

Phylogeny of the fern family Aspleniaceae in Australasia and the south-western Pacific
Daniel J. Ohlsen A , Leon R. Perrie B , Lara D. Shepherd B , Patrick J. Brownsey B and Michael J. Bayly A CA School of Botany, The University of Melbourne, Parkville, Vic. 3010, Australia.
B Museum of New Zealand Te Papa Tongarewa, PO Box 467, Wellington 6140, New Zealand.
C Corresponding author. Email: mbayly@unimelb.edu.au
Australian Systematic Botany 27(6) 355-371 https://doi.org/10.1071/SB14043
Submitted: 3 November 2014 Accepted: 11 March 2015 Published: 29 June 2015
Abstract
Aspleniaceae is one of the largest fern families. It is species-rich in Australasia and the south-western Pacific (ASWP), where approximately 115 species occur. In the current study, the chloroplast regions rbcL, trnL–trnF and rps4–trnS were sequenced for 100 Aspleniaceae samples from ASWP. These data were combined with published sequences for species from New Zealand and other regions for phylogenetic analyses. Species of Aspleniaceae from ASWP were placed in six of the eight previously identified inter-continental clades. The majority of species from ASWP were placed in two of these clades, with the remaining four clades each being represented by three or fewer species. Strong biogeographic affinities with South-east Asia were observed and immigration, rather than local radiations of endemic taxa, appears to have made a more important contribution to patterns of diversity in ASWP. This study supports the current taxonomic practice of recognising two genera, Asplenium L. and Hymenasplenium Hayata, in Aspleniaceae, and identifies future taxonomic work required for the family in this region, including potential synonymising of species, and revision of species complexes or widespread species that are demonstrably non-monophyletic.
Introduction
Aspleniaceae is one of the largest fern families, including ~700 species (Kramer and Viane 1990; Smith et al. 2006). It is diverse in Australasia and the south-western Pacific (here abbreviated to ‘ASWP’ and used to encompass Australia, New Guinea, Vanuatu, Fiji, New Caledonia and New Zealand). Approximately 115 species of Aspleniaceae occur in this region, many of which are local endemics (Table 1).

|
The family principally comprises a large cosmopolitan genus, Asplenium L. (Smith et al. 2006). Many taxonomic treatments have also recognised several morphologically identifiable groups as segregate genera or sections (e.g. Copeland 1947; Pichi Sermolli 1977; Tryon and Tryon 1982), some of which occur in ASWP, including Asplenium section Thamnopteris C.Presl, Hymenasplenium Hayata, Loxoscaphe T.Moore and Pleurosorus Fée (Brownsey 1998; Brownsey and Perrie 2011). However, molecular studies are encouraging retention of only one segregate genus, Hymenasplenium, which is resolved as sister to all other Aspleniaceae, in contrast to the other segregate genera that are nested within Asplenium (Murakami 1995; Murakami et al. 1999a; Schneider et al. 2004).
Molecular systematic studies have included many species and groups of Aspleniaceae from various parts of the world (e.g. Murakami 1995; Murakami et al. 1999a, 1999b; Pinter et al. 2002; Van den heede et al. 2003; Schneider et al. 2004, 2005; Bellefroid et al. 2010; Dong et al. 2012), including New Zealand (Perrie and Brownsey 2005; Shepherd et al. 2008a, 2008b; Perrie et al. 2010). However, the phylogenetic positions of very few Aspleniaceae from other areas of ASWP have been studied through DNA sequencing (Schneider et al. 2004; Schneider et al. 2005; Yatabe et al. 2009).
Morphological classifications (e.g. circumscriptions of segregate genera) and molecular phylogenetic analyses both support the notion that Aspleniaceae in Australia includes several widespread lineages. Schneider et al. (2004) established a global Aspleniaceae chloroplast phylogeny incorporating species from most parts of the world, and defined eight major clades. That study included some New Zealand Aspleniaceae, and some widespread species shared with ASWP. Fourteen species (of ~115) that occur in ASWP were included in their study and were placed in four of the major molecular groups. Among these species were A. nidus (species authorities provided in Tables 1 and 2) and A. phyllitidis of section Thamnopteris, and Hymenasplenium unilaterale (as A. unilaterale). In addition, Pleurosorus hispanicus (as A. hispanicum) from Europe was placed in a separate group from any species of ASWP included in the study. These findings indicated that at least five of the eight major groups occur in ASWP. However, it remains to be confirmed whether Australian Pleurosorus is closely related to the European Pleurosorus species, and whether species shared between ASWP and other regions are truly conspecific. With limited sampling from ASWP, it also remains to be determined whether other clades are represented in this region, and to which clades the majority of local species belong.

|
The most detailed molecular analysis of Aspleniaceae from ASWP is that of Perrie and Brownsey (2005), which focussed on Aspleniaceae in New Zealand. They sequenced the chloroplast regions rbcL and trnL–trnF for most Aspleniaceae of New Zealand, confirming relationships established by Schneider et al. (2004) and elucidating the close relationship of many previously non-sampled New Zealand species to lineages detected by Schneider et al. (2004). New Zealand Aspleniaceae was found to be polyphyletic, with species being placed in four separate parts of the global phylogeny of Schneider et al. (2004). The majority were placed in a clade equivalent to the ‘Austral’ group of regularly hybridising species (Brownsey 1977a), nested within Clade V of Schneider et al. (2004). The Austral group has been found to contain four clades, including the ‘Obtusatum’, ‘Bulbiferum’ and ‘Flaccidum’ groups (Perrie and Brownsey 2005; Shepherd et al. 2008a), whose species are shared between at least south-eastern Australia and New Zealand, and a further group of two Lord Howe Island species (Shepherd et al. 2008a). However, the full geographic extent of the Austral group remains to be determined.
Despite past studies, the relationships of many species of the Aspleniaceae of ASWP, to each other and to those elsewhere, remain to be assessed by molecular phylogenetic analyses. This is a substantial gap in understanding the systematics of Aspleniaceae, given taxa of ASWP are a large proportion of its worldwide diversity. The present study uses sequences of the chloroplast regions rbcL, rps4–trnS and trnL–trnF for all Australian species recognised by Brownsey (1998), and many Aspleniaceae from nearby New Guinea, and the south-western Pacific, with the aim of constructing a well resolved molecular phylogeny of the family in this region. Its goals are to better understand the evolutionary and biogeographic history of this group in ASWP, including the contributions to Aspleniaceae diversity in the ASWP made by in situ radiations versus accumulation of lineages through immigration. Additionally, the present study tests the current generic placement of taxa from the region and assesses the delimitation of some species, especially widely distributed species that are disjunct between different parts of ASWP.
Materials and methods
Sampling for molecular study
In total, 296 new DNA sequences were generated from 100 recent collections from wild populations in New Guinea, Australia, Vanuatu, Fiji and New Caledonia (Table 2). This included all Australian species recognised by Brownsey (1998), and several species from the south-western Pacific and New Guinea. Some collections could not be confidently assigned to a current species and are putatively new; these include Australian A. sp. ‘Windsor Tableland’ and New Caledonian A. sp. ‘New Caledonia’. In addition to the new sequences generated here, sequences for 70 further accessions were harvested from GenBank. For Aspleniaceae of ASWP this included any accessions sequenced for at least rbcL and trnL–trnF; for taxa from other parts of the world, this included accessions sequenced for all loci. Care was taken to include the closest sequenced relatives of all ASWP species, on the basis of previous studies and preliminary analyses, as well as a representative sample of Aspleniaceae diversity worldwide.
DNA isolation, amplification, and sequencing
DNA was extracted from 20 mg of silica gel-dried young leaf tissue. For all samples, apart from those from Fiji, leaf tissue was ground using a mortar and pestle with the aid of acid-washed grinding sand (Ajax Finechem, Sydney, NSW, Australia). DNA was isolated from ground samples using a DNeasy Plant Mini Kit (QIAGEN, Melbourne, Vic., Australia), following the manufacturer’s instructions and with a final elution volume of 100 µL.
The chloroplast DNA markers rbcL, trnL–trnF (trnL intron and trnL–trnF intergenic spacer), and rps4–trnS (rps4 gene and rps4–trnS intergenic spacer) were sequenced. These regions were chosen because (1) they are routinely used in fern systematics, enabling comparison of sequences from other studies, including Schneider et al. (2004, 2005) and (2) good resolution of both higher- and lower-level relationships can be achieved in Aspleniaceae employing these regions.
