

Provenance modulates sensitivity of stored seeds of the Australian native grass Neurachne alopecuroidea to temperature and moisture availability
Friday Gray A , Anne Cochrane B D and Pieter Poot CA 22 Lakeview Boulevard, Vasse, WA 6280, Australia.
B Biodiversity and Conservation Science, Department of Biodiversity, Conservation and Attractions, Locked Bag 104, Bentley Delivery Centre, Perth, WA 6983, Australia.
C School of Biological Sciences, Faculty of Science, University of Western Australia, 35 Stirling Highway, Crawley, WA 6009, Australia.
D Corresponding author. Email: anne.cochrane@dbca.wa.gov.au
Australian Journal of Botany 67(2) 106-115 https://doi.org/10.1071/BT18240
Submitted: 11 December 2018 Accepted: 24 February 2019 Published: 4 April 2019
Abstract
In the present study we assessed the sensitivity of stored seeds of the common grass Neurachne alopecuroidea R.Br. from south-western Australian sources to varying temperature and moisture conditions as a tool to anticipate potential adaptability of seeds to climate change. We examined among-population germination responses, focusing on germination of excised seeds to overcome possible dormancy imparted by the lemma and palea. We hypothesised that temperature above and below the optimum and low moisture potentials would adversely affect germination, and that conditions for successful germination would be associated with the local climate at each seed source site. Experiment 1 used a bi-directional temperature gradient plate to measure responses to constant and alternating temperatures (5–40°C). Experiment 2 examined responses to moisture availability using polyethylene glycol (PEG 8000) solutions at different temperatures. Temperature optima varied among populations with significant reductions in germination occurring only below 15°C. Germination speed and success declined with decreasing moisture availability, with greater impact at higher temperatures. Significant population variation was observed. Tolerance to temperature and moisture availability was higher than expected and some of this variation suggests adaptation to local climates across the species Western Australian distribution. We discuss these results in the context of seed use in restoration and global warming.
Additional keywords: climate change, germination, moisture availability, Poaceae, seed bank, temperature, Western Australia.
Introduction
Seed germination is a vulnerable stage in the life cycle of a flowering plant (Silvertown 2009; Walck et al. 2011), with temperature and moisture availability key variables that drive germination, emergence and establishment success (Fenner 2000). Species tend to germinate over a range of temperatures, but also have an optimum temperature (or temperature range) that elicits the most complete and rapid germination (Probert 2000). Decreases in final germination percentage and increases in time to germination generally occur when temperatures are either above or below species optima (e.g. Cochrane 2016) and rising temperatures can lead to shifts in germination timing (Mondoni et al. 2012; Fernández-Pascual et al. 2015). Declining soil water potential affects a seed’s ability to take up water (Hegarty 1978), and like sub- and supra-optimal temperatures, can cause delays and declines in germination (Hagon and Chan 1977; Cochrane et al. 2014). The close relationship between germination and the prevailing environment thus indicates that some seed traits have the potential to be influenced by changing climatic conditions (Walck et al. 2011). Changes to seed traits and their functions can impact on species distributions and abundances, ecosystem functioning and community dynamics (Walck et al. 2011). For these reasons, it is critical we understand the effects of environmental factors on this early stage of the plant life cycle.
Grasses are vitally important as food sources for the world’s population and their abundance and role in water catchments, biodiversity reserves and as carbon sinks have been major areas of research (Tilman 1997; Boval and Dixon 2012). Studies have demonstrated that sub- and supra-optimal temperatures can be detrimental to grass seed germination (McGinnies 1960; Hagon and Chan 1977; van den Berg and Zeng 2006). Temperature tolerance can also vary significantly between species and populations (McGinnies 1960; Lodge and Whalley 1981; Yang et al. 2014). Additionally, interactions between moisture availability and temperature during germination occur and can influence germination success (Tadmor et al. 1969; Sharma 1976). Although heat stress can be alleviated by moisture, moisture stress is exacerbated by high temperatures through the process of evapotranspiration.
Although the germination ecology of several important grasses has been well investigated, few studies have examined species restricted to Mediterranean-climate zones. The genus Neurachne R.Br. or ‘mulga grass’ has an Australasian distribution and is represented by 14 perennial species (Australian Plant Name Index, https://biodiversity.org.au/nsl/services/APNI, accessed 20 February 2018), with five species found in Western Australia. The common and widespread species Neurachne alopecuroidea R.Br. (foxtail mulga grass) occurs over a large climatic gradient in the winter wet, summer dry regions of the South West Australian Floristic Region (SWAFR), and in south-eastern South Australia and western Victoria (Prendergast and Hattersley 1985). It is a tufted perennial grass growing to 40 cm tall and the only species of Neurachne to grow in this Mediterranean region of Western Australia (https://florabase.dpaw.wa.gov.au/, accessed 6 March 2019). Plants are assumed to have a C3 photosynthetic pathway (Hattersley and Roksandic 1983), and by definition, germination and growth should be tied to the autumn–winter period when temperatures are cool and water availability is high (Hattersley 1983). Plants also have a highly flexible regeneration strategy and can resprout after disturbance (Harris et al. 2010).
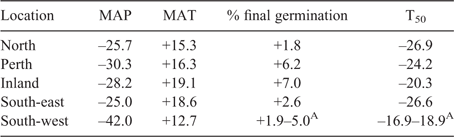
The SWAFR has a long history of a relatively stable and buffered Mediterranean climate that has resulted in the native flora being adapted for survival under local conditions (Hopper and Gioia 2004), including regeneration during the winter wet season when conditions are cool and moist (Bell et al. 1993; Bolger et al. 1999). If species are adapted to current local conditions then many species may now be at risk as the climate changes (Anderson 2016; CSIRO 2016). Temperatures in the region have already increased by ~1.1°C since 1910 (Hope et al. 2015) and rainfall has declined by around 20% since the mid-1970s (Bates et al. 2012). By 2070 mean temperatures are expected to further increase by 1–6°C with a 5–60% decrease in mean annual rainfall, relative to 1960–1990 averages (IOCI 2011). These warmer, drier conditions are likely to impact on the health and functioning of many species and ecosystems in the SWAFR (Anderson 2016; CSIRO 2016), and potentially on the grass N. alopecuroidea.
The objective of our study was to examine the sensitivity of seeds of N. alopecuroidea to temperature and moisture availability under laboratory conditions, and across multiple populations sourced from a latitudinal gradient in the SWAFR. We used seeds that had been stored cool and dry, and examined germination rather than dormancy. We hypothesised that (i) seeds have low temperature and high moisture requirements to coincide germination with the cool wet season; and (ii) that germination responses vary among seed populations and reflect the local climate conditions experienced at the different seed source locations. We expected that seed responses would follow a geographic cline with seeds from the warmer northern population being more successful than seeds from cooler climates when exposed to high temperatures. Similarly, seeds from the inland population, where average annual rainfall is low, were expected to perform better than seeds from wetter climates when exposed to low moisture potentials. Performance was measured in terms of speed and success of germination.
Methodology
Seed sources and preparation
Mature seeds of Neurachne alopecuroidea R.Br. were sourced from a latitudinal gradient in the SWAFR spanning ~600 km (Fig. 1; Table 1). Collections were made over several years by the Western Australian Department of Biodiversity, Conservation and Attractions’ Threatened Flora Seed Centre (TFSC) and stored dry at 15°C and 15% RH post-harvest, with the exception of the Perth and Inland collections, which had been dried and frozen at –20°C until a few weeks before the first experiment commenced. Seeds were removed from the fruiting spikelets before use as many grasses as considered to be dormant at seed shed as a result of the constrictions of the palea and lemma (Simpson 1990; Baskin and Baskin 2014; Bellairs et al. 2015). In this way we tried to separate out the processes of dormancy and germination, focusing solely on the latter. Whole seeds that were plump with no discolouration were assumed to be viable and were used in the germination experiments.

|

|
Experiment 1: effect of temperature on seed germination
We examined germination responses simultaneously over a gradient of constant and alternating temperatures between 5 and 40°C using a bi-directional temperature gradient plate (Model GRD1, Grant Instruments, Cambridge, UK). Seeds from four of the five populations (North, Perth, Inland and South-east) were plated onto 5 mL of 7.5 gm L–1 water agar in 35 mm diameter Petri dishes, with each dish containing 10 seeds. Each population was represented by 49 dishes and each cell of the gradient plate experienced a unique diurnal temperature combination. All cells were exposed to light for 12 h each day.
Germination, which was defined as radicle emergence to at least 3 mm, was scored daily for three weeks. All germinants and soft/mouldy seeds were removed each day. After 3 weeks, the remaining ungerminated seeds were transferred from the gradient plate to a constant 25°C incubator with a 12 h day/12 h night photoperiod and germination was scored for a further 3 weeks. This temperature was chosen as seeds demonstrated most rapid and complete germination under these conditions and we wished to determine whether seeds could ‘recover’ and continue to germinate after three weeks incubation under sub- and supra-optimal temperatures.
Experiment 2: effect of water availability on seed germination
Using all five populations (North, Perth, Inland, South-east and South-west), we examined the response of seeds to a range of moisture potentials (–0.25, –0.5, –0.75, –1.0, –1.25 or –1.5 MPa plus a control of 0 MPa) at three different constant temperatures (20, 25 and 30°C). Seeds access to moisture was constrained using aqueous solutions of polyethylene glycol (PEG 8000). PEG is a non-ionic water polymer that is widely used to create solutions of specific water potential values and therefore induce water stress (Michel and Kaufmann 1973; Ourry et al. 1992). The water potential of aqueous solutions of PEG is curvilinearly related to concentration and, at a given concentration, increases linearly with temperature (Michel and Kaufmann 1973). Solutions were made up to the appropriate strength with distilled (DI) water, according to an empirical equation that accounted for temperature differences (Michel and Kaufmann 1973; Hardegree and William 1990). The higher the concentration of solutes, the more negative the osmotic potential and the greater the moisture stress. The water potential of each solution was verified using a DecagonWP4 Dewpoint Potential Meter (Decagon Devices, Pullman, WA, USA). Five replicates of 10 seed from each of the five populations were plated onto 60 mm Petri dishes on Whatman (Maidstone, UK) filter paper over sponge. Each dish was irrigated with 7 mL of the appropriate solution and sealed in plastic zip lock bags to prevent desiccation. Following three weeks of daily germination scoring, seeds were transferred to new plates and irrigated only with DI water for a further three weeks. The viability of any remaining ungerminated seed was determined by examining seeds for emerging radicles or healthy internal embryos under a microscope.
Data analyses
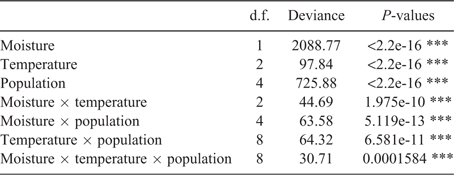
The data obtained from daily germination scoring of each cell on the temperature gradient plate and treatment in the incubators was used to calculate final percent germination and time to 20 or 50 percent germination (T20 and T50 respectively). A generalised linear model (GLM) with binomial error was used to determine the significance of the effects of mean temperature, magnitude of diurnal temperature fluctuations and population and their interaction on percentage germination on the temperature gradient plate. The most important attributes of fluctuating temperatures are the mean and magnitude (Roberts 1988). Over-dispersion was accounted for in this model with use of the quasibinomial function. A second model was used to determine the effects of mean temperatures, the magnitude of the diurnal temperature fluctuations and population on T50. The effects of diurnal temperature variability on germination rate are non-linear and best described by a polynomial curve. A quadratic model was therefore created to explain time to germination with T50 as the dependent variable and the temperature parameters and the square of the magnitude of the diurnal temperature fluctuations and population used as the explanatory variables.
A second GLM (binomial error) was used to relate percent germination (dependent variable) to moisture availability, temperature and population for Experiment 2. A linear model without quadratic functions was used to analyse the influence of moisture availability, temperature and population on T20 (dependent variable).
For both sets of data four GLMs, each with a different link (logit, cauchit, probit or cloglog), were originally created. The models with the lowest Akaike information criterions (cauchit model for the first dataset and probit for the second) were used for any further analysis. Heteroscedasticity was detected in both linear models and thus accounted for with the use of the vcovHC function. All four models included two-way interaction effects and were used to generate analysis of variance (ANOVA) tables. Pairwise t-tests with Holm adjustments were used to determine which populations had significantly different percent germination and T50 values (Experiment 1) or differences in percent germination and T20 between populations and moisture and temperature treatments (Experiment 2). Statistical analysis was conducted in R (ver. 3.3.2; R Core Team 2016).
Predictions for current and 2070 climate conditions
Temperature and precipitation values for current (1970–2000 averages) and future (2061–2080) conditions at the seed source sites were sourced from BIOCLIM (www.worldclim.com). Future values used the high greenhouse gas emission scenario for an increase in radiative forcing of 8.5 W m–2 by 2070, compared with pre-industrial values (RCP8.5). Current and future mean monthly minimum and maximum temperature values were used in the germination temperature response models to predict changes to germination percentage and speed. The differences between current and future temperature and rainfall values were determined to allow for calculation of percentage change. Predictions of percent germination and T50 under current and 2070 mean monthly temperatures were generated using the predict function in R. The predictions were made with temperature response GLM and Linear models similar to those described above but without temperature change magnitude as a variable. As there was no temperature response data for the South-west population predictions were generated from a simple GLM (binomial error, cauchit link) where average temperature was the only predictor variable and percent germination or T50 was the dependent variable. The range of predicted germination values generated by the models for the four populations tested on the temperature gradient plate was applied to the SW population as this population was not included in Experiment 1.
Results
Experiment 1: effect of temperature on seed germination
Germination ranged from 0 to 100% with only 8% of Petri dishes scoring less than 50% after 3 weeks. Time to initiate germination depended on the temperature regime in the cell and commenced at day 2 under optimal temperature conditions. It took 3–21 days for seeds to reach 50% germination (T50) and this was dependent on the temperature regime. The most rapid and complete germination (i.e. 100%) occurred between a mean temperature of 24 (Inland population) or 27°C (North population). The range of diurnal temperatures and times to 50% germination over which germination occurred are illustrated by contour plots (Fig. 2).

|
The mean of the diurnal temperature fluctuations, but not their magnitude, had a highly significant effect on germination levels (Table 2). Population alone was only marginally significant (P = 0.079), but the interaction between population and mean temperature was highly significant, with percentage germination of seeds of the South-east population significantly different to seeds from the Perth and Inland populations (P = 0.025). The germination proportion increased with rising mean temperatures and this response did not begin to decline until temperatures exceeded 30°C. For North and South-east populations, percent germination was 80% or greater across most temperature combinations, except where mean temperature was below 15°C (Fig. 2). Germination of seeds from the Perth and Inland populations declined at 25°C but was 80% or higher in most cells of the temperature gradient plate except where the mean temperature was below 15°C or above 30°C.

|
Both mean temperature and population had highly significant effects on T50 and there was a significant interaction between these variables (Table 2). Only the Inland and South-east populations differed from each other (P = 0.0042). Seeds of the North, South-east and Perth populations took 6 or fewer days to reach T50 when the mean temperature exceeded 20°C, except where the magnitude of diurnal fluctuations exceeded 15°C. The number of days to reach T50 was generally higher for the Inland population but this population was similarly affected by low temperatures and high magnitude temperature fluctuations. When temperatures exceeded 30°C, the speed of germination (as measured by T50) declined more rapidly for seeds of the Inland population than for the other populations (Fig. 2). At the conclusion of the experiment, 15% of seeds remained ungerminated. Less than 6% of these seeds did not germinate after an additional three weeks in a 25°C incubator, indicating that most of seeds used in the experiment were viable and capable of recovering from sub- and supra-optimal temperature conditions.
Experiment 2: effect of water availability on seed germination
Reductions in water potential caused a significant decline in percent germination and the effect of moisture availability was strongly dependent on population (Table 3): the North and South-east populations were least impacted by low moisture potentials, the Perth population was moderately affected and the Inland and South-west populations were strongly affected (Fig. 3). There was a significant interaction between temperature and moisture availability with declining moisture having a greater impact on germination at higher temperatures: germination was significantly lower at 25 and 30°C than at 20°C. For the North and South-east populations, the negative effect of decreasing water potential only became slightly more detrimental with increasing temperature, whereas this negative effect was strong for the Perth and South-west populations and very strong for the Inland population.
|

|
Time to 20% germination (T20) occurred between 3 and 21 days and was dependent on moisture potential, with the number of days to T20 increasing as water potential declined (Fig. 3). More than 30% of Petri dishes however, did not attain 20% germination. Population had a significant effect on T20: North and South-east seed populations generally had the lowest T20 values and the Inland and South-west seed had the highest. The effect of moisture availability also depended on population (interaction effect P < 0.001). The T20 of North, Perth and South-east populations tended to be less affected by low water potentials than the other populations (Fig. 3). Temperature was not a significant factor affecting T20 and the effect of moisture availability was not dependent on temperature. All seeds that remained ungerminated after the completion of Experiment 2 were confirmed to be viable and most had an emerging radicle, although not large enough to be considered ‘germinated’ (i.e. less than 3 mm).
Predictions for current and 2070 climate conditions
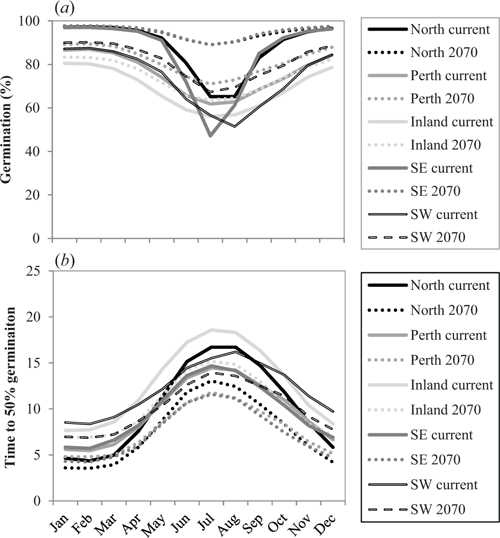
Mean monthly minimum and maximum temperatures are expected to increase significantly across the SWAFR in the next 50 years. The largest predicted changes are in mean annual temperature for the Inland population and in mean annual precipitation for the South-west population (Table 4). The models indicate that increases in temperature by 2070 may lead to marked changes in both percent germination and T50 (Fig. 4a, b), with the largest changes predicted for May–September. During this late autumn–winter-early spring period germination will increase and T50 will decline compared with current conditions.
|
|
Discussion
This study examined the germination of seeds of the common Australian grass, Neurachne alopecuroidea, under differing environmental conditions as a means to anticipate the potential adaptability of populations to changing temperature and moisture conditions. A major finding of this study was that N. alopecuroidea seeds have significant resilience to high temperatures and some resistance to reduced water availability, higher than previously reported for other Australian C3 grasses (Hagon 1976; Hagon and Chan 1977; Lodge and Whalley 1981). Our assumption that seeds of N. alopecuroidea would be negatively impacted by high temperatures based on this classification as a C3 plant, due to its distribution in a winter wet, summer dry Mediterranean-climate zone and the assumption of winter germination, was unfounded. Seeds showed an exceptional capacity to recover from sub- and supra-optimal temperature conditions, demonstrating that seeds are highly resilient to adverse conditions. Although seeds were negatively impacted by low water potential, we consider it likely that most seeds will germinate outside the currently assumed winter temperature conditions, provided sufficient moisture is available in the soil.
There is a clear indication that warmer temperatures are required for most rapid and complete germination of N. alopecuroidea with the optimal timing for germination occurring during the summer months (December–March). It could be assumed that this would only occur if sufficient moisture were available. Declining rainfall may therefore have a greater impact on N. alopecuroidea germination at locations where average temperatures are higher as final germination at warmer sites was more dependent on water availability. This finding corresponds well with results from previous studies that investigated other native and agricultural grass species however not our study species (Tadmor et al. 1969; Sharma 1976). North, Perth and South-east seed populations exhibited higher temperature and moisture tolerance that may be related to adaptation to the warmer drier local climates of these source populations. In contrast, the Inland population was not well adapted to dry conditions despite the lower rainfall experienced at its source site, and seeds originating from the highest rainfall zone (i.e. the South-west population) also exhibited limited adaptation to dry conditions possibly related to the high mean annual and mean monthly precipitation experienced at the seed source site.
Based on our modelling, the forecast changes to temperature and rainfall may have significant positive impacts on germination of some populations of N. alopecuroidea in the future. It is likely that germination overall will be more rapid and reach higher levels under rising temperatures. Cold temperatures during May–September (when moisture availability is greatest) and low precipitation during summer months will likely limit germination with germination more likely to occur during autumn and spring when water availability is adequate but temperatures are warmer, or indeed even occur following rains during summer. One explanation for the discrepancy between our results and those from other studies on Australian C3 grasses is the fact that we removed the surrounding structures from the seeds before incubation, thereby addressing some of the potential seed dormancy issues. Dormancy can prevent grass seeds from germinating under unsuitable conditions but can be alleviated artificially in several ways (for example, by removing the lemma and palea). In many studies seed dormancy was not addressed before experimentation and dormancy was overcome (or not) through the various temperature and moisture treatments provided to the seeds (e.g. Groves et al. 1982). Subsequent studies may need to conduct similar experiments on fresh seeds with their covering structures intact for comparative purposes and to isolate dormancy breaking treatments as compared with germination requirements for temperature and moisture.
We have demonstrated that populations of N. alopecuroidea varied in their responses to the applied treatments, but that the responses were not necessarily clinal. Species that occur across latitudinal, altitudinal or climatic gradients have a tendency to exhibit intra-specific variation in seed traits, though not always as a result of local adaptation to the prevailing conditions (Cochrane et al. 2015). There are some obvious differences between populations but most populations appear to have germination favoured by forecast conditions for 2070, given sufficient water availability. However, as mean annual rainfall is predicted to decline we may expect to see marked reductions in soil moisture availability which may well have negative impacts on germination. Our modelling suggests that the North, Perth and South-east populations will have greatest tolerance to climate conditions forecast for 2070 while the South-west population will be least resilient. This latter population has the greatest forecast rainfall decline and its seeds exhibit the least tolerance to reductions in water availability. However, we recognise the discrepancy between environmental data obtained from models such as BioClim and soil temperature and soil water potential sensed by the seed in the soil. We also acknowledge the inherent difficulties in inferring climate-germination relationships along an environmental gradient when only one population per point on the gradient is used. Nevertheless, it seems unlikely that seeds of N. alopecuroidea will be significantly affected by increasing temperatures during germination per se. In itself, ability to thrive under high temperatures is promising, but seeds were less resilient to declining water availability, and reductions in total rainfall are expected across the region. How our laboratory moisture stress treatments and the modelled changes in rainfall relate to field soil moisture availability is uncertain, and how likely seeds would be subjected to such conditions remains unknown. Changes to temperature and rainfall conditions may lead to a seasonal shift in germination timing. This can have far-reaching consequences for seedling establishment, and hence for population size and structure and community composition in species that rely on seed germination for population persistence and colonisation. However, when dormancy is removed (as in this study when we removed the lemma and palea), it is difficult to predict germination timing or the seasonal changes in dormancy and germination and their interaction with the environment that occurs in the field. This is particularly pertinent under climate change scenarios as dormancy is a main driver of germination timing in nature. We also did not focus on fresh seeds’ response to environmental variables and the subsequent ecological interpretations gained for natural populations.
Nevertheless, understanding how widely distributed plant species, including those that may be crucial to ecosystem functioning, will respond to changing environmental conditions across their populations can provide information to direct the focus of conservation and restoration objectives. Although we gained no insight into the germination or dormancy mechanisms at play in fresh seeds, we learnt that seeds are capable of germinating well after being stored for up to12 years under cool dry conditions. This confirms that ex-situ seed banking is an important and effective means to conserve genetic diversity for future use, and thus contributes to planning for restoration projects. However, our understanding of the impacts of higher temperatures on seedling growth and survival is still poorly understood and requires further investigation, including obtaining data on changes in surface soil moisture availability as rainfall declines.
We have demonstrated that seed sensitivity to temperature and moisture availability in N. alopecuroidea varies among populations, and that some of this variation appears linked to differences in environmental conditions experienced at the seed source locations, but not all. Adaptation to warm, dry conditions should increase the survival of those populations but where there is preference for cooler, moist conditions vulnerability may be increased. The considerable variation in germination response across the species Western Australian distribution suggests that some sites may be better seed sources for future restoration than others. Restoration initiatives should take into account the resilience of certain seed populations to altered climatic conditions. In the past, local seed has been widely advocated for such projects (O’Brien et al. 2007), but the use of ecotypes that are better adapted to predicted climates may increase the resilience of restoration plantings (Bischoff et al. 2006; Broadhurst et al. 2008). This climate-adjusted seed sourcing is likely to increase the chance of species survival (Prober et al. 2015). Mixing populations may increase the genetic diversity of plantings, which can improve adaptability and decrease the risk of inbreeding depression (Broadhurst et al. 2008). Such an approach could aid in conserving plant diversity across a range of other common species.
Conflicts of interest
The authors declare no conflict of interest.
Funding statement
This research did not receive any specific funding.
Acknowledgements
We thank the Threatened Flora Seed Centre (Department of Biodiversity, Conservation and Attractions) for providing seeds and assistance with seed processing. The temperature gradient plate was purchased by the South Coast Natural Resource Management Inc. (Project 04SC1-13 h) from a 2008 Australian Government Natural Heritage Trust grant. We would like to acknowledge David Merritt and Amber Bateman for providing instruction and access to the Decagon WP4 Dewpoint Potential Meter and other equipment and facilities at the Biodiversity Conservation Centre at Kings Park, Perth, Western Australia. This research was part of an Honours degree at the University of Western Australia, Perth. We thank two anonymous reviewers for their comments which have improved the manuscript. All experiments conducted as part of this research comply with current Australian laws. The datasets generated and/or analysed during this study are available from the primary author on reasonable request.
References
Anderson JT (2016) Plant fitness in a rapidly changing world. New Phytologist 210, 81–87.| Plant fitness in a rapidly changing world.Crossref | GoogleScholarGoogle Scholar | 26445400PubMed |
Barrow CJ (1992) ‘World atlas of desertification. United Nations Environment Programme.’ (Edward Arnold: London)
Baskin CC, Baskin JM (2014) ‘Seeds: ecology, biogeography, and evolution of dormancy and germination.’ 2nd edn. (Academic Press: San Diego, CA, USA)
Bates B, Frederiksen C, Wormworth J (2012) ‘Western Australia’s weather and climate: a synthesis of Indian Ocean Climate Initiative Stage 3 research’. (CSIRO: Canberra, ACT)
Bell DT, Plummer JA, Taylor SK (1993) Seed germination ecology in south-western Western Australia. Botanical Review 59, 24–73.
| Seed germination ecology in south-western Western Australia.Crossref | GoogleScholarGoogle Scholar |
Bellairs SM, Wurm PAS, Kernich B (2015) Temperature affects the dormancy and germination of sympatric annual (Oryza meridionalis) and perennial (O. rufipogon) native Australian rices (Poaceae) and influences their emergence in introduced para grass (Urochloa mutica) swards. Australian Journal of Botany 63, 687–695.
| Temperature affects the dormancy and germination of sympatric annual (Oryza meridionalis) and perennial (O. rufipogon) native Australian rices (Poaceae) and influences their emergence in introduced para grass (Urochloa mutica) swards.Crossref | GoogleScholarGoogle Scholar |
Bischoff A, Vonlanthen B, Steinger T, Muller-Scharer H (2006) Seed provenance matters – effects on germination of four plant species used for ecological restoration. Basic and Applied Ecology 7, 347–359.
| Seed provenance matters – effects on germination of four plant species used for ecological restoration.Crossref | GoogleScholarGoogle Scholar |
Bolger TP, Chapman R, Le Coultre IF (1999) Seed dormancy release in three common pasture grasses from a Mediterranean-type environment under contrasting conditions. Australian Journal of Experimental Agriculture 39, 143–147.
| Seed dormancy release in three common pasture grasses from a Mediterranean-type environment under contrasting conditions.Crossref | GoogleScholarGoogle Scholar |
Boval M, Dixon RM (2012) The importance of grasslands for animal production and other functions: a review on management and methodological progress in the tropics. Animal 6, 748–762.
| The importance of grasslands for animal production and other functions: a review on management and methodological progress in the tropics.Crossref | GoogleScholarGoogle Scholar | 22558923PubMed |
Broadhurst LM, Lowe A, Coates DJ, Cunningham SA, McDonald M, Vesk PA, Yates C (2008) Seed supply for broadscale restoration: maximizing evolutionary potential. Evolutionary Applications 1, 587–597.
Cochrane A (2016) Can sensitivity to temperature during germination help predict global warming vulnerability? Seed Science Research 26, 14–29.
| Can sensitivity to temperature during germination help predict global warming vulnerability?Crossref | GoogleScholarGoogle Scholar |
Cochrane JA, Hoyle GL, Yates CJ, Wood J, Nicotra AB (2014) Evidence of population variation in drought tolerance during seed germination in four Banksia (Proteaceae) species from Western Australia. Australian Journal of Botany 62, 481–489.
| Evidence of population variation in drought tolerance during seed germination in four Banksia (Proteaceae) species from Western Australia.Crossref | GoogleScholarGoogle Scholar |
Cochrane A, Yates CJ, Hoyle GL, Nicotra AB (2015) Will among-population variation in seed traits improve the chance of species persistence under climate change? Global Ecology and Biogeography 24, 12–24.
| Will among-population variation in seed traits improve the chance of species persistence under climate change?Crossref | GoogleScholarGoogle Scholar |
CSIRO (2016) State of the climate. Available at: https://www.csiro.au/en/Research/OandA/Areas/Assessing-our-climate/State-of-the-Climate-2018/Previous-State-of-the-Climate-reports/State-of-the-Climate-2016 (accessed 6 March 2019).
DPIRD (2017) Weather stations and radar. The Government of Western Australia, Department of Primary Industries and Regional Development. Available at: https://www.agric.wa.gov.au/weather-stations-and-radar (accessed 20 April 2018).
Fenner M (2000) ‘Seeds: the ecology of regeneration in plant communities.’ 2nd edn. (CABI International: Wallingford, UK)
Fernández-Pascual E, Seal CE, Pritchard HW (2015) Simulating the germination response to diurnally alternating temperatures under climate change scenarios: comparative studies on Carex diandra seeds. Annals of Botany 115, 201–209.
| Simulating the germination response to diurnally alternating temperatures under climate change scenarios: comparative studies on Carex diandra seeds.Crossref | GoogleScholarGoogle Scholar | 25564469PubMed |
Groves R, Hagon M, Ramakrishnan P (1982) Dormancy and germination of seed of eight populations of Themeda australis. Australian Journal of Botany 30, 373–386.
| Dormancy and germination of seed of eight populations of Themeda australis.Crossref | GoogleScholarGoogle Scholar |
Hagon M (1976) Germination and dormancy of Themeda australis, Danthonia spp., Stipa bigeniculata and Bothriochloa macra. Australian Journal of Botany 24, 319–327.
| Germination and dormancy of Themeda australis, Danthonia spp., Stipa bigeniculata and Bothriochloa macra.Crossref | GoogleScholarGoogle Scholar |
Hagon M, Chan C (1977) The effects of moisture stress on germination of some Australian native grass seeds. Australian Journal of Experimental Agriculture 17, 86–89.
| The effects of moisture stress on germination of some Australian native grass seeds.Crossref | GoogleScholarGoogle Scholar |
Hancock N, Hughes L (2014) Turning up the heat on the provenance debate: Testing the ‘local is best’ paradigm under heatwave conditions. Austral Ecology 39, 600–611.
| Turning up the heat on the provenance debate: Testing the ‘local is best’ paradigm under heatwave conditions.Crossref | GoogleScholarGoogle Scholar |
Hardegree SP, William EE (1990) Effect of polyethylene glycol exclusion on the water potential of solution-saturated filter paper. Plant Physiology 92, 462–466.
| Effect of polyethylene glycol exclusion on the water potential of solution-saturated filter paper.Crossref | GoogleScholarGoogle Scholar | 16667298PubMed |
Harris RJ, Mioduszewski P, Molony LN (2010) Vegetation responses to chaining in an isolated remnant in Western Australia’s wheatbelt. Journal of the Royal Society of Western Australia 93, 1–11.
Hattersley PW (1983) The distribution of C3 and C4 grasses in Australia in relation to climate. Oecologia 57, 113–128.
| The distribution of C3 and C4 grasses in Australia in relation to climate.Crossref | GoogleScholarGoogle Scholar | 28310164PubMed |
Hattersley P, Roksandic Z (1983) δ13C values of C3 and C4 species of Australian Neurachne and its allies (Poaceae). Australian Journal of Botany 31, 317–321.
| δ13C values of C3 and C4 species of Australian Neurachne and its allies (Poaceae).Crossref | GoogleScholarGoogle Scholar |
Hattersley PW, Watson L, Johnston CR (1982) Remarkable leaf anatomical variations in Neurachne and its allies (Poaceae) in relation to C3 and C4 photosynthesis. Botanical Journal of the Linnean Society 84, 265–272.
| Remarkable leaf anatomical variations in Neurachne and its allies (Poaceae) in relation to C3 and C4 photosynthesis.Crossref | GoogleScholarGoogle Scholar |
Hegarty TW (1978) The physiology of seed hydration and dehydration, and the relation between water stress and the control of germination: a review. Plant, Cell & Environment 1, 101–119.
| The physiology of seed hydration and dehydration, and the relation between water stress and the control of germination: a review.Crossref | GoogleScholarGoogle Scholar |
Hope P, Abbs D, Bhend J, Chiew F, Church J, Ekström M, Kirono D, Lenton A, Lucas C, McInnes K, Moise A, Monselesan D, Mpelasoka F, Timbal BLW, Whetton P (2015) ‘Southern and South-Western Flatlands cluster report, climate change in Australia: projections for Australia’s natural resource management regions.’ (CSIRO: Canberra, ACT)
Hopper SD, Gioia P (2004) The south-west Australian floristic region: evolution and conservation of a global hot spot of biodiversity. Annual Review of Ecology, Evolution, and Systematics 35, 623–650.
| The south-west Australian floristic region: evolution and conservation of a global hot spot of biodiversity.Crossref | GoogleScholarGoogle Scholar |
IOCI (2011) Projected changes in our climate. Indian Ocean Climate Initiative. Available at: http://www.ioci.org.au/our-climate.html?start=1 (accessed 12 July 2017).
Lodge G, Whalley R (1981) Establishment of warm- and cool-season native perennial grasses on the north-west slopes of New South Wales. Australian Journal of Botany 29, 111–119.
| Establishment of warm- and cool-season native perennial grasses on the north-west slopes of New South Wales.Crossref | GoogleScholarGoogle Scholar |
McGinnies WJ (1960) Effects of moisture stress and temperature on germination of six range grasses. Agronomy Journal 52, 159–162.
| Effects of moisture stress and temperature on germination of six range grasses.Crossref | GoogleScholarGoogle Scholar |
Michel BE, Kaufmann MR (1973) The osmotic potential of polyethylene glycol 6000. Plant Physiology 51, 914–916.
| The osmotic potential of polyethylene glycol 6000.Crossref | GoogleScholarGoogle Scholar | 16658439PubMed |
Mondoni A, Rossi G, Orsenigo S, Probert RJ (2012) Climate warming could shift the timing of seed germination in alpine plants. Annals of Botany 110, 155–164.
| Climate warming could shift the timing of seed germination in alpine plants.Crossref | GoogleScholarGoogle Scholar | 22596094PubMed |
O’Brien EK, Mazanec RA, Krauss SL (2007) Provenance variation of ecologically important traits of forest trees: implications for restoration. Journal of Applied Ecology 44, 583–593.
| Provenance variation of ecologically important traits of forest trees: implications for restoration.Crossref | GoogleScholarGoogle Scholar |
Ourry A, Mesle S, Boucaud J (1992) Effects of osmotic stress (NaCl and polyethylene glycol) on nitrate uptake, translocation, storage and reduction in ryegrass (Lolium perenne L.). New Phytologist 120, 275–280.
| Effects of osmotic stress (NaCl and polyethylene glycol) on nitrate uptake, translocation, storage and reduction in ryegrass (Lolium perenne L.).Crossref | GoogleScholarGoogle Scholar |
Prendergast H, Hattersley P (1985) Distribution and cytology of Australian Neurachne and its allies (Poaceae), a group containing C3, C4 and C3-C4 intermediate species. Australian Journal of Botany 33, 317–336.
| Distribution and cytology of Australian Neurachne and its allies (Poaceae), a group containing C3, C4 and C3-C4 intermediate species.Crossref | GoogleScholarGoogle Scholar |
Prober S, Byrne M, McLean E, Steane D, Potts B, Vaillancourt R, Stock W (2015) Climate-adjusted provenancing: a strategy for climate-resilient ecological restoration. Frontiers in Ecology and Evolution 3, 65
| Climate-adjusted provenancing: a strategy for climate-resilient ecological restoration.Crossref | GoogleScholarGoogle Scholar |
Probert RJ (2000) The role of temperature in the regulation of seed dormancy and germination. In ‘Seeds: the ecology of regeneration in plant communities’. 2nd edn. (Ed. M Fenner) pp. 261–292. (CABI International: Wallingford, UK)
R Core Team (2016) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Available at: https://www.R-project.org/ (accessed 10 August 2017).
Ravenscroft CH, Fridley JD, Grime JP (2014) Intraspecific functional differentiation suggests local adaptation to long-term climate change in a calcareous grassland. Journal of Ecology 102, 65–73.
| Intraspecific functional differentiation suggests local adaptation to long-term climate change in a calcareous grassland.Crossref | GoogleScholarGoogle Scholar |
Roberts EH (1988) Temperature and seed germination. Symposia of the Society for Experimental Biology 42, 109–132.
Sharma ML (1976) Interaction of water potential and temperature effects on germination of three semi-arid plant species. Agronomy Journal 68, 390–394.
| Interaction of water potential and temperature effects on germination of three semi-arid plant species.Crossref | GoogleScholarGoogle Scholar |
Silvertown J (2009) ‘Introduction to plant population biology.’ 4 edn. (John Wiley & Sons Inc.: Hoboken, NJ, USA)
Simpson GM (1990) ‘Seed dormancy in grasses.’ (Cambridge University Press: Cambridge, UK)
Tadmor NH, Cohen Y, Harpaz Y (1969) Interactive effects of temperature and osmotic potential on the germination of range plants. Crop Science 9, 771–774.
| Interactive effects of temperature and osmotic potential on the germination of range plants.Crossref | GoogleScholarGoogle Scholar |
Tilman D (1997) Community invasibility, recruitment limitation, and grassland biodiversity. Ecology 78, 81–92.
| Community invasibility, recruitment limitation, and grassland biodiversity.Crossref | GoogleScholarGoogle Scholar |
van den Berg L, Zeng YJ (2006) Response of South African indigenous grass species to drought stress induced by polyethylene glycol (PEG) 6000. South African Journal of Botany 72, 284–286.
| Response of South African indigenous grass species to drought stress induced by polyethylene glycol (PEG) 6000.Crossref | GoogleScholarGoogle Scholar |
Walck JL, Hidayati SN, Dixon KW, Thompson K, Poschlod P (2011) Climate change and plant regeneration from seed. Global Change Biology 17, 2145–2161.
| Climate change and plant regeneration from seed.Crossref | GoogleScholarGoogle Scholar |
WorldClim – Global Climate Data (2016) WorldClim version2. Available at: http://worldclim.org/version2 (accessed 22 May 2018).
Yang S, Li X, Yang Y, Yin X, Yang Y (2014) Comparing the relationship between seed germination and temperature for Stipa species on the Tibetan Plateau. Botany 92, 895–900.
| Comparing the relationship between seed germination and temperature for Stipa species on the Tibetan Plateau.Crossref | GoogleScholarGoogle Scholar |