

Experimentally testing the response of feral cats and their prey to poison baiting
Tim S. Doherty A B F , Michelle L. Hall C D E , Ben Parkhurst C and Vanessa Westcott C
A B F , Michelle L. Hall C D E , Ben Parkhurst C and Vanessa Westcott C
A School of Life and Environmental Sciences, The University of Sydney, Camperdown, NSW 2006, Australia.
B Centre for Integrative Ecology, School of Life and Environmental Sciences, Deakin University, 221 Burwood Highway, Burwood, Vic. 3125, Australia.
C Bush Heritage Australia, 1/395 Collins Street, Melbourne, Vic. 3000, Australia.
D School of Biological Sciences, The University of Western Australia, 35 Stirling Highway, Perth, WA 6009, Australia.
E School of BioSciences, The University of Melbourne, Melbourne, Vic. 3010, Australia.
F Corresponding author. Email: tim.doherty@sydney.edu.au
Wildlife Research 49(2) 137-146 https://doi.org/10.1071/WR21008
Submitted: 13 January 2021 Accepted: 16 June 2021 Published: 20 August 2021
Journal Compilation © CSIRO 2022 Open Access CC BY
Abstract
Context: Feral cats, Felis catus, have caused the decline and extinction of many species worldwide, particularly on islands and in Australia where native species are generally naïve to the threat of this introduced predator. Effectively reducing cat populations to protect wildlife is challenging because cats have a cryptic nature, high reproductive rate and strong reinvasion ability.
Aims: We experimentally tested the response of feral cats and their native prey to an Eradicat® poison baiting program at a conservation reserve.
Methods: Baits were distributed by hand along roads and tracks every 50 m (~10 baits km−2). We used camera traps to monitor the response of cats to baiting using a repeated before–after, control–impact design over 6 years. We also measured introduced rabbit, Oryctolagus cuniculus, activity by using sand pads and small mammal and reptile captures by using pitfall trapping.
Key results: Dynamic occupancy modelling showed only modest effects of baiting on cats in 2 of 6 years, with occupancy in the baited area decreasing from 54% to 19% in 2014 (–35%) and from 89% to 63% in 2017 (–26%). Baiting effectiveness was not related to antecedent rainfall or prey availability. Bait availability was reduced by non-target interference; 73% of 41 monitored baits were removed by non-target species. We found no evidence for persistent changes in small mammal or reptile capture rates in the baited area relative to the unbaited area over the life of the project.
Conclusions: Relatively low baiting density and non-target interference with baits are likely to have reduced baiting efficacy. Further testing and refinement of ground baiting is needed, including trialling higher baiting densities and/or frequencies.
Implications: We highlight key areas for future research that should benefit feral cat management not only in Australia, but also on the many islands worldwide where cats threaten native wildlife.
Keywords: cat baiting, dynamic occupancy model, impact evaluation, invasive predator, lethal control, pest control.
Introduction
Invasive predators are a major driver of global biodiversity loss, having contributed to more than 50% of bird, mammal and reptile extinctions worldwide (Doherty et al. 2016). Their impacts have been greatest on islands, where prey species are typically naïve to the threat of introduced predators (Salo et al. 2007; Medina et al. 2011). Feral cats, Felis catus, are one of the most damaging species because humans have spread them across the world, they are highly adaptable to varying environmental conditions (Bengsen et al. 2016), and they prey on a range of birds, mammals, reptiles, amphibians and invertebrates (Bonnaud et al. 2011). We use the term ‘feral cats’ to refer to animals that live in the wild and have no direct dependence on humans. Reducing the impacts of cats on native wildlife is a key concern of conservation practitioners and scientists globally (Nogales et al. 2013; Shionosaki et al. 2015; Loss and Marra 2017; Doherty et al. 2017).
Management of feral cats has typically focussed on lethal control, including trapping, shooting, and poison baiting. Each of these methods rely on removing individual predators from a system to reduce or eliminate predation pressure on prey species. Cats have a high reproductive rate and ability to reinvade, so lethal control must be intensive and sustained so as to effectively reduce cat population densities (Leo et al. 2018; Lohr and Algar 2020). Demographic studies have indicated that, on average, more than 57% of a cat population must be removed annually to reduce population densities (Hone et al. 2010). However, achieving this in practice has been challenging because of the cryptic nature of cats and their aversion to entering traps and consuming baits (Fisher et al. 2015). For instance, the huntability of cats (number of cats shot at as a percentage of those sighted) on Marion Island ranged from 25% to 44% over 4 years (Bloomer and Bester 1992). Low-intensity trapping and removal of cats in Tasmania, Australia, actually caused an increase in cat activity and relative abundance at removal sites, possibly owing to immigration by neighbouring cats into vacated territories (Lazenby et al. 2014). In the Flinders Ranges, South Australia, the number of cat detections on cameras did not change after 40 cats were removed through trapping over 2 months (Stobo-Wilson et al. 2020). On Rota Island, spotlight hunting of cats caused a modest knockdown within the first 18 months, but the population stabilised over the next 11 months (Leo et al. 2018). These examples illustrate that effective cat control can be very time and labour intensive, meaning that resource and financial constraints can also hinder success.
In Australia, where cats have contributed to the extinction of more than 20 native mammal species and threaten many other birds, mammals and reptiles (Woinarski et al. 2015, 2019), there are two specially designed poison baits that can effectively reduce cat populations, dependent on environmental conditions (e.g. Johnston et al. 2014; Comer et al. 2018; Lohr and Algar 2020). Eradicat® and Curiosity® are small sausage-style baits comprising kangaroo meat, chicken fat, and digest and flavour enhancers (Algar et al. 2007; Johnston et al. 2013). Eradicat® is registered for use in parts of Western Australia and contains 4.5 mg of 1080 poison (sodium fluoroacetate) injected into the bait (Algar et al. 2007). Curiosity® is designed for use in southern and eastern Australia and contains a hard capsule of para-aminopropiophenone (PAPP) poison (Johnston et al. 2013, 2014). Eradicat® is usually deployed aerially at a rate of 50 baits km−2, which can reduce cat populations, although effectiveness varies among years (Algar et al. 2007, 2011, 2013; Richards and Algar 2010; Comer et al. 2018; Lohr and Algar 2020; Palmer et al. 2021). The baits are also readily consumed by dingoes, Canis dingo, and introduced red foxes, Vulpes vulpes, and thus can also reduce their population densities (Richards and Algar 2010; Berry et al. 2013; Wysong et al. 2020b).
In addition to aerial baiting, baits can also be distributed along tracks and roads (‘ground baiting’), although this approach has received less attention (but see Doherty and Algar 2015; Burrows et al. 2018; Lohr and Algar 2020; Fancourt et al. 2021). Ground baiting may be a more accessible option for smaller landholders that do not have adequate resources for aerial operations (e.g. some non-government organisations and private landholders). However, because ground baiting relies on roads and tracks for bait distribution, potential baiting densities are reduced, which may limit effectiveness. Placing baits along tracks may increase encounter rates by animals that preferentially move along tracks (Geyle et al. 2020; Wysong et al. 2020a), but by the same token, the baits will be biased away from animals that rarely use tracks (Fancourt et al. 2021).
In the present study, we experimentally tested the impacts of annual Eradicat® ground baiting on feral cat occupancy and activity over 6 years (2013–19, excluding 2015) at a conservation reserve in Western Australia. We monitored impacts of baiting on both feral cats and their prey, comparing the baited area with an unbaited control area (BACI experimental design). This project began under an experimental permit before Eradicat® was registered by the national regulator (Australian Pesticides and Veterinary Medicines Authority, PER14102). The baiting involved an annual application of baits at a density of ~10 per km2 (one bait every 50 m), which is 20% of that used in aerial operations. This is the maximum density that we could use, and we were limited to applying baits once per year under the experimental permit. Results from the first 2 years of this project showed that baiting reduced cat activity in 2014, but not 2013 (Doherty and Algar 2015). We re-analyse that data here, along with additional data, to address the following research questions for 2013–19:
-
Which species consume Eradicat® baits and how often?
-
Does annual ground baiting with Eradicat® reduce cat occupancy and activity?
-
Is baiting effectiveness related to rainfall or prey availability?
-
Have capture rates of small mammals and reptiles in the baited area changed over time relative to the unbaited area?
Materials and methods
Study site and design
This study was conducted at Charles Darwin Reserve, a ~68 000 ha property managed for conservation by Bush Heritage Australia (–29.65, 116.97; Fig. 1). The climate is semiarid Mediterranean and mean annual rainfall is ~300 mm (Wanarra weather station; Bureau of Meteorology 2020). Vegetation primarily consists of Acacia shrublands and Eucalyptus woodlands, with smaller areas of salt lakes, and granite outcrops and breakaways. The property is surrounded by Unallocated Crown Land to the south, pastoral leases to the west and north, and the Mount Gibson Wildlife Sanctuary to the east. The reserve does not have predator or herbivore exclusion fences. Cats are common in the study area and dingoes less so, whereas foxes are uncommon (Doherty 2015b).
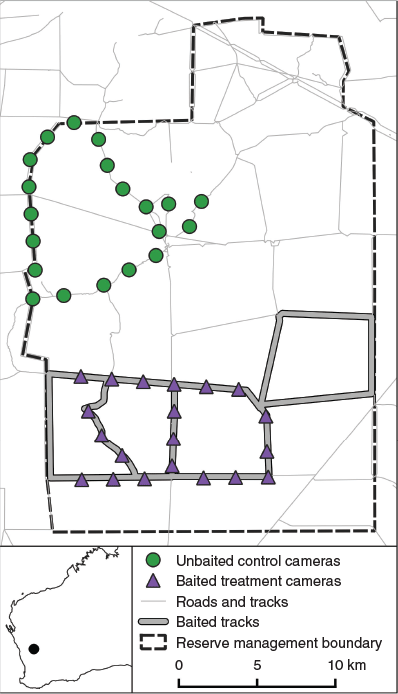
|
The baited treatment area (~150 km2) was located in the southern half of the reserve and the unbaited experimental control area (~100 km2) was located in the northern half (Fig. 1). Toxic Eradicat® baits were distributed in the baited area once annually from 2013 to 2019. Baits were laid by hand along alternating sides of vehicle tracks at a rate of one bait every 50 m, with ~1500 baits being laid each year, resulting in a density of ~10 baits km−2. Baits were not replaced once they were removed. Baiting was conducted once in each of September 2013, May 2014, June 2015, and May 2016–19. However, we do not present the 2015 data because of multiple issues with camera trap operation that year that compromised data collection. As detailed below, our analyses include using dynamic occupancy models and generalised linear mixed models (GLMMs) to assess cat responses to baiting (Question 2), general linear models to assess drivers of baiting effectiveness (Question 3), and GLMMs to model changes in small mammal and reptile capture rates (Question 4). Data collection in 2013–14 was approved by the Edith Cowan University Animal Ethics Committee (8501), and in 2015–19 by the Department of Parks and Wildlife (08-000407-1).
Camera set-up and bait uptake trials
Cats were monitored before and after baiting each year using motion-sensing cameras. Twenty cameras each separated by ≥2 km were deployed in each of the baited and unbaited treatments. At this spacing, the cameras are unlikely to have been spatially independent for the purposes of occupancy modelling, thus the occupancy results should be interpreted as probability of site use, rather than true occupancy (MacKenzie et al. 2018). Most of the unbaited cameras (75%) were >9.50 km from the baited area, with the minimum distance being 5.50 km. We considered this distance large enough for the two treatments to be considered independent for cats, given that home range studies from similar Australian environments recorded mean home ranges of 2.48–22.10 km2 (Jones and Coman 1982; Edwards et al. 2001; Molsher et al. 2005; Hilmer 2010; Bengsen et al. 2012), which corresponds to a home range diameter of 1.80–5.30 km (if assumed to be a circle). Although dingoes were recorded on our cameras, we do not present the data here because the treatments were too close together to be independent for dingoes, given their much larger home ranges (Harden 1985; Robley et al. 2010; Newsome et al. 2013; Allen et al. 2014).
The cameras used in 2013–15 were a mixture of Moultrie i60 and Scoutguard 560PV units, whereas the cameras used in 2016–19 were Reconyx HC600 Hyperfire. Equal numbers of the two camera models were deployed in each treatment in 2013–15 and assignment to locations was randomised, to reduce any bias. Cameras were fixed to steel posts ~30 cm above the ground and next to vehicle tracks. Cameras were programmed to take three photographs each time the sensor was triggered, with a minimum delay of 1 min between triggers. To measure bait uptake (Question 1), each of the 20 cameras in the baited area had a bait placed in front of it during the baiting period in 2015–19. Memory cards were collected from cameras 1–3 weeks after the baiting and photos were inspected to assess whether baits were taken, which species were responsible, and how long after bait placement removal took place.
Feral cat occupancy and activity (Question 2)
Each pre- and post-baiting monitoring session lasted for 4–6 weeks (Table S1, available as Supplementary material to this paper), although some individual cameras stopped working prematurely because of battery failure or memory cards reaching capacity. Post-baiting monitoring began 1–4 weeks after baiting, except in 2016 when it began 2 months after baiting. In 2013–15, half of the cameras had a scent lure and the other half an audio lure, which were swapped between cameras halfway through each monitoring session. Scent lures were a fresh chicken wing encased in a PVC tube pegged to the ground, with a fresh chicken wing provided when lures were swapped. The audio lure was a small electronic device that emitted the sound of a bird tweeting (Lucky Duck, WI, USA). Only scent lures (chicken wings) were used at all cameras in 2016–19.
Photos were manually inspected, and the presence of animal species was recorded, along with the site number, date and time. We created dataframes relating to sampling effort and detection histories using the camtrapR package in R (Niedballa et al. 2016; R Core Team 2019). The sampling effort dataframes recorded when cameras stopped functioning before the end of survey periods. Two cameras were also stolen in 2013. Detection histories represented the presence or absence of a species at each camera during each successive 3-day period throughout a survey. For instance, a 31-day survey would have 10 3-day sampling occasions, with the extra day excluded. We chose a 3-day, rather than daily, sampling period to improve model convergence.
We used dynamic occupancy models in the unmarked package in R (Fiske and Chandler 2011) to assess changes in cat occupancy in response to baiting each year. Dynamic occupancy models use data from multiple primary periods (pre-baiting and post-baiting here) that comprise multiple secondary periods (3-day blocks in this case) and do not assume that the system is closed between the primary periods. This approach enables the estimation of detectability, initial site occupancy (first primary period), and colonisation and extinction probabilities, which represent changes in site occupancy between primary periods, i.e. from before to after baiting.
We analysed each year separately and used a multi-step approach by first conducting all subset model selection on the detection component and then fitting a specific set of hypothesis-based models for the occupancy and extinction components. First, we determined whether detection probabilities were influenced by vegetation composition at each site by using model selection. We calculated the proportion of the following five vegetation types within a 500-m radius of each camera: young shrublands (<20 years since fire), old shrublands (≥20 years since fire), woodlands, salt lakes and granite outcrops. We excluded young shrublands from the modelling because they were negatively correlated with old shrublands (Pearson’s r = –0.58) and woodlands (–0.43). We fitted a global detection model containing main effects for each of the four vegetation variables, and held occupancy, colonisation and extinction probabilities constant. For 2013 and 2014, we also included camera model (Moultrie or Scoutguard) as a potential predictor of detectability. We used the dredge function in the MuMIn package (Bartoń 2019) to fit all possible model combinations and ranked the models by using Akaike’s information criterion corrected for small sample size (AICc). For the subsequent modelling of occupancy and extinction probabilities, we used the detection variables from the model with the highest weight, which in some cases was the null model (Table S2).
To determine whether baiting affected cat occupancy, we fitted and ranked a series of models that included treatment (baited/unbaited) as a predictor of initial site occupancy, extinction probability, or both. These models also included the detection covariates as per Table S2. We compared these three models to a null model that included only the detection covariates (if applicable). Because the experiment used a before–after, control–impact (BACI) design, an effect of baiting would be supported if there was an effect of treatment on extinction probability. Specifically, we would expect extinction probability from pre- to post-baiting to be higher in the baited than in the unbaited area. We discuss the results from any models with a ΔAICc ≤ 2. We used parametric bootstrapping with 1000 simulations to derive pre- and post-baiting occupancy estimates for each treatment, which we present as means with 95% confidence intervals.
We also used GLMMs with a Gaussian distribution to assess changes in cat activity in response to baiting. The activity index was calculated by dividing the number of independent detections of cats on each camera by the number of nights the camera was active and multiplying this by 100 (i.e. number of detections per 100 trap-nights). Independent detections were photographs on the same camera that were at least 60 min apart. The models included fixed effects of Time (pre-/post-baiting), Treatment (baited/unbaited), and Time × Treatment. A significant interaction would support an impact of baiting on cat activity. Models also included a random effect of Site to account for repeat sampling. We fitted a separate model for each year and present parameter estimates and 95% confidence intervals. To provide further context to the occupancy and activity results, we also present in the Supplementary material plots of naïve occupancy, which represents the proportion of sites within each treatment where each species was detected, disregarding varying sampling effort.
Drivers and outcomes of baiting effectiveness (Questions 3 and 4)
We used data on rainfall, small mammal capture rates, and introduced rabbit, Oryctolagus cuniculus, activity to assess whether baiting effectiveness was related to the availability of potential prey (Question 3). We calculated total rainfall for the 6 months (Rain_6m) and 12 months (Rain_12m) before each baiting event, using data from a rain gauge at the reserve. We used pitfall trapping data to estimate capture rates of small mammals (Mammal_CR: number of individuals captured per trap-night) in the spring (September–November) before each baiting event (n = 8–16 sites; Table S3). The small mammal species captured were Notomys mitchellii, Pseudomys hermanns burgensis, Mus musculus and Sminthopsis spp. (see Doherty et al. 2015 for further details). As per previous studies (e.g. Coates 2008; Weston et al. 2009), we used sand pads to monitor rabbits and calculated an index of rabbit activity for both the spring (Rab_spr) and winter (Rab_win) before each baiting event (i.e. in the previous year). The index was calculated as the proportion of days rabbits were detected on each sand pad (n = 69), averaged across all sand pads for each season. Sand pad data were not available for winter 2013 and spring 2012 and 2013. To further assess the relationship between prey availability and baiting effectiveness, we calculated a ratio (PP_ratio) by dividing Mammal_CR by mean pre-baiting cat activity across all cameras for each year, following Christensen et al. (2013). Pitfall trapping and sand pad monitoring sites were spread across baited and unbaited areas (Fig. S1), although we pooled all data for analysis to assess broadscale inter-annual variation in prey availability, and because there were few differences between treatments across years (see Results). Full details of the field sampling are provided in the Supplementary material.
To create a measure of baiting effectiveness for each year, we subtracted the mean difference in activity or occupancy between baited and unbaited sites pre-baiting, from the mean difference after baiting, i.e. (AfterUnbaited – AfterBaited) – (BeforeUnbaited – BeforeBaited) (Christie et al. 2019). For the occupancy metric, we used the estimates from the top-ranked model for each year. Using this approach, values of 0 indicate that the difference between treatments is equal for before and after baiting. Positive values indicate a greater difference in favour of unbaited sites, i.e. occupancy decreased more in baited than unbaited sites and/or unbaited sites increased more than baited sites. Negative values indicate a greater difference in favour of baited sites, i.e. occupancy in unbaited sites decreased more than in baited sites and/or baited sites increased more than unbaited sites. As such, more positive values are indicative of a greater impact of baiting.
To test the relationship between baiting effectiveness and environmental variables, we fitted general linear models with either the occupancy or activity baiting effectiveness metric as the response variable and Rain_6m, Rain_12m, Mammal_CR, PP_ratio, Rab_spr or Rab_win as the predictor variable. We included only one predictor variable per model because of the small sample size (n = 4–6 years). We present model parameter estimates, 95% confidence intervals and plots of the data.
We also used GLMMs assuming a Gaussian distribution to assess whether capture rates of small mammals and reptiles have changed over time in baited and unbaited areas (Question 4). Data were derived from the pitfall trapping as described above and in the Supplementary material. The response variable was small mammal or reptile capture rate for each spring 2012–18 (excluding 2014, when sampling was not undertaken). Year 2012 represents a baseline from before baiting began. We included fixed effects of Year, Treatment, and Year × Treatment, and a random effect of Site to account for repeat sampling.
Results
Bait uptake
Of the 100 baits laid in front of cameras in 2015–19, 30 could not be seen on camera because of poor placement or camera malfunction, and a further 29 baits disappeared without the event being recorded on camera. Of the remaining 41 baits, we recorded an animal interaction for 34 baits. Records of cats included one bait consumed 9 h after being laid, another bait inspected but not removed 7 days after being laid, and another bait that a cat walked past without seeming to inspect it, 3 days after it was laid. Twenty baits were either eaten or taken away by emus, Dromaius novaehollandiae (49% of visible baits), seven by ravens, Corvus coronoides (17%), and one each by a fox, hopping mouse, Notomys mitchellii, and grey currawong, Strepera versicolor. Emus removed baits within 0–9 days of baits being laid (mean = 4.05). In one case, the bait was removed 5 h after being laid. Ravens removed baits within 1–22 days (mean = 6.43) and the fox, hopping mouse and currawong removed baits 3 days, 2 days and 5 min after being laid, respectively. Twenty-six minutes after the fox consumed a bait, a fox also inspected but did not remove a bait at a neighbouring camera. One dingo was also recorded walking past a bait without seeming to inspect it. Ten baits remained in place at the end of the trial period.
Occupancy and activity
Over the 6-year study, feral cats were detected at 3–18 treatment cameras and 12–19 control cameras each year (mean = 11.67 and 14.83 of 20 cameras, respectively). There was uncertainty in the impacts of baiting on cat occupancy in most years, with multiple models having ΔAICc values of ≤2 (Table S4, Fig. S2). There was only 1 year with a clear impact of baiting on cat occupancy; in 2017 occupancy decreased in the baited area post-baiting (from 0.89 to 0.63 in the top model), whereas it increased slightly in the unbaited area (from 0.89 to 1.00; Fig. 2). The changes in activity and naïve occupancy also support an impact of baiting in 2017, including a significant Time × Treatment interaction in the mixed model (Figs 2, S3, Table S5).
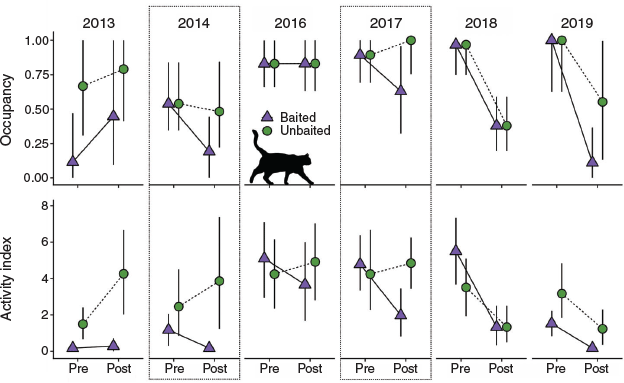
|
The best-supported occupancy models for 2014 and 2019 also indicated greater declines in occupancy in the baited than in the unbaited area (Fig. 2), although the null models were also well supported (Table S4, Fig. S2). Changes in naïve occupancy and the activity index support an impact of baiting in 2014, but not 2019 (Figs 2, S3). The Time × Treatment interaction for activity in 2014 had confidence intervals slightly overlapping zero (–0.48–5.56; Table S5). Estimated occupancy from the top model for 2014 was 0.54 in both treatments pre-baiting and 0.48 in the unbaited area and 0.19 in the baited area post-baiting (Fig. 2). The activity confidence intervals for Time × Treatment in 2013 and 2018 also only slightly overlapped zero (Table S5), but when considered together with the occupancy data, there is little support for an impact of baiting.
Drivers and outcomes of baiting effectiveness
Pre-baiting occupancy, activity and naïve occupancy were similar between baited and unbaited treatments each year from 2014 to 2019 (Figs 2, S3), indicating that there was no cumulative effect of baiting on cat activity or occupancy over time. There was no relationship between baiting effectiveness (based on changes in cat activity/occupancy) and rainfall, small mammal capture rate, rabbit activity, or the predator–prey ratio index (Table S6, Fig. S4). Small mammal capture rates were similar between treatments every year, except in 2015 and 2016 when average capture rates were 2.1- and 1.8-fold higher respectively, at baited than at unbaited sites (Fig. 3, Table S7). Mean capture rates of reptiles did not vary between treatments, but overall capture rates in 2013 and 2016–18 were lower than those in the baseline year of 2012 (Fig. 3, Table S7).
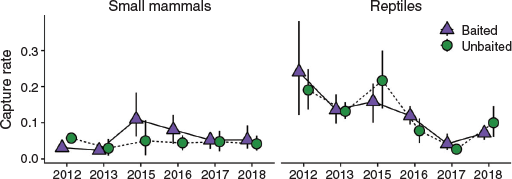
|
Discussion
We experimentally tested the impact of poison baiting on feral cats and potential benefits for their native prey at a conservation reserve over 6 years. Annual ground baiting using Eradicat® at ~10 baits km−2 was mostly ineffective at reducing cat occupancy, with there being only weak treatment effects in two of the 6 years tested (absolute decreases in occupancy of 35% in 2014 and 26% in 2017). Consistent with the limited effect of baiting on feral cats, we found no evidence of benefits for their native prey. There are several factors that could have limited the efficacy of the baiting program, including baiting density, prey availability and non-target uptake of baits.
The baiting density achieved in this project was ~10 baits km−2, which is much lower than the rate of 50 baits km−2 used in aerial operations. The lower baiting density may be insufficient for the majority of cats to detect baits or to encounter a bait when they are likely to eat it. At Matuwa, in central Western Australia, ground baiting at ~2.80 baits km−2 was less effective than aerial baiting at 50 baits km−2 (Lohr and Algar 2020). In the Gibson Desert, Burrows et al. (2003) found that a fresh meat bait designed for cats reduced activity by 75% and 100% at densities of 10 and 22 baits km−2 respectively, during low-rainfall periods, whereas baiting at 11 baits km−2 during a high-rainfall period reduced activity by 25%. Ground application of fresh meat baits at a density of 7.3 km−2 was ineffective at reducing cat densities in Queensland (Fancourt et al. 2021). In South Australia, Moseby and Hill (2011) tested aerial Eradicat® baiting at densities of 10 and 25 baits km−2 and found that cat activity declined in response to baiting in just one of seven trials. When taken together, these studies suggest that a baiting density of 10 baits km−2 is not in itself insufficient, but may be when combined with other factors, as discussed below.
The effective baiting density in our study was likely to be reduced by non-target consumption of baits. Our uptake trials in 2013–14 (reported in Doherty and Algar 2015) showed that corvids, Corvus spp., removed the most baits (12 of 30), followed by cats (6) and varanids, Varanus spp. (2). In 2015–19, 73% of the baits where fate could be determined were removed by non-target species, primarily emus, which removed 49% of visible baits. We recorded only one bait being removed by a cat. Our effective sample size was modest (41 baits) because fate could not be determined for a large number of baits, but if we assume that none of those baits was taken by non-targets (which is unlikely), then an absolute minimum of 30% of baits were removed by non-target species. Of the baits removed by non-targets, 47% were removed within 3 days of being laid and 90% within 7 days. This means that the window of bait availability to cats is very narrow and when combined with the already low propensity of cats to consume baits, the chances of bait uptake are very low.
Many other studies have also recorded high non-target uptake of cat baits, including 22% of baits at Peron Peninsula (Algar et al. 2007), 14–57% at Arid Recovery (Moseby et al. 2011), 71% at Kangaroo Island (Hohnen et al. 2020), and 97% at Dryandra and Tutanning (Friend et al. 2020). Only one of those studies recorded emus removing baits (Algar et al. 2007), although it is not clear what proportion of baits were interfered with. Emus commonly travel long distances along tracks at our study site and elsewhere (T. S. Doherty, pers. obs.), which would provide them with ample opportunity to encounter baits, which were placed at 50-m intervals. However, it is also worth noting that baits for the uptake trials were placed in the open to facilitate camera monitoring, which may have increased their detection and, hence, removal by non-target species. Moseby et al. (2011) found that bait removal by corvids was higher for baits in the open than for those under bushes. In practice, most of the 1500 baits deployed each year were placed underneath or close to shrubs; thus, actual rates of non-target removal may have been lower. We recommend that future work quantify rates of non-target removal for baits that are placed in the open and under shrubs, and on and off tracks.
Prey availability is one of the strongest determinants of the efficacy of cat baiting programs, which itself is primarily driven by rainfall in arid and semiarid Australia (Letnic and Dickman 2010). Using data from three sites in Western Australia, Christensen et al. (2013) showed that the efficacy of aerial cat baiting was predicted by a predator–prey ratio index. Reductions in cat activity were greatest when the amount of prey available per cat was lowest (as inferred from indices; Christensen et al. 2013). Short et al. (1997) also found that bait uptake by cats was inversely related to rabbit density. In contrast, we found no relationship between baiting effectiveness and a similar predator–prey index, nor with measures of rainfall, rabbit activity, or small mammal capture rate. However, this does not mean that prey availability is not important in our study area. It may just be that small sample sizes and low bait availability (and hence, effectiveness) inhibited the detection of any trends. A larger sample size covering a greater range of baiting effectiveness would provide more detailed insights.
Other factors that could have limited the efficacy on ground baiting in this study include the small area that was baited, relative to aerial baiting operations, and the fact that baits were applied only once per year. The size of our baited area (~15 000 ha) was dictated by the need to fit both the treatment and control areas within the one property. Over time, any effects of baiting would have been reversed because cats from surrounding unbaited areas would have reinvaded the baited area (Algar et al. 2013; Lazenby et al. 2014). However, this does not preclude the detection of short-term effects of baiting (within 1–2 months) and we did indeed detect modest effects of baiting in 2 of 6 years. Increasing the frequency at which baits are laid may improve the chances that cats encounter and consume baits. However, only a single annual application of baits was permitted under our experimental permit, and autumn baiting is recommended in the semiarid zone because of the lower prey availability at that time of year. Applying baits more frequently during this time could increase the amount of time that fresh baits are available and thus the likelihood that cats encounter baits when they are hungry or are using tracks (Algar et al. 2007).
A strength of this study was the inclusion of an unbaited reference area for comparison, allowing treatment effects to be separated from seasonal effects that could otherwise bias conclusions. For instance, if we did not survey an unbaited control area, we could have concluded that baiting was effective in 2018 and 2019, even though decreases in activity and occupancy were similar between baited and unbaited areas. We recommend that future work, where feasible, include an unbaited reference area to maximise inferential strength (Christie et al. 2019). This is especially important where control techniques are still being developed, or widespread and consistent effectiveness has not yet been demonstrated. However, where inclusion of an unbaited reference area is not feasible, GPS/VHF tracking of target animals to record survival post-baiting can provide complementary information about baiting efficacy (e.g. Palmer et al. 2021).
Controlling predators is only a means to an end and the ultimate aim of any invasive predator control program should be to produce positive outcomes for the target asset, such as increased species richness, abundance, or threatened species survival. Small mammals and reptiles, along with rabbits, are the major components of cat diets in our study area (Doherty 2015a). We found no consistent differences in capture rates of small mammals and reptiles between baited and unbaited areas over the life of this project. Small mammal captures were higher in baited than in unbaited areas in 2015 and 2016, but because we were unable to measure the impact of baiting on cats in 2015, it is difficult to contextualise these results. However, the overall lack of differences is not surprising, given that there were only modest effects of baiting on cats in 2 of 6 years (2014 and 2017). For cat control to benefit prey populations, reductions in cat populations must be intense and sustained, although there is little precise information about the minimum level of suppression required (Norbury et al. 2015). The clearest examples are where cats (and foxes) have been eradicated from islands and fenced exclosures, resulting in dramatic improvements in the survival of reintroduced mammals (Legge et al. 2018) and increases in the abundance and richness of resident fauna (Moseby et al. 2009; Roshier et al. 2020). Although there are many studies demonstrating reductions in cat activity or density in response to baiting, there is very little evidence available regarding the outcomes for prey populations (but see Stewart et al. 2019; Comer et al. 2020; Palmer et al. 2021). We recommend that control programs for cats, and other pest species more generally, incorporate clear objectives and monitoring programs for the species they are trying to protect (see also Reddiex and Forsyth 2006). Such monitoring can enable assessment of return-on-investment and adaptive management to refine interventions over time, including detecting and addressing unexpected negative consequences (Ruscoe et al. 2011; Walsh et al. 2012).
Feral cat management is challenging, and the effectiveness of different approaches varies among locations and years. Further work is clearly needed to improve the effectiveness of ground baiting of feral cats. Key areas for future research are investigation of methods to reduce non-target removal of baits, investigation of methods that improve bait detection and consumption by cats, and assessment of different baiting densities and frequencies. Furthermore, since effective baiting may selectively target only a subset of the population (e.g. poor hunters or bait naïve cats; Lohr and Algar 2020), complementing baiting with other control methods such as trapping and shooting may be important to maximise the impacts of control efforts and to reduce the risk of selecting for bait resistance (Allsop et al. 2017; Lohr and Algar 2020). In addition, research on the effects of managing the environment by either reducing the resources supporting cat populations (e.g. rabbit populations; McGregor et al. 2020), or managing fire and grazing to conserve habitat structure that provides refuges for native species (Leahy et al. 2015; Legge et al. 2019), could help identify the most effective approaches for feral cat management. Such work would benefit feral cat management not only in Australia, but also on the many islands worldwide where cats threaten native wildlife.
Data availability
Data and code are available at FigShare https://doi.org/10.6084/m9.figshare.13240637.v1.
Conflicts of interest
The authors declare no conflicts of interest.
Declaration of funding
Financial support for this project was provided by Earthwatch Institute Australia, Bush Heritage Australia and Edith Cowan University. T. S. Doherty was supported by Edith Cowan University, Deakin University and the Australian Research Council (DE200100157) over the life of the project.
Author contributions
T. S. Doherty conceived and designed the research; all authors collected the data; T. S. Doherty analysed the data and wrote the manuscript, with input from all co-authors.
Acknowledgements
We acknowledge the Badimia People as the Traditional Owners of the land on which this research was conducted. We thank Luke Bayley and Will Hansen for conducting the baiting, Nic Dunlop and the Conservation Council of Western Australia for providing the rainfall data and assisting with pitfall trapping, Brian Crute for conducting the sand-pad monitoring, Dave Algar, Matt Appleby, Rob Davis and Jim Radford for advice on initial study design, and the many volunteers who assisted with fieldwork. We thank two reviewers for their comments on a previous version of the manuscript.
References
Algar, D., Angus, G. J., Williams, M. R., and Mellican, A. E. (2007). Influence of bait type, weather and prey abundance on bait uptake by feral cats (Felis catus) on Peron Peninsula, Western Australia. Conservation Science Western Australia 6, 109–149.Algar, D., Hamilton, N., Onus, M., Hilmer, S., Comer, S., Tiller, S., Bell, L., Pinder, J., Adams, E., and Butler, S. (2011). Field trial to compare baiting efficacy of Eradicat® and Curiosity® baits. Department of Environment and Conservation, Perth, WA, Australia.
Algar, D., Onus, M., and Hamilton, N. (2013). Feral cat control as part of Rangelands Restoration at Lorna Glen (Matuwa), Western Australia: the first seven years. Conservation Science Western Australia 8, 367–381.
Allen, B. L., Engeman, R. M., and Leung, L. K. P. (2014). The short-term effects of a routine poisoning campaign on the movements and detectability of a social top-predator. Environmental Science and Pollution Research International 21, 2178–2190.
| The short-term effects of a routine poisoning campaign on the movements and detectability of a social top-predator.Crossref | GoogleScholarGoogle Scholar | 24043505PubMed |
Allsop, S. E., Dundas, S. J., Adams, P. J., Kreplins, T. L., Bateman, P. W., and Fleming, P. A. (2017). Reduced efficacy of baiting programs for invasive species: Some mechanisms and management implications. Pacific Conservation Biology 23, 240–257.
| Reduced efficacy of baiting programs for invasive species: Some mechanisms and management implications.Crossref | GoogleScholarGoogle Scholar |
Bartoń, K. (2019). ‘MuMIn: Multi-Model Inference. R package version 1.43.15.’ Available at https://CRAN.R-project.org/package=MuMIn.
Bengsen, A. J., Butler, J. A., and Masters, P. (2012). Applying home-range and landscape-use data to design effective feral-cat control programs. Wildlife Research 39, 258–265.
| Applying home-range and landscape-use data to design effective feral-cat control programs.Crossref | GoogleScholarGoogle Scholar |
Bengsen, A. J., Algar, D., Ballard, G., Buckmaster, T., Comer, S., Fleming, P. J. S., Friend, J. A., Johnston, M., McGregor, H., Moseby, K., and Zewe, F. (2016). Feral cat home-range size varies predictably with landscape productivity and population density. Journal of Zoology 298, 112–120.
| Feral cat home-range size varies predictably with landscape productivity and population density.Crossref | GoogleScholarGoogle Scholar |
Berry, O., Tatler, J., Hamilton, N., Hilmer, S., Hitchen, Y., and Algar, D. (2013). Slow recruitment in a red-fox population following poison baiting: a non-invasive mark–recapture analysis. Wildlife Research 40, 615–623.
| Slow recruitment in a red-fox population following poison baiting: a non-invasive mark–recapture analysis.Crossref | GoogleScholarGoogle Scholar |
Bloomer, J. P., and Bester, M. N. (1992). Control of feral cats on sub-Antarctic Marion Island, Indian Ocean. Biological Conservation 60, 211–219.
| Control of feral cats on sub-Antarctic Marion Island, Indian Ocean.Crossref | GoogleScholarGoogle Scholar |
Bonnaud, E., Medina, F. M., Vidal, E., Nogales, M., Tershy, B., Donlan, C. J., and Corre, M. (2011). The diet of feral cats on islands: a review and a call for more studies. Biological Invasions 13, 581–603.
| The diet of feral cats on islands: a review and a call for more studies.Crossref | GoogleScholarGoogle Scholar |
Bureau of Meteorology (2020). ‘Climate data online.’ Australian Government Bureau of Meteorology. Available at http://www.bom.gov.au/climate/data/.
Burrows, N. D., Algar, D., Robinson, A. D., Sinagra, J., Ward, B., and Liddelow, G. (2003). Controlling introduced predators in the Gibson Desert of Western Australia. Journal of Arid Environments 55, 691–713.
| Controlling introduced predators in the Gibson Desert of Western Australia.Crossref | GoogleScholarGoogle Scholar |
Burrows, N., Liddelow, G., Thoomes, E., and Ambrose, D. (2018). ‘Aerial and ground baiting, Matuwa 2018.’ Department of Biodiversity, Conservation and Attractions: Perth, WA, Australia.
Christensen, P. E. S., Ward, B. G., and Sims, C. (2013). Predicting bait uptake by feral cats, Felis catus, in semi‐arid environments. Ecological Management & Restoration 14, 47–53.
| Predicting bait uptake by feral cats, Felis catus, in semi‐arid environments.Crossref | GoogleScholarGoogle Scholar |
Christie, A. P., Amano, T., Martin, P. A., Shackelford, G. E., Simmons, B. I., and Sutherland, W. J. (2019). Simple study designs in ecology produce inaccurate estimates of biodiversity responses. Journal of Applied Ecology 56, 2742–2754.
| Simple study designs in ecology produce inaccurate estimates of biodiversity responses.Crossref | GoogleScholarGoogle Scholar |
Coates, T. D. (2008). The effect of fox control on mammal populations in an outer urban conservation reserve. Australian Mammalogy 30, 51–63.
| The effect of fox control on mammal populations in an outer urban conservation reserve.Crossref | GoogleScholarGoogle Scholar |
Comer, S., Speldewinde, P., Tiller, C., Clausen, L., Pinder, J., Cowen, S., and Algar, D. (2018). Evaluating the efficacy of a landscape scale feral cat control program using camera traps and occupancy models. Scientific Reports 8, 5335.
| Evaluating the efficacy of a landscape scale feral cat control program using camera traps and occupancy models.Crossref | GoogleScholarGoogle Scholar | 29593271PubMed |
Comer, S., Clausen, L., Cowen, S., Pinder, J., Thomas, A., Burbidge, A. H., Tiller, C., Algar, D., and Speldewinde, P. (2020). Integrating feral cat (Felis catus) control into landscape-scale introduced predator management to improve conservation prospects for threatened fauna: a case study from the south coast of Western Australia. Wildlife Research 47, 762–778.
| Integrating feral cat (Felis catus) control into landscape-scale introduced predator management to improve conservation prospects for threatened fauna: a case study from the south coast of Western Australia.Crossref | GoogleScholarGoogle Scholar |
Doherty, T. S. (2015a). Dietary overlap between sympatric dingoes and feral cats at a semiarid rangeland site in Western Australia. Australian Mammalogy 37, 219–224.
| Dietary overlap between sympatric dingoes and feral cats at a semiarid rangeland site in Western Australia.Crossref | GoogleScholarGoogle Scholar |
Doherty, T. S. (2015b). Ecology of feral cats Felis catus and their prey in relation to shrubland fire regimes. Ph.D. thesis, Edith Cowan University, Perth, WA, Australia.
Doherty, T. S., and Algar, D. (2015). Response of feral cats to a track-based baiting programme using Eradicat baits. Ecological Management & Restoration 16, 124–130.
| Response of feral cats to a track-based baiting programme using Eradicat baits.Crossref | GoogleScholarGoogle Scholar |
Doherty, T. S., Davis, R. A., Van Etten, E. J. B., Collier, N., and Krawiec, J. (2015). Response of a shrubland mammal and reptile community to a history of landscape-scale wildfire. International Journal of Wildland Fire 24, 534–543.
| Response of a shrubland mammal and reptile community to a history of landscape-scale wildfire.Crossref | GoogleScholarGoogle Scholar |
Doherty, T. S., Glen, A. S., Nimmo, D. G., Ritchie, E. G., and Dickman, C. R. (2016). Invasive predators and global biodiversity loss. Proceedings of the National Academy of Sciences of the United States of America 113, 11261–11265.
| Invasive predators and global biodiversity loss.Crossref | GoogleScholarGoogle Scholar | 27638204PubMed |
Doherty, T. S., Dickman, C. R., Johnson, C. N., Legge, S. M., Ritchie, E. G., and Woinarski, J. C. Z. (2017). Impacts and management of feral cats Felis catus in Australia. Mammal Review 47, 83–97.
| Impacts and management of feral cats Felis catus in Australia.Crossref | GoogleScholarGoogle Scholar |
Edwards, G. P., De Preu, N., Shakeshaft, B. J., Crealy, I. V., and Paltridge, R. M. (2001). Home range and movements of male feral cats (Felis catus) in a semiarid woodland environment in central Australia. Austral Ecology 26, 93–101.
Fancourt, B. A., Augusteyn, J., Cremasco, P., Nolan, B., Richards, S., Speed, J., Wilson, C., and Gentle, M. N. (2021). Measuring, evaluating and improving the effectiveness of invasive predator control programs: feral cat baiting as a case study. Journal of Environmental Management 280, 111691.
| Measuring, evaluating and improving the effectiveness of invasive predator control programs: feral cat baiting as a case study.Crossref | GoogleScholarGoogle Scholar | 33272660PubMed |
Fisher, P., Algar, D., Murphy, E., Johnston, M., and Eason, C. (2015). How does cat behaviour influence the development and implementation of monitoring techniques and lethal control methods for feral cats? Applied Animal Behaviour Science 173, 88–96.
| How does cat behaviour influence the development and implementation of monitoring techniques and lethal control methods for feral cats?Crossref | GoogleScholarGoogle Scholar |
Fiske, I., and Chandler, R. (2011). unmarked: an RPackage for fitting hierarchical models of wildlife occurrence and abundance. Journal of Statistical Software 43, 1–23.
| unmarked: an RPackage for fitting hierarchical models of wildlife occurrence and abundance.Crossref | GoogleScholarGoogle Scholar |
Friend, J. A., Hill, R., Macmahon, B., Bell, L., Button, T., Mosen, C., and Hill, S. (2020). Are red-tailed phascogales (Phascogale calura) at risk from Eradicat® cat baits? Wildlife Research 47, 747–761.
| Are red-tailed phascogales (Phascogale calura) at risk from Eradicat® cat baits?Crossref | GoogleScholarGoogle Scholar |
Geyle, H. M., Stevens, M., Duffy, R., Greenwood, L., Nimmo, D. G., Sandow, D., Thomas, B., White, J., and Ritchie, E. G. (2020). Evaluation of camera placement for detection of free‐ranging carnivores; implications for assessing population changes. Ecological Solutions and Evidence 1, e12018.
| Evaluation of camera placement for detection of free‐ranging carnivores; implications for assessing population changes.Crossref | GoogleScholarGoogle Scholar |
Harden, R. H. (1985). The ecology of the dingo in north-eastern New South Wales I. Movements and home range. Wildlife Research 12, 25–37.
| The ecology of the dingo in north-eastern New South Wales I. Movements and home range.Crossref | GoogleScholarGoogle Scholar |
Hilmer, S. (2010). Ecophysiology of feral cats (Felis catus) in Australia. Ph.D. thesis, Goethe University, Frankfurt, Germany.
Hohnen, R., Murphy, B. P., Legge, S. M., Dickman, C. R., and Woinarski, J. C. Z. (2020). Uptake of ‘Eradicat’ feral cat baits by non-target species on Kangaroo Island. Wildlife Research 47, 547–556.
| Uptake of ‘Eradicat’ feral cat baits by non-target species on Kangaroo Island.Crossref | GoogleScholarGoogle Scholar |
Hone, J., Duncan, R. P., and Forsyth, D. M. (2010). Estimates of maximum annual population growth rates (rm) of mammals and their application in wildlife management. Journal of Applied Ecology 47, 507–514.
| Estimates of maximum annual population growth rates (rm) of mammals and their application in wildlife management.Crossref | GoogleScholarGoogle Scholar |
Johnston, M. J., O’Donoghue, M., Holdsworth, M., Robinson, S., Herrod, A., Eklom, K., Gigliotti, F., Bould, L., and Little, N. (2013). Field assessment of the Curiosity® bait for managing feral cats in the Pilbara. Department of Sustainability and Environment, Melbourne, Vic., Australia.
Johnston, M. J., Bould, L., O’Donoghue, M., Holdsworth, M., Marmion, P., Bilney, R., Reside, A. E., Caldwell, D., Gaborov, R., and Gentles, T. (2014). Field efficacy of the Curiosity® bait for management of a feral cat population at Roxby Downs, South Australia. Department of Environment and Primary Industries, Melbourne, Vic., Australia.
Jones, E., and Coman, B. J. (1982). Ecology of the feral cat, Felis catus (L.), in south-eastern Australia III.* Home ranges and population ecology in semiarid north-west Victoria. Australian Wildlife Research 9, 409–420.
| Ecology of the feral cat, Felis catus (L.), in south-eastern Australia III.* Home ranges and population ecology in semiarid north-west Victoria.Crossref | GoogleScholarGoogle Scholar |
Lazenby, B. T., Mooney, N. J., and Dickman, C. R. (2014). Effects of low-level culling of feral cats in open populations: a case study from the forests of southern Tasmania. Wildlife Research 41, 407–420.
| Effects of low-level culling of feral cats in open populations: a case study from the forests of southern Tasmania.Crossref | GoogleScholarGoogle Scholar |
Leahy, L., Legge, S. M., Tuft, K., McGregor, H. W., Barmuta, L. A., Jones, M. E., and Johnson, C. N. (2015). Amplified predation after fire suppresses rodent populations in Australia’s tropical savannas. Wildlife Research 42, 705–716.
| Amplified predation after fire suppresses rodent populations in Australia’s tropical savannas.Crossref | GoogleScholarGoogle Scholar |
Legge, S. M., Woinarski, J. C. Z., Burbidge, A. A., Palmer, R., Ringma, J., Radford, J. Q., Mitchell, N., Bode, M., Wintle, B., Baseler, M., Bentley, J., Copley, P., Dexter, N., Dickman, C. R., Gillespie, G. R., Hill, B., Johnson, C. N., Latch, P., Letnic, M., Manning, A., McCreless, E. E., Menkhorst, P., Morris, K., Moseby, K., Page, M., Pannell, D., and Tuft, K. (2018). Havens for threatened Australian mammals: the contributions of fenced areas and offshore islands to the protection of mammal species susceptible to introduced predators. Wildlife Research 45, 627–644.
| Havens for threatened Australian mammals: the contributions of fenced areas and offshore islands to the protection of mammal species susceptible to introduced predators.Crossref | GoogleScholarGoogle Scholar |
Legge, S. M., Smith, J. G., James, A., Tuft, K. D., Webb, T., and Woinarski, J. C. Z. (2019). Interactions among threats affect conservation management outcomes: livestock grazing removes the benefits of fire management for small mammals in Australian tropical savannas. Conservation Science and Practice 1, e52.
| Interactions among threats affect conservation management outcomes: livestock grazing removes the benefits of fire management for small mammals in Australian tropical savannas.Crossref | GoogleScholarGoogle Scholar |
Leo, B. T., Anderson, J. J., Ha, J., Phillips, R. B., and Ha, R. R. (2018). Modeling impacts of hunting on control of an insular feral cat population. Pacific Science 72, 57–67.
| Modeling impacts of hunting on control of an insular feral cat population.Crossref | GoogleScholarGoogle Scholar |
Letnic, M., and Dickman, C. R. (2010). Resource pulses and mammalian dynamics: conceptual models for hummock grasslands and other Australian desert habitats. Biological Reviews of the Cambridge Philosophical Society 85, 501–521.
| Resource pulses and mammalian dynamics: conceptual models for hummock grasslands and other Australian desert habitats.Crossref | GoogleScholarGoogle Scholar | 20015313PubMed |
Lohr, C. A., and Algar, D. (2020). Managing feral cats through an adaptive framework in an arid landscape. The Science of the Total Environment 720, 137631.
| Managing feral cats through an adaptive framework in an arid landscape.Crossref | GoogleScholarGoogle Scholar | 32325591PubMed |
Loss, S. R., and Marra, P. P. (2017). Population impacts of free-ranging domestic cats on mainland vertebrates. Frontiers in Ecology and the Environment 15, 502–509.
| Population impacts of free-ranging domestic cats on mainland vertebrates.Crossref | GoogleScholarGoogle Scholar |
MacKenzie, D. I., Nichols, J. D., Royle, J. A., Pollock, K. H., Bailey, L. L., and Hines, J. E. (2018). ‘Occupancy estimation and modeling: Inferring patterns and dynamics of species occurrence’, 2nd edn. (Academic Press: London, UK.)
McGregor, H., Moseby, K., Johnson, C. N., and Legge, S. M. (2020). The short-term response of feral cats to rabbit population decline: are alternative native prey more at risk? Biological Invasions 22, 799–811.
| The short-term response of feral cats to rabbit population decline: are alternative native prey more at risk?Crossref | GoogleScholarGoogle Scholar |
Medina, F. M., Bonnaud, E., Vidal, E., Tershy, B. R., Zavaleta, E. S., Josh Donlan, C., Keitt, B. S., Corre, M., Horwath, S. V., and Nogales, M. (2011). A global review of the impacts of invasive cats on island endangered vertebrates. Global Change Biology 17, 3503–3510.
| A global review of the impacts of invasive cats on island endangered vertebrates.Crossref | GoogleScholarGoogle Scholar |
Molsher, R., Dickman, C. R., Newsome, A. E., and Müller, W. (2005). Home ranges of feral cats (Felis catus) in central-western New South Wales, Australia. Wildlife Research 32, 587–595.
| Home ranges of feral cats (Felis catus) in central-western New South Wales, Australia.Crossref | GoogleScholarGoogle Scholar |
Moseby, K. E., and Hill, B. M. (2011). The use of poison baits to control feral cats and red foxes in arid South Australia I. Aerial baiting trials. Wildlife Research 38, 338–349.
| The use of poison baits to control feral cats and red foxes in arid South Australia I. Aerial baiting trials.Crossref | GoogleScholarGoogle Scholar |
Moseby, K. E., Hill, B. M., and Read, J. L. (2009). Arid Recovery: a comparison of reptile and small mammal populations inside and outside a large rabbit, cat and fox-proof exclosure in arid South Australia. Austral Ecology 34, 156–169.
| Arid Recovery: a comparison of reptile and small mammal populations inside and outside a large rabbit, cat and fox-proof exclosure in arid South Australia.Crossref | GoogleScholarGoogle Scholar |
Moseby, K. E., Read, J. L., Galbraith, B., Munro, N., Newport, J., and Hill, B. M. (2011). The use of poison baits to control feral cats and red foxes in arid South Australia II. Bait type, placement, lures and non-target uptake. Wildlife Research 38, 350–358.
| The use of poison baits to control feral cats and red foxes in arid South Australia II. Bait type, placement, lures and non-target uptake.Crossref | GoogleScholarGoogle Scholar |
Newsome, T. M., Ballard, G.-A., Dickman, C. R., Fleming, P. J. S., and van de Ven, R. (2013). Home range, activity and sociality of a top predator, the dingo: a test of the Resource Dispersion Hypothesis. Ecography 36, 914–925.
| Home range, activity and sociality of a top predator, the dingo: a test of the Resource Dispersion Hypothesis.Crossref | GoogleScholarGoogle Scholar |
Niedballa, J., Sollmann, R., Courtiol, A., and Wilting, A. (2016). camtrapR: an R package for efficient camera trap data management. Methods in Ecology and Evolution 7, 1457–1462.
| camtrapR: an R package for efficient camera trap data management.Crossref | GoogleScholarGoogle Scholar |
Nogales, M., Vidal, E., Medina, F. M., Bonnaud, E., Tershy, B. R., Campbell, K. J., and Zavaleta, E. S. (2013). Feral cats and biodiversity conservation: the urgent prioritization of island management. Bioscience 63, 804–810.
| Feral cats and biodiversity conservation: the urgent prioritization of island management.Crossref | GoogleScholarGoogle Scholar |
Norbury, G. L., Pech, R. P., Byrom, A. E., and Innes, J. (2015). Density-impact functions for terrestrial vertebrate pests and indigenous biota: guidelines for conservation managers. Biological Conservation 191, 409–420.
| Density-impact functions for terrestrial vertebrate pests and indigenous biota: guidelines for conservation managers.Crossref | GoogleScholarGoogle Scholar |
Palmer, R., Anderson, H., Richards, B., Craig, M. D., and Gibson, L. A. (2021). Does aerial baiting for controlling feral cats in a heterogeneous landscape confer benefits to a threatened native meso-predator? PLoS One 16, e0251304.
| Does aerial baiting for controlling feral cats in a heterogeneous landscape confer benefits to a threatened native meso-predator?Crossref | GoogleScholarGoogle Scholar | 33961676PubMed |
R Core Team (2019). ‘R: a language and environment for statistical computing.’ (R Foundation for Statistical Computing: Vienna, Austria.) Available at https://www.r-project.org/.
Reddiex, B., and Forsyth, D. M. (2006). Control of pest mammals for biodiversity protection in Australia. II. Reliability of knowledge. Wildlife Research 33, 711–717.
| Control of pest mammals for biodiversity protection in Australia. II. Reliability of knowledge.Crossref | GoogleScholarGoogle Scholar |
Richards, J., and Algar, D. (2010). Sustained introduced predator control in the southern rangelands of Western Australia. Unpublished report, Australian Wildlife Conservancy.
Robley, A., Gormley, A., Forsyth, D. M., Wilton, A. N., and Stephens, D. (2010). Movements and habitat selection by wild dogs in eastern Victoria. Australian Mammalogy 32, 23–32.
| Movements and habitat selection by wild dogs in eastern Victoria.Crossref | GoogleScholarGoogle Scholar |
Roshier, D. A., Hotellier, F. L., Carter, A., Kemp, L., Potts, J., Hayward, M. W., and Legge, S. M. (2020). Long-term benefits and short-term costs: small vertebrate responses to predator exclusion and native mammal reintroductions in south-western New South Wales, Australia. Wildlife Research 47, 570–579.
| Long-term benefits and short-term costs: small vertebrate responses to predator exclusion and native mammal reintroductions in south-western New South Wales, Australia.Crossref | GoogleScholarGoogle Scholar |
Ruscoe, W. A., Ramsey, D. S. L., Pech, R. P., Sweetapple, P. J., Yockney, I., Barron, M. C., Perry, M., Nugent, G., Carran, R., Warne, R., Brausch, C., and Duncan, R. P. (2011). Unexpected consequences of control: competitive vs. predator release in a four-species assemblage of invasive mammals. Ecology Letters 14, 1035–1042.
| Unexpected consequences of control: competitive vs. predator release in a four-species assemblage of invasive mammals.Crossref | GoogleScholarGoogle Scholar | 21806747PubMed |
Salo, P., Korpimaki, E., Banks, P. B., Nordstrom, M., and Dickman, C. R. (2007). Alien predators are more dangerous than native predators to prey populations. Proceedings. Biological Sciences 274, 1237–1243.
| Alien predators are more dangerous than native predators to prey populations.Crossref | GoogleScholarGoogle Scholar | 17360286PubMed |
Shionosaki, K., Yamada, F., Ishikawa, T., and Shibata, S. (2015). Feral cat diet and predation on endangered endemic mammals on a biodiversity hot spot (Amami-Ohshima Island, Japan). Wildlife Research 42, 343–352.
| Feral cat diet and predation on endangered endemic mammals on a biodiversity hot spot (Amami-Ohshima Island, Japan).Crossref | GoogleScholarGoogle Scholar |
Short, J., Turner, B., Risbey, D. A., and Carnamah, R. (1997). Control of feral cats for nature conservation. II. Population reduction by poisoning. Wildlife Research 24, 703–714.
| Control of feral cats for nature conservation. II. Population reduction by poisoning.Crossref | GoogleScholarGoogle Scholar |
Stewart, A., Portelli, D., and Nano, C. (2019). ‘Central Rock rat monitoring and feral cat control in the West MacDonnell Ranges, 2019.’ (Northern Territory Department of Environment and Natural Resources Flora and Fauna Division: Alice Springs, NT, Australia.)
Stobo-Wilson, A. M., Brandle, R., Johnson, C. N., and Jones, M. E. (2020). Management of invasive mesopredators in the Flinders Ranges, South Australia: effectiveness and implications. Wildlife Research 47, 720–730.
| Management of invasive mesopredators in the Flinders Ranges, South Australia: effectiveness and implications.Crossref | GoogleScholarGoogle Scholar |
Walsh, J. C., Wilson, K. A., Benshemesh, J., and Possingham, H. P. (2012). Unexpected outcomes of invasive predator control: the importance of evaluating conservation management actions. Animal Conservation 15, 319–328.
| Unexpected outcomes of invasive predator control: the importance of evaluating conservation management actions.Crossref | GoogleScholarGoogle Scholar |
Weston, M. A., Antos, M. J., and Tzaros, C. L. (2009). Sand pads: a promising technique to quantify human visitation into nature conservation areas. Landscape and Urban Planning 89, 98–104.
| Sand pads: a promising technique to quantify human visitation into nature conservation areas.Crossref | GoogleScholarGoogle Scholar |
Woinarski, J. C. Z., Burbidge, A. A., and Harrison, P. L. (2015). Ongoing unraveling of a continental fauna: decline and extinction of Australian mammals since European settlement. Proceedings of the National Academy of Sciences of the United States of America 112, 4531–4540.
| Ongoing unraveling of a continental fauna: decline and extinction of Australian mammals since European settlement.Crossref | GoogleScholarGoogle Scholar |
Woinarski, J. C. Z., Legge, S. M., and Dickman, C. R. (2019). ‘Cats in Australia: companion and killer.’ (CSIRO Publishing: Melbourne, Vic., Australia.)
Wysong, M. L., Hradsky, B. A., Iacona, G. D., Valentine, L. E., Morris, K., and Ritchie, E. G. (2020a). Space use and habitat selection of an invasive mesopredator and sympatric, native apex predator. Movement Ecology 8, 18.
| Space use and habitat selection of an invasive mesopredator and sympatric, native apex predator.Crossref | GoogleScholarGoogle Scholar | 32391154PubMed |
Wysong, M. L., Iacona, G. D., Valentine, L. E., Morris, K., and Ritchie, E. G. (2020b). On the right track: placement of camera traps on roads improves detection of predators and shows non-target impacts of feral cat baiting. Wildlife Research 47, 557–569.
| On the right track: placement of camera traps on roads improves detection of predators and shows non-target impacts of feral cat baiting.Crossref | GoogleScholarGoogle Scholar |