Chloroplast DNA markers were amplified by polymerase chain reaction (PCR), performed on a MyCycler thermal cycler (Bio-Rad, Sydney, NSW). Reaction mixtures comprised 5 µL of 5× MyTaq reaction buffer containing 5 mM of each dNTP and 15 mM MgCl2 (Bioline, Sydney, NSW), 0.125 µL (0.625 units) MyTaq DNA Polymerase (Bioline), 10 ρmol of each primer, 2.0 µL of extracted DNA, and ultrapure water added to make a total volume of 25 µL. The rbcL gene was amplified using the primers ESRBCL1F (5ʹ-ATG TCA CCA CAA ACG GAG ACT AAA GC-3ʹ) and ESRBCL1361R (5ʹ-TCA GGA CTC CAC TTA CTA GCT TCA CG-3ʹ) (Schuettpelz and Pryer 2007). Asplenium pellucidum, A. caudatum and A. longissimum were amplified using aF (5ʹ-ATG TCA CCA CAA ACA GAG ACT AAA GC-3ʹ) and cR (5ʹ-GCA GCA GCT AGT TCC GGG CTC CA-3ʹ) (Hasebe et al. 1994), because of unsuccessful amplification with ESRBCL1F and ESRBCL1361R. The trnL intron, trnL 3ʹ exon and trnL–trnF intergenic spacer were amplified using the primers F (5ʹ-ATT TGA ACT GGT GAC ACG AG-3ʹ; Taberlet et al. 1991) and Fern1 (5ʹ-GGC AGC CCC CAR ATT CAG GGR AAC C-3ʹ; Trewick et al. 2002). For Australian A. baileyanum, A. polyodon and A. tenerum and New Caledonian A. polyodon, amplification with the internal primers D (5ʹ-GGG GAT AGA GGG ACT TGA AC-3ʹ) and E (5ʹ-GGT TCA AGT CCC TCT ATC CC-3ʹ) (Taberlet et al. 1991) was required for sequencing reactions to obtain adequate trnL–trnF sequence for analysis. The rps4 gene and the rps4–trnS intergenic spacer were amplified using the forward primer 5ʹ-ATG TCM CGT TAY CGA GGR CCT CGT-3ʹ and the reverse primer 5ʹ-TAC CGA GGG TTC GAA TC-3’ (Schneider et al. 2005).
Polymerase chain reaction thermocycling conditions for rbcL and trnL–trnF involved an initial denaturation step of 95°C for 1 min, followed by 33 cycles of 95°C for 1 min, 55°C for 1 min, and 65°C for 4 min, and a final extension of 65°C for 5 min. PCR thermocycling conditions for rps4–trnS involved an initial denaturation step of 95°C for 1 min, followed by 37 cycles of 94°C for 1 min, 56°C for 1 min, and 72°C for 1 min, and a final extension of 72°C for 5 min. DNA concentrations were quantified by electrophoresis against Hyperladder I and EasyLadder I (Bioline) and PCR products were purified using illustra ExoSTAR 1-step enzymatic purification (GE Healthcare Life Sciences, Freiburg, Germany). Purified PCR products were then sent to the Australian Genome Research Facility (AGRF), Melbourne Branch, where sequencing reactions and capillary separation, using the 96-capillary analyser AB 3730xl sequencing platform (Applied Biosystems, Foster City, CA, USA), were performed.
For the Fijian samples, genomic DNA was extracted, PCR amplified and the PCR products purified and sequenced following Shepherd et al. (2007).
Sequence editing, alignment and analysis
Sequences were edited using Sequencher v. 4.9 (Gene Codes Corporation, Ann Arbor, MI, USA). Sequences were aligned manually in Se-Al Sequence Alignment Editor v. 2.0a11 (Rambaut 2002). Regions of ambiguous alignment were excluded from analyses.
A single rbcL, trnL–trnF and rps4–trnS alignment (submitted to TreeBase) was analysed. Congruence of phylogenetic signal between cpDNA markers was tested by comparing preliminary maximum-parsimony (MP) analyses of individual markers. These analyses showed that any difference in signal among loci affected only some poorly supported nodes that are not shown in the figures of the present study, providing justification for concatenation of loci. The following three analyses were conducted: a MP, a maximum likelihood (ML) and a Bayesian analysis (BA). Athyrium filix-femina (Athyriaceae), Rhachidosorus consimilis (Rhachidosoraceae) and Diplaziopsis cavaleriana (Diplaziopsidaceae) were chosen as outgroups. Rhachidosorus and Diplaziopsis are the closest related genera to Aspleniaceae that have all the chloroplast loci employed in the present study publically available (Rothfels et al. 2012). Athyrium was also included to give a more distant relative in Eupolypods II (Rothfels et al. 2012).
Maximum-parsimony analyses were run in PAUP* v 4.0β10 (Swofford 2002). Gap characters in the alignment were treated as a fifth character state. For multiple-base indels, characters were excluded from analyses so that indels were each represented only by a single gap character, when variability did not occur within indels. Question marks, the character recognised in PAUP for missing data, were used to fill gaps, where needed, when indels fell across otherwise variable regions of the alignment. A heuristic tree search was used, with delayed character-state optimisation (DELTRAN) and starting trees were obtained by a closest addition sequence, followed by tree bisection-reconnection (TBR) branch swapping. Bootstrap support for nodes was determined using 1000 ‘full heuristic’ replicates.
MrModel Test v2 (Nylander 2004) was used with the Akaike information criterion (AIC) to assess the most appropriate model of sequence evolution for BA and ML analyses. Separate models of sequence evolution were assessed for the different coding and non-coding for BA and for the entire dataset for ML analyses. BA was performed in MrBayes 3.2.1 (Huelsenbeck and Ronquist 2001). For BA, two concurrent analyses were run, each with four Markov chains of 3 000 000 generations. This number of generations allowed the standard deviation of split frequencies to become less than 0.01. Trees were sampled every 1000 generations. A GTR+I+G substitution model was chosen for rbcL, a GTR+G substitution model was chosen for the trnL intron, trnL–trnF spacer, rps4, and the rps4–trnS spacer, and a K80+I substitution model was chosen for the trnL exon. Default priors were used, and the substitution-model parameters were unlinked across the loci. Tracer v1.5 (Rambaut and Drummond 2009) was used to determine samples to be discarded as ‘burn-in’. After the ‘burn-in’ was discarded, 2700 samples remained for consensus trees to be calculated from. Gap characters were coded in a separate binary dataset as restriction data.
Maximum likelihood (ML) analyses were performed with the PhyML 3.0 web server (http://www.atgc-montpellier.fr/phyml, accessed 16 October 2014; Guindon et al. 2010). A GTR+I+G substitution model with nst = 6, Γ = 1.3768 and pinvar = 0.2685 was chosen. ML-optimised equilibrium frequencies, subtree pruning and regrafting (SPR) branch-swapping and 10 random starting trees were applied. Branch support was assessed using an approximate likelihood-ratio test (aLRT, Anisimova and Gascuel 2006). Gap characters were treated as missing data and a single model of sequence evolution was chosen for the entire alignment in ML analyses because the PhyML 3.0 web server cannot process coded indels or apply separate models to different sections of the dataset in one analysis.
Results
Phylogeny of Aspleniaceae
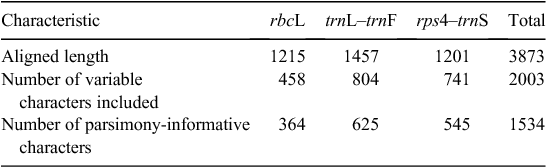
Sequence characteristics for the alignment are summarised in Table 3. Thirteen strongly supported clades, I–XIII, are here defined within the Aspleniaceae phylogeny. Seven of these clades occur in ASWP. These are Clade I or Hymenasplenium, Clade III, Clade V, Clade VI, Clade VIII, Clade IX, Clade XI and Clade XIII (Fig. 1). The distributions of these clades in ASWP are shown in Fig. 2. The majority of ASWP species were placed in Clades V or VI. Topologies of trees generated by the different tree-building methods were very similar and the consensus tree resulting from the BA is presented and described. Clade I (Hymenasplenium) and Clade II were successive sisters to the remaining Aspleniaceae. Relationships among Clades III–XIII were not well resolved, and most nodes among these clades had poor support (<80% bootstrap support (BS); <0.95 posterior probability (PP) and aLRT support values).
|

|

|
Australasian and south-western Pacific species in Clades I, III, VII, VIII and IX
Clade I (Hymenasplenium) is represented in ASWP by the following three species: H. excisum, H. unilaterale and A. wildii. Clade I is absent from temperate Australasia (Fig. 2A).
Clade III (Pleurosorus clade) is represented in ASWP by A. subglandulosum (which has been previously known as Pleurosorus rutifolius and P. subglandulosus), and is widespread in temperate and subtropical areas of Australia and New Zealand (Fig. 2B). Australasian accessions form a clade with A. hispanicum (P. hispanicus) from Europe and A. papaverifolium (P. papaverifolius) from Chile (Fig. 1).
Clade IX is known only from ASWP and comprises A. flabellifolium and A. pauperequitum. Clades VIII, XI and XIII are represented in ASWP by single species. Clade XI is represented in ASWP by A. trichomanes and Clade XIII by A. normale. Clades XI and XIII were part of a larger clade that was strongly supported in BA and ML analyses. Asplenium trichomanes was not resolved as monophyletic but formed two clades on the basis of the substrate of samples, i.e., those from limestone in one clade (samples labelled 4n Australia, 4n Romania, 2n France; Fig. 1), and those not restricted to limestone in another (samples labelled 2n Australia, 6n New Zealand). Representatives of ASWP A. normale formed a clade and were closely related to A. normale and A. hobdyi samples from Hawaii. ASWP distributions of species in Clades IX, XI and XIII are shown in Fig. 2E. Clade VIII is represented in the south-western Pacific by a single species, A. dognyense from New Caledonia (Figs 1, 2F).
Relationships among species of Clade V
Clade V contained the largest number of species from ASWP sequenced in the present study, with 39 species being represented (Fig. 3A). Several strongly supported clades were resolved (Fig. 3A). These include the following: a clade referred to here as Thamnopteris 1 (not strongly supported in MP analysis), formed entirely of section Thamnopteris collections; the Thamnopteris 2 clade, comprising A. antiquum and a New Guinean accession labelled as A. nidus; the Austral group (sensu Perrie and Brownsey 2005); the A. tenerum clade, comprising A. carruthersii, A. decorum, A. tenerum and A. tenuiculum; a clade of A. marattioides and A. scandens; and a clade of A. simplicifrons and A. subemarginatum. Relationships between these clades were not resolved with strong support (Fig. 3A), but relationships within them were mostly congruent between analyses.

|
The Thamnopteris 1 clade included two clades, namely, a clade of mainly A. australasicum-like species, and another composed of A. nidus and A. phyllitidis. Within the latter clade, a New Guinean population of A. phyllitidis was sister to a clade of A. nidus samples from Australia and Vanuatu. In the clade comprising mainly A. australasicum-like species, the Australian species A. goudeyi, A. harmanii, and Australian A. australasicum and New Caledonian A. australasicum formed a clade. This Australian and New Caledonian clade was sister to a clade including A. australasicum from Vanuatu and Fiji, a sample not identified to species level from New Guinea, and a New Guinean population identified as A. nidus. This is one of four clades in which A. nidus was placed by the analyses, making material identified as A. nidus markedly polyphyletic. Asplenium australasicum was also placed in three separate clades and is polyphyletic.
Within the Austral group, four strongly supported principal clades were recovered. These were a clade of three Lord Howe Island endemic species (A. milnei, A. surrogatum and A. pteridioides) and the Obtusatum, Flaccidum and Bulbiferum groups. Relationships among these strongly supported clades of the Austral group were not well supported. Within the Obtusatum group, a New Caledonian clade of A. vieillardii and A. oligolepidum was sister to species that occur in New Zealand. Within the Flaccidum group, Fijian samples of A. gibberosum and A. stenolobum formed a clade sister to a clade comprising New Zealand A. shuttleworthianum, Vanuatu A. gibberosum and New Caledonian A. subflexuosum.
Some species in Clade V lack close relatives among the sampled species. Samples of both A. amboinense and A. bipinnatifidum (each strongly supported as monophyletic) do not form well supported clades with any other species. Likewise, A. scandens and A. marattioides, although shown as sister species, are not closely related to other clades and are highly divergent from each other; i.e. terminal branch lengths in MP analyses of 58 and 65 steps respectively. The same is true of the pair A. subemarginatum and A. simplicifrons (terminal branch lengths 56 and 46).
Relationships among species of Clade VI
Clade VI was the second-most diverse clade in ASWP. At least 25 species of ASWP are placed in this clade (Fig. 3B). Several clades within clade VI were well supported (Fig. 3B). Sister to other groups of Clade VI was the A. aethiopicum clade, which included A. aethiopicum, Australian and New Guinean A. parvum and New Caledonian A. filidens. Also well supported was a sister relationship between the A. caudatum clade and the rest of Clade VI, excluding the A. aethiopicum clade. The A. caudatum clade comprised A. caudatum, A. longissimum (A. sp. ‘Litchfield’ Russell-Smith 6470 sensu Short et al. 2003), and A. pellucidum.
A sister relationship between the A. cuneatum and A. capitisyork clades was well supported. The A. capitisyork clade comprised A. capitisyork from Cape York, far-northern Australia, which was sister to A. keyserrianum and A. cromwellianum, both from New Guinea. The A. cuneatum clade was a large clade of species from northern Australia, New Guinea, and tropical Pacific Islands, including A. affine, A. athertonense, A. baileyanum, A. cuneatum, A. cromwellianum, A. insiticium, A. lobulatum, A. nitidum, A. setisectum and New Guinean samples unassigned to species. Samples identified as A. cuneatum were not monophyletic: an Australian A. cuneatum accession was sister to A. sp. ‘Windsor Tableland’ from north-eastern Australia, whereas Fijian and New Caledonian A. cuneatum were sister to Fijian A. insiticium. None of the analyses revealed with certainty the closest relatives of Australian A. baileyanum, New Guinean A. affine and A. sp. ‘New Caledonia’. Resolution of the relationships between A. lobulatum, A. sp. indet. ‘New Guinea 1’ and A. athertonense was poor because of low sequence variation among these species (pair-wise differences across all loci no more than five base pairs) and variation among A. lobulatum populations.
The rest of Clade VI comprised species or groups of species whose phylogenetic positions were not well resolved. These included A. laserpitiifolium, A. novoguineense, a clade comprising an Australian and a New Caledonian accession of A. polyodon, New Zealand A. polyodon, the A. paleaceum clade, and the A. listeri clade. The A. paleaceum clade comprised A. attenuatum, A. bicentenniale, A. carnarvonense and A. paleaceum. These species were resolved as a polytomy. The A. listeri clade comprised epiphytic A. polyodon from Christmas Island, north-west of Australia, a coastal limestone-dwelling sample of A. listeri from Christmas Island, and coastal limestone-dwelling samples from the Pacific attributed to A. polyodon. Consequently, A. polyodon is not monophyletic.
Discussion
The Aspleniaceae phylogeny: major clades and generic limits
Results here are congruent with previously published Aspleniaceae phylogenies (Murakami 1995; Murakami et al. 1999a; Schneider et al. 2004, 2005). All major clades have been recovered in previous studies (Schneider et al. 2004, 2005; Chang et al. 2013) and the relationships among these clades also match what has been previously found (Schneider et al. 2004, 2005). An important congruent finding was the placement of Clade I, Hymenasplenium, as sister to other Aspleniaceae. This provides further support for treating the Hymenasplenium clade as a separate genus, as has been suggested (Smith et al. 2006), and generally followed in recent treatments (e.g. Murakami 1995; Murakami et al. 1998, 1999a; Sasaki 2008; Brownsey and Perrie 2011; Gabancho and Prada 2011). The Australian endemic, A. wildii, has been previously noted as morphologically similar and most likely related to species of Hymenasplenium on the basis of its creeping rhizome and pinnate fronds (Brownsey 1998). This is confirmed by placement of A. wildii in the Hymenasplenium clade by the chloroplast phylogeny of the present study, and a new combination for A. wildii in Hymenasplenium will be made elsewhere.
All species of Pleurosorus and the section Thamnopteris were nested well within Asplenium in Clades III and V respectively in agreement with previous molecular phylogenetic studies (Schneider et al. 2004, 2005), and supporting inclusion of these taxa in Asplenium. Section Thamnopteris was previously found not to be monophyletic with predominantly South-east Asian taxon sampling (Murakami et al. 1999a; Yatabe et al. 2009). In the present study, monophyly of section Thamnopteris was not supported. Section Thamnopteris accessions form two clades, one including the majority of accessions, including some A. nidus samples, and the other with A. antiquum and a New Guinean accession supplied to us as A. nidus. Placement of South-east Asian A. nidus populations in both these clades has previously been demonstrated (Yatabe et al. 2001, 2009), and this emphasises that morphological features used for identification in section Thamnopteris do not delimit monophyletic species (Murakami et al. 1999b; Yatabe et al. 2001; Dong et al. 2012). Furthermore, the species with entire fronds not belonging to section Thamnopteris (i.e. without an anastomosing vein near the margin), A. simplicifrons and A. amboinense, were placed in two other separate positions in Clade V, indicating that the transition between simple and divided fronds has occurred multiple times even within Clade V.
The present study has shown that the segregate genus Loxoscaphe, defined by short, submarginal and pouch-like sori (Gastony and Johnson 2001), is also not monophyletic, having arisen on at least two independent occasions. Bellefroid et al. (2010) showed that African species of the Loxoscaphe group were monophyletic. One of these African species, A. loxoscaphoides, is included in the present study and does not form a clade with south-western Pacific species, A. gibberosum and A. stenolobum (Fig. 3A), previously included in Loxoscaphe (Brownlie 1977). The close relationship of these south-western Pacific species to A. shuttleworthianum from New Zealand is supported by the similar ‘loxoscaphoid’ morphology of A. shuttleworthianum (Perrie and Brownsey 2005).
Relationships found here within Clade V are congruent with analyses of Perrie and Brownsey (2005), who focussed on this clade in New Zealand with a smaller sample of species. Among the Austral group are the well supported Obtusatum, Flaccidum and Bulbiferum groups of Perrie and Brownsey (2005). However, the larger number of species sampled in the present study did not resolve uncertainty regarding relationships among these three groups (Perrie and Brownsey 2005; Shepherd et al. 2008a).
Several species in Clade V, such as A. carruthersii, A. scandens, A. subemarginatum, A. simplicifrons and A. marattioides, had particularly long branch lengths when compared with other species (Fig. 3A). It is possible that these lengths were artefacts of low taxon sampling in New Guinea, where most of these species occur and possibly many of their close relatives. The morphologically distinct Malesian group of approximately four species sometimes recognised as the segregate genus Diplora Baker (see Mickel 1976; Copeland 1913, 1949) likely contributes to this undetected diversity. Partial rbcL sequence obtained from herbarium material (WELT P017103) of a putatively new Diplora species from the Solomon Islands is placed with the A. marattioides+A. scandens clade (L. Shepherd, unpub. data).
Biogeography of Aspleniaceae in ASWP
The present study has highlighted that the Aspleniaceae of ASWP is an assemblage of different lineages, some of which are widespread throughout the world. Molecular dating of Aspleniaceae by Perrie and Brownsey (2005), using rbcL, supported comparatively recent exchange of ASWP Aspleniaceae with those of other regions. The longest isolation of a clade in and around this region may be that of the Austral group. With the exception of a possibly recently formed circum-antarctic distribution in A. obtusatum (Brownlie 1965; Parris 2001), this clade appears to be endemic to southern Australasia and the islands of the south Pacific (Fig. 2C). Otherwise, much more recent exchange with other regions is prevalent, with divergence between the other Aspleniaceae of ASWP and their closest relatives in other regions not nearly as deep as that of the Austral group. This is especially the case for populations of species shared among various regions of the world (see Table 1).
All clades detected within ASWP, including the Austral group, are nested within clades defined by Schneider et al. (2004), which are all widespread in other regions. The presence of five of these clades in ASWP had been previously shown by molecular data or suspected on the basis of morphology (e.g. Clade III – Pleurosorus, see Fig. 1). However, our study has extended the known distribution of Clade VIII. Clade VIII was previously known only from Africa, Europe and South America (Schneider et al. 2004), but its distribution is here shown to include also the south-western Pacific (Fig. 2F), where its single known representative is the New Caledonian species A. dognyense. This species, or a close relative, may also occur in Western Samoa (WELT P015959!). Furthermore, A. dognyense is morphologically similar and presumably closely related to the unsequenced A. macraei Hook. & Grev., differing mainly by its proliferous fronds (Brownlie 1969; Palmer 2003). Asplenium macraei is known from the Austral Islands, and Hawaii (Palmer 2003), suggesting that Clade VIII may be widespread in the Pacific.
Some subclades within the major clades defined by Schneider et al. (2004) and the present study are also widespread in several regions of the world. The A. aethiopicum clade in Clade VI is likely to be pantropical. This clade contains the A. aethiopicum/A. praemorsum complex, with Australian and African samples shown to be closely related to each other and to cultivated material of A. praemorsum, which is generally considered to be an American species (Fig. 3B). The A. aethiopicum clade is also likely to contain Hawaiian A. contiguum Kaulf., Asian A. lepturus J.Smith ex C.Presl and South American A. serra Langsdorff & Fisher. These species have narrow and caudate pinnae, long-creeping rhizomes and reticulate perispores without ridges (Youxing and Viane 2013), a combination of characters so far known only from species of the A. aethiopicum clade.
A subclade within Clade III is also widespread in several regions of the world. The subclade of species previously attributed to Pleurosorus (A. hispanicum, A. papaverifolium, A. subglandulosum; Fig. 1) positioned within Clade III confirms a particularly remarkable disjunction. No intervening Pleurosorus species are known, making this disjunction comparable to that seen in the trans-continental distributions of A. dalhousiae Hook. (Van den heede et al. 2003), A. platyneuron (L.) Britton (Wagner et al. 1993; Moran and Smith 2001; Crouch et al. 2011) and A. exiguum Beddome (Wagner et al. 1993).
The greatest sharing of clades between ASWP and another region of the world is with adjacent areas of Malesia and South-east Asia (Fig. 4). Thirty-one species from ASWP also occur in that region (see Table 1). Most of the species shared between ASWP and adjacent Malesia and South-east Asia were placed in Clades V and VI. In addition, several other South-east Asian and Malesian species not sampled in the present study are morphologically similar to some Clade V and Clade VI species and also undoubtedly belong to these clades (e.g. several section Thamnopteris species). The high diversity of these clades in this adjacent region helps explain the higher diversity of these clades in ASWP, particularly Clade VI, which predominantly occurs in tropical areas of ASWP adjacent to Indonesia (Fig. 2D). Of the Clade VI species in ASWP, only A. aethiopicum, A. attenuatum of the A. paleaceum clade, and the A. polyodon complex occur south of the tropics (Fig. 2D). Apart from the Austral group, other clades in Clade V also predominantly occur in tropical areas of ASWP, with A. australasicum of the Thamnopteris I clade being the only species that extends into temperate areas (Fig. 2C).

|
Clades less well represented or absent in ASWP (i.e. Clades II, III, IV, VII, VIII, X, XI, XII and XIII) are predominant in regions more distant from Australia (Fig. 1). Previous studies have also shown several species not included in the present study and that do not occur in ASWP to belong to one of these clades (Van den heede et al. 2003; Schneider et al. 2004, 2005; Li and Lu 2006; Dyer et al. 2012; Chang et al. 2013). The influence of surrounding regions on Aspleniaceae diversity in the Pacific is further demonstrated by Hawaii. All the major clades present in ASWP are likely to be present in Hawaii on the basis of previous studies (Schneider et al. 2004, 2005; Schuettpelz and Pryer 2007). However, the ‘black-stemmed spleenwort’ clade comprising Clades XI, XII and XIII, well represented in surrounding continents of the northern hemisphere (Schneider et al. 2004), contains more species in Hawaii than any of the other major clades with at least 10 species (Schneider et al. 2004, 2005), demonstrating the importance of the closer northern hemisphere continents to the fern diversity of Hawaii (also see Geiger et al. 2007). In contrast, Clade V, predominant in the southern hemisphere (Schneider et al. 2004) and south Pacific, may be represented in Hawaii only by A. nidus.
The importance of immigration to patterns of diversity in Aspleniaceae of ASWP and surrounding areas can also be appreciated from the high proportion of species in each area of ASWP whose closest relatives are from other areas (Fig. 4). For instance, among the Vanuatu and Fijian species, more have a sister relationship with species of Malesia than with other local species. Even among taxa from the large continent of Australia, a high proportion have their closest relatives in other areas, especially in Malesia (Fig. 4). The exception to this trend is New Zealand where a high proportion of species had their closest relatives in New Zealand, and more sister relationships were shared with Australia rather than Malesia (Fig. 4).
In general, even when local speciation has apparently occurred in parts of ASWP, it appears to have given rise to only minor species radiations. The largest radiation of species wholly endemic to a single land area comprises only the following three species: A. milnei, A. pteridoides and A. surrogatum endemic to Lord Howe Island, east of mainland Australia. This Lord Howe Island clade supports a previous suggestion that these species have evolved in geographic sympatry (Papadopulos et al. 2011). The New Caledonian clade that includes A. vieillardii and A. oligolepidum may also comprise three species, given that partial rbcL sequence from New Caledonian A. novae-caledoniae is apparently placed with this clade (L. Shepherd, unpub. data). The A. paleaceum clade contains four Australian species, but one of its species, A. attenuatum, is also known from New Caledonia (not sampled in the present study), making this clade not wholly endemic to Australia.
The majority of island endemic species in ASWP appear to have evolved on the same island where their closest relatives occur. Of the 19 island endemic species whose phylogenetic positions are known, possibly only six (A. dognyense, A. goudeyi, A. pauperequitum, A. tenuiculum, and possibly two cryptic species of section Thamnopteris) may not have their closest relative also occurring on the same island. These six may be the result of anagenetic speciation, i.e. where an island immigrant evolves into a separate species in isolation from its disjunct progenitor (Stuessy et al. 2006; Gehrke and Linder 2011; Patiño et al. 2014). This differs from most fern groups in which anagenetic speciation appears to be the prevalent source of island endemics (Patiño et al. 2014), including the Blechnaceae ferns of the south-western Pacific (Perrie et al. 2014).
Implications for species-level taxonomy
The present study resolves an outstanding taxonomic issue regarding the distinctiveness of A. carnarvonense from A. capitisyork. Brownsey (1998) considered that these two Queensland endemic species were very similar and, on the basis of spore and overall morphology, placed them together as the only members of the ‘Asplenium capitisyork’ group. In contrast, the chloroplast phylogenies presented here placed A. capitisyork and A. carnarvonense into separate clades with strong support (the A. capitisyork and A. paleaceum clades respectively). This result provides significant new evidence of the distinctiveness of these taxa, and supports their continued recognition as separate species. However, in Aspleniaceae, hybridisation and allopolyploidy are frequent (Lovis 1973; Reichstein 1981), and it is possible that A. capitisyork and A. carnarvonense are allopolyploids that share the same parental species but different maternal parents and, hence, different chloroplast haplotypes (Vogel et al. 1998). Nuclear DNA markers are required to test this hypothesis.
The present study also identifies the need for taxonomic revision for several members of ASWP Aspleniaceae. In particular, several species as currently circumscribed are not monophyletic (i.e. A. australasicum, A. cromwellianum, A. cuneatum, A. gibberosum, A. nidus, A. polyodon and A. tenerum) and potentially contain cryptic species (Paris 1989); taxonomic revision of these groups of species will require molecular study of populations from across their ranges, including outside ASWP. Non-monophyly of these species may be due to these species being allopolyploids with multiple and reciprocal origins (see above). However, a reciprocal polyploidy origin is an unlikely explanation for the Thamnopteris species and A. polyodon. Multiple chromosome counts have shown only tetraploid counts so far (Manton 1954; Manton and Sledge 1954; Bir 1960; Abraham et al. 1962; Kawakami 1970; Brownsey 1977b; Nakato 1987; Yatabe et al. 2001, 2002, 2009; Tindale and Roy 2002; Yatabe and Murakami 2003) and diploid species have not yet been recorded in Clade VI and only a few species in Clade V have been reported as diploid (Bir 1960; Mehra 1961; Tindale and Roy 2002). Instead, the prevalent ploidy level in Clades V and VI is tetraploid (Perrie and Brownsey 2005), suggesting that these species are likely derived from tetraploid ancestors and that non-monophyly of these taxa in chloroplast trees is not an artefact of reciprocal allopolyploidy (see Yatabe et al. 2009).
Some accessions in the dataset were genetically distinct, and are not easily assigned to a current species; they potentially represent undescribed species. Australian A. sp. ‘Windsor Tableland’ most closely resembles A. athertonense morphologically, but its chloroplast sequences were more closely related to Australian A. cuneatum. The genetic distinction of A. sp. ‘Windsor Tableland’ from both of these species was greater than that between other pairs of morphologically distinctive species such as A. athertonense and A. baileyanum (Fig. 3B). Likewise, A. sp. ‘New Caledonia’ was more genetically divergent from its closest relatives and the morphologically similar A. lobulatum than were several other well established and morphologically distinctive species pairs (Fig. 3B). This gives support towards treating these accessions as new species.
Conversely, some currently accepted species of Aspleniaceae from ASWP are both genetically (Fig. 3) and morphologically similar and might best be considered conspecific. This includes: A. parvum and A. filidens; limestone-dwelling A. polyodon in the Pacific and limestone-dwelling A. listeri from the Australian Territory of Christmas Island in the Indian Ocean; A. athertonense and A. lobulatum.
For some groups of morphologically distinctive species, only minor chloroplast variation was detected here (e.g. A. paleaceum clade, Fig. 3B). These groups potentially represent species complexes (Viane and Reichstein 2003) in which chloroplast similarity has resulted from hybridisation. Study of nuclear genetic variation is needed to more fully understand the relationships of these taxa and assess their taxonomic status.
The present study has clearly highlighted some unresolved taxonomic issues among Aspleniaceae of ASWP. Some of these, including the status of A. sp. ‘Windsor Tableland’, A. polyodon and relatives (including A. listeri), and the A. paleaceum clade are the subjects of more detailed works (Ohlsen 2014; Ohlsen et al. 2014a, 2014b) using additional genetic markers or morphological comparisons. The results of those studies, combined with the present one, will form the basis for taxonomic treatment for all Australian Aspleniaceae.
Conclusions
The present study has provided further support for many relationships within the Aspleniaceae recovered from previous studies (Murakami et al. 1999a; Schneider et al. 2004; Perrie and Brownsey 2005). It has also established relationships among most Aspleniaceae in ASWP, which were previously largely unknown. On comparison with taxa from other regions of the world, it has revealed that ASWP species belong to several large inter-continental clades. The widespread distribution of most species in the region and the close relationship of the locally endemic species to more widespread species or species endemic elsewhere indicate the importance of immigration as a source of diversity in ASWP. The present study has also exposed taxonomic issues at the genus and species level in Aspleniaceae. Although taxon sampling for some of the ASWP region was comprehensive, many species and groups of Aspleniaceae in Malesia were not sampled in the study. Molecular study involving more taxon sampling in Malesia could provide a focus for future study. Such study will further complete understanding of Aspleniaceae systematics in general, as well as elucidate more detailed biogeographic histories in ASWP and improve taxonomic circumscriptions among species of Aspleniaceae of this region.
Acknowledgements
We thank Bruce Gray, Alasdair Grigg, Dirk Karger, Michael Kessler, Jurgen Kluge, Braydon Maloney and Adele Neale for their assistance with collecting and Alan Smith for identifications of New Guinean material. Molecular work was performed in the Cookson Laboratory, School of Botany, The University of Melbourne, and the School of Biological Sciences, Victoria University of Wellington. This study was funded by a Bush Blitz research grant (BBR21-25) from the Australian Biological Resources Study, with support from The University of Melbourne School of Botany Foundation. This study forms part of Daniel J. Ohlsen’s PhD Thesis, titled Molecular systematics, biogeography, and taxonomy of the fern family Aspleniaceae in Australasia and the south-west Pacific, supported by an Australian Postgraduate Award.
References
Abraham A, Ninan AC, Mathew PH (1962) Studies on the cytology and phylogeny of the pteridophytes. VII. Observations on one hundred species of south Indian ferns. The Journal of the Indian Botanical Society 41, 339–421.Anisimova M, Gascuel O (2006) Approximate likelihood-ratio test for branches: a fast, accurate, and powerful alternative. Systematic Biology 55, 539–552.
| Approximate likelihood-ratio test for branches: a fast, accurate, and powerful alternative.Crossref | GoogleScholarGoogle Scholar | 16785212PubMed |
Bellefroid E, Rambe SK, Leroux O, Viane RLL (2010) The base number of ‘loxoscaphoid’ Asplenium species and its implication for cytoevolution in Aspleniaceae. Annals of Botany 106, 157–171.
| The base number of ‘loxoscaphoid’ Asplenium species and its implication for cytoevolution in Aspleniaceae.Crossref | GoogleScholarGoogle Scholar | 20498038PubMed |
Bir SS (1960) Cytological observations on the east Himalayan members of Asplenium Linn. Current Science 29, 445–447.
Brownlie G (1965) The geographical affinities of the south Pacific island fern floras. Pacific Science 19, 219–223.
Brownlie G (1969) Aspleniaceae. In ‘Flore de la Nouvelle-Caledonie et Dependances. No. 3 Pteridophytes’. (Ed. A Aubreville) pp. 184–201. (Museum of Natural History: Paris)
Brownlie G (Ed.) (1977) Aspleniaceae. In ‘The Pteridophyte Flora of Fiji’. (Ed. G Brownlie) pp. 208–228. (Cramer: Vaduz)
Brownsey PJ (1977a) Asplenium hybrids in the New Zealand flora. New Zealand Journal of Botany 15, 601–637.
| Asplenium hybrids in the New Zealand flora.Crossref | GoogleScholarGoogle Scholar |
Brownsey PJ (1977b) A taxonomic revision of the New Zealand species of Asplenium. New Zealand Journal of Botany 15, 39–86.
| A taxonomic revision of the New Zealand species of Asplenium.Crossref | GoogleScholarGoogle Scholar |
Brownsey PJ (1998) Aspleniaceae. In ‘Flora of Australia, Ferns, Gymnosperms and Allied Groups. Vol. 48’. (Ed. PM McCarthy) pp. 295–327. (ABRS: Canberra; and CSIRO, Melbourne)
Brownsey PJ, Perrie LR (2011) A revised checklist of Fijian ferns and lycophytes. Telopea 13, 513–562.
Brownsey PJ, Smith-Dodsworth JC (2000) ‘New Zealand ferns and allied plants.’ (David Bateman: Auckland)
Chang Y, Li J, Lu S, Schneider H (2013) Species diversity and reticulate evolution in the Asplenium normale complex (Aspleniaceae) in China and adjacent areas. Taxon 62, 673–687.
| Species diversity and reticulate evolution in the Asplenium normale complex (Aspleniaceae) in China and adjacent areas.Crossref | GoogleScholarGoogle Scholar |
Copeland EB (1913) On Phyllitis in Malaya and the supposed genera Diplora and Triphlebia. Philippine Journal of Science 8, 147–155.
Copeland EB (1947) ‘Genera Filicum. (Annales Cryptogamici et Phytophaologici vol. 5).’ (Chronica Botanica: Waltham, Massachusetts)
Copeland EB (1949) Aspleniaceae and Blechnaceae of New Guinea. Philippine Journal of Science 78, 207–229.
Crouch NR, Klopper RR, Burrows JE, Burrows SM (2011) Aspleniaceae. In ‘Ferns of Southern Africa: a comprehensive guide’. pp. 578–663. (Struik Publishers: Cape Town)
Dong SY, Mugahidin , Wei LL, Chao YS (2012) A new species of Asplenium section Thamnopteris (Aspleniaceae) from Indonesia. Blumea 57, 190–194.
| A new species of Asplenium section Thamnopteris (Aspleniaceae) from Indonesia.Crossref | GoogleScholarGoogle Scholar |
DuPuy DJ (1993) Aspleniaceae. In ‘Flora of Australia: Oceanic Islands 2. Vol. 50’. (Eds AS George, AE Orchard, HJ Hewson) pp. 554–558. (Australian Government Publishing Service: Canberra)
Dyer RJ, Savolainen V, Schneider H (2012) Apomixis and reticulate evolution in the Asplenium monanthes fern complex. Annals of Botany 110, 1515–1529.
| Apomixis and reticulate evolution in the Asplenium monanthes fern complex.Crossref | GoogleScholarGoogle Scholar | 22984165PubMed |
Gabancho LR, Prada C (2011) The genus Hymenasplenium (Aspleniaceae) in Cuba, including new combinations for the neotropical species. American Fern Journal 101, 265–281.
| The genus Hymenasplenium (Aspleniaceae) in Cuba, including new combinations for the neotropical species.Crossref | GoogleScholarGoogle Scholar |
Gastony GJ, Johnson WP (2001) Phylogenetic placements of Loxoscaphe thecifera (Aspleniaceae) and Actiniopteris radiata (Pteridaceae) based on analysis of rbcL sequence. American Fern Journal 91, 197–213.
| Phylogenetic placements of Loxoscaphe thecifera (Aspleniaceae) and Actiniopteris radiata (Pteridaceae) based on analysis of rbcL sequence.Crossref | GoogleScholarGoogle Scholar |
Gehrke B, Linder HP (2011) Time, space and ecology: why some clades have more species than others. Journal of Biogeography 38, 1948–1962.
| Time, space and ecology: why some clades have more species than others.Crossref | GoogleScholarGoogle Scholar |
Geiger JMO, Ranker TA, Ramp Neale JM, Klimas ST (2007) Molecular biogeography and origins of the Hawaiian fern flora. Brittonia 59, 142–158.
| Molecular biogeography and origins of the Hawaiian fern flora.Crossref | GoogleScholarGoogle Scholar |
Green PS (1994) Aspleniaceae. In ‘Flora of Australia: Oceanic Islands 2. Vol. 50’. (Ed. AJG Wilson) pp. 591–597. (Australian Government Publishing Service: Canberra)
Guindon S, Dufayard JF, Lefort V, Anisimova M, Hordijk W, Gascuel O (2010) New algorithms and methods to estimate maximum-likelihood phylogenies: assessing the performance of PhyML 3.0. Systematic Biology 59, 307–321.
| New algorithms and methods to estimate maximum-likelihood phylogenies: assessing the performance of PhyML 3.0.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXks1Kms7s%3D&md5=2e3e202e8279618465b24e4d9cc74b41CAS | 20525638PubMed |
Hasebe M, Omori T, Nakazawa M, Sano T, Kato M, Iwatsuki K (1994) rbcL gene sequences provide evidence for the evolutionary lineages of leptosporangiate ferns. Proceedings of the National Academy of Sciences of the United States of America 91, 5730–5734.
| rbcL gene sequences provide evidence for the evolutionary lineages of leptosporangiate ferns.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK2cXksFCju7Y%3D&md5=585d59f9a1be845db1d38a93bd18b1c6CAS | 8202555PubMed |
Holttum RE (1974) Asplenium Linn., sect. Thamnopteris Presl. The Gardens’ Bulletin Singapore 27, 143–154.
Huelsenbeck JP, Ronquist FR (2001) MrBayes: Bayesian inference of phylogeny. Biometrics 17, 754–755.
Jones DL (1996) A new species of Asplenium L. section Thamnopteris C.Presl (Aspleniaceae) from Lord Howe Island. Muelleria 9, 37–40.
Kawakami S (1970) Karyological studies on Aspleniaceae. II. Chromosomes of seven species in Aspleniaceae. Botanical Magazine, Tokyo 83, 74–81.
| Karyological studies on Aspleniaceae. II. Chromosomes of seven species in Aspleniaceae.Crossref | GoogleScholarGoogle Scholar |
Kramer KU, Viane R (1990) Aspleniaceae. In ‘The Families and Genera of Vascular Plants: Pteridophytes and Gymnosperms 1’. (Eds. KU Kramer, PS Green) pp 52–56. (Springer-Verlag: Berlin)
Li C-X, Lu S-G (2006) Relationship of Asplenium yunnanense and A. lushanense inferred from the sequence analysis of chloroplast rbcL, trnL–F and rps4–trnS. Acta Phytotaxonomica Sinica 44, 296–303.
| Relationship of Asplenium yunnanense and A. lushanense inferred from the sequence analysis of chloroplast rbcL, trnL–F and rps4–trnS.Crossref | GoogleScholarGoogle Scholar |
Lovis JD (1973) A biosystematic approach to phylo-genetic problems and its application to the Aspleniaceae. In ‘The Phylogeny and Classification of the Ferns’. (Eds AC Jermy Crabbe JA, Thomas BA) pp. 211–228. (Academic Press: London)
Manton I (1954) Cytological notes on one hundred species of Malayan ferns. In ‘A Revised Flora of Malaya. Vol. II’. (Ed. RE Holttum) p. 643.(Government Printing Office: Singapore)
Manton I, Sledge WA (1954) Observations on the cytology and taxonomy of the pteridophyte flora of Ceylon. Philosophical Transactions of the Royal Society of London. B, Biological Sciences 238, 127–185.
| Observations on the cytology and taxonomy of the pteridophyte flora of Ceylon.Crossref | GoogleScholarGoogle Scholar |
Mehra PN (1961) Chromosome numbers in Himalayan ferns. Research Bulletin of the Panjab University, Science 12, 139–164.
Mickel JT (1976) Sinephropteris, a new genus of scolopendroid ferns. Brittonia 28, 326–328.
| Sinephropteris, a new genus of scolopendroid ferns.Crossref | GoogleScholarGoogle Scholar |
Moran RC, Smith AR (2001) Phytogeographic relationships between neotropical and African–Madagascan pteridophytes. Brittonia 53, 304–351.
| Phytogeographic relationships between neotropical and African–Madagascan pteridophytes.Crossref | GoogleScholarGoogle Scholar |
Murakami N (1995) Systematics and evolutionary biology of the fern genus Hymenasplenium (Aspleniaceae). Journal of Plant Research 108, 257–268.
| Systematics and evolutionary biology of the fern genus Hymenasplenium (Aspleniaceae).Crossref | GoogleScholarGoogle Scholar |
Murakami N, Yokoyama J, Cheng X, Iwasaki H, Imaichi R, Iwatsuki K (1998) Molecular alpha-taxonomy of Hymenasplenium obliquissimum complex (Aspleniaceae) based on rbcL sequence comparisons. Plant Species Biology 13, 51–56.
| Molecular alpha-taxonomy of Hymenasplenium obliquissimum complex (Aspleniaceae) based on rbcL sequence comparisons.Crossref | GoogleScholarGoogle Scholar |
Murakami N, Nogami S, Watanabe M, Iwatsuki K (1999a) Phylogeny of Aspleniaceae inferred from rbcL nucleotide sequences. American Fern Journal 89, 232–243.
| Phylogeny of Aspleniaceae inferred from rbcL nucleotide sequences.Crossref | GoogleScholarGoogle Scholar |
Murakami N, Watanabe M, Yokoyama J, Yatabe Y, Iwasaki H, Serizawa S (1999b) Molecular taxonomic study and revision of the three Japanese species of Asplenium sect. Thamnopteris. Journal of Plant Research 112, 15–25.
| Molecular taxonomic study and revision of the three Japanese species of Asplenium sect. Thamnopteris.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1MXjvFShsLg%3D&md5=1c950e110a7be8f6f15f3716a9572cfaCAS |
Nakato N (1987) Notes on chromosomes of Japanese pteridophytes. The Journal of Japanese Botany 62, 261–267.
Nylander JAA (2004) ‘MrModeltest v2.’ (Program distributed by the author. Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden)
Ogle CC (1987) Taxonomic changes in Asplenium (Aspleniaceae; Filicales) in New Zealand. New Zealand Journal of Botany 25, 591–593.
| Taxonomic changes in Asplenium (Aspleniaceae; Filicales) in New Zealand.Crossref | GoogleScholarGoogle Scholar |
Ohlsen DJ (2014) Molecular systematics, biogeography, and taxonomy of the fern family Aspleniaceae in Australasia and the south-west Pacific. PhD Thesis, The University of Melbourne.
Ohlsen DJ, Perrie LR, Shepherd LD, Brownsey PJ, Bayly MJ (2014a) [2015] Investigation of species boundaries and relationships in the Asplenium paleaceum complex (Aspleniaceae) using AFLP fingerprinting and chloroplast and nuclear DNA sequences. Australian Systematic Botany 27, 378–394.
| [2015] Investigation of species boundaries and relationships in the Asplenium paleaceum complex (Aspleniaceae) using AFLP fingerprinting and chloroplast and nuclear DNA sequences.Crossref | GoogleScholarGoogle Scholar |
Ohlsen DJ, Perrie LR, Shepherd LD, Bayly MJ (2014b) [2015] Taxonomic status and distribution of the critically endangered Christmas Island spleenwort (Asplenium listeri, Aspleniaceae): it is not as rare as we thought. Australian Systematic Botany 27, 372–377.
| [2015] Taxonomic status and distribution of the critically endangered Christmas Island spleenwort (Asplenium listeri, Aspleniaceae): it is not as rare as we thought.Crossref | GoogleScholarGoogle Scholar |
Palmer DD (2003) Aspleniaceae In ‘Hawaii’s ferns and fern allies’. pp. 52–76. (University of Hawaii Press: Honolulu, HI)
Papadopulos AST, Baker WJ, Crayn D, Butlin RK, Kynast RG, Hutton I, Savolainen V (2011) Speciation with gene flow on Lord Howe Island. Proceedings of the National Academy of Sciences of the United States of America 108, 13188–13193.
| Speciation with gene flow on Lord Howe Island.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXhtVKitL7M&md5=1bbd35929e987a0263d3983b63e6dc78CAS |
Paris CA (1989) Cryptic species, species delimitation, and taxonomic practice in the homosporous ferns. American Fern Journal 79, 46–54.
| Cryptic species, species delimitation, and taxonomic practice in the homosporous ferns.Crossref | GoogleScholarGoogle Scholar |
Parris BS (2001) Circum-Antarctic continental distribution patterns in pteridophyte species. Brittonia 53, 270–283.
| Circum-Antarctic continental distribution patterns in pteridophyte species.Crossref | GoogleScholarGoogle Scholar |
Patiño J, Carine M, Fernandez-Palacios JM, Otto R, Schaefer H, Vanderpoorten A (2014) The anagenetic world of spore-producing land plants. New Phytologist 201, 305–311.
| The anagenetic world of spore-producing land plants.Crossref | GoogleScholarGoogle Scholar | 24010958PubMed |
Perrie LR, Brownsey PJ (2005) Insights into the biogeography and polyploid evolution of New Zealand Asplenium from chloroplast DNA sequence data. American Fern Journal 95, 1–21.
| Insights into the biogeography and polyploid evolution of New Zealand Asplenium from chloroplast DNA sequence data.Crossref | GoogleScholarGoogle Scholar |
Perrie LR, Shepherd LD, de Lange PJ, Brownsey PJ (2010) Parallel polyploidy speciation: distinct sympatric gene-pools of recurrently derived allooctoploid Asplenium ferns. Molecular Ecology 19, 2916–2932.
| Parallel polyploidy speciation: distinct sympatric gene-pools of recurrently derived allooctoploid Asplenium ferns.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXhtVyktb3J&md5=51f8746765bc0eb70f72175d49c53a3dCAS | 20579287PubMed |
Perrie LR, Wilson RK, Shepherd LD, Ohlsen DJ, Batty EL, Brownsey PJ, Bayly MJ (2014) Molecular phylogenetics and generic taxonomy of Blechnaceae ferns. Taxon 63, 745–758.
| Molecular phylogenetics and generic taxonomy of Blechnaceae ferns.Crossref | GoogleScholarGoogle Scholar |
Pichi Sermolli REG (1977) Tentamen pteridophytorum taxonomic ordinem redigeni. Webbia 31, 313–512.
| Tentamen pteridophytorum taxonomic ordinem redigeni.Crossref | GoogleScholarGoogle Scholar |
Pinter I, Bakker F, Barrett J, Cox C, Gibby M, Henderson S, Morgan-Richards M, Rumsey F, Russell S, Trewick S, Schneider H, Vogel J (2002) Phylogenetic and biosystematic relationships in four highly disjunct polyploid complexes in the subgenera Ceterach and Phyllitis in Asplenium (Aspleniaceae). Organisms, Diversity & Evolution 2, 299–311.
| Phylogenetic and biosystematic relationships in four highly disjunct polyploid complexes in the subgenera Ceterach and Phyllitis in Asplenium (Aspleniaceae).Crossref | GoogleScholarGoogle Scholar |
Rambaut A (2002) ‘Se-Al: sequence alignment editor.’ Available at http://tree.bio.ed.ac.uk/. [Verified 6 May 2015]
Rambaut A, Drummond AJ (2009) Tracer MCMC trace analysis tool, version v1.5.0. 2003–2009. Available at http://tree.bio.ed.ac.uk/software/tracer/ [Verified 2013]
Reichstein T (1981) Hybrids in European Aspleniceae (Pteridophyta). Botanica Helvetica 91, 89–139.
Rothfels CJ, Larsson A, Kuo L-Y, Korall P, Chiou W-L, Pryer KM (2012) Overcoming deep roots, fast rates, and short internodes to resolve the ancient rapid radiation of Eupolypod II ferns. Systematic Biology 61, 490–509.
| Overcoming deep roots, fast rates, and short internodes to resolve the ancient rapid radiation of Eupolypod II ferns.Crossref | GoogleScholarGoogle Scholar | 22223449PubMed |
Salvo AE, Prada C, Diaz T (1982) Revision del genero Asplenium L., subgenera Pleurosorus (Fee) Salvo, Prada & Diaz. Candollea (Genève) 37, 457–484.
Sasaki A (2008) Aspleniaceae. In ‘Illustrated flora of ferns and fern allies of South Pacific Islands’. (Ed. M Nakamura) pp. 133–146. (Tokai University Press: Tokyo, Japanm)
Schneider H, Russell SJ, Cox CJ, Bakker F, Henderson S, Rumsey F, Barrett J, Gibby M, Vogel JC (2004) Chloroplast phylogeny of asplenioid ferns based on rbcL and trnL–F spacer sequences (Polypodiidae, Aspleniaceae) and its implications for biogeography. Systematic Botany 29, 260–274.
| Chloroplast phylogeny of asplenioid ferns based on rbcL and trnL–F spacer sequences (Polypodiidae, Aspleniaceae) and its implications for biogeography.Crossref | GoogleScholarGoogle Scholar |
Schneider H, Ranker TA, Russell SJ, Cranfill R, Geiger JM, Aguraiuja R, Wood KR, Grundmann M, Kloberdanz K, Vogel JC (2005) Origin of the endemic fern genus Diella coincides with the renewal of Hawaiian terrestrial life in the Miocene. Proceedings. Biological Sciences 272, 455–460.
| Origin of the endemic fern genus Diella coincides with the renewal of Hawaiian terrestrial life in the Miocene.Crossref | GoogleScholarGoogle Scholar |
Schuettpelz E, Pryer KM (2007) Fern phylogeny inferred from 400 leptosporangiate species and three plastid genes. Taxon 56, 1037–1050.
| Fern phylogeny inferred from 400 leptosporangiate species and three plastid genes.Crossref | GoogleScholarGoogle Scholar |
Shepherd LD, Perrie LR, Brownsey PJ (2007) Fire and ice: volcanic and glacial impacts on the phylogeography of the New Zealand forest fern Asplenium hookerianum. Molecular Ecology 16, 4536–4549.
| Fire and ice: volcanic and glacial impacts on the phylogeography of the New Zealand forest fern Asplenium hookerianum.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXhsVSitrrJ&md5=05520b4b8b72e3a5933711770b8642e6CAS | 17877716PubMed |
Shepherd LD, Holland BR, Perrie LR (2008a) Conflict amongst chloroplast DNA sequences obscures the phylogeny of a group of Asplenium ferns. Molecular Phylogenetics and Evolution 48, 176–187.
| Conflict amongst chloroplast DNA sequences obscures the phylogeny of a group of Asplenium ferns.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXntlCktb8%3D&md5=8c90351dfc6cf6f523b3934556635086CAS | 18462954PubMed |
Shepherd LD, Perrie LR, Brownsey PJ (2008b) Low-copy nuclear DNA sequences reveal a predominance of allopolyploids in a New Zealand Asplenium fern complex. Molecular Phylogenetics and Evolution 49, 240–248.
| Low-copy nuclear DNA sequences reveal a predominance of allopolyploids in a New Zealand Asplenium fern complex.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXhtFOltr3M&md5=2d3225a4ddb5d0f215fd3087340ccf4dCAS | 18640280PubMed |
Short P, Dixon D, Osterkamp MM (2003) A review of ferns and fern allies of the Northern Territory. The Beagle 19, 7–80.
Smith AR, Pryer KM, Schuettpelz E, Korall P, Schneider H, Wolf PG (2006) A classification for extant ferns. Taxon 55, 705–731.
| A classification for extant ferns.Crossref | GoogleScholarGoogle Scholar |
Stuessy TF, Jakubowsky G, Gomez RS, Pfosser M, Schluter PM, Fer T, Sun BY, Kato H (2006) Anagenetic evolution in island plants. Journal of Biogeography 33, 1259–1265.
| Anagenetic evolution in island plants.Crossref | GoogleScholarGoogle Scholar |
Swofford DL (2002) ‘PAUP* 4.0: phylogenetic analysis using parsimony (*and other methods).’ (Sinauer Associates: Sunderland, MA)
Taberlet P, Gielly L, Pautou G, Bouvet J (1991) Universal primers for amplification of three non-coding regions of chloroplast DNA. Plant Molecular Biology 17, 1105–1109.
| Universal primers for amplification of three non-coding regions of chloroplast DNA.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK38Xhslel&md5=7da114d7dcd800cbbe3423fa69803643CAS | 1932684PubMed |
Tindale MD, Roy SK (2002) A cytotaxonomic survey of the pteridophyta of Australia. Australian Systematic Botany 15, 839–937.
| A cytotaxonomic survey of the pteridophyta of Australia.Crossref | GoogleScholarGoogle Scholar |
Trewick SA, Morgan-Richards M, Russell SJ, Henderson S, Rumsey FJ, Pinter I, Barrett JA, Gibby M, Vogel JC (2002) Polyploidy, phylogeography and Pleistocene refugia of the rockfern Asplenium ceterach: evidence from chloroplast DNA. Molecular Ecology 11, 2003–2012.
| Polyploidy, phylogeography and Pleistocene refugia of the rockfern Asplenium ceterach: evidence from chloroplast DNA.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38XosVWrsLk%3D&md5=e63990b6b846ddbe3c2b5f4cac27dd91CAS | 12296944PubMed |
Tryon RM, Tryon AF (1982) ‘Ferns and allied plants with special reference to tropical America.’ (Springer-Verlag: New York)
Van den heede CJ, Viane RLL, Chase MW (2003) Phylogenetic analysis of Asplenium subgenus Ceterach (Pteridophyta: Aspleniaceae) based on plastid and nuclear ribosomal ITS DNA. American Journal of Botany 90, 481–495.
| Phylogenetic analysis of Asplenium subgenus Ceterach (Pteridophyta: Aspleniaceae) based on plastid and nuclear ribosomal ITS DNA.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3sXivFegs78%3D&md5=dc8b55286004d4e16da0526790c63f97CAS | 21659141PubMed |
Viane RLL, Reichstein T (2003) Notes on new or interesting Asplenium species from Western Asia, including comments on Ching and Wu (1985), and Fraser-Jenkins (1992) Reliquiae Reichsteinianae 1. In ‘Pteridology in the New Millennium’. (Eds S Chandra, M Srivastava) pp. 73–105. (The University of Chicago Press: Chicago, IL)
Vogel JC, Russell SJ, Rumsey FJ, Barrett JA, Gibby M (1998) Evidence for maternal transmission of chloroplast DNA in the genus Asplenium (Aspleniaceae, Pteridophyta). Botanica Acta 111, 247–249.
| Evidence for maternal transmission of chloroplast DNA in the genus Asplenium (Aspleniaceae, Pteridophyta).Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1cXktVaquro%3D&md5=c01d14168f4ebd3261344454478ee236CAS |
Wagner WH, Moran RC, Werth CR (1993) Aspleniaceae. In ‘Flora of North America. Pteridophytes and gymnosperms, vol. 2’. pp. 228–245. (Oxford University Press: New York)
Yatabe Y, Murakami N (2003) Recognition of cryptic species in the Asplenium nidus complex using molecular data – a progress report. Telopea 10, 487–496.
Yatabe Y, Masuyama S, Darnaedi D, Murakami N (2001) Molecular systematics of the Asplenium nidus complexes from Mt Halimum National Park, Indonesia: evidence for reproductive isolation among three sympatric rbcL sequence types. American Journal of Botany 88, 1517–1522.
| Molecular systematics of the Asplenium nidus complexes from Mt Halimum National Park, Indonesia: evidence for reproductive isolation among three sympatric rbcL sequence types.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3MXmsVyhtbk%3D&md5=ed4c554b7ab1dff0ef111aca8f68035fCAS | 21669684PubMed |
Yatabe Y, Darnaedi D, Murakami N (2002) Allozyme analysis of cryptic species in the Asplenium nidus complex from West Java, Indonesia. Journal of Plant Research 115, 483–490.
| Allozyme analysis of cryptic species in the Asplenium nidus complex from West Java, Indonesia.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3sXitFehtL0%3D&md5=40816dd70b1a73887a80a977152170f7CAS | 12579452PubMed |
Yatabe Y, Shinohara W, Matsumoto S, Murakami N (2009) Patterns of hybrid formation among cryptic species of bird-nest fern, Asplenium nidus complex (Aspleniaceae), in West Malesia. Botanical Journal of the Linnean Society 160, 42–63.
| Patterns of hybrid formation among cryptic species of bird-nest fern, Asplenium nidus complex (Aspleniaceae), in West Malesia.Crossref | GoogleScholarGoogle Scholar |
Youxing L, Viane R (2013) Aspleniaceae. In ‘Flora of China’. (Eds ZY Wu, PH Raven, DY Hong) pp. 267–316. (Science Press: Beijing; and Missouri Botanical Garden Press: St Louis, MO)