the 15-day acclimatisation period, a baseline blood sample was taken from each rat before treatment. The rat was placed under general anaesthesia using inhalational isoflurane gas in oxygen, delivered via induction anaesthetic chamber initially and then via facemask for blood sampling. Blood was collected from the lateral saphenous vein using a 25-gauge needle. Up to 20 μL of blood were collected, and blood-clotting time was measured immediately using the blood-clotting time analyser. The analyser measures clotting times within the range of of 9.6–96 s; therefore, for rats with clotting times less than 9.6 s, we were only able to determine that this was the maximum clotting time.
Treatment
Immediately following the pre-treatment blood sample, the rats in each group were provided with the appropriate subcutaneous dose of brodifacoum dissolved in PEG. The control group of rats was administered with PEG200 only. In all cases the total volume injected was 0.08 mL per 10 g bodyweight of rat. The rats were returned to their cages while they were still under influence of anaesthesia, and recovered 2–3 min later.
Post-treatment
Up to 20 μL of blood was collected from each rat 24 h after administering the dose of brodifacoum. Rats were anaesthetised as described above, then the post-treatment blood sample was collected via cardiac puncture with a 22-gauge needle. After blood collection, the rats were euthanased by cervical dislocation while still under general anaesthesia. Blood-clotting time was measured immediately as for the pre-treatment sample. The control group was also sampled 24 h after the PEG was administered, but at this stage 20 μL of blood was collected from each rat in same manner as described for pre-treatment blood sampling. These rats were monitored twice daily for at least 7 days to study any ill effects of PEG on them. They were then anaesthetised and euthanased by cervical dislocation. All procedures involving the use of animals were approved by the Massey University Animal Ethics Committee (Protocol 15/97).
Statistical analysis
Probit analysis is used commonly to calculate the ED50 of different rodenticides (Prescott et al. 2007; Akçay 2013). In probit analysis, each animal’s blood-clotting times are reduced to a binary variable, indicating whether it is considered to be a responder. The approach is therefore wasteful of data, and inappropriate for a sample of 36 animals. We therefore used linear regression to model the relationship between dose and INR, and used the resulting model to estimate the ED50 dose, i.e. the dose yielding a median INR of 3.6. We tested whether the slope of the relationship differed between males and females. Because we only had maximum pre-treatment clotting times for some animals, we used multiple imputation to model these missing values (Carpenter and Kenward 2013), with the maximum set as the upper limit and assuming that the pre-treatment clotting times were normally distributed. We used OpenBUGS (Spiegelhalter et al. 2014) to model the data, both because this approach facilitated use multiple imputation, and because it allowed the effective dose to be easily calculated as a derived parameter. Parameter estimates are reported ±1 s.e.
Results
BCR test
The 36 rats used in the experiment were healthy mature adults. The average weight of males was 169 g (range = 142–219) and the average weight of females was 148 g (range = 130–162).
The average normal blood-clotting time in ship rats was estimated to be 9.6 s ± 0.1, including modelling of the missing values, and this was consistent for males and females (Table 3). No clinical signs of lethargy, distress or haemorrhage were identified before the end of the 24-h period in any of the rats. BCR tests for the control group showed no difference in clotting times of blood collected before and after the administration of PEG (Table 4). No ill effects of PEG were observed in the 7-day period after the dose of PEG was administered.

|

|
Male and female rats administered with the initial dose of 0.72 mg/kg produced an INR ranging between 1.86–2.38 and 1.86–2.13 respectively, so were all below the value of 3.6 considered a threshold response for susceptible ship rats to a given anticoagulant dose (Fig. 2). The dose of brodifacoum was increased successively to 0.8, 1.0 and 2.0 mg/kg for subsequent male ship rat groups, and still no rats had INR values ≥3.6. The dose was further increased to 3 mg/kg, to which all rats responded with an INR ≥ 3.6, with values ranging between 3.63 and 4.00. A subsequent reduction to 2.5 mg/kg in the final group again resulted in none of the male rats having INR values ≥3.6.

|
The brodifacoum dose was increased directly to 3 mg/kg in the second group of female rats because all the male rats in the group responded to this dose with INR ≥ 3.6. However, none of female rats responded to this dose. An increase to 4 mg/kg resulted in all three females responding with INR values ranging from 3.84 to 4.25. A reduction to 3.5 mg/kg for the final group resulted in one of the three females having an INR ≥ 3.6.
Estimation of effective dose
The data indicated that it was reasonable to assume a linear relationship between INR and dose (Fig. 3). Because there was no apparent effect of the PEG, it was also assumed in the model that INR = 1 when the dose = 0. There was a significant difference in the slope of the relationship for male and female ship rats (P = 0.0091; Fig. 3). The ED50 of brodifacoum was estimated to be 2.88 ± 0.23 mg/kg for males and 3.81 ± 0.32 mg/kg for females.

|
Discussion and conclusions
In the present study, successive doses of brodifacoum were administered to each group of rats, ranging from 0.72 to 3.00 mg/kg in male ship rats and 0.72 to 4.00 mg/kg in female ship rats, until an INR ≥ 3.6 was obtained. From this experiment, the ED50 of brodifacoum for ship rats at Akatarawa forest was estimated to be 2.88 mg/kg for males and 3.81 mg/kg for females. The effective dose is a sublethal dose that produces a critical measurable response to an administered quantity of anticoagulant; therefore, it is always a less-than-lethal dose (LD50) of brodifacoum, i.e. the dose that kills 50% individuals in a susceptible population. Reported LD50 doses of brodifacoum vary in rodent species populations around the world, and may be affected by several factors including historic exposure of poison in ancestors, age, sex, nutritional status and different experimental approaches to calculate lethal dose (Kaukeinen and Rampaud 1986; Moussa 2005). The published data suggest a range of 0.46–0.77 mg/kg for LD50 values of brodifacoum for ship rats (Dubock and Kaukeinen 1978; Fisher 2005), which is lower than the ED50 values found in this study, possibly indicating a relative tolerance, or outright resistance, to brodifacoum in our study population.
The highest published LD50 value of brodifacoum for ship rats, 0.77 mg/kg, was from India (Mathur and Prakash 1981). In New Zealand, the LD50 value of ship rats was found to be 0.46 mg/kg in a study carried out in a brodifacoum-free area on the west coast of the South Island (O’Connor and Booth 2001). The west coast of the South Island is more remote in terms of human habitation than Akatarawa forest, from which rats were captured for the present study. The effective dose estimated for male ship rats in this study (2.88 mg/kg) is nearly 3.7 times the highest published lethal dose of brodifacoum, and 6.2 times the published lethal dose in New Zealand. In females, the value of 3.81 mg/kg is nearly five times the highest published lethal dose in India and 8.2 times the published lethal dose in New Zealand. The calculated effective doses in this study are very high compared with the two published LD50 values.
In the present study, all the ship rats with INR values < 3.6 at a given dose were categorised as non-responsive to standard doses of anticoagulants based on Garg and Singla’s (2015) research, indicating this was a suitable criterion for R. rattus. Prescott et al. (2007) earlier suggested that an INR > 5.0 was a suitable indicator of responsiveness to an anticoagulant dose in rodents (Norway rats and house mice). However, none of the rats in our study were given a high enough dose to achieve an INR > 5.0, so our conclusions are not sensitive to this difference in cut-off INR value. In any case, our estimated ED50 values were clearly still higher than initially expected, based on known LD50 values of brodifacoum for ship rats.
Most of the studies involving blood-clotting response tests have been focused on Norway rats and house mice. There is no BCR test standardised for ship rats providing us with an ED50 value for any known anticoagulant (Berny et al. 2018). The only previous BCR test on ship rats was for detecting resistance to bromadialone in India (Garg et al. 2017). However, in that study, the dose used for the BCR test was calculated based on a published lethal dose rather than conducting an experiment to determine the effective dose. Therefore, we do not have any data to compare the effective dose and lethal dose for any anticoagulant in ship rats, but we can make comparisons with published data for LD50 and ED50 values of brodifacoum in Norway rats and house mice (Table 4). Both species are major pests in most parts of the world where anticoagulant resistance has been studied extensively. There are published effective doses of brodifacoum available for Norway rats from studies around the world, and those effective doses are far less than the highest published lethal dose. Similarly, for house mice, published effective doses are always lower than the published lethal doses.
The second dose administered to the rats, 0.8 mg/kg, exceeded the highest published lethal dose of 0.77 mg/kg. Therefore, all the animals were expected to respond to this dose and produce an INR ≥ 3.6. A recent study undertaken on Lord Howe Island in Australia, in preparation for a rodent eradication programme, found doses of 0.8 mg/kg administered to ship rats achieved 100% mortality, irrespective of the long history of use of FGARs and SGARs on the island since the 1960s. Although there is no formal record of brodifacoum use by residents on the island, it is thought to have exceeded 400 kg per annum since 2000 (Wheeler et al. 2019). Contrary to these results, none of the ship rats from Akatarawa forest with no recent history of anticoagulant use responded to 0.8 mg/kg brodifacoum dose with INR ≥ 3.6.
Susceptibility may vary among populations of ship rats due to differences in historic exposure to the toxin, but also to natural variation. Wild rodents are expected to be more variable in their responses than laboratory strains (Prescott et al. 2007), thus laboratory strains are generally preferred for evaluating the effective dose of a poison. Due to the absence of laboratory ship rats in New Zealand, we attempted to measure the effective dose for susceptible ship rats using a wild population with no known history of anticoagulant exposure. The apparently high level of anticoagulant tolerance or resistance in this population means we were unable to confirm that our estimates represented baseline ED50 values for susceptible ship rats in the New Zealand context.
One potential explanation for this resistance would be a high intake of vitamin K in diet in Akatarawa forest. Vitamin K is an antidote of anticoagulants and would result in no or low response to the administered brodifacoum dose. However, the acclimatisation period of 15 days should have led to utilisation of any extra vitamin K in the body, and the diet provided to ship rats during this period only contained the minimum amount of vitamin K required by rats.
Although the lack of response of ship rats to lower doses like 0.72–1.00 mg/kg could be explained based on natural variance in susceptibility to brodifacoum, it is unclear how a general low susceptibility or tolerance due to natural variance contributes to lack of response to substantially doses in the 2.5–3.0 mg/kg range. The lack of blood-clotting inhibition in response to such doses is plausibly due a high prevalence of some form of resistance to brodifacoum within the ship rat population at Akatarawa forest.
Although brodifacoum or any other anticoagulant was never used in the part of the forest from where rats were captured, it is used extensively in the neighbouring areas by private landowners as well as Greater Wellington Regional Council. Ship rats exposed to brodifacoum from these areas may have been invading the forest and may be the cause of this resistance. Sodium fluoroacetate (1080) has been the only poison used in pest control operations in north-western Akatarawa forest, applied every 4–6 years since 2001. 1080 is an acute metabolic poison and has a mode of action different from anticoagulants. Because 1080 is a very effective rodenticide, there is a sharp decline in rat population in the forest each time it is used (Innes and Williams 1991; Innes et al. 1995). This may be facilitating a more rapid invasion of ship rats into the forest. The ship rat population in the area is known to recover quickly after the use of 1080 pellets, and in general, ship rat populations are known to recover in about 6–12 months (Barron et al. 2014). This time window provides opportunity for ship rats to move to the forest with abundant food and space with little competition. Both male and female ship rats have a home range of 0.5–1.0 ha, but males increase their home ranges during the breeding season (Dowding and Murphy 1994; Harper and Rutherford 2016). In beech forests, ship rats have been found to have a home range of up to 11.4 ha (Pryde et al. 2005). Also, ship rats are faster and more agile than Norway rats (Foster et al. 2011). Breeding in ship rats is polyoestrous, and the gestation period is usually 20–22 days long with a litter size ranging from 3–10 pups. Growth of pups is rapid, and they reach maturity in 3–4 months (Bentley and Taylor 1965). Taking into consideration the fast generation time, high mobility of ship rats and high effective dose evaluated from an area of historically negligible anticoagulant use, it seems possible that anticoagulant resistance is present in ship rats in the Wellington region. It is also likely that ship rats from areas of long-term brodifacoum use (20+ years) may have even higher resistance to brodifacoum than that observed in this study.
Further investigation needs to be done to confirm physiological and/or genetic basis for possible resistance in New Zealand ship rats. Genetic analysis of the rat tail samples preserved from this experiment may indicate whether the resistance to brodifacoum in ship rats at Akatarawa Forest is due to heritable resistance. If this is the case, rodent pest management by anticoagulants may become a challenge in near future. A rodent resistant to a more toxic anticoagulant is known to develop resistance to less toxic anticoagulants to which it may have even never been exposed (Pelz and Prescott 2015). This removes the option of using alternative anticoagulants to control rodent pest population with lowered susceptibility to brodifacoum. In this situation, effective rodent control may be achieved only by increasing the concentration of brodifacoum in the baits, leading to an increase in secondary and non-target poisoning, or by using alternative poisons like cholecalciferol or 1080. Continual use of brodifacoum may also encourage further development of resistance in ship rat populations.
Another factor affecting pest control operation may be the higher effective dose found in females than males. This may be due to two factors. First, basal activity of vitamin K-dependent blood clotting factors (II, VII and X) is higher in females than in males, and there is also higher plasmic half life of prothrombin and blood factor X in females (Lefebvre et al. 2016). Anticoagulants are known to inhibit vitamin K recycling, leading to reduced blood clotting that may prove to be fatal. Higher activity of vitamin-K dependent blood clotting factors would reduce susceptibility of females to brodifacoum. Second, there is sexually dimorphic expression of cytochrome P450 complex in rats. Cytochrome P450 plays a crucial role in metabolism of anticoagulants in the body. In a study done on bromadialone-resistant rats, different CYP450 isoforms have been known to express 5–80-fold higher in bromadialone-resistant female rats as compared with susceptible female rats, meaning the resistant females can metabolise bromadialone 5–80 times faster as compared with susceptible females. In contrast, these enzymes expressed only 1.7–3.0-fold in resistant male rats (Markussen et al. 2008).
If brodifacoum resistance in female ship rats is accompanied by one or more heritable mutations, it may pose a future threat to effective rodent control in New Zealand because resistant female rats are likely harder to control than resistant males (Rymer 2017). This is because only one male is required to mate with several females, and therefore a reduction in male density alone is expected to have little effect on population dynamics. Female ship rats can reach maturity at an age of 3–4 months and are polyoestrous, producing up to 10 pups in a litter (Bentley and Taylor 1965). Therefore, with brodifacoum resistant females passing resistance to the next generations, resistance can spread at alarming rates and ship rat populations may recover rapidly after application of an anticoagulant.
At present, there is a need to consider poisons other than anticoagulants for pest management in New Zealand. Also, regular monitoring of susceptibility of each anticoagulant poison used for rodent species in New Zealand needs to be done. Assessment of susceptibility in areas with long-term use of anticoagulant poisons must be a priority to ensure effective rodent control in the future. Although we were unable to evaluate the effective dose of a susceptible population, the estimated effective doses in this study are still useful for comparing with areas of known brodifacoum use (i.e. to assess whether the apparent level of resistance is even greater in these areas). We also recommend evaluating the effective dose using ship rats captured from isolated areas with no possible brodifacoum use, such as the few remaining populations on offshore islands, as a comparison with the ship rats used in this study.
Data availability
All the data that supports this study are available in the article.
Conflicts of interest
Penny Fisher is an Associate Editor of Wildlife Research and did not at any stage have editor-level access to this manuscript while in peer review, as is the standard practise when handling manuscripts submitted by an editor of this journal. The authors have no further conflicts of interest to declare.
Declaration of funding
This research was funded by the Massey University research fund and GWRC Science and Research Fund. This funding contributed towards expenses related to field and laboratory trials.
Acknowledgements
We are greatly thankful to GWRC for providing information about the study site and Landcare Research, Lincoln, for providing cages for acclimatisation period of animals.
References
Akçay, A (2013). The calculation of LD50 using probit analysis. The FASEB Journal 27, 1217.28.| The calculation of LD50 using probit analysis.Crossref | GoogleScholarGoogle Scholar |
Andru, J, Cosson, JF, Caliman, JP, and Benoit, E (2013). Coumatetralyl resistance of Rattus tanezumi infesting oil palm plantations in Indonesia. Ecotoxicology 22, 377–386.
| Coumatetralyl resistance of Rattus tanezumi infesting oil palm plantations in Indonesia.Crossref | GoogleScholarGoogle Scholar | 23264020PubMed |
Barron M, Griffiths J, Thomson C, Perry M, Clarke D, Sweetapple P (2014) How rapidly do rat populations recover after control with 1080? Kararehe Kino Issue 23. Manaaki Whenua, New Zealand.
Beausoleil, NJ, Fisher, P, Littin, KE, Warburton, B, Mellor, DJ, Dalefield, RR, and Cowan, P (2016). A systematic approach to evaluating and ranking the relative animal welfare impacts of wildlife control methods: poisons used for lethal control of brushtail possums (Trichosurus vulpecula) in New Zealand. Wildlife Research 43, 553–565.
| A systematic approach to evaluating and ranking the relative animal welfare impacts of wildlife control methods: poisons used for lethal control of brushtail possums (Trichosurus vulpecula) in New Zealand.Crossref | GoogleScholarGoogle Scholar |
Bentley, EW (1968). The warfarin resistance problem in England and Wales. Schriftenreihe des Vereins fur Wasser-, Boden- und Lufthygiene 32, 19–25.
Bentley, EW, and Taylor, EJ (1965). Growth of laboratory-reared ship rats (Rattus rattus L.). Annals of Applied Biology 55, 193–205.
| Growth of laboratory-reared ship rats (Rattus rattus L.).Crossref | GoogleScholarGoogle Scholar |
Berny P, Esther A, Jacob J, Prescott C (2018) Development of resistance to anticoagulant rodenticides in rodents in anticoagulant rodenticides and wildlife. In ‘Emerging Topics in Ecotoxicology’. (Eds NW Van den Brick, JE Elliot, RF Shore, BA Rattner) pp. 259–286. (Springer International Publishing AG)
Boyle, CM (1960). Case of apparent resistance of Rattus norvegicus Berkenhout to anticoagulant poisons. Nature 188, 517.
| Case of apparent resistance of Rattus norvegicus Berkenhout to anticoagulant poisons.Crossref | GoogleScholarGoogle Scholar |
Bruce, RD (1985). An up-and-down procedure for acute toxicity testing. Fundamental Applied Toxicology 5, 151–157.
| An up-and-down procedure for acute toxicity testing.Crossref | GoogleScholarGoogle Scholar | 3987991PubMed |
Buckle, A, Endepols, S, Klemann, N, and Jacob, J (2013). Resistance testing and the effectiveness of difenacoum against Norway rats (Rattus norvegicus) in a tyrosine139cysteine focus of anticoagulant resistance, Westphalia, Germany. Pest Management Science 69, 233–239.
| Resistance testing and the effectiveness of difenacoum against Norway rats (Rattus norvegicus) in a tyrosine139cysteine focus of anticoagulant resistance, Westphalia, Germany.Crossref | GoogleScholarGoogle Scholar | 23208747PubMed |
Carpenter JR, Kenward MG (2013) ‘Multiple Imputation and its Application.’ (John Wiley & Sons Ltd: Chichester, UK)
Cowan, PE, Gleeson, DM, Howitt, RLJ, Ramón-Laca, A, Estherd, A, and Pelz, H-J (2017). Vkorc1 sequencing suggests anticoagulant resistance in rats in New Zealand. Pest Management Science 73, 262–266.
| Vkorc1 sequencing suggests anticoagulant resistance in rats in New Zealand.Crossref | GoogleScholarGoogle Scholar | 27117082PubMed |
Cunningham DM, Moors PJ (1996) ‘Guide to the Identification and Collection of New Zealand Rodents.’ (Department of Conservation: Wellington, New Zealand)
Desidiri, D, Aldighieri, R, Le Louet, M, and Tardieu, A (1978). Supplementary report on the study of resistance to coumafene in Rattus rattus observed at Marseilles. Bulletin de la Société de Pathologie Exotique 71, 301–303.
Dowding, JE, and Murphy, EC (1994). Ecology of ship rats (Rattus rattus) in a kauri (Agathis australis) forest in Northland, New Zealand. New Zealand Journal of Ecology 18, 19–27.
Dubock AC, Kaukeinen DE (1978) Brodifacoum (Talon rodenticide), a novel concept. In ‘Proceedings of the 8th Vertebrate Pest Conference’, 7–9 March 1978, Sacramento, CA, USA. (Ed. AWE Howard) pp. 127–137. (University of California: Davis, CA, USA)
Eason, CT, Murphy, EC, Wright, GRG, and Spurr, EB (2002). Assessment of risks of brodifacoum to non-target birds and mammals in New Zealand. Ecotoxicology 11, 35–48.
| Assessment of risks of brodifacoum to non-target birds and mammals in New Zealand.Crossref | GoogleScholarGoogle Scholar | 11898799PubMed |
Erica Gill, J, Kerins, GM, Langton, SD, and MacNicoll, AD (1993). The development of a blood clotting response test for discriminating between difenacoum resistant and susceptible Norway rats (Rattus norvegicus Berk.). Comparative Biochemistry and Physiology Part C: Comparative Pharmacology 104, 29–36.
| The development of a blood clotting response test for discriminating between difenacoum resistant and susceptible Norway rats (Rattus norvegicus Berk.).Crossref | GoogleScholarGoogle Scholar |
Fisher P (2005) Review of house mouse (Mus musculus) susceptibility to anticoagulant poisons. Department of Conservation Science Internal Series 198. (Department of Conservation: Wellington, New Zealand)
Foster, S, King, C, Patty, B, and Miller, S (2011). Tree-climbing capabilities of Norway and ship rats. New Zealand Journal of Zoology 38, 285–296.
| Tree-climbing capabilities of Norway and ship rats.Crossref | GoogleScholarGoogle Scholar |
Fu, X, Booth, SL, and Smith, DE (2007). Vitamin K contents of rodent diets: a review. Journal of the American Association for Laboratory Animal Science 46, 8–12.
| 17877321PubMed |
Garg, N, and Singla, N (2015). Blood clotting response test for detecting resistance to second generation anticoagulant bromadialone in house rat (Rattus rattus). Indian Journal of Animal Research 49, 607–611.
Garg, N, Singla, N, Jindal, V, and Babbar, BK (2017). Studies on bromadiolone resistance in Rattus rattus populations from Punjab, India. Pesticide Biochemistry and Physiology 139, 24–31.
| Studies on bromadiolone resistance in Rattus rattus populations from Punjab, India.Crossref | GoogleScholarGoogle Scholar | 28595918PubMed |
Goulois, J, Chapeuzet, A, Lambert, V, Chatron, N, Tchartanov, L, Legros, L, Benoît, E, and Lattard, V (2016). Evidence of a target resistance to antivitamin K rodenticides in the roof rat Rattus rattus: identification and characterisation of a novel Y25F mutation in the Vkorc1 gene. Pest Management Science 72, 544–550.
| Evidence of a target resistance to antivitamin K rodenticides in the roof rat Rattus rattus: identification and characterisation of a novel Y25F mutation in the Vkorc1 gene.Crossref | GoogleScholarGoogle Scholar | 25847836PubMed |
Greaves JH, Cullen-Ayers PB (1988) Genetics of difenacoum resistance in the rat. In ‘Current advances in Vitamin K Research’. (Ed. JW Suttie) pp. 389–397. (Elsevier: New York, NY, USA)
Greaves, JH, and Rennison, BD (1973). Population aspects of warfarin resistance in the brown rat, Rattus norvegicus. Mammal Review 3, 27–29.
| Population aspects of warfarin resistance in the brown rat, Rattus norvegicus.Crossref | GoogleScholarGoogle Scholar |
Hadler, MR, and Shadbolt, RS (1975). Novel 4-hydroxycoumarin anticoagulants active against resistant rats. Nature 253, 275–277.
| Novel 4-hydroxycoumarin anticoagulants active against resistant rats.Crossref | GoogleScholarGoogle Scholar | 1113846PubMed |
Hadler, MR, Redfern, R, and Rowe, FP (1975). Laboratory evaluation of difenacoum as a rodenticide. Journal of Hygiene 74, 441–448.
| Laboratory evaluation of difenacoum as a rodenticide.Crossref | GoogleScholarGoogle Scholar | 1056964PubMed |
Harper, GA, and Rutherford, M (2016). Home range and population density of black rats (Rattus rattus) on a seabird island: a case for a marine subsidised effect? New Zealand Journal of Ecology 40, 219–228.
| Home range and population density of black rats (Rattus rattus) on a seabird island: a case for a marine subsidised effect?Crossref | GoogleScholarGoogle Scholar |
Innes JG, Williams D (1991) The impact of aerial 1080 poisoning on ship rat populations at Mapara and Kaharoa. Forest Research institute Contract Report FWE 91/30. (Forest Research Institute: Rotorua)
Innes, J, Warburton, B, Williams, D, Speed, H, and Bradfield, P (1995). Large-scale poisoning of ship rats (Rattus rattus) in Indigenous Forests of the North-Island, New Zealand. New Zealand Journal of Ecology 19, 5–17.
Ishizuka, M, Okajima, F, Tanikawa, T, Min, H, Tanaka, KD, Sakamoto, KQ, and Fujita, S (2007). Elevated warfarin metabolism in warfarin-resistant roof rats (Rattus rattus) in Tokyo. Drug Metabolism and Disposition 35, 62–66.
| Elevated warfarin metabolism in warfarin-resistant roof rats (Rattus rattus) in Tokyo.Crossref | GoogleScholarGoogle Scholar | 17012541PubMed |
Jackson WB, Ashton AD (1979) Present distribution of anticoagulant resistance in the United States. In ‘Vitamin K Metabolism and Vitamin K-dependent Protiens’. (Ed. JW Suttie) pp. 392–397. (University Park Press: Baltimore, MD, USA)
Kaukeinen D, Rampaud MA (1986) A review of brodifacoum efficacy in the U.S. and worldwide. In ‘Proceedings of the 12th Vertebrate Pest Conference, 4–6 March 1986, San Diego, CA, USA’. (Ed. TP Salmon) pp. 9–85. (University of California: Davis, CA, USA)
Lefebvre, S, Rannou, B, Besse, S, Benoit, E, and Lattard, V (2016). Origin of the gender differences of the natural resistance to antivitamin K anticoagulants in rats. Toxicology 344, 34–41.
| Origin of the gender differences of the natural resistance to antivitamin K anticoagulants in rats.Crossref | GoogleScholarGoogle Scholar | 26860702PubMed |
Li, T, Chang, CY, Jin, DY, Lin, PJ, Khvorova, A, and Stafford, DW (2004). Identification of the gene for vitamin K epoxide reductase. Nature 427, 541–544.
| Identification of the gene for vitamin K epoxide reductase.Crossref | GoogleScholarGoogle Scholar | 14765195PubMed |
Liang, L (2005). The resistance of Rattus flavipectus and R. norvegicus to Anticoagulant Rodenticide in Zhanjiang proper. Chinese Journal of Vector Biology and Control 16, 21–22.
Lund, M (1969). Resistance to anticoagulants in Denmark. Schriftenreihe des Vereins für Wasser-, Boden- und Lufthygiene 32, 27–38.
| 5293929PubMed |
Lund M (1984) Resistance to the second generation anticoagulant rodenticides. In ‘Proceedings of the 11th Vertebrate Pest Conference, 6–8 March 1984, Sacramento, CA, USA’. (Ed. DO Clark) (University of California: David, CA, USA)
Markussen, M, Heiberg, A, Fredholm, M, and Kristensen, M (2008). Differential expression of cytochrome P450 genes between bromadiolone-resistant and anticoagulant-susceptible Norway rats: a possible role for pharmacokinetics in bromadiolone resistance. Pest Management Science 64, 239–248.
| Differential expression of cytochrome P450 genes between bromadiolone-resistant and anticoagulant-susceptible Norway rats: a possible role for pharmacokinetics in bromadiolone resistance.Crossref | GoogleScholarGoogle Scholar | 18080289PubMed |
Marsh, RE (1977)). Bromadiolone, a new anticoagulant rodenticide. EPPO Bulletin 7, 495–502.
Mathur, RP, and Prakash, I (1981). Evaluation of brodifacoum against T. indica, M. hurrianae and R. rattus. Journal of Hygiene 87, 179–184.
| Evaluation of brodifacoum against T. indica, M. hurrianae and R. rattus.Crossref | GoogleScholarGoogle Scholar | 7288172PubMed |
Moussa BR (2005) Biological and toxicology studies on some rodents. Masters thesis, Zagazig University, Zagazig, Egypt. Available at https://bu.edu.eg/portal/uploads/discussed_thesis/finalabsrtract/10812645.pdf
Myllymäki, A (1995). Anticoagulant resistance in Europe: appraisal of the data from the 1992 EPPO Questionnaire. Pesticide Science 43, 69–72.
| Anticoagulant resistance in Europe: appraisal of the data from the 1992 EPPO Questionnaire.Crossref | GoogleScholarGoogle Scholar |
O’Connor CE, Booth LH (2001) Palatability of rodent baits to wild house mice. In ‘Science for Conservation, Vol. 184’, p. 11. (Department of Conservation: Wellington, New Zealand)
Pelz HJ, Prescott C (2015) Resistance to anticoagulant rodenticides. In ‘Rodent Pests and their Control’. (Ed. AP Buckle, RH Smith) pp. 195–201. (CAB International: Wallingford, Oxfordshire, UK)
Pelz, H-J, Rost, S, Hünerberg, M, Fregin, A, Heiberg, A-C, Baert, K, Macnicoll, AD, Prescott, CV, Walker, A-S, Oldenburg, J, and Müller, CR (2005). The genetic basis of resistance to anticoagulants in rodents. Genetics 170, 1839–1847.
| The genetic basis of resistance to anticoagulants in rodents.Crossref | GoogleScholarGoogle Scholar | 15879509PubMed |
Prescott, CV, and Buckle, AP (2000). Blood-clotting response tests for resistance to diphacinone and chlorophacinone in the Norway rat (Rattus Norvegicus Berk.). Crop Protection 19, 291–296.
| Blood-clotting response tests for resistance to diphacinone and chlorophacinone in the Norway rat (Rattus Norvegicus Berk.).Crossref | GoogleScholarGoogle Scholar |
Prescott, CV, Buckle, AP, Hussain, I, and Endepols, S (2007). A standardised BCR resistance test for all anticoagulant rodenticides. International Journal of Pest Management 53, 265–272.
| A standardised BCR resistance test for all anticoagulant rodenticides.Crossref | GoogleScholarGoogle Scholar |
Pryde, M, Dilks, P, and Fraser, I (2005). The home range of ship rats (Rattus rattus) in beech forest in the Eglinton Valley, Fiordland, New Zealand: a pilot study. New Zealand Journal of Zoology 32, 139–142.
| The home range of ship rats (Rattus rattus) in beech forest in the Eglinton Valley, Fiordland, New Zealand: a pilot study.Crossref | GoogleScholarGoogle Scholar |
Quy, RJ, Cowan, DP, Prescott, CV, Gill, JE, Kerins, GM, Dunsford, G, Jones, A, and Macnicoll, AD (1995). Control of a population of Norway rats resistant to anticoagulant rodenticides. Pesticide Science 45, 247–256.
| Control of a population of Norway rats resistant to anticoagulant rodenticides.Crossref | GoogleScholarGoogle Scholar |
Redfern, R, and Gill, JE (1978). The development and use of a test to identify resistance to the anticoagulant difenacoum in the Norway rat (Rattus norvegicus). Journal of Hygiene 81, 427–431.
| The development and use of a test to identify resistance to the anticoagulant difenacoum in the Norway rat (Rattus norvegicus).Crossref | GoogleScholarGoogle Scholar | 731023PubMed |
Redfern, R, Gill, JE,, and Hadler, MR (1976). Laboratory evaluation of WBA 8119 as a rodenticide for use against warfarin-resistant and non-resistant rats and mice. Journal of Hygiene 77, 419–426.
| 1069820PubMed |
Rost, S, Fregin, A, Ivaskevicius, V, Conzelmann, E, Hortnagel, K, and Pelz, HJ (2004). Mutations in VKORC1 cause warfarin resistance and multiple coagulation factor deficiency type 2. Nature 427, 537–541.
| Mutations in VKORC1 cause warfarin resistance and multiple coagulation factor deficiency type 2.Crossref | GoogleScholarGoogle Scholar | 14765194PubMed |
Rost, S, Pelz, H-J, Menzel, S, MacNicoll, VL, León, V, Song, K-J, Jäkel, T, Oldenburg, J, and Müller, C (2009). Novel mutations in the VKORC1 gene of wild rats and mice – a response to 50 years of selection pressure buy warfarin. BMC Genetics 10, 4.
| Novel mutations in the VKORC1 gene of wild rats and mice – a response to 50 years of selection pressure buy warfarin.Crossref | GoogleScholarGoogle Scholar | 19200363PubMed |
Rymer DJ (2017) Anticoagulant resistance in Norway rats conferred by VKORC1 mutations in South-East England. PhD Thesis, University of Reading, UK.
Spiegelhalter D, Thomas A, Best N, Lunn D (2014) ‘OpenBUGS User Manual, Version 3.2.3.’ (MRC Biostatistics Unit: Cambridge, UK)
Wheeler, R, Priddel, D, O’Dwyer, T, Carlile, N, Portelli, D, and Wilkinson, I (2019). Evaluating the susceptibility of invasive black rats (Rattus rattus) and house mice (Mus musculus) to brodifacoum as a prelude to rodent eradication on Lord Howe Island. Biological Invasions 21, 833–845.
| Evaluating the susceptibility of invasive black rats (Rattus rattus) and house mice (Mus musculus) to brodifacoum as a prelude to rodent eradication on Lord Howe Island.Crossref | GoogleScholarGoogle Scholar |
Wodzicki KA (1978). Review of existing control methods. In ‘The Ecology and Control of Rodents in New Zealand Nature Reserves’. (Eds PR Dingwall, IAE Atkinson, C Hay) pp. 195–205. (Department of Lands and Survey: Wellington, New Zealand